Abstract
Shoot apical meristem (SAM) consists of stem cells that act as a reservoir for the aerial growth. It plays an important role in the differential architectural development in plants. SAM actively performs parallel functions by maintaining the pluripotent of stem cells and continuous organogenesis throughout the plant’s life cycle. Molecular mechanisms regulating the signaling networks of this dual function of the SAM have been progressively understood. In the SAM, the feedback loop of WUSCHEL (WUS)-CLAVATA (CLV) has been found to be the key regulator in stabilizing stem cell proliferation and differentiation. In general, WUS migrates into central zone (CZ) from organizing center (OC) and activates the expression of CLV3 by binding to the promoter elements. CLV3 acts as a ligand to interact with the CLV1, leucine rich repeats (LRR) receptor-like kinase (RLK) and LRR receptor-like protein CLV2, and protein kinase coryne (CRN) (CLV2/CRN) to restrict WUS transcription to the OC. Evolution of CLV3 is one of the main factors contributing to the transformation of two-dimensional (2D) to 3D plants. WUS-CLV loop is involved in several pathways and networks that integrate on meristem maintenance and cell identity transition. WUS-CLV maintains stem cells with simultaneous differentiation signals by the spatial-temporal signaling of the phytohormones. WUS-CLV loop has an interaction with reactive oxygen species (ROS), an important signaling molecules regulating cell proliferation and developmental transition. WUS also forms feedback loop with AGAMOUS (AG) for differentiation, proliferation, and termination of floral meristem. These loops might also involve in interaction with vernalization and its regulatory factors that oversees the precise timing of flowering after exposure to cold temperatures. In this review, we highlight the evolutionary and developmental importance of the WUS-CLV feedback loop on SAM maintenance and cell identity transition for inflorescence and floral meristem development.
1. Introduction
Plants encompass a small population of stem cells at both shoot apical meristem (SAM) and root apical meristem (RAM) throughout their life cycle. Stem cells are pluripotent and self-renewing [1]. Stem cells have the ability to differentiate into various types of cells; meanwhile, daughter cells will be maintained as stem cell in the ‘niche’. Stem cells present in both the shoot and root apices are specified during the embryogenesis stage. In early stages, it is mostly in an ordered pattern, whereas, in the later developmental stages, stem cell fate has more of a spatial and a regulatory-dependent pattern [2]. Stem cells has the ability to renew themselves. One set of cells remains as stem cells (undifferentiated) and other set differentiate into organs such as leaves, flowers, and stems indicate intrinsic complex networks of the stem cell niche [3]. All pathways responsible for formation or maintenance of stem cells are extensively regulated by phytohormones, such as cytokinin (CK), auxin, transcription factors (TFs), and external environmental stimuli [1]. These cells are distributed in different kinds of meristems, including the embryonic and post-embryonic meristem. SAM and RAM are specified in the embryogenesis [4] and activated at favorable conditions. The post-embryonic meristem is the tissue formed after the development of the embryo, including the intercalary meristem and lateral meristem. SAM and RAM are the primary meristems, forming stem cells responsible for formation of the whole plant. Whereas, intercalary and lateral meristems are the secondary meristems, accountable for internode, internal tissue layers and other lateral growth of the plant [2,3].
Stem cells with various differentiation potencies are present at the shoot and root apex. RAM gives rise to entire root system whereas SAM consists of a pool of pluripotent stem cells that give rise to the above-ground parts of the plant [1]. Stem cells attain different identities due to the interactive effects of various gene regulatory pathways. SAM is structurally organized into three clonal layers: L1 (epidermal) layer, L2 (sub-epidermal) layer, and L3 (innermost corpus) layer(s). Outer L1 and L2 layers are called as tunica. Zones that are allocated radially are named as the central zone (CZ), containing pluripotent stem cells; peripheral zone (PZ), containing transit-amplifying stem cells; and organizing center (OC) that organizes and maintains the stem cell population [1,3]. The CZ contains pluripotent stem cells and the transit amplifying cells within the PZ are recruited to form the organ (Figure 1).
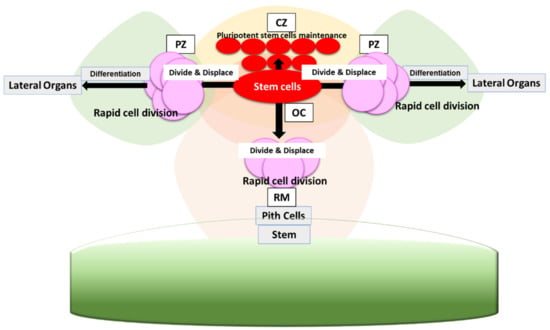
Figure 1.
Stem cell organization in SAM. Stem cells present in the central zone (CZ) provides mainly two subsets of daughter cells. The one subset of cells rarely divides and maintains the stem cell pool. The other subset of cells rapidly divides and differentiates in peripheral zone (PZ) to give rise to lateral organs. Organizing center (OC) maintains stem cell population. Cells that are displaced basally to the rib meristem (RM) give rise to pith cells, eventually forming stem.
Both the SAM and floral meristem (FM) determine the size, number, and shape of agronomic traits such as the flowers, fruits, seeds, kernels, etc [5]. A key regulatory pathway involved in the maintenance of the stem cell population and specification of stem cell identity in the SAM is the WUSCHEL (WUS)–CLAVATA (CLV) feedback loop. The WUS-CLV is a feedback loop/system that plays a crucial role in maintaining stem cell homeostasis as well as organ development [1,2,3,4,5]. Post-embryogenic development occurs from the apical meristem present in shoot and root tips. Leaves, inflorescences, and floral organs are differentiated from the SAM [4,5]. It is interesting to note that WUS-CLV pathway has been evolutionarily conserved across land plants [6]. This pathway is extensively studied in the model plant Arabidopsis thaliana and also in maize, rice, and tomato. WUS activity is required for the identity, structural, and functional integrity of the SAM and FM. The WUS and CLV genes are mandatory for the progressing meristem cells toward organ initiation [7,8]. In tomato, orthologs of the WUS and CLV genes have been elucidated as the determining factor for the carpel number, floral growth, and fruit size [9,10]. A study on the WUS showed it plays a major role in the course of monocot and dicot evolution [11]. Previous reports on maize suggested that crop yield and plant architecture are strongly influenced by WUS-CLV orthologs [12,13,14]. The importance of WUS and CLV on floral organ numbers, subfunctionalization, and diversification was proved in rice [15,16,17].
The following table shows the orthologs of the Arabidopsis genes with their functions (Table 1).

Table 1.
WUS, CLV, and AGAMOUS (AG) orthologs in tomato, maize and rice.
It is curious to know that, in the meristem, a small set of populations renew themselves and the rest undergoes differentiation to form the new organs (Figure 1). Though several studies were carried out in the model plants as well as crops, still some of the areas in stem cell research have to be addressed to open new avenues in the near future such as (i) transcriptional networks involved in defining the cell types, (ii) regulatory factors involved in determining stem cell fate, and (iii) cell identity transition.
