
Growth and Nutritional Responses of Juvenile Wild and Domesticated Cacao Genotypes to Soil Acidity
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Localization and Cacao Clones
2.2. Cacao Seedling Propagation
2.3. Acidity-Stress Pot Experiment
2.4. Determination of Soil Physicochemical Properties
2.5. Determination of Biometric Parameters and Tolerance Index in Cacao Seedlings
- B0: Total dry biomass of each genotype without lime
- B1: Total dry biomass of each genotype with lime
- G0: Total dry biomass of each genotype without lime
- G0*: Mean total dry biomass of all assessed genotypes without lime
- G1: Total dry biomass of each genotype with lime
- G1*: Mean total dry biomass of all assessed genotypes with lime
- B0: Total dry biomass of each genotype without lime
- B1: Total dry biomass of each genotype with lime
- B0max: Maximum value of total dry biomass observed above all genotypes in conditions without lime.
2.6. Physiological Variables
2.7. Determination of Macro and Micronutrients Concentrations and Uptake in Cacao Shoots
- Element concentration = g kg−1 for macronutrients and mg kg−1 for micronutrients
- f = 1 for macronutrients and 1000 for micronutrients.
- Element concentration = g kg−1 for macronutrients and mg kg−1 for micronutrients
- NUE = g shoot g−1 for macronutrients and g shoot mg−1 for micronutrients.
2.8. Statistical Analysis
3. Results
3.1. Soil Physical and Chemical Attributes before and after Liming
3.2. Biometric and Physiological Characteristics of Cacao Plants under Acid Soils
3.3. Concentration and Uptake of Macro-Micro Nutrients in Juvenile Cacao Genotypes Grown on Limed and Unlimed Acid Soil
3.4. Comparison of Tolerance Indices to Acidic Soils in Juvenile Cacao
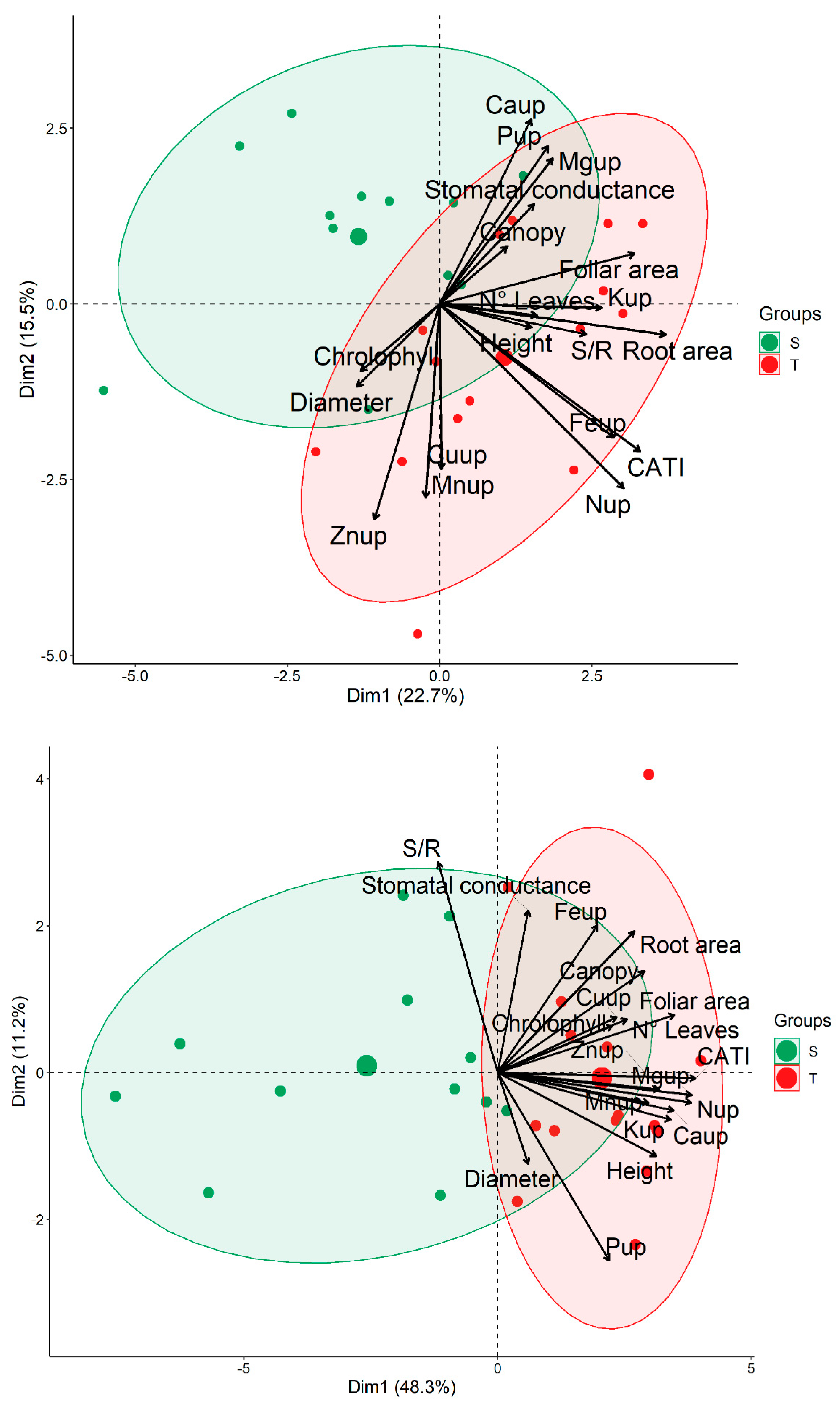
3.5. Relationship between Biometric Parameters and Nutrient Uptake in Different Clones
4. Discussion
4.1. Responses of Cacao Plants under Acid Soils
4.2. Nutrition and Tolerance to Soil Acidity of Cacao Plants
4.3. Relationship between Biometric, Physiology and Nutrition and Tolerance to Soil Acidity of Cacao Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Motamayor, J.C.; Lachenaud, P.; da Silva e Mota, J.W.; Loor, R.; Kuhn, D.N.; Brown, J.S.; Schnell, R.J. Geographic and genetic population differentiation of the Amazonian chocolate tree (Theobroma cacao L). PLoS ONE 2008, 3, 3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICCO Production of Cocoa Beans. Available online: https://www.icco.org/wp-content/uploads/Production_QBCS-XLVII-No.-2.pdf (accessed on 5 July 2021).
- Voora, V.; Bermudéz, S.; Larrea, C. Global Market Report: Cocoa. Sustainable Commodities Marketplace Series 2019; IISD-The International Institute for Sustainable Development: Winnipeg, MB, Canada, 2019; 12p, Available online: https://www.iisd.org/system/files/publications/ssi-global-market-report-cocoa.pdf (accessed on 6 September 2022).
