
SNP and Haplotype Variability in the BnP5CR2 Gene and Association with Resistance and Susceptible Cultivars for Sclerotinia sclerotiorum in Brassica napus


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of the Pathogen of Sclerotinia sclerotiorum in Brassica napus
2.2. Field Experiments on Identification of Disease Resistance
2.3. Genomic DNA Extraction, Primer Synthesis, and PCR Amplification
2.4. Ligation and Sequencing
2.5. SNP Polymorphism of BnP5CR2
2.6. Evolutionary Relationships Analysis of BnP5CR2
2.7. BnP5CR2-CDS-Haplotype Diversity and Network Analysis
2.8. Expression Features Analysis of the BnP5CR2
3. Results
3.1. Identification of Sclerotinia sclerotiorum Resistance in Brassica napus
3.2. SNP Polymorphisms of BnP5CR2
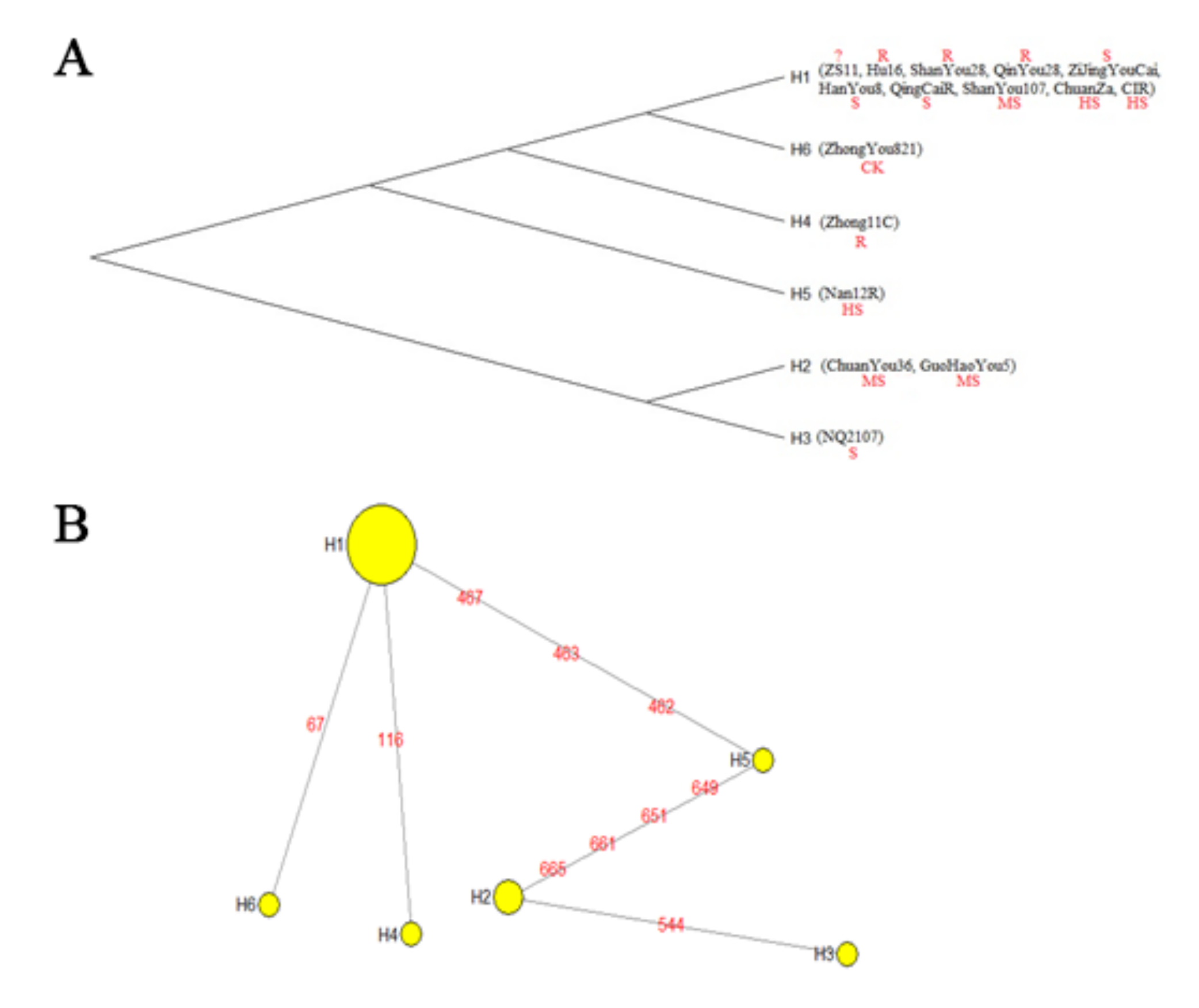
3.3. Evolutionary Tree of BnP5CR2
3.4. BnP5CR2-CDS-Haplotype Diversity and Network
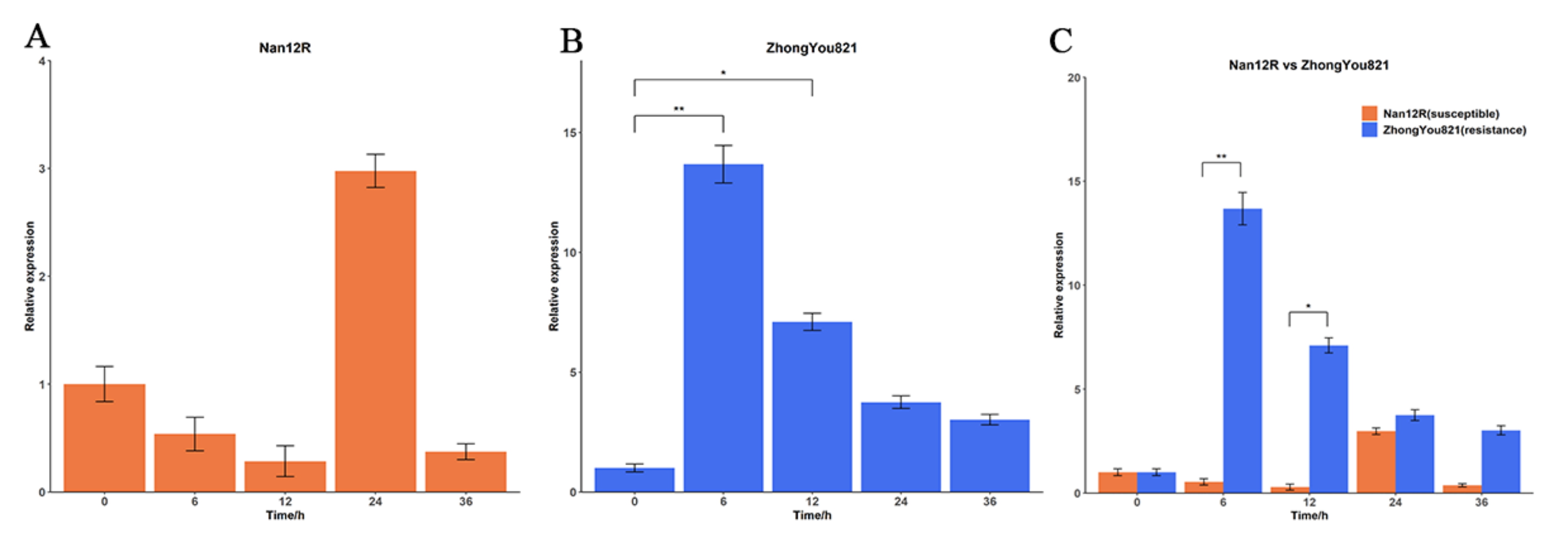
3.5. Expression Features of the BnP5CR2 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Wang, Y.X.; Zhou, W.Y.; Zheng, S.M.; Ye, R.Z. Detection of candidate gene networks involved in resistance to Sclerotinia sclerotiorum in soybean. J. Appl. Genet. 2021, 63, 1–14. [Google Scholar] [CrossRef]
- Mattioni, C.; Lacerenza, N.G.; Troccoli, A.; Deleonardis, M.A.; Difonzo, N. Water and salt stress-induced alterations in proline metabolism of Triticum durum seedlings. Physiol. Plant. 1997, 101, 787–792. [Google Scholar] [CrossRef]
- Roosens, N.H.; Thu, T.T.; Iskandar, H.M.; Jacobs, M. Isolation of the ornithine-delta-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis thaliana. Plant Physiol. 1998, 117, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.G.; Sun, C.; Liu, Y.L. Ornithine Pathway in Proline Biosynthesis Activated by Salt Stress in Barley Seedlings. Chin. Bull. Bot. 2001, 43, 36–40. Available online: https://www.jipb.net/EN/Y2001/V43/I1/36 (accessed on 3 August 2022).