In this review, we present the paradigm of the WUS-CLV feedback loop in the stem cell niche and framework of the identity transition, floral development, and phytohormones cross-talk. Importantly, ROS on regulating the SAM and also genetic network of vernalization in meristematic tissue are discussed.
2. WUS-CLV Feedback Loop in SAM Maintenance
WUS-CLV signaling is crucial for the development of meristem and considered to play a major role in crop domestication. Evolution, proliferation, and differentiation of stem cells in the SAM are synchronized by a primary auto-regulatory WUS-CLV signaling pathway [3]. WUSCHEL-related homeobox (WOX) encodes the homeodomain TF. Rodriguez et al. reported that last 63-aa stretch present at the C-terminus of Arabidopsis holds transcriptional regulatory domains namely acidic region, WUS-box and ethylene-responsive element binding factor-associated amphiphilic repression (EAR-like) domain. WUS-box is required for the nuclear retention of WUS, while EAR-like domain is responsible for its nuclear export, as it resembles the nuclear export signals (NESs). EAR-like domain determines high levels of the WUS inside the nucleus in the cells of the rib meristem (RM) whereas lower nuclear levels in adjoining cells. Additionally, WUS is constricted by the N-terminal DNA binding homeodomain and homodimerization sequence present in the central part of the protein which are responsible for homodimer formation [23]. WUS move between the cells via plasmodesmata. Subsequent to the migration to the outer layer with the activation of its negative regulator, WUS restrict its own accumulation [24].
A study of wus mutants showed that the SAM failed to generate proper leaf primordial and FM terminate prematurely [7]. On the contrary, ectopic expression of WUS trigger pluripotency which results in altered SAM, activate shoot stem cell marker in root as well as induction of shoot tissue from root, and initiated new vegetative buds in Arabidopsis [25,26,27]. WUS and hairy meristem (HAM) family of GRAS domain transcriptional regulator controls stem cell production and ensure that CLV3 transcription is activated only in the outermost apical layers of the SAM. Activation of CLV3 by WUS occurs only in the absence of HAM [28].
CLV3 is a member of the clavata 3/embryo surrounding region (CLE) family of the polypeptide, formed as a pre-propeptide in the cells of L1 and L2 that encode 96 amino acid, processed into 12 amino acid or arabinosylate 13 amino acids peptides [29]. All members of the CLE protein family contain a conserved 14 amino acids sequence motif, termed as the CLE motif [30]. CLV1 encodes leucine-rich repeat (LRR) receptor-like kinase (RLK) and CLV2 encodes LRR receptors-like protein (RLP), and the CORYNE (CRN) protein encodes membrane-associated receptor-like cytoplasmic kinase (RLCK) (CLV2/CRN) form complex with the CLV3 peptide [4,5]. According to the signaling cascade, CLV1, CLV2/CRN receptors recognize the CLV3 peptide and are activated to restrict the expression of the WUS to a small subset of cells in the center (Figure 2).
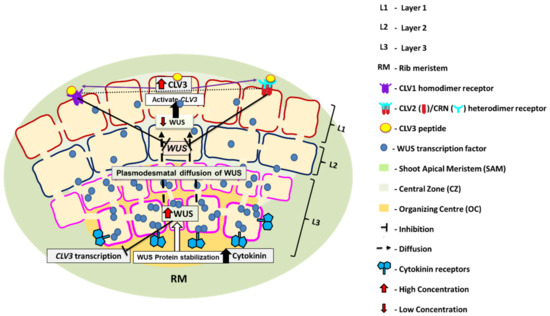
Figure 2.
WUS-CLV feedback loop. WUS is mainly expressed in OC-containing cells of RM and L3. WUS acts non-autonomously in L1 and L2. High levels of WUS are maintained by high cytokinin concentration and absence of CLV3 suppression on WUS transcription. At higher concentration of WUS, it represses CLV3 transcription in L3 and RM. WUS concentration decreases as it moves towards the apex because of the absence of cytokinin in L1 and L2. At low concentration of WUS, CLV3 transcription is activated. CLV3 then represses the transcription of WUS in L1 and L2 by interacting with homodimer of CLV1 and heterodimer of CLV2/CRN, receptors of CLV3. In this way, WUS-CLV maintains a feedback loop. CLV, CLAVATA; WUS, WUSCHEL.
The WUS acts as an inducer of the CLV3 at its low concentration, while at high concentration it represses the CLV3 by binding to the cis-regulatory elements of the CLV3 gene [23,31]. WUS forms homodimers and inhibit CLV3 [31]. It was hypothesized that formation of homodimer could limit the mobility of WUS [32]. While in L1 and L2, due to the low WUS concentration, it remains in its monomeric form and binds to the cis-regulatory elements of the CLV3 gene, but, this time, it activates CLV3 expression. Therefore, the WUS gradient determines the activation or repression of the CLV3, thus forming the feedback loop [31]. Transcription of the WUS is inhibited by the activated CLV3 and its receptor complexes after the subsequent CLV3 signaling cascade formation. Fate of the stem cell is entirely governed by this regulatory feedback loop [33]. Loss of clv function leads to the enlarged meristem might be due to the increase in rates of cell division in the meristem and cells on the boundary of CZ-PZ, remains as CZ cells after division [34].
Therefore, the WUS-CLV is inevitable on the stem cell niche in SAM maintenance and inflorescence transition for reproductive development. There is further involvement of the WUS-CLV on the reactive oxygen species (ROS) signaling and vernalization for cell proliferation and organ development. This review gives an overview on the regulatory mechanisms of WUS-CLV in cellular differentiation and cell identity transition.
3. WUS-CLV in Identity Transition
The WUS concentration plays a decisive role in stem cell homeostasis. CLE peptides are vital for SAM maintenance and differentiation. [30] WUS concentration regulates the transcription of CLV3 through an activation or repression switch. The CLV3 inhibits the WUS transcript in upper layers. The WUS is restricted to the center of the OC while CLV3 has a high expression in outermost layers. Regulation of WUS synthesis and its subcellular partitioning by the CLV3 maintains a nuclear WUS concentration in-turn regulating CLV3 levels. This regulatory network fine-tunes the WUS accumulation [23,35]. Expression of CLV3 in apex prevents conversion of CZ into PZ cells in the surrounding area of CZ [34]. This gradient-dependent expression or repression determines the maintenance and differentiation of stem cells, ultimately leading to the transition from the vegetative to reproductive phase.
