
Effect of Short-Term Phosphorus Supply on Rhizosphere Microbial Community of Tea Plants

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Field Site and Experiment Design
2.2. Tea tree and Soil Sampling and Preparation
2.3. Soil Chemical Properties and Plant Elemental Analyses
2.4. Soil DNA Extraction, Amplification, High Throughput Sequencing and Bioinformatics
2.5. Bioinformatics and Statistical Analysis
3. Results
3.1. Soil Properties and ACP Activity Change under Different Rates of P Input
3.2. Rhizosphere Microbial Community Diversity and Composition under Different Rates of P Input
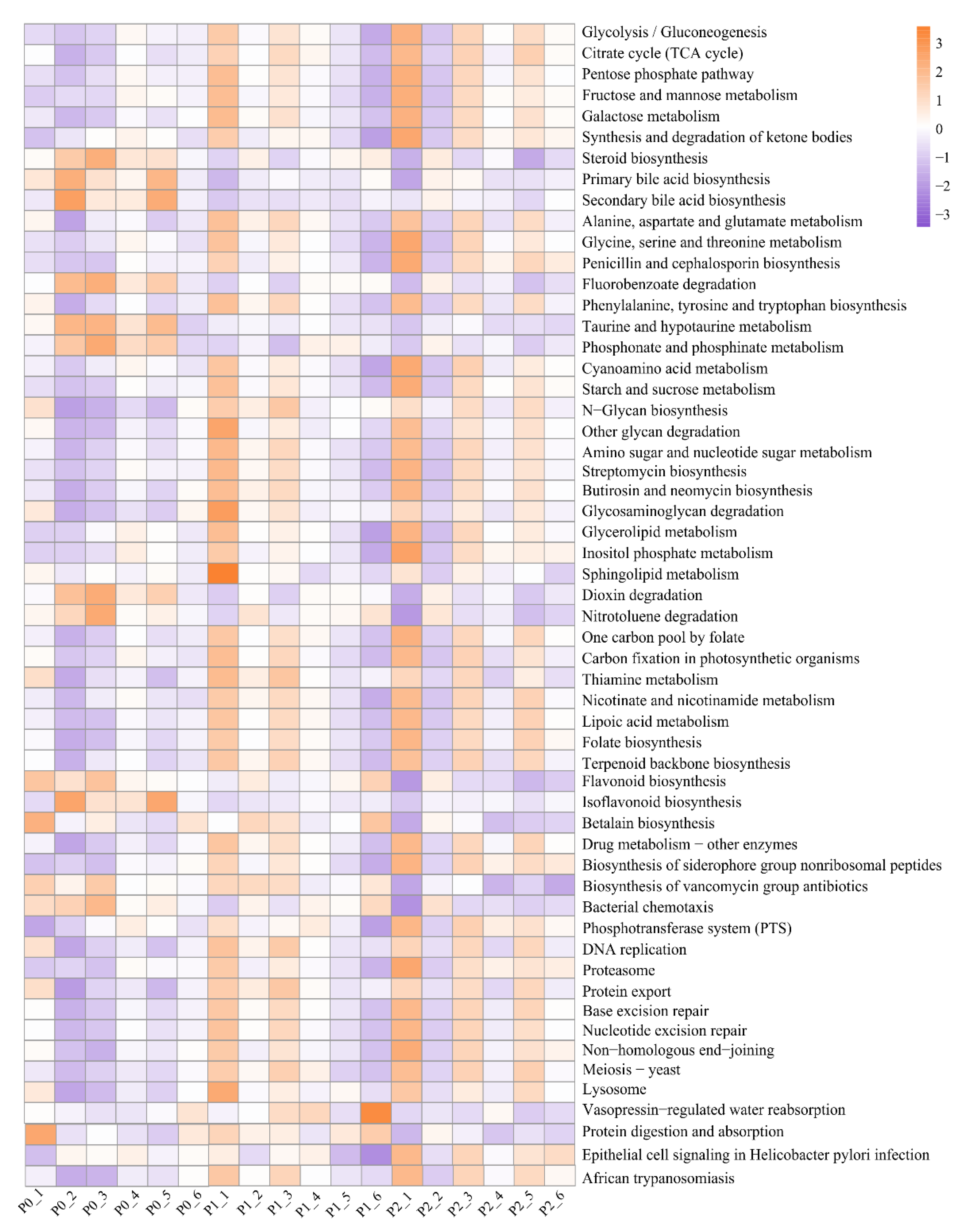
3.3. Rhizosphere Microbial Function Profiles under Different Rates of P Input
3.4. The Response of Tea Plant Biomass and P Content to Different Rates of P Input
4. Discussion
4.1. Effects of P Input Rate on Soil Physicochemical Properties, Tea Plant Biomass and P Content
4.2. Effects of P Input Rate on Rhizosphere Microbiome of Tea Plant
4.3. Effects of Rhizosphere Microbiome on Tea Biomass and Phosphorus Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bünemann, E.K.; Oberson, A.; Frossard, E. Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling; Springer: Berlin/Heidelberg, Germany, 2011; p. 483. [Google Scholar]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Sindhu, S.S.; Phour, M.; Choudhary, S.R.; Chaudhary, D. Phosphorus Cycling: Prospects of Using Rhizosphere Microorganisms for Improving Phosphorus Nutrition of Plants. Geomicrobiol. Biogeochem. 2014, 39, 199–237. [Google Scholar]
- Bergkemper, F.; Schöler, A.; Engel, M.; Lang, F.; Krüger, J.; Schloter, M.; Schulz, S. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 1988–2000. [Google Scholar] [CrossRef] [Green Version]
- Beauregard, M.S.; Hamel, C.; Atul, N.; St-Arnaud, M. Long-term phosphorus fertilization impacts soil fungal and bacterial diversity but not AM fungal community in alfalfa. Microb. Ecol. 2010, 59, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Gumiere, T.; Rousseau, A.N.; da Costa, D.P.; Cassetari, A.; Cotta, S.R.; Andreote, F.D.; Gumiere, S.J.; Pavinato, P.S. Phosphorus source driving the soil microbial interactions and improving sugarcane development. Sci. Rep. 2019, 9, 4400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, J.; Liu, X.; Hui, X.; Ma, Q.; Liu, J. Differentiating bacterial community responses to long-term phosphorus fertilization in wheat bulk and rhizosphere soils on the Loess Plateau. Appl. Soil Ecol. 2021, 166, 104090. [Google Scholar] [CrossRef]
- Ding, Z.; Jia, S.; Wang, Y.; Xiao, J.; Zhang, Y. Phosphate stresses affect ionome and metabolome in tea plants. Plant Physiol. Biochem. 2017, 120, 30–39. [Google Scholar] [CrossRef]