- Ambikapathy, J.; Marshall, J.S.; Hocart, C.H.; Hardham, A.R. The role of proline in osmoregulation in Phytophthora nicotianae. Fungal Genet. Biol. 2001, 35, 287–299. [Google Scholar] [CrossRef]
- Quan, X.Q.; Zhang, Y.J.; Shan, L.; Bi, Y.P. Advances in Proline Metabolism Resear ches of Higher Plants. Biotechnol. Bull. 2007, 1, 14–18. [Google Scholar] [CrossRef]
- Funck, D.; Winter, G.; Baumgarten, L.; Forlani, G. Requirement of proline synthesis during Arabidopsis reproductive development. BMC Plant Biol. 2012, 12, 191. [Google Scholar] [CrossRef]
- Sun, C.C.; Zhao, H.Y.; Zheng, C.X. Effects of NaCl stress on osmolyte and proline metabolism in Ginkgo biloba seedling. Plant Physiol. J. 2017, 53, 186–192. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, X.Z.; Yang, J.H.; Zhang, S.; Zheng, C.S. Effect of grafting on osmolytes and expression of related metabolic enzyme genes in chrysanthemum under water stress. Plant Physiol. J. 2018, 363, 208–216. [Google Scholar] [CrossRef]
- Verbruggen, N.; Villarroel, R.; Van Montagu, M. Osmoregulation of a pyrroline-5-carboxylate reductase gene in Arabidopsis thaliana. Plant Physiol. 1993, 103, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Delauney, A.J.; Verma, D.P. A soybean gene encoding delta 1-pyrroline-5-carboxylate reductase was isolated by functional complementation in Escherichia coli and is found to be osmoregulated. Mol. Gen. Genet. 1990, 221, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.L.; Slocum, R.D. Molecular cloning and evidence for osmoregulation of the delta 1-pyrroline-5-carboxylate reductase (proC) gene in pea (Pisum sativum L.). Plant Physiol. 1992, 100, 1464–1470. [Google Scholar] [CrossRef]
- Walton, E.F.; Podivinsky, E.; Wu, R.M.; Rrynolds, P.H.S.; Young, L.W. Regulation of proline biosynthesis in kiwifruit buds with and without hydrogen cyanamide treatment. Physiol. Plant. 2002, 102, 171–178. [Google Scholar] [CrossRef]
- Sripinyowanich, S.; Klomsakul, P.; Boonburapong, B.; Bangyeekhun, B.; Asami, T.; Gu, H.Y. Exogenous ABA induces salt tolerance in indica rice (Oryza sativa L.): The role of OsP5CS1 and OsP5CR gene expression during salt stress. Environ. Exp. Bot. 2013, 86, 94–105. [Google Scholar] [CrossRef]
- Cao, L.; Wei, S.Q.; Han, L.; Qian, Y.Q.; Zhang, H.L.; Xin, H.B. Gene cloning and expression of the pyrroline-5-carboxylate reductase gene of perennial ryegrass (Lolium perenne). Hortic. Plant J. 2015, 1, 113–120. [Google Scholar] [CrossRef]
- Fu, L.L.; Han, B.Y.; Tan, D.G.; Sun, X.P.; Zhang, J.M. Gene Cloning of MeP5CS and MeP5CR in Cassava and Their Expression Analysis under Drought Stress. Hubei Agric. Sci. 2016, 55, 4024–4028. [Google Scholar] [CrossRef]