Whitewoods et al. reported the role of the conserved CLV and RLK pathways in the switch from the two-dimensional (2D) to 3D growth of land plants by comparing moss, Physcomitrella, and Arabidopsis. Modern mosses have leafy shoots that grow from a filamentous (2D) precursor. Genome data indicated the appearance of core elements of the CLV pathway in the last common ancestor of land plants, together with the evolutionary innovation of 3D growth. Their experiments showed that Physcomitrella CLE functions similarly to CLV3 in Arabidopsis in regulating growth and proliferation through the conserved receptor machinery [6]. Further comparative studies between plants with different inflorescence structures such as Arabidopsis, rice, and maize showed the existence of homologs with similar functions for SAM maintenance and inflorescence meristem (IM) transition among plant lineages [36]. CLV3 is also recognized by the CLV1-related barely any meristem (BAM) LRR-RLKs and receptor-like protein kinase 2 (RPK2) via BAM1 [5,6]. Arabidopsis clv1/bam1/bam2/bam3 quadruple mutants showed strongly disordered planes in the stem cell niche and ground tissue layers of mutant roots [6]. BAM1, BAM2, and BAM3 promote shoot and flower meristem. It was suggested the BAM genes can play complementary or opposite role of CLV1 [37]. It is also clear that BAM regulated by CLV1 and interaction of CLV1 with WUS is distinct to each other [38]. During the Physcomitrella gametophore there was a high dynamic expression of CLEs, CLVs, and RPK2 genes, and hence it can be deduced that CLEs also act via RPK2 in regulating the 3D growth [6]. However, still the inflorescence complexity needs to be explored by comparative analysis between different plant species.
In shoot meristem, CLV1/BAM1 and CLV2/CRN complexes act in separate but in the parallel pathways. For the fruit development, CLV1, CLV2, and CRN receptors all act in a linear pathway [39]. WUS also involved in FM determinacy through epigenetic mechanisms [40]. In this context, Sun et al. [41] recently showed that the WUS is repressed by AGAMOUS (AG) via KNUCKLES (KNU) through histone deacetylation at floral stage 6. An earlier study revealed that the WUS is expressed in immature stomium cells and is associated with anther development. As a result of this interaction, anthers of wus mutants had fewer and malformed lobes compared to the wild type, showing that the WUS is essential for normal anther development [42]. Another study in Chrysanthemum morifolium also indicated that the WUS interacts with cycloidea 2 (CYC2) TF to regulate reproductive organ development [43].
Flowering of plants starts with the transition of the SAM into the IM. Further differentiation of the FM is regulated by environmental cues. From the indeterminate shoot to the development of determinate floral organs such as sepal, petal, stamen, and carpel, cell identity transition and stem cell reprogramming occur with various level of developmental regulation in the apical region [44,45]. Therefore, the regulation of the FM by WUS, CLV, and AG is a dynamic switch that ensures successful reproduction.
4. WUS-CLV in Flower Development
Flowers are formed from the proliferating SAM where the meristem size depends greatly on the WUS-CLV feedback loop. Unlike WUS, shoot meristemless (STM) express throughout the meristematic region. STM is mainly required for maintenance of undifferentiated cells in SAM and FM. For SAM fate, STM can be used as marker gene as its expression is extinguished during the leaf primordia specification [46,47]. Aintegumenta (ANT) is specific to the floral organogenesis and patterning of other regulatory genes [48]. In floral primordia, expression of ANT is found in peripheral regions on stage 0 and 1 [47]. It was observed that mutant of clv showed supernumerary floral structure [49] and wus mutant terminate prematurely [7]. This observation suggests that the WUS-CLV loop plays an important role in floral organ differentiation rather than the transition.
Termination of the FM occurs at stage 6, in a WUS-AG dependent manner to ascertain proper development of the floral organs [50]. AG is a C-class MADS domain-containing TF, positively regulating FM determinacy [51]. The ag mutants exhibit indeterminate nature in FM containing interior flowers inside third whorl [52]. Induction of AG by the WUS is dependent on the leafy (LFY). It is mandatory that WUS required LFY to acts on cis-regulatory element of AG to confer floral identity in the FM after the transition from the IM. During the formation of the FM, WUS together with LFY binds sequences to activate AG expression. AG only mildly represses the WUS after floral stage 3 [50,51,52]. For the termination of the WUS at stage 6 after carpel initiated, KNU encodes C2-H2 zinc-finger TF plays a pivotal role in FM determinacy [53]. AG completely represses WUS transcription through KNU [54] and also directly through the terminal flower 2 (TFL2)-AG complex that brings chromatin loop formation at the WUS locus [55]. Thereby terminating stem cell activity and start determinate floral growth. WUS is required in developing stamens at stages 7–8 [51] and in developing ovules [56]. It can be seen that the AG-WUS and WUS-CLV pathways function are at least partially independent since the effects of ag and clv1 mutations on FM determinacy are additive. Consistently, WUS expression increased strongly in ag-1 clv1-4 in comparison to ag-1 [50]. Thus, this complex regulatory network is mandatory for FM development to produce flowers with specified numbers and whorls of floral organs.
5. Phytohormones Crosstalk with WUS-CLV Regulatory Pathway in SAM
Plant hormone levels are modulated during various developmental stages causing differential expression of SAM maintenance, cell identity, and cell fate transition-related genes. Phytohormones play a major role in cell division and differentiation during organogenesis in plants. Spatial and temporal signaling of auxin is dynamic to specify distinct cell identities in the SAM and organogenesis in a concentration-dependent manner [4,5]. Auxin accumulates at the positions of floral anlagen through the polar transport by pinformed 1 (PIN 1) [57]. Auxin maxima then activates genes that lead to organogenesis. The model for auxin-mediated organogenesis in the SAM is based on the inhibitory field theory. According to this theory, pre-existing organs restrict the initiation of new organs in their vicinity by chemical inhibitors. Also, polar auxin transport deplete auxin around organs along to trigger organ initiation [58].
Auxin and CK play antagonistic roles in division and differentiation at the shoot apex. In the SAM, auxin maxima induce cellular differentiation and organ outgrowth at the site of the primordial formation, whereas CK maxima are found to undifferentiated state in SAM [59]. The RM-enriched CK stabilizes the WUS protein by acting on the nuclear retention signal. The WUS is stabilized through the CK signaling action on the acidic domain and the WUS-box. Stability of the WUS is achieved when the degron activity of WUS-box is masked by the combined effect of the nuclear retention machinery and the CK signaling to regulate CLV3 transcription [60]. The cells in the PZ have a high auxin concentration, and therefore they start to differentiate, whereas cells in the OC can maintain stem cell properties because of the presence of the WUS and CK [61]. WUS limits auxin signaling through histone acetylation, preventing cells at the CZ from differentiating [62]. Previously formed leaf primordia act as a sink of auxin. It can travel to certain distance for phyllotaxis position and protrusion [63]. The WUS represses the expression of type-A ARR genes such as ARR5/6/7/15 to increase the CK activity in the OC [64]. WUS protein stabilization depends on the CK concentration until WUS can be independently stable [60]. The CK receptors such as ARABIDOPSIS HISTIDINE KINASE 2/3/4 (AHK2/3/4) play an important role in CK response by sensing the environment [6,65]. Crosstalk between auxin and CK is necessary for SAM maintenance. Auxin response factor 3 (ARF3) inhibit CK for FM determinacy [66] whereas auxin response factor 5 (ARF5)/monopteros (MP) repress type-A ARRs such as ARR7/15, a negative regulator of CK [67]. This process fine tune auxin and CK level. Moreover, by repressing the expression of target genes, WUS restricts auxin-mediated cell differentiation which enhances the CK-mediated WUS expression. Auxin through ARF5/MP represses (at least partly) the expression of the dornroschen/enhancer of shoot regeneration 1 (DRN/ESR1). DRN strongly regulate CLV3 which proved to interfere with WUS-CLV feedback loop [61]. Changes in the auxin and CK signaling could affect the behaviour of stem cell fate. The interaction between the WUS-CLV loop and auxin and CK clearly highlights strong relations between stem cell maintenance and hormone signaling (Figure 3).