- Lin, Z.-H.; Qi, Y.-P.; Chen, R.-B.; Zhang, F.-Z.; Chen, L.-S. Effects of phosphorus supply on the quality of green tea. Food Chem. 2012, 130, 908–914. [Google Scholar] [CrossRef]
- Ni, K.; Liao, W.; Yi, X.; Niu, S.; Ma, L.; Shi, Y.; Zhang, Q.; Liu, M.; Ruan, J. Analysis on current situation and potential of fertilization reduction in Tea garden in China. J. Plant Nutr. Fertil. 2019, 25, 421–432. [Google Scholar]
- Ma, L.; Chen, H.; Shan, Y.; Jiang, M.; Zhang, G. Status and suggestion of tea garden fertilization on main green tea producing counties in Zhejiang Province. J. Tea Sci. 2013, 33, 74–84. [Google Scholar]
- Wen, Q.; Guo, Q.; Zhu, Y.; Dong, C. Phosphorus adsorption and desorption characteristics and pH value of agglomerates in acidic soils in southern China J. Nat. Sci. Heilongjiang Univ. 2014, 31, 800–805. [Google Scholar]
- Prescott, C.E.; Katzensteiner, K.; Weston, C. Soils and restoration of forested landscapes. In Soils and Landscape Restoration; Stanturf, J.A., Callaham, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 299–331. [Google Scholar]
- Liu, J.; Ma, Q.; Hui, X.; Ran, J.; Ma, Q.; Wang, X.; Wang, Z. Long-term high-P fertilizer input decreased the total bacterial diversity but not phoD-harboring bacteria in wheat rhizosphere soil with available-P deficiency. Soil Biol. Biochem. 2020, 149, 107918. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.O.; Pandey, R. Exploring phosphorus fertilizers and fertilization strategies for improved human and environmental health. Biol. Fertil. Soils 2020, 56, 299–317. [Google Scholar] [CrossRef] [Green Version]
- D’Haene, K.; Hofman, G. Phosphorus offtake and optimal phosphorus fertilisation rate of some fodder crops and potatoes in temperate regions. Agrokémia Talajt. 2015, 64, 403–420. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.S.; Kumar, G.; Bajpai, R.; Teli, B.; Rashid, M.; Sarma, B.K. PGPR formulations and application in the management of pulse crop health. In Biofertilizers; Rakshit, A., Meena, V.S., Parihar, M., Singh, H.B., Singh, A.K., Eds.; Woodhead Publishing: Cambridge, UK, 2021; pp. 239–251. [Google Scholar]
- Huang, D.; Ma, M.; Wang, Q.; Zhang, M.; Jing, G.; Li, C.; Ma, F. Arbuscular mycorrhizal fungi enhanced drought resistance in apple by regulating genes in the MAPK pathway. Plant Physiol. Biochem. 2020, 149, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Bucking, H.; Liepold, E.; Ambilwade, P. The Role of the Mycorrhizal Symbiosis in Nutrient Uptake of Plants and the Regulatory Mechanisms Underlying These Transport Processes. In Plant Science; Dhal, N.K., Sahu, S.C., Eds.; IntechOpen: London, UK, 2012; Chanpter 4. [Google Scholar]