- Instituto Nacional de Estadística e Informática-INEI. Resultados definitivos. IV Censo Nacional Agropecuario; INEI: Lima, Perú, 2013; 63p. [Google Scholar]
- Sombroek, W.G. Soils of the Amazon region. In Monographiae Biologicae; Springer: Dordrecht, The Netherlands, 1984; Volume 56, pp. 521–535. [Google Scholar]
- Rahman, M.A.; Lee, S.H.; Ji, H.C.; Kabir, A.H.; Jones, C.S.; Lee, K.W. Importance of mineral nutrition for mitigating aluminum toxicity in plants on acidic soils: Current status and opportunities. Int. J. Mol. Sci. 2018, 19, 3073. [Google Scholar] [CrossRef] [Green Version]
- Rengel, Z. Soil Acidification: The World Story. In Handbook of Soil Acidity; Rengel, Z., Ed.; CRC Press: Boca Raton, FL, USA, 2003; Volume 1. [Google Scholar]
- Miheretu, B. Soil acid management using biochar: Review. Int. J. Agric. Sci. Food Technol. 2020, 6, 211–217. [Google Scholar] [CrossRef]
- Takala, B. Soil acidity and its management options in Western Ethiopia: Review. JEES 2019, 9, 27–35. [Google Scholar] [CrossRef]
- Samac, D.A.; Tesfaye, M. Plant improvement for tolerance to aluminum in acid soils-A review. Plant Cell. Tissue Organ Cult. 2003, 75, 189–207. [Google Scholar] [CrossRef]
- Long, A.; Zhang, J.; Yang, L.T.; Ye, X.; Lai, N.W.; Tan, L.L.; Lin, D.; Chen, L.S. Effects of low pH on photosynthesis, related physiological parameters, and nutrient profiles of citrus. Front. Plant Sci. 2017, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baligar, V.C.; Fageria, N.K. Soil aluminum effects on growth and nutrition of cacao. Soil Sci. Plant Nutr. 2005, 51, 709–713. [Google Scholar] [CrossRef]
- Ribeiro, M.A.Q.; Almeida, A.-A.F.; Mielke, M.S.; Gomes, F.P.; Pires, M.V.; Baligar, V.C. Aluminum effects on growth, photosynthesis, and mineral nutrition of cacao genotypes. J. Plant Nutrition. 2013, 36, 1161–1179. [Google Scholar] [CrossRef]
- Baligar, V.C.; Schaffert, R.E.; Dos Santos, H.L.; Pitta, G.V.E.; Bahia Filho, A.F.D.C. Soil aluminium effects on uptake, influx, and transport of nutrients in sorghum genotypes. Plant Soil 1993, 150, 271–277. [Google Scholar] [CrossRef]
- Dogbatse, J.A.; Arthur, A.; Padi, F.K.; Konlan, S.; Quaye, A.K.; Owusu-Ansah, F.; Awudzi, G.K. Influence of acidic soils on growth and nutrient uptake of cocoa (Theobroma cacao L.) varieties. Commun. Soil Sci. Plant Anal. 2020, 51, 2280–2296. [Google Scholar] [CrossRef]
- Foy, C.D. Physiological effects of hydrogen, aluminum, and manganese toxicities in acid soil. In Soil Acidity and Liming; John Wiley & Sons, Ltd.: New York, NY, USA, 1984; pp. 57–97. ISBN 9780891182078. [Google Scholar]