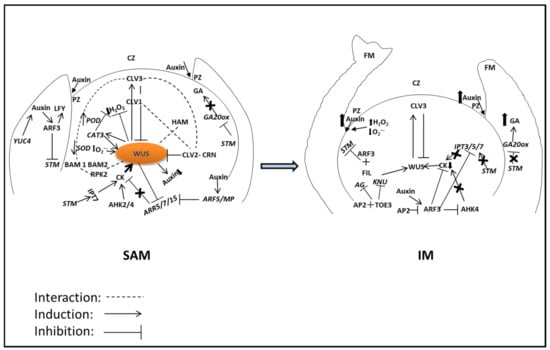
Figure 3.
Regulatory network of WUS-CLV feedback loop in SAM maintenance and its transition to the inflorescence meristem (IM). Genes involved in the SAM, IM, and floral meristem (FM) are spatial and temporally regulated by several factors including phytohormones. AG, AGAMOUS; AHK, ARABIDOPSIS HISTIDINE KINASE; AP, APETALA; ARF, AUXIN RESPONSE FACTOR; ARR, ARABIDOPSIS RESPONSE REGULATORS; BAM, BARELY ANY MERISTEM; CAT, CATALASE; CK, CYTOKININ; CLV, CLAVATA; CRN, CORYNE; FIL, FILAMENTOUS; GA, GIBERELLIC ACID; GA20OX, GA20 OXIDASE; H2O2, HYDROGEN PEROXIDE; HAM, HAIRY MERISTEM; IPT, ISOPENTENYL TRANSFERASE; KNU, KNUCKLES; LFY, LEAFY; MP, MONOPTEROS; O2•−, SUPEROXIDE; POD, PEROXIDASE; RPK, RECEPTOR-LIKE PROTEIN KINASE; SOD, SUPEROXIDE DISMUTASE; STM, SHOOT MERISTEMLESS; TOE, TARGET OF EAT; WUS, WUSCHEL; YUC, YUCCA. CZ, central zone; PZ, peripheral zone. Other regulatory factors/genes have been avoided due to its complexity and out of focus of this review.
Along with auxin, gibberellin (GA) is another key hormone in connection with cell growth and differentiation. Higher level of auxins and GA are required for outgrowth at the flanks of SAM. It regulates phyllotaxis, floral transition, and flowering [68]. Suppression of GA biosynthesis and induction of CK are carried out by KNOX TFs including KNOTTED1 (KN1) in maize, STM in A. thaliana, leading to the inhibition of differentiation in the CZ of the SAM [69]. STM also induces isopentenyl transferase 7 (IPT7) which increase the cytokinin level for WUS maintenance [70]. Chang et al. reviewed the role of ARF3 in induction of LFY and YUCCA4 (YUC4) to increase auxin that can leads to differentiation in PZ [4]. MP and FIL indirectly and ARF4 and ARFs ETTIN (ETT)/ARF3 directly reduces expression of STM to promote flowering [71]. For FM initiation, APETALA 2 (AP2) and TARGET OF EAT 3 (TOE3) inhibits AG for floral patterning [4,72]. GA level is reduced in SAM at CZ by KNOX. Suppression of GA signaling is achieved by two functions of the KNOX TF. Primarily, KNOX suppressing GA20 oxidase is involved in the biosynthesis of GA and, secondly, by enhancing the activity of GA2 oxidase which in turn is responsible for inactivation of active forms of GAs. The low levels of GA are required for undifferentiated vegetative meristematic activity, whereas its high level supports floral transition [73]. AGAMOUS-LIKE15 (AGL15) regulate endogenous GA levels by regulating GA2ox6 [74]. During floral transition GA levels are generally high based on spatial- and –temporal expression of GA20ox2 [69]. Interaction with brassinazole-resistant 1 (BZR1) and DELLA was required by both brassinosteroids and GA3 to regulate each other. DELLA mediates various hormonal and signaling pathways. [75]. Bao et al. [68] summarized GA signaling as the main component of the crosstalk between different hormones during FM development and its interaction with LFY, flowering locus d (FD), flowering locus T (FT) and suppressor of the overexpression of constans 1 (SOC1), involved in the process. As the WUS interacts with LFY [52], these hormones may also have their effects on WUS expression directly or indirectly. Jasmonic acid (JA) is also involved in plant developmental processes but majorly in defense-related responses [76]. MYC2, MYC3, and MYC4 plays major role in most of the JA-dependent regulation processes [77]. For the JA-mediated inhibition of flowering above three TFs are most vital [78].
Ethylene is another vital hormone participating in the stress response, senescence, regulation of floral transition, and fruit ripening. JA and ethylene were found to induce the expression of HISTONE DEACETYLASE 6 (HDA6) and HDA19 [79,80]. Especially, HDA6 is vital for the deacetylation of FLC chromatin [79]. This chromatin remodeling is essential for repression of FLC. Abscisic acid (ABA) acts antagonistically to GA, as GA promotes seed germination, while ABA restricts it [81]. Abscisic acid (ABA)-insensitive mutant 5 (ABI5), a basic leucine zipper (bZIP) transcription factor, is involved in floral transition in Arabidopsis. Induction of ABI5 could delay flowering initiation via upregulating FLC expression [82] whereas ABA-dependent GIGANTEA signaling induces expression of FT which accelerates flowering to escape drought conditions [83].
Only few reports are available on the direct interaction of phytohormones such as GA, ethylene, and ABA with the WUS-CLV pathway. Further research needs to be conducted with respect to hormonal signaling in the SAM involving cell identity genes which would fetch information on how hormonal levels and their signaling alter the SAM maintenance and development by affecting major genes involved the WUS and CLV.