- Ling, N.; Chen, D.; Guo, H.; Wei, J.; Bai, Y.; Shen, Q.; Hu, S. Differential responses of soil bacterial communities to long-term N and P inputs in a semi-arid steppe. Geoderma 2017, 292, 25–33. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.; Cao, F.; Shen, D.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Luo, G.; Xue, C.; Jiang, Q.; Xiao, Y.; Zhang, F.; Guo, S.; Shen, Q.; Ling, N. Soil Carbon, Nitrogen, and Phosphorus Cycling Microbial Populations and Their Resistance to Global Change Depend on Soil C:N:P Stoichiometry. mSystems 2020, 5, e00162-20. [Google Scholar] [CrossRef]
- Peng, Y.; Duan, Y.; Huo, W.; Zhang, Z.; Huang, D.; Xu, M.; Wang, X.; Yang, X.; Wang, B.; Kuzyakov, Y.; et al. C:P stoichiometric imbalance between soil and microorganisms drives microbial phosphorus turnover in the rhizosphere. Biol. Fertil. Soils 2022, 58, 421–433. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, G.; Chen, H.; Chen, C.; Wang, J.; Ai, S.; Wei, D.; Li, D.; Ma, B.; Tang, C.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, V.; Ray, P.; Craven, K.D. Rhizosphere Sampling Protocols for Microbiome (16S/18S/ITS rRNA) Library Preparation and Enrichment for the Isolation of Drought Tolerance-Promoting Microbes. In Plant Stress Tolerance: Methods and Protocols; Sunkar, R., Ed.; Springer: New York, NY, USA, 2017; pp. 349–362. [Google Scholar]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Geng, Y.; Zhou, H.; Yang, Y. Comparison of soil acid phosphatase activity determined by different methods. Chin. J. Eco-Agric. 2016, 24, 98–104. [Google Scholar]
- Hamalovâ, M.; Hodslavská, J.; Janos, P.; Kanický, V. Determination of Phosphorus, Potassium, and Magnesium in Fertilizers by Inductively Coupled Plasma–Atomic Emission Spectroscopy and Comparison with Other Techniques. J. AOAC Int. 2020, 80, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Lu, R. Methods for Soil Agrochemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Ji, L.; Wu, Z.; You, Z.; Yi, X.; Ni, K.; Guo, S.; Ruan, J. Effects of organic substitution for synthetic N fertilizer on soil bacterial diversity and community composition: A 10-year field trial in a tea plantation. Agric. Ecosyst. Environ. 2018, 268, 124–132. [Google Scholar] [CrossRef]
- Ji, L.; Ni, K.; Wu, Z.; Zhang, J.; Yi, X.; Yang, X.; Ling, N.; You, Z.; Guo, S.; Ruan, J. Effect of organic substitution rates on soil quality and fungal community composition in a tea plantation with long-term fertilization. Biol. Fertil. Soils 2020, 56, 633–646. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glockner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Caicedo, H.H.; Hashimoto, D.A.; Caicedo, J.C.; Pentland, A.; Pisano, G.P. Overcoming barriers to early disease intervention. Nat. Biotechnol. 2020, 38, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Huang, H. linkET: Everything is Linkable, R Package Version 0.0.3. Available online: http://github.com/Hy4m/linkET (accessed on 28 September 2022).