- Bhuyan, M.H.M.B.; Hasanuzzaman, M.; Nahar, K.; Mahmud, J.A.; Parvin, K.; Bhuiyan, T.F.; Fujita, M. Plants behavior under soil acidity stress: Insight into morphophysiological, biochemical, and molecular responses. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches, 1st ed.; Springer: Cham, Switzerland, 2019; pp. 35–82. [Google Scholar] [CrossRef]
- Zhang, Y.-K.; Zhu, D.-F.; Zhang, Y.-P.; Chen, H.-Z.; Xiang, J.; Lin, X.-Q. Low pH-induced changes of antioxidant enzyme and ATPase activities in the roots of rice (Oryza sativa L.) seedlings. PLoS ONE 2015, 10, e0116971. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Di, T.; Xu, G.; Chen, X.; Zeng, H.; Yan, F.; Shen, Q. Adaptation of plasma membrane H+-ATPase of rice roots to low pH as related to ammonium nutrition. Plant Cell Environ. 2009, 32, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Iuchi, S.; Koyama, H.; Iuchi, A.; Kobayashi, Y.; Kitabayashi, S.; Kobayashi, Y.; Ikka, T.; Hirayama, T.; Shinozaki, K.; Kobayashi, M. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc. Natl. Acad. Sci. USA 2007, 104, 9900–9905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawaki, Y.; Iuchi, S.; Kobayashi, Y.; Kobayashi, Y.; Ikka, T.; Sakurai, N.; Fujita, M.; Shinozaki, K.; Shibata, D.; Kobayashi, M.; et al. STOP1 regulates multiple genes that protect Arabidopsis from proton and aluminum toxicities. Plant Physiol. 2009, 150, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arévalo-Gardini, E.; Farfán, A.; Barraza, F.; Arévalo-Hernández, C.O.; Zúñiga-Cernades, L.B.; Alegre, J.; Baligar, V.C. Growth, physiological, nutrient-uptake-efficiency and shade-tolerance responses of cacao genotypes under different shades. Agronomy 2021, 11, 1536. [Google Scholar] [CrossRef]
- Arévalo-Gardini, E.; Canto, M.; Alegre, J.; Loli, O.; Julca, A.; Baligar, V. Changes in soil physical and chemical properties in long term improved natural and traditional agroforestry management systems of cacao genotypes in Peruvian Amazon. PLoS ONE 2015, 10, e0132147. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods; CAB International: Wallingford, UK, 1993; Volume 78, ISBN 0851988210. [Google Scholar]
- Yuan, T.L. Determination of exchangeable hydrogen in soils by a titration method. Soil Sci. 1959, 88, 164–167. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Melo, L.C.; de Oliveira, J.P. Differential soil acidity tolerance of dry bean genotypes. Commun. Soil Sci. Plant Anal. 2012, 43, 1523–1531. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Souza Júnior, J.O. De Cacau: Cultivo, Pesquisa e Inovação; EDITUS: Ilheús, Bahia, Brasil, 2018. [Google Scholar]
- Baligar, V.C.; Fageria, N.K. Aluminum influence on growth and uptake of micronutrients by cacao. J. Food Agric. Environ. 2005, 33, 173–177. [Google Scholar]
- Tandzi, L.N.; Mutengwa, C.S.; Ngonkeu, E.L.M.; Gracen, V. Breeding maize for tolerance to acidic soils: A review. Agronomy 2018, 8, 84. [Google Scholar] [CrossRef]