6. ROS in SAM Regulation
Similar to the phytohormones, ROS are also vital for the stem cell maintenance and differentiation. Redox state decides the stem cell fate. Especially low proliferation of OC could be overlapped with the redox status [84]. ROS such as super oxide (O2•−), hydrogen peroxide (H2O2), hydroxyl ion (•OH), etc. are a natural by-product generated during the normal aerobic metabolism process. Excessive accumulation of ROS damages macromolecules and nucleic acids. Equilibrium of ROS generation and oxidation was maintained by the antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), peroxidases (POD) etc. [85]. ROS act as a secondary messengers in regulating the balance between cell division and differentiation. ROS status is one of the determining factors for cell fate.
Recent findings by Zeng et al. [86] showed that equilibrium between the H2O2 and O2•− regulate WUS transcription. A high concentration of O2•− maintains WUS expression to inhibit differentiation of stem cells in OC, while higher signals of H2O2 in the PZ repress WUS for differentiation. It can be inferred that ROS direct stem cell fate, and the balance between O2•− and H2O2 is crucial for SAM maintenance and differentiation. Wang et al. established a putative model for ROS-mediated regulation of age-dependent developmental programmed cell death (dPCD) involving the WUS-CLV pathway. At the proliferative stage, WUS expression is maintained by O2•− and also WUS might promote CAT3 to keep the H2O2 in lesser concentration. WUS expression averted the activation of the ORESARA 1–BIFUNCTIONAL NUCLEASE 1 (ORE1-BFN1) senescence cascade, hence preventing dPCD of stem cell. Promoting the CAT3 and repressing acyl-CoA oxidase 1 (ACX1) helps to maintain low level of H2O2. Production of O2•− persuade WUS to maintain stem cells. In contrast, CLV3 could inhibit O2•− for proliferation [87]. ROS play a critical role in abiotic and biotic responses to plant development and maintaining homeostasis [88]. Recent studies mentioned that spatial distribution of the O2•−/H2O2 balance serves as a key switch for stem cell maintenance and differentiation. Antioxidant enzymes such as SOD and POD plays a major role in keeping ROS under controlled level [86,87]. Decrease in the O2•− and increase in H2O2 could lead to the differentiation rather than stem cell maintenance [86]. These results reveal that the WUS-CLV participates in the determination of stem cell fate through ROS, as O2•− promotes WUS activity and stem cell maintenance, and H2O2 induces differentiation. Due to this interaction of the WUS-CLV with ROS, it can be stated that the pathway articulates cell proliferation and differentiation homeostasis and involved in thermomorphogenesis.
7. Vernalization
Ambient temperature is the most crucial for the precise timing of flowering. For instance, several Brassicaceae plants need to be exposed to low temperatures (few weeks) for flowering which is generally termed as vernalization. Appropriate vernalization triggers genetic and developmental changes in the SAM resulting in the transition to the IM and further development of either terminal or lateral FMs. Flowering in response to vernalization is not immediately occur in low temperatures instead it accelerates flowering when the low temperature is removed, i.e., in the warm temperatures of the following spring after winter [89].
The molecular genetic network of vernalization in Arabidopsis majorly involves suppression of flowering locus C (FLC) during cold exposure leads to flowering [90]. Frigida (FRI) repress the flowering is correlated with its induction of FLC gene. However, natural allelic variation exists between these two key genes contribute for difference in flowering time [91]. FRI act as a scaffold protein that holds the FRI-LIKE 1 (FRL 1), FRI essential 1 (FES 1), suppressor OF FRI 4 (SUF 4), and FLC expressor (FLX) to form a transcription activator complex for increasing the transcription of FLC [92]. FLC repress transcription of flowering locus D (FD), FT, and SOC1 resulting in delayed flowering. It was reported that FLC repressed by the cold exposure [89,93]. It is involved with several pathways such as prolonged cold exposure, photoperiod via CONSTANS (CO), and autonomous pathways to suppress the FLC gene. In exposure to cold, activation of VRN1 and increase in VIN3 expression and association with VIL1/VRN5 induce chromatin repression of FLC [93,94,95]. This silencing involves two repressive histone modifications such as trimethylation of Lys9 and Lys27 of histone H3 (H3K9me3 and H3K27me3) resulting in stable epigenetic repression of FLC in response to cold temperatures. Before vernalization, high levels of FLC block the floral transition in winter-annual strains of Arabidopsis as its chromatin is enriched with active histone marks [93]. To prove that Yang et al. reported that H3K36me3 and H3K27me3 rarely coexist [96]. During vernalization, it can be considered that main factor of FLC gradual silencing could be due to increasing H3K9me3 and H3K27me3 (repressive histone marks) and decreasing H3K4me3 and H3K36me3 (active histone marks) at the nucleation level of FLC (Figure 4). Meanwhile, GA activates the promoter of LFY [97]. Simultaneously, SOC1 induced by both GA and FD and FT [93]. SOC1 and AGL24 possessed positive regulation to each other which is required for LFY activation [98].
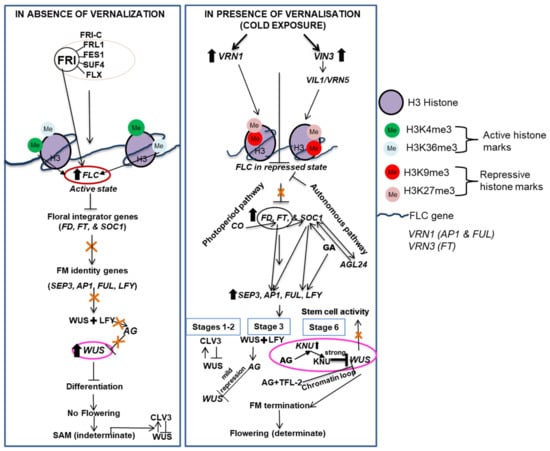
Figure 4.
Simple heuristic model of the FRI-mediated FLC activation and vernalization-induced flowering pathway of Arabidopsis. In the absence of vernalization, FLC remains activated by a scaffold protein known as FRI. Because of the allelic variation in FLC and FRI, there is existence of natural variation in the requirement of vernalization. It must be noted that not all plants required to exhibit above mentioned model. AG, AGAMOUS; AGL24, AGAMOUS-LIKE 24; APETALA 1 (AP1); CLV, CLAVATA; CONSTANS (CO); FES 1, FRI essential 1; FLC, flowering locus C; FD, FLOWERING LOCUS D; FLX, FLC expressor; FRI, frigida; FRL 1, FRI-LIKE 1; FUL, FRUITFUL; FT, FLOWERING LOCUS T; LFY, LEAFY; SEP, SEPALLATA; SOC1, SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1; SUF 4, suppressor OF FRI 4; TFL, TERMINAL FLOWER; VRN1, VERNALIZATION 1; VIN3, VERNALIZATION INSENSITIVE; VIL1, VRN5/VIN3-LIKE 1; WUS, WUSCHEL; H, Histone; K, Lysine; me3, trimethylation.