- Zhang, W.; Tang, X.; Feng, X.; Wang, E.; Li, H.; Shen, J.; Zhang, F. Management Strategies to Optimize Soil Phosphorus Utilization and Alleviate Environmental Risk in China. J. Environ. Qual. 2019, 48, 1167–1175. [Google Scholar] [CrossRef]
- Hui-min, G.; Bo-lang, C.; Qing-hui, W. Effects of phosphorus application on soil phosphorus availability and phosphorus fertilizer utilization rate in different cotton fields. Soil Fertil. China 2019, 3, 100–108. [Google Scholar]
- Ge, S.; Zhu, Z.; Jiang, Y. Long-term impact of fertilization on soil pH and fertility in an apple production system. J. Soil Sci. Plant Nutr. 2018, 18, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, X.; Guo, Z.; Jia, Z.; Wang, S.; Ding, K. Response of soil microbes to a reduction in phosphorus fertilizer in rice-wheat rotation paddy soils with varying soil P levels. Soil Tillage Res. 2018, 181, 127–135. [Google Scholar] [CrossRef]
- Zhao, M.; Zhao, J.; Yuan, J.; Hale, L.; Wen, T.; Huang, Q.; Vivanco, J.M.; Zhou, J.; Kowalchuk, G.A.; Shen, Q. Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ. 2021, 44, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-J.; Chen, Q.; Shi, W.-C.; Gao, Z.; Sun, X.; Dong, J.-J.; Li, J.; Wang, H.-T.; Gao, J.-G.; Liu, Z.-G.; et al. Interactions between phosphorus availability and microbes in a wheat–maize double cropping system: A reduced fertilization scheme. J. Integr. Agric. 2022, 21, 840–854. [Google Scholar] [CrossRef]
- Bui, E.N.; Henderson, B.L. C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 2013, 373, 553–568. [Google Scholar] [CrossRef]
- Song, Y.; Ai, Z.; Qiao, L.; Zhai, J.; Li, Y.; Li, Y. Effects of Fertilization on Ecological Stoichiometric Ratio Soil Carbon, Nitrogen and Nitrogen in Farmland of the Loess Plateau. Res. Soil Water Conserv. 2019, 26, 38–45, 52. [Google Scholar]
- Yang, F.; He, Y.; Li, C.; Wang, Y.; Lin, T. Effect fertilization on phosphorus fixation in upland red soil and its affecting factors. Acta Pedol. Sin. 2006, 43, 793–799. [Google Scholar]
- Tan, H.; Barret, M.; Mooij, M.J.; Rice, O.; Morrissey, J.P.; Dobson, A.; Griffiths, B.; O’Gara, F. Long-term phosphorus fertilisation increased the diversity of the total bacterial community and the phoD phosphorus mineraliser group in pasture soils. Biol. Fertil. Soils 2012, 49, 661–672. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Condron, L.M.; Gerard, E.; Dignam, B.E.A.; Black, A.; O’Callaghan, M. Long-term P fertilisation of pasture soil did not increase soil organic matter stocks but increased microbial biomass and activity. Biol. Fertil. Soils 2017, 53, 511–521. [Google Scholar] [CrossRef]