- Tang, F.F.; Zhao, Y.L.; Wang, P.L.; Feng, D.M.; Song, Y.; Gao, C.Q. Cloning and Stress Tolerance Analysis of ThP5CR from Tamarix hispida. Sci. Silvae Sin. 2017, 531–539. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Zhang, X.J.; Sun, X.M.; Ma, Z.; Zhang, Y. Polymorphic Study of P5CR in Rapeseed (B. napus). Genomics Appl. Biol. 2020, 39, 5678–5683. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, C.C.; Li, M.; Cui, Y.R.; Shi, Y.Y.; Wu, Z.C. The landscape of gene–CDS–haplotype diversity in rice: Properties, population organization, footprints of domestication and breeding, and implications for genetic improvement. Mol. Plant 2021, 14, 787–804. [Google Scholar] [CrossRef]
- Zhang, J.L.; Zhang, L.X.; Yang, K.M.; Yan, L.Z. Comparison Study on Extraction Methods for Genomic DNA from Tissues of Camellia Plants with Modified CTAB Method. Acta Agric. Univ. Jiangxiensis 2017, 39, 785–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Polzin, T.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2002; Volume 25, pp. 567–568. [Google Scholar] [CrossRef]
- Gao, N.X.; Mu, Y.C.; Jiang, B.X.; Li, M.M.; Gao, Z.M. Physiological and Biochemical Analyses of Resistance of Different Rape Varieties to Sclerotinia sclerotiorum; Aahui Agricultural University: Hefei, China, 2012. [Google Scholar]
- Chen, X.; Truksa, M.; Shah, S.; Weselake, R.J. A survey of quantitative real-time polymerase chain reaction internal reference genes for expression studies in Brassica napus. Anal. Biochem. 2010, 405, 138–140. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 27.0; IBM Corp: Armonk, NY, USA, 2020. [Google Scholar]
- Liu, L.Y. Method Evaluation of Sclerotinia Resistance Identification and Allelic Variation Analysis of Resistance-Related Genes and The Other Traits Genes in An Oilseed Rape Pedigree; Chinese Academy of Agricultural Sciences: Beijing, China, 2013. [Google Scholar]
- Li, H.; Wen, L.; Liu, K. Research progress on resistance mechanism of Brassica napus to Sclerotinia sclerotiorum. Crop Res. 2015, 29, 84–90. [Google Scholar] [CrossRef]
- Mao, W.; Ho, Y.M.; Liu, Z.W. Analysis of several enzymes activity induced by Sclerotinia sclerotiorum and oxalic acid in rapeseed. J. Dalian Polytech. Univ. 2011, 30, 39–42. [Google Scholar]
- Wan, H.F.; Liu, Y.; Mei, J.Q.; Ding, Y.J.; Liang, Y.; Qu, C.M. Relationship Between the Expression of Genes Encoding Resistance-related Enzymes and the Resistance to Sclerotinia sclerotiorum in Resynthesized Brassica napus with High Level of Resistance. Sci. Agric. Sin. 2012, 45, 4543–4551. [Google Scholar]
- Mwape, V.W.; Mobegi, F.M.; Regmi, R.; Newman, T.E.; Kamphuis, L.G.; Derbyshire, M.C. Analysis of differentially expressed Sclerotinia sclerotiorum genes during the interaction with moderately resistant and highly susceptible chickpea lines. BMC Genomics 2021, 22, 333. [Google Scholar] [CrossRef]