- Ofori-Frimpong, K.; Afrifa, A.A.; Appiah, M.R.A. Growth and nutrient uptake of some cocoa varieties grown in contrasting soils. Ghana Jnl. Agric. Sci. 2002, 35, 41–48. [Google Scholar] [CrossRef]
- Siecińska, J.; Nosalewicz, A. Aluminium toxicity to plants as influenced by the properties of the root growth environment affected by other co-stressors: A review. Rev. Environ. Contam. Toxicol. 2017, 243, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Sposito, G. The Chemistry of Soils, 2nd ed.; Oxford University Press: New York, NY, USA, 2008; ISBN 9780195313697. [Google Scholar]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2011; ISBN 9780123849052. [Google Scholar]
- de Almeida, N.M.; de Almeida, A.A.F.; Mangabeira, P.A.O.; Ahnert, D.; Reis, G.S.M.; de Castro, A.V. Molecular, biochemical, morphological and ultrastructural responses of cacao seedlings to aluminum (Al3+) toxicity. Acta Physiol. Plant. 2015, 37, 5. [Google Scholar] [CrossRef]
- Meriño-Gergichevich, C.; Alberdi, M.; Ivanov, A.G.; Reyes-Díaz, M. Al3+-Ca2+ interaction in plants growing in acid soils: Al-phytotoxicity response to calcareous amendments. J. Soil Sci. Plant Nutr. 2010, 10, 217–243. [Google Scholar]
- Fageria, N.K.; Morais, O.P.; Carvalho, M.C.S.; Colombari Filho, J.M. Upland rice genotype evaluations for acidity tolerance. Commun. Soil Sci. Plant Anal. 2015, 46, 1076–1096. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Chapter 7 Ameliorating soil acidity of tropical oxisols by liming for sustainable crop production. Adv. Agron. 2008, 99, 345–399. [Google Scholar]
- Sodré, G.A.; Gomes, A.R.S. Cocoa propagation, technologies for production of seedlings. Rev. Bras. Frutic. 2019, 41, 1–22. [Google Scholar] [CrossRef]
- Roberto, A.; Gomes, S.; Sodré, G.A.; Guiltinan, M.; Lockwood, R.; Maximova, S. Supplying New Cocoa Planting Material to Farmers: A Review of Propagation Methodologies; Bioversity International: Rome, Italy, 2015; 189p. [Google Scholar]
- Kraiser, T.; Gras, D.E.; Gutié Rrez, A.G.; Gonzá Lez, B.; Gutié Rrez, R.A. A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 2011, 62, 1455–1466. [Google Scholar] [CrossRef]
- Rengel, Z. Books in soils, plants, and the environment CRCNET books. In Handbook of Soil Acidity; CRC PRESS: Boca Raton, FL, USA, 2003; Volume 94, 512p. [Google Scholar]
- Zhao, X.Q.; Guo, S.W.; Shinmachi, F.; Sunairi, M.; Noguchi, A.; Hasegawa, I.; Shen, R.F. Aluminium tolerance in rice is antagonistic with nitrate preference and synergistic with ammonium preference. Ann. Bot. 2013, 111, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.Q.; Shen, R.F. Aluminum–nitrogen interactions in the soil–plant system. Front. Plant Sci. 2018, 9, 807. [Google Scholar] [CrossRef] [Green Version]
- Baligar, V.C.; Fageria, N.K. Influence of nitrogen forms and levels on the growth and nutrition of cacao. J. Plant Nutr. 2017, 40, 709–718. [Google Scholar] [CrossRef]