Repression of FLC is stable even after the cold exposure is over. However, this repression is only stable throughout mitosis, and FLC is activated again. It transiently expressed during the gametogenesis, and mid-to-late embryogenesis [93,99,100]. During cold, stable silencing of FLC is mediated by H3K27me3. In the following warm period, epigenetic genetic process opposing the histone modification such as silencing H3K27me3 quantitatively based on the cold exposure period [101]. Active chromatin-modifying complex for epigenetic reprogramming and FRI-C and PAF1 complexes, are crucial for the reactivation of FLC [101,102].
Vernalization oversees the precise timing of flowering majorly through FLC repression which activates the floral pathway integrator genes such as FD, FT and SOC1. This in turn upregulates the FM identity genes such as SEPALLATA3 (SEP3), FRUITFUL (FUL), APETALA 1 (AP1), and LFY [93]. As the stages of flower development progress, stage 1–2 WUS-CLV loop is active, in stage 3 WUS induce AG along with the activated LFY, and, at stage 6, AG indirectly represses the WUS through KNU and also directly with TFL-2 to modulate chromatin loop for the FM termination [51]. Hence, it can be proposed that a direct/indirect relation can be established between vernalization and the WUS-CLV loop (Figure 4). The loop maintains stem cells in the SAM in vegetative state could also guides them for further organ development, transition, and termination. However, more research on this area needs to be conducted to elucidate the role of the WUS-CLV in vernalization, repression and reactivation of FLC.
8. Conclusions
Functional importance of the WUS-CLV feedback loop can be understood from the studies as this loop is active from the embryogenic stage to FM termination. According to the phylogenetic and functional analyses, the WUS-CLV feedback loop is conserved in plant lineages. These genes play pivotal role in embryo patterning, stem cell regulation, SAM maintenance, organ differentiation, and cell identity transition. During this transition, other genes such as LFY become interlinked with the WUS loop and contribute to the cell identity transition for the IM and FM development. FM determinacy control requires a precise regulatory network of these genes to arrest floral stem cell activity for proper sepal, petal, stamen, and carpel development. Based on the environmental cues such as vernalization, photoperiod, phytohormones and ROS signaling interact with the WUS-CLV and WUS-AG for optimal flowering and reproduction growth. However, knowledge of the gene regulatory networks on the stem cell niche for SAM maintenance and its transition mechanism on distinct floral development between plant species is still incomplete.
Our future investigation will be focused on the evolution of the flower, the conserved and distinct adaptation mechanism existing between the plant species during evolution and domestication. The results of our works can be utilized for plant architecture developmental studies and crop improvement programes.
Author Contributions
Conceptualization and framework, P.S.; Writing and original draft preparation, Y.A. and B.S.; Review, editing, and corrections, A.M. and P.S.; Validation, P.S. All authors have read and agreed to the published version of the manuscript.
Funding
Y.A. and B.S. were supported by a short-term research fellowship from the NIPGR core research grant. We acknowledge the NIPGR core research grant to support this study.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This study was supported by the core research grant of the National Institute of Plant Genome Research, New Delhi, India. AM, DST-INSPIRE faculty (DST/INSPIRE/04/2021/003731) acknowledges the support from Department of Science and Technology (DST), Government of India.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaillochet, C.; Lohmann, J.U. The never-ending story: From pluripotency to plant developmental plasticity. Development 2015, 142, 2237–2249. [Google Scholar] [CrossRef] [PubMed]
- Scheres, B. Stem cells: A Plant biology perspective. Cell 2005, 122, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Jürgens, G. Stem cells that make stems. Nature 2002, 415, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Guo, Y.; Zhang, H.; Liu, X.; Guo, L. Same actor in different stages: Genes in shoot apical meristem maintenance and floral meristem determinacy in Arabidopsis. Front. Ecol. Evol. 2020, 8, 89. [Google Scholar] [CrossRef]
- Fletcher, J.C. The CLV-WUS Stem cell signaling pathway: A roadmap to crop yield optimization. Plants 2018, 7, 87. [Google Scholar] [CrossRef]
- Whitewoods, C.D.; Cammarata, J.; Venza, Z.N.; Sang, S.; Crook, A.D.; Aoyama, T.; Wang, X.Y.; Waller, M.; Kamisugi, Y.; Cuming, A.C.; et al. CLAVATA was a genetic novelty for the morphological innovation of 3D growth in land plants. Curr. Biol. 2018, 28, 2365–2376.e5. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.F.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer KF, X.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Ren, J.; Ma, Y.; Yin, J. Characterization of tomato transcription factor WUSCHEL and functional study in Arabidopsis. J. Integr. Agric. 2012, 11, 1257–1265. [Google Scholar] [CrossRef]
- Xu, C.; Liberatore, K.L.; MacAlister, C.A.; Huang, Z.; Chu, Y.H.; Jiang, K.; Brooks, C.; Ogawa-Ohnishi, M.; Xiong, G.; Pauly, M.; et al. A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat. Genetic. 2015, 47, 784–792. [Google Scholar] [CrossRef]
- Nardmann, J.; Werr, W. The shoot stem cell niche in angiosperms: Expression patterns of WUS orthologues in rice and maize imply major modifications in the course of mono-and dicot evolution. Mol. Biol. Evol. 2006, 23, 2492–2504. [Google Scholar] [CrossRef]
- Bommert, P.; Lunde, C.; Nardmann, J.; Vollbrecht, E.; Running, M.; Jackson, D.; Hake, S.; Werr, W. Thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptor-like kinase. Development 2005, 132, 1235–1245. [Google Scholar] [CrossRef]
- Taguchi-Shiobara, F.; Yuan, Z.; Hake, S.; Jackson, D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev. 2001, 15, 2755–2766. [Google Scholar] [CrossRef]