- Xu, B.; Gong, X.; Chen, S.; Hu, M.; Zhang, J.; Peng, Q. Transcriptome Analysis Reveals the Complex Molecular Mechanisms of Brassica napus–Sclerotinia sclerotiorum Interactions. Front. Plant Sci. 2021, 12, 716935. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.R.; Wan, L.L.; Zhang, X.H.; Xin, Q.; Song, Y.X.; Hong, D.F. Interaction between Brassica napus polygalacturonase inhibition proteins and Sclerotinia sclerotiorum polygalacturonase: Implications for rapeseed resistance to fungal infection. Planta 2021, 253, 34. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| BnP5CR2 | Forward primer 1: CAGTCAATAATTTATTTTGCGATGG; Reverse primer 1: AACTGCAGCTGGTGTATTAG. | 56 |
| Forward primer 2: TCAGTGTTTGCATCTAACATGCTCC; Reverse primer 2: TAACAAAGTAGCTGAGATCTGAACC. | 60 |
| Gene Name | NCBI Accession No. | Forward Primer Sequence (5′–3′) | Reverse Primers Sequence (5′–3′) |
|---|---|---|---|
| BnP5CR2 | Gene ID: 106416421 | GCTAATCGAAGCCGTGAACTC | AGTTTTCACATCTAC-CAAAGCAATA |
| UBC21 | Gene ID: 832645 | CCTCTGCAGCCTCCTCAAGT | CATATCTCCCCTGTCT-TGAAATGC |
| Sample | Disease Index | Relative Resistance Index | Resistance Rating | Sample | Disease Index | Relative Resistance Index | Resistance Rating |
|---|---|---|---|---|---|---|---|
| Hu16 | 17.14 | −0.432 | R | QingCaiR | 27.72 | 0.186 | S |
| ShanYou 28 | 22.02 | −0.121 | R | ShanYou 107 | 50.69 | 1.172 | MS |
| Zhong 11C | 21.66 | −0.142 | R | ChuanYou 36 | 55.48 | 1.364 | MS |
| QinYou 28 | 23.44 | −0.040 | R | GuoHaoYou 5 | 60.64 | 1.576 | MS |
| ZhongYou 821 | 24.16 | 0.000 | R | ChuanZa | 71.25 | 2.052 | HS |
| ZiJingYouCai | 24.87 | 0.038 | S | Nan12R | 85.16 | 2.891 | HS |
| HanYou 8 | 24.53 | 0.020 | S | CIR | 88.15 | 3.151 | HS |
| NQ2107 | 26.51 | 0.124 | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, Y.; Wu, D.; Qu, D.; Sun, X.; Zhang, X. SNP and Haplotype Variability in the BnP5CR2 Gene and Association with Resistance and Susceptible Cultivars for Sclerotinia sclerotiorum in Brassica napus. Agronomy 2022, 12, 2956. https://doi.org/10.3390/agronomy12122956
Zhang Y, Wang Y, Wu D, Qu D, Sun X, Zhang X. SNP and Haplotype Variability in the BnP5CR2 Gene and Association with Resistance and Susceptible Cultivars for Sclerotinia sclerotiorum in Brassica napus. Agronomy. 2022; 12(12):2956. https://doi.org/10.3390/agronomy12122956
Chicago/Turabian StyleZhang, Yu, Yu Wang, Dong Wu, Dong Qu, Xiaomin Sun, and Xiaojuan Zhang. 2022. "SNP and Haplotype Variability in the BnP5CR2 Gene and Association with Resistance and Susceptible Cultivars for Sclerotinia sclerotiorum in Brassica napus" Agronomy 12, no. 12: 2956. https://doi.org/10.3390/agronomy12122956
APA StyleZhang, Y., Wang, Y., Wu, D., Qu, D., Sun, X., & Zhang, X. (2022). SNP and Haplotype Variability in the BnP5CR2 Gene and Association with Resistance and Susceptible Cultivars for Sclerotinia sclerotiorum in Brassica napus. Agronomy, 12(12), 2956. https://doi.org/10.3390/agronomy12122956