- Hartemink, A.E. Nutrient Stocks, Nutrient Cycling, and Soil Changes in Cocoa Ecosystems: A Review. Adv. Agron. 2005, 86, 227–253. [Google Scholar]
- Marrocos, P.C.L.; Loureiro, G.A.D.A.; Araujo, Q.R.D.; Sodré, G.A.; Ahnert, D.; Escalona-Valdez, R.A.; Baligar, V.C. Mineral nutrition of cacao (Theobroma cacao L.): Relationships between foliar concentrations of mineral nutrients and crop productivity. J. Plant Nutr. 2020, 43, 1498–1509. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Thor, K. Calcium—Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Builes, V.H.; Küsters, J.; de Souza, T.R.; Simmes, C. Calcium nutrition in coffee and its influence on growth, stress tolerance, cations uptake, and productivity. Front. Agron. 2020, 2, 590892. [Google Scholar] [CrossRef]
- Husain, S.H.; Mohammed, A.; Ch’ng, H.Y.; Khalivulla, S.I. Residual effects of calcium amendments on oil palm growth and soil properties. In Proceedings of the IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 756, p. 012060. [Google Scholar]
- Calva, C.; Espinosa, J. Efecto de la aplicación de cuatro materiales de encalado en control de la acidez de un suelo de Loreto, Orellana. Siembra 2017, 4, 110–120. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| n | Wild Cacao Genotypes from the River Basins of Peruvian Amazon | Peruvian Grower’s Cacao Genotypes (ICT Collection) | Brazilian Cacao Genotypes | National and International Cacao Genotypes |
|---|---|---|---|---|
| 1 | AYP-15 | ICT-1112 | BN-34 | CCN-10 * |
| 2 | AYP-20 | ICT-1292 | BS-01 | CCN-51 |
| 3 | AYP-22 | ICT-2171 | CA-14 | EET-400 |
| 4 | PAS-91 | ICT-1092 | CEPEC 2002 | H-10 |
| 5 | PAS-93 | ICT-2172 | CP-2005 C10 | IMC-67 |
| 6 | PAS-100 | ICT-2653 | CP-49 C10 | POUND-12 |
| 7 | PAS-105 | ICT-2161 | CP-53 C10 | SCA-6 |
| 8 | NUC-156 | ICT-2142 | IPIRANGA-1 | ICS-1 |
| 9 | UGU-112 | ICT-1506 | PH-09 | ICS-6 |
| 10 | UGU-126 | ICT-1026 | PH-15 | ICS-39 |
| 11 | UGU-130 | ICT-2173 | PH-16 | ICS-95 |
| 12 | UNG-53 | ICT-1087 | PH-17 | TSH-565 |
| 13 | UNG-73 | ICT-1281 | PH-21 | TSH-1188 |
| 14 | UNG-76 | ICT-1251 | PH-144 | UF-613 |
| 15 | UNG-77 | ICT-1189 | PH-990 | UF-667 |
| Variables | Before Liming | After Liming |
|---|---|---|
| pH | 4.46 | 5.88 |
| EC dS/m | 0.08 | 0.37 |
| O.M.% | 1.77 | 1.57 |
| CaCO3% | <0.3 | <0.3 |
| N% | 0.08 | 0.07 |
| P mg kg−1 | 2.19 | 4.13 |
| Ca cmolc kg−1 | 1.22 | 3.37 |
| Mg cmolc kg−1 | 0.6 | 1.69 |
| K cmolc kg−1 | 0.19 | 0.25 |
| Al cmolc kg−1 | 2 | 0.00 |
| Effective CEC cmolc kg−1 | 4.44 | 5.31 |
| CEC pH 7.0 | 6.34 | 7.88 |
| Base saturation% | 31.70 | 67.38% |
| Al saturation% | 45.05% | 0.00% |
| Fe mg kg−1 | 141.1 | 110.3 |
| Cu mg kg−1 | 1.5 | 0.7 |
| Mn mg kg−1 | 6.2 | 4.4 |
| Zn mg kg−1 | 1.2 | 0.9 |