- Je, B.; Gruel, J.; Lee, Y.K.; Bommert, P.; Arevalo, E.D.; Eveland, A.L.; Wu, Q.; Goldshmidt, A.; Meeley, R.; Bartlett, M.; et al. Signaling from maize organ primordia via FASCIATED EAR3 regulates stem cell proliferation and yield traits. Nat. Genet. 2016, 48, 785–791. [Google Scholar] [CrossRef]
- Tanaka, W.; Ohmori, Y.; Ushijima, T.; Matsusaka, H.; Matsushita, T.; Kumamaru, T.; Kawano, S.; Hirano, H.Y. Axillary meristem formation in rice requires the WUSCHEL ortholog TILLERS ABSENT1. Plant Cell 2015, 27, 1173–1184. [Google Scholar] [CrossRef]
- Suzaki, T.; Sato, M.; Ashikari, M.; Miyoshi, M.; Nagato, Y.; Hirano, H.Y. The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis CLAVATA1. Development 2004, 131, 5649–5657. [Google Scholar] [CrossRef]
- Chu, H.; Qian, Q.; Liang, W.; Yin, C.; Tan, H.; Yao, X.; Yuan, Z.; Yang, J.; Huang, H.; Luo, D.; et al. The FLORAL ORGAN NUMBER4 Gene encoding a putative ortholog of Arabidopsis CLAVATA3 regulates apical meristem size in rice. Plant Physiol. 2006, 142, 1039–1052. [Google Scholar] [CrossRef]
- Liu, X.; Kim, Y.J.; Müller, R.; Yumul, R.E.; Liu, C.; Pan, Y.; Cao, X.; Goodrich, J.; Chen, X. AGAMOUS terminates floral stem cell maintenance in Arabidopsis by directly repressing WUSCHEL through recruitment of Polycomb Group proteins. Plant Cell 2011, 23, 3654–3670. [Google Scholar] [CrossRef]
- Itkin, M.; Seybold, H.; Breitel, D.; Rogachev, I.; Meir, S.; Aharoni, A. TOMATO AGAMOUS-LIKE 1 is a component of the fruit ripening regulatory network. Plant J. 2009, 60, 1081–1095. [Google Scholar] [CrossRef]
- Schmidtiai’, R.J.; Veit, B.; Mandel, M.A.; Mena, M.; Hakeyb, S.; Yanofsky’, M.F. Identification and molecular characterization of ZAG1, the maize homolog of the Arabidopsis floral homeotic gene AGAMOUS. Plant Cell 1993, 5, 729–737. [Google Scholar] [CrossRef]
- Suzaki, T.; Toriba, T.; Fujimoto, M.; Tsutsumi, N.; Kitano, H.; Hirano, H.Y. Conservation and diversification of meristem maintenance mechanism in Oryza sativa: Function of the FLORAL ORGAN NUMBER2 gene. Plant Cell Physiol. 2006, 47, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Lee, D.Y.; Miyao, A.; Hirochika, H.; An, G.; Hirano, H.Y. Functional diversification of the two C-class MADS box genes OSMADS3 and OSMADS58 in Oryza sativa. Plant Cell 2006, 18, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, K.; Perales, M.; Snipes, S.; Yadav, R.K.; Diaz-Mendoza, M.; Reddy, G.V. DNA-dependent homodimerization, sub-cellular partitioning, and protein destabilization control WUSCHEL levels and spatial patterning. Proc. Natl. Acad. Sci. USA 2016, 113, E6307–E6315. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Perales, M.; Gruel, J.; Girke, T.; Jönsson, H.; Reddy, G.V. WUSCHEL protein movement mediates stem cell homeostasis in the Arabidopsis shoot apex. Genes Dev. 2011, 25, 2025–2030. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Gallois, J.L.; Nora, F.R.; Mizukami, Y.; Sablowski, R. WUSCHEL induces shoot stem cell activity and developmental plasticity in the root meristem. Genes Dev. 2004, 18, 375–380. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Wang, X.M.; Li, J.; Li, J.H.; Wu, J.S.; Walker, J.C.; Xu, Z.H.; Chong, K. Activation of the WUS gene induces ectopic initiation of floral meristems on mature stem surface in Arabidopsis thaliana. Plant Mol. Bio. 2005, 57, 773–784. [Google Scholar] [CrossRef]
- Zhou, Y.; Yan, A.; Han, H.; Li, T.; Geng, Y.; Liu, X.; Meyerowitz, E.M. HAIRY MERISTEM with WUSCHEL confines CLAVATA3 expression to the outer apical meristem layers. Science 2018, 361, 502–506. [Google Scholar] [CrossRef]
- Kim, H.-J.; Wu, C.-Y.; Yu, H.-M.; Sheen, J.; Lee, H. Dual CLAVATA3 peptides in Arabidopsis shoot stem cell signaling. J. Plant Biol. 2017, 60, 506–512. [Google Scholar] [CrossRef]
- Mitchum, M.G.; Wang, X.; Davis, E.L. Diverse and conserved roles of CLE peptides. Curr. Op. Plant Biol. 2008, 11, 75–81. [Google Scholar] [CrossRef]
- Perales, M.; Rodriguez, K.; Snipes, S.; Yadav, R.K.; Diaz-Mendoza, M.; Reddy, G.V. Threshold-dependent transcriptional discrimination underlies stem cell homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, E6298–E6306. [Google Scholar] [CrossRef]
- Daum, G.; Medzihradszky, A.; Suzaki, T.; Lohmann, J.U. A mechanistic framework for noncell autonomous stem cell induction in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14619–14624. [Google Scholar] [CrossRef]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 2000, 289, 617–619. [Google Scholar] [CrossRef]
- Reddy, G.V.; Meyerowitz, E.M. Stem-cell homeostasis and growth dynamics can be uncoupled in the Arabidopsis shoot apex. Science 2005, 310, 663–667. [Google Scholar] [CrossRef]
- Plong, A.; Rodriguez, K.; Alber, M.; Chen, W.; Reddy, G.V. CLAVATA3 mediated simultaneous control of transcriptional and post-translational processes provides robustness to the WUSCHEL gradient. Nat. Commun. 2021, 12, 6361. [Google Scholar] [CrossRef]
- Chongloi, G.L.; Prakash, S.; Vijayaraghavan, U. Regulation of meristem maintenance and organ identity during rice reproductive development. J. Exp. Bot. 2019, 70, 1719–1736. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Bickle, K.L.; Schrage, K.J.; Muskett, P.; Patel, K.; Clark, S.E. The CLAVATA1-related BAM1, BAM2 and BAM3 receptor kinase-like proteins are required for meristem function in Arabidopsis. Plant J. 2006, 45, 1–16. [Google Scholar] [CrossRef]
- Nimchuk, Z.L.; Zhou, Y.; Tarr, P.T.; Peterson, B.A.; Meyerowitz, E.M. Plant stem cell maintenance by transcriptional cross-regulation of related receptor kinases. Development 2015, 142, 1043–1049. [Google Scholar] [CrossRef]
- Durbak, A.R.; Tax, F.E. CLAVATA signaling pathway receptors of Arabidopsis regulate cell proliferation in fruit organ formation as well as in meristems. Genetics 2011, 189, 177–194. [Google Scholar] [CrossRef]
- Cao, X.; He, Z.; Guo, L.; Liu, X. Epigenetic mechanisms are critical for the regulation of WUSCHEL expression in floral meristems. Plant Physiol. 2015, 168, 1189–1196. [Google Scholar] [CrossRef]