- Kaminsky, L.M.; Thompson, G.L.; Trexler, R.V.; Bell, T.H.; Kao-Kniffin, J. Medicago sativa has Reduced Biomass and Nodulation When Grown with Soil Microbiomes Conditioned to High Phosphorus Inputs. Phytobiomes J. 2018, 2, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Yuan, M.; Tang, L.; Shen, Y.; Yu, Q.; Li, S. Integrated microbiology and metabolomics analysis reveal responses of soil microorganisms and metabolic functions to phosphorus fertilizer on semiarid farm. Sci. Total Environ. 2022, 817, 152878. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, S.D.; Palmer, A.S.; Winsley, T.; Lamb, E.; Bissett, A.; Brown, M.V.; van Dorst, J.; Ji, M.; Ferrari, B.C.; Grogan, P.; et al. Soil fertility is associated with fungal and bacterial richness, whereas pH is associated with community composition in polar soil microbial communities. Soil Biol. Biochem. 2014, 78, 10–20. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, S.; Li, C.; Zhao, L.; Feng, H.; Yue, G.; Ren, Z.; Cheng, G. The soil carbon/nitrogen ratio and moisture affect microbial community structures in alkaline permafrost-affected soils with different vegetation types on the Tibetan plateau. Res. Microbiol. 2014, 165, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Samaddar, S.; Chatterjee, P.; Truu, J.; Anandham, R.; Kim, S.; Sa, T. Long-term phosphorus limitation changes the bacterial community structure and functioning in paddy soils. Appl. Soil Ecol. 2019, 134, 111–115. [Google Scholar] [CrossRef]
- Li, P.; Shen, C.; Jiang, L.; Feng, Z.; Fang, J. Difference in soil bacterial community composition depends on forest type rather than nitrogen and phosphorus additions in tropical montane rainforests. Biol. Fertil. Soils 2019, 55, 313–323. [Google Scholar] [CrossRef]
- Hanrahan-Tan, D.G.; Henderson, L.; Kertesz, M.A.; Lilje, O. The Effects of Nitrogen and Phosphorus on Colony Growth and Zoospore Characteristics of Soil Chytridiomycota. J. Fungi 2022, 8, 341. [Google Scholar] [CrossRef]
- Bonfante, P.; Venice, F. Mucoromycota: Going to the roots of plant-interacting fungi. Fungal Biol. Rev. 2020, 34, 100–113. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]
- Qin, H.; Lu, K.; Strong, P.J.; Xu, Q.; Wu, Q.; Xu, Z.; Xu, J.; Wang, H. Long-term fertilizer application effects on the soil, root arbuscular mycorrhizal fungi and community composition in rotation agriculture. Appl. Soil Ecol. 2015, 89, 35–43. [Google Scholar] [CrossRef]
- Urcoviche, R.C.; Gazim, Z.C.; Dragunski, D.C.; Barcellos, F.G.; Alberton, O. Plant growth and essential oil content of Mentha crispa inoculated with arbuscular mycorrhizal fungi under different levels of phosphorus. Ind. Crops Prod. 2015, 67, 103–107. [Google Scholar] [CrossRef]
- Wang, F.Y.; Hu, J.L.; Lin, X.G.; Qin, S.W.; Wang, J.H. Arbuscular mycorrhizal fungal community structure and diversity in response to long-term fertilization: A field case from China. World J. Microbiol. Biotechnol. 2011, 27, 67–74. [Google Scholar] [CrossRef]
- Sabater, S.; Artigas, J.; Gaudes, A.; MuÑOz, I.; Urrea, G.; RomanÍ, A.M. Long-term moderate nutrient inputs enhance autotrophy in a forested Mediterranean stream. Freshw. Biol. 2011, 56, 1266–1280. [Google Scholar] [CrossRef]