| Texture class | sandy loam | sandy loam |
| Variables | Units | Limed (n = 60) | Unlimed (n = 60) | p |
|---|---|---|---|---|
| Shoot length | cm | 50.74 ± 9.61 | 49.86 ± 10.11 | 0.3978 |
| Root maximum length | mm | 76.81 ± 16.48 | 67.81 ± 15.54 | <0.001 |
| Stem diameter | cm | 11.99 ± 1.41 | 11.40 ± 1.24 | <0.001 |
| Crown diameter | cm | 40.23 ± 4.02 | 40.02 ± 5.59 | 0.6937 |
| Number of branches | count | 7.07 ± 2.69 | 6.51 ± 2.79 | 0.059 |
| Number of leaves | count | 41.42 ± 11.99 | 38.02 ± 11.29 | <0.001 |
| Leaf area | cm2 | 1114.30 ± 310.80 | 875.10 ± 244.72 | <0.001 |
| Root area | cm2 | 364.61 ± 97.72 | 293.45 ± 74.01 | <0.001 |
| Shoot weight | g | 31.89 ± 5.22 | 29.48 ± 6.08 | <0.001 |
| Root weight | g | 12.27 ± 3.39 | 11.05 ± 3.13 | <0.001 |
| Shoot/root | ratio | 38.55 ± 14.97 | 27.77 ± 8.60 | <0.001 |
| gs * | mmol m−2 s−1 | 244.52 ± 115.93 | 232.46 ± 96.32 | 0.2835 |
| Chlorophyll | SPAD | 12.63 ± 3.25 | 12.35 ± 2.73 | 0.3841 |
| Variables | Unit | Concentration | ||
|---|---|---|---|---|
| Lime (n = 60) | Unlimed (n = 60) | p | ||
| N * | dag kg−1 | 1.08 ± 0.16 | 1.13 ± 0.16 | 0.300 |
| P | dag kg−1 | 0.08 ± 0.04 | 0.06 ± 0.03 | <0.001 |
| K | dag kg−1 | 2.31 ± 0.49 | 1.82 ± 0.5 | <0.001 |
| Ca | dag kg−1 | 1.51 ± 0.61 | 1.16 ± 0.21 | <0.001 |
| Mg | dag kg−1 | 0.84 ± 0.34 | 0.59 ± 0.1 | <0.001 |
| Cu | mg kg−1 | 5.38 ± 3.4 | 6.42 ± 2.44 | <0.001 |
| Fe | mg kg−1 | 112.36 ± 42.1 | 133.15 ± 56.19 | <0.001 |
| Mn | mg kg−1 | 120.7 ± 71.84 | 1172.32 ± 421.78 | <0.001 |
| Zn | mg kg−1 | 38.2 ± 16.41 | 52.95 ± 12.13 | <0.001 |
| Uptake | ||||
| N | mg plant−1 | 343.37 ± 79.23 | 334.47 ± 88.54 | 0.3152 |
| P | mg plant−1 | 24.09 ± 14.18 | 17.96 ± 7.82 | <0.001 |
| K | mg plant−1 | 674.09 ± 177.12 | 579.33 ± 194.56 | <0.001 |
| Ca | mg plant−1 | 484.6 ± 223.02 | 339.07 ± 79.04 | <0.001 |
| Mg | mg plant−1 | 269.15 ± 128.03 | 173.31 ± 45.56 | <0.001 |
| Cu | mg plant−1 | 1.7 ± 1.16 | 1.84 ± 0.7 | 0.144 |
| Fe | mg plant−1 | 35.78 ± 14.31 | 38.88 ± 18 | 0.072 |
| Mn | mg plant−1 | 39.28 ± 26.11 | 348.23 ± 143.71 | <0.001 |
| Zn | mg plant−1 | 12.07 ± 5.52 | 15.41 ± 4.04 | <0.001 |
| Nutrient Use Efficiency | ||||
| N | g shoot g−1 | 95.88 ± 19.66 | 91.23 ± 17.73 | 0.003 |
| P | g shoot g−1 | 1635.52 ± 911.82 | 1919.40 ± 1000.92 | <0.001 |
| K | g shoot g−1 | 59.87 ± 20.37 | 45.08 ± 8.74 | <0.001 |
| Ca | g shoot g−1 | 87.47 ± 65.15 | 88.40 ± 14.78 | <0.001 |
| Mg | g shoot g−1 | 148.86 ± 88.67 | 175.15 ± 29.41 | <0.001 |
| Cu | g shoot mg−1 | 274.56 ± 191.12 | 185.85 ± 111.65 | <0.001 |
| Fe | g shoot mg−1 | 10.47 ± 5.80 | 14.66 ± 74.19 | <0.001 |
| Mn | g shoot mg−1 | 11.37 ± 7.31 | 1.00 ± 0.53 | <0.001 |
| Zn | g shoot mg−1 | 29.65 ± 11.18 | 20.15 ± 6.88 | <0.001 |
| Tolerance Indices | Tolerant | Non-Tolerant |
|---|---|---|