- Richardson, A.; Lynch, J.; Ryan, P.; Delhaize, E.; Smith, F.; Smith, S.; Harvey, P.; Ryan, M.; Veneklaas, E.; Lambers, H.; et al. Plant and microbial strategies to improve phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Jin, X.; Zeng, X.-Y.; Qi, C.-G.; Yin, L.-Y.; Deng, Y. Influences of phosphorus application level on maize arbuscular mycorrhizal colonization and hyphal acquisition to heterogeneous phosphorus supply. J. Plant Nutr. Fertil. 2018, 24, 163–169. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Zhang, H.; Wu, X.; Li, G.; Qin, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Ozimek, E.; Hanaka, A. Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture 2020, 11, 7. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Huang, X.-F.; Chaparro, J.M.; Badri, D.V.; Manter, D.K.; Vivanco, J.M.; Guo, J. Root and bacterial secretions regulate the interaction between plants and PGPR leading to distinct plant growth promotion effects. Plant Soil 2015, 401, 259–272. [Google Scholar] [CrossRef]
- Yi, X.; Ji, L.; Hu, Z.; Yang, X.; Li, H.; Jiang, Y.; He, T.; Yang, Y.; Ni, K.; Ruan, J. Organic amendments improved soil quality and reduced ecological risks of heavy metals in a long-term tea plantation field trial on an Alfisol. Sci Total Environ. 2022, 838, 156017. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | P0 | P1 | P2 |
|---|---|---|---|
| ACP (mg g−1 h−1 dry soil) | 0.94 ± 0.06 a | 0.76 ± 0.04 a | 0.84 ± 0.07 a |
| AP (mg kg−1) | 0.77 ± 0.08 b | 0.94 ± 0.1 ab | 1.57 ± 0.36 a |
| C:N Ratio | 7.20 ± 0.13 a | 7.21 ± 0.05 a | 6.90 ± 0.07 b |
| C:P Ratio | 4.68 ± 0.13 ab | 5.04 ± 0.14 a | 4.48 ± 0.25 b |
| N:P Ratio | 0.65 ± 0.01 a | 0.70 ± 0.02 a | 0.65 ± 0.03 a |
| pH | 3.31 ± 0.02 a | 3.32 ± 0.02 a | 3.23 ± 0.01 b |
| TC (g kg−1) | 6.82 ± 0.27 a | 6.92 ± 0.16 a | 6.79 ± 0.18 a |
| TN (g kg−1) | 0.95 ± 0.02 a | 0.96 ± 0.02 a | 0.98 ± 0.02 a |
| TP (g kg−1) | 1.46 ± 0.02 ab | 1.38 ± 0.01 b | 1.53 ± 0.06 a |
| Biomass (g pot−1) | 20.64 ± 3.54 b | 15.58 ± 0.66 b | 38.52 ± 2.26 a |
| P concentration (mg g−1 DW) | 1.26 ± 0.17 b | 1.87 ± 0.11 a | 1.91 ± 0.15 a |
| P content (mg pot−1) | 26.56 ± 6.84 b | 29.00 ± 2.02 b | 72.66 ± 5.35 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Ji, L.; Long, L.; Ni, K.; Yang, X.; Ma, L.; Guo, S.; Ruan, J. Effect of Short-Term Phosphorus Supply on Rhizosphere Microbial Community of Tea Plants. Agronomy 2022, 12, 2405. https://doi.org/10.3390/agronomy12102405
Yang H, Ji L, Long L, Ni K, Yang X, Ma L, Guo S, Ruan J. Effect of Short-Term Phosphorus Supply on Rhizosphere Microbial Community of Tea Plants. Agronomy. 2022; 12(10):2405. https://doi.org/10.3390/agronomy12102405
Chicago/Turabian StyleYang, Haoyu, Lingfei Ji, Lizhi Long, Kang Ni, Xiangde Yang, Lifeng Ma, Shiwei Guo, and Jianyun Ruan. 2022. "Effect of Short-Term Phosphorus Supply on Rhizosphere Microbial Community of Tea Plants" Agronomy 12, no. 10: 2405. https://doi.org/10.3390/agronomy12102405