| ATI * | ICT-1506, TSH-565, PAS-91, UGU-112, ICT-1281, TSH-1188, PAS-93, CCN-51, BN-34, PH-09, ICT-1251, PH-21, CP-49-C10, CA-14, CCN-10, EET-400, ICT-1189, PH-144, IMC-67, SCA-6, PH-15, AYP-22, ICT-2161, CP-53-C10, ICT-1087, ICT-1026, ICT-1112, UF-613, ICS-95, ICS-39, ICT-1292, UF-667, POUND-12, ICT-2142, BS-01, PAS-100, PAS-105, ICT-1092, ICS-6, CEPEC-2002, NUC-156, PH-16 | PH-17, UNG-53, AYP-15, H-10, ICT-2172, ICT-2653, ICS-1, CP-2005-C10, PH-990, ICT-2171, UNG-77, ICT-2173, IPIRANGA-1, UGU-130, UNG-73, AYP-20, UGU-126, UNG-76 |
| CATI | CCN-51, PH-21, CCN-10, PAS-91, ICT-1087, ICS-95, UF-667, TSH-565, PH-144, ICT-1189 | CA-14, PAS-93, PH-09, SCA-6, BN-34, ICT-1026, CP-49-C10, EET-400, PH-15, ICT-1281, ICS-39, BS-01, ICT-1506, TSH-1188, CP-53-C10, UGU-112, ICT-1112, ICT-1251, PH-990, ICT-2142, PH-16, IMC-67, AYP-22, AYP-15, UF-613, ICT-1292, ICT-2172, ICT-2161, ICS-6, ICT-2173, CEPEC-2002, UNG-53, ICT-2171, ICT-1092, PH-17, IPIRANGA-1, UNG-77, PAS-105, CP-2005-C10, H-10, PAS-100, ICT-2653, POUND-12, NUC-156, ICS-1, UNG-73, UGU-126, UGU-130, AYP-20, UNG-76 |
| GTI | ICT-1087, CCN-51, CCN-10, UF-667, PH-990, TSH-565, IPIRANGA-1, PH-21, PH-144, PAS-91, PH-16, ICT-1026, CP-49-C10, ICS-95, ICT-2171, PH-15, ICT-2173, PH-09, SCA-6, ICT-1189, ICT-2172, ICT-2142, UNG-77, BN-34, CA-14, AYP-15, ICS-39, ICT-1292, ICS-6, CEPEC-2002, BS-01, ICT-1112, PAS-93, CP-53-C10 | EET-400, CP-2005-C10, H-10, ICT-2161, UNG-53, TSH-1188, IMC-67, PH-17, ICT-2653, UNG-73, PAS-105, ICT-1281, UF-613, AYP-22, ICT-1506, ICT-1092, ICT-1251, ICS-1, POUND-12, UGU-112, UGU-126, PAS-100, NUC-156, UGU-130, UNG-76, AYP-20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arévalo-Hernández, C.O.; Arévalo-Gardini, E.; Farfan, A.; Amaringo-Gomez, M.; Daymond, A.; Zhang, D.; Baligar, V.C. Growth and Nutritional Responses of Juvenile Wild and Domesticated Cacao Genotypes to Soil Acidity. Agronomy 2022, 12, 3124. https://doi.org/10.3390/agronomy12123124
Arévalo-Hernández CO, Arévalo-Gardini E, Farfan A, Amaringo-Gomez M, Daymond A, Zhang D, Baligar VC. Growth and Nutritional Responses of Juvenile Wild and Domesticated Cacao Genotypes to Soil Acidity. Agronomy. 2022; 12(12):3124. https://doi.org/10.3390/agronomy12123124
Chicago/Turabian StyleArévalo-Hernández, César O., Enrique Arévalo-Gardini, Abel Farfan, Martin Amaringo-Gomez, Andrew Daymond, Dapeng Zhang, and Virupax C. Baligar. 2022. "Growth and Nutritional Responses of Juvenile Wild and Domesticated Cacao Genotypes to Soil Acidity" Agronomy 12, no. 12: 3124. https://doi.org/10.3390/agronomy12123124
APA StyleArévalo-Hernández, C. O., Arévalo-Gardini, E., Farfan, A., Amaringo-Gomez, M., Daymond, A., Zhang, D., & Baligar, V. C. (2022). Growth and Nutritional Responses of Juvenile Wild and Domesticated Cacao Genotypes to Soil Acidity. Agronomy, 12(12), 3124. https://doi.org/10.3390/agronomy12123124