- Sun, B.; Zhou, Y.; Cai, J.; Shang, E.; Yamaguchi, N.; Xiao, J.; Looi, L.S.; Wee, W.Y.; Gao, X.; Wagner, D.; et al. Integration of transcriptional repression and polycomb-mediated silencing of WUSCHEL in floral meristems. Plant Cell 2019, 31, 1488–1505. [Google Scholar] [CrossRef] [PubMed]
- Deyhle, F.; Sarkar, A.K.; Tucker, E.J.; Laux, T. WUSCHEL regulates cell differentiation during anther development. Dev. Biol. 2007, 302, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, M.; Yuan, C.; Han, Y.; Zheng, T.; Cheng, T.; Wang, J.; Zhang, Q. Interactions between WUSCHEL-and CYC2-like transcription factors in regulating the development of reproductive organs in Chrysanthemum morifolium. Int. J. Mol. Sci. 2019, 20, 1276. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Prunet, N.; Morel, P.; Negrutiu, I.; Trehin, C. Time to stop: Flower meristem termination. Plant Physiol. 2009, 150, 1764–1772. [Google Scholar] [CrossRef]
- Endrizzi, K.; Moussian, B.; Haecker, A.; Levin, J.Z.; Laux, T. The SHOOT MERISTEMLESS gene is required for maintenance of undifferentiated cells in Arabidopsis shoot and floral meristems and acts at a different regulatory level than the meristem genes WUSCHEL and ZWILLE. Plant J. 1996, 10, 967–979. [Google Scholar] [CrossRef]
- Long, J.; Barton, M.K. Initiation of axillary and floral meristems in Arabidopsis. Dev. Biol. 2000, 218, 341–353. [Google Scholar] [CrossRef]
- Krizek, B.A.; Blakley, I.C.; Ho, Y.Y.; Freese, N.; Loraine, A.E. The Arabidopsis transcription factor AINTEGUMENTA orchestrates patterning genes and auxin signaling in the establishment of floral growth and form. Plant J. 2020, 103, 752–768. [Google Scholar] [CrossRef]
- Rojo, E.; Sharma, V.K.; Kovaleva, V.; Raikhel, N.V.; Fletcher, J.C. CLV3 is localized to the extracellular space, where it activates the Arabidopsis CLAVATA stem cell signaling pathway. Plant Cell 2002, 14, 969–977. [Google Scholar] [CrossRef]
- Lohmann, J.U.; Hong, R.L.; Hobe, M.; Busch, M.A.; Parcy, F.; Simon, R.; Weigel, D. A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell 2001, 105, 793–803. [Google Scholar] [CrossRef]
- Sun, B.; Ito, T. Regulation of floral stem cell termination in Arabidopsis. Front. Plant Sci. 2015, 6, 17. [Google Scholar] [CrossRef]
- Lenhard, M.; Bohnert, A.; Jürgens, G.; Laux, T. Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell 2001, 105, 805–814. [Google Scholar] [CrossRef]
- Payne, T.; Johnson, S.D.; Koltunow, A.M. KNUCKLES (KNU) encodes a C2H2 zinc-finger protein that regulates development of basal pattern elements of the Arabidopsis gynoecium. Development 2004, 131, 3737–3749. [Google Scholar] [CrossRef]
- Sun, B.; Xu, Y.; Ng, K.H.; Ito, T. A timing mechanism for stem cell maintenance and differentiation in the Arabidopsis floral meristem. Genes Dev. 2009, 23, 1791–1804. [Google Scholar] [CrossRef]
- Guo, L.; Cao, X.; Liu, Y.; Li, J.; Li, Y.; Li, D.; Zhang, K.; Gao, C.; Dong, A.; Liu, X. A chromatin loop represses WUSCHEL expression in Arabidopsis. Plant J. 2018, 94, 1083–1097. [Google Scholar] [CrossRef]
- Gross-Hardt, R.; Lenhard, M.; Laux, T. WUSCHEL signaling functions in interregional communication Arabidopsis ovule development. Genes Dev. 2002, 16, 1129–1138. [Google Scholar] [CrossRef]
- Heisler, M.G.; Ohno, C.; Das, P.; Sieber, P.; Reddy, G.V.; Long, J.A.; Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 2005, 15, 1899–1911. [Google Scholar] [CrossRef]
- Landrein, B.; Vernoux, T. Auxin, Chief Architect of the Shoot Apex. In Auxin and Its Role in Plant Development; Zažímalová, E., Petrášek, J., Benková, E., Eds.; Springer-Verlag: Wien, Austria, 2014; pp. 191–212. [Google Scholar] [CrossRef]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The yin-yang of hormones: Cytokinin and auxin interactions in plant development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef]
- Snipes, S.A.; Rodriguez, K.; DeVries, A.E.; Miyawaki, K.N.; Perales, M.; Xie, M.; Reddy, G.V. Cytokinin stabilizes WUSCHEL by acting on the protein domains required for nuclear enrichment and transcription. PLoS Genet. 2018, 14, e1007351. [Google Scholar] [CrossRef]
- Luo, L.; Zeng, J.; Wu, H.; Tian, Z.; Zhao, Z. A molecular framework for auxin-controlled homeostasis of shoot stem cells in Arabidopsis. Mol. Plant 2018, 11, 899–913. [Google Scholar] [CrossRef]
- Ma, Y.; Miotk, A.; Šutiković, Z.; Ermakova, O.; Wenzl, C.; Medzihradszky, A.; Gaillochet, C.; Forner, J.; Utan, G.; Brackmann, K.; et al. WUSCHEL acts as an auxin response rheostat to maintain apical stem cells in Arabidopsis. Nat. Commun. 2019, 10, 5093. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, D.; Pesce, E.R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Leibfried, A.; To, J.P.C.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL controls meristem function by direct regulation of cytokinin-inducible response regulators. Nature 2005, 438, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miwa, K.; Ishikawa, K.; Yamada, H.; Aiba, H.; Mizuno, T. The arabidopsis sensor His-kinase, AHK4, can respond to cytokinins. Plant Cell Physiol. 2001, 42, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, R.; Zi, H.; Li, Y.; Cao, X.; Li, D.; Lin, D.; Tong, J.; Pan, Y.; Jiao, Y.; et al. AUXIN RESPONSE FACTOR3 regulates floral meristem determinacy by repressing cytokinin biosynthesis and signaling. Plant Cell 2018, 30, 324–346. [Google Scholar] [CrossRef]
- Zhao, Z.; Andersen, S.U.; Ljung, K.; Dolezal, K.; Miotk, A.; Schultheiss, S.J.; Lohmann, J.U. Hormonal control of the shoot stem-cell niche. Nature 2010, 465, 1089–1092. [Google Scholar] [CrossRef]
- Bao, S.; Hua, C.; Shen, L.; Yu, H. New insights into gibberellin signaling in regulating flowering in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 118–131. [Google Scholar] [CrossRef]
- Bolduc, N.; Hake, S. The maize transcription factor KNOTTED1 directly regulates the gibberellin catabolism gene ga20x1. Plant Cell 2009, 21, 1647–1658. [Google Scholar] [CrossRef]
- Yanai, O.; Shani, E.; Dolezal, K.; Tarkowski, P.; Sablowski, R.; Sandberg, G.; Samach, A.; Ori, N. Arabidopsis KNOXI proteins activate cytokinin biosynthesis. Curr. Biol. 2005, 15, 1566–1571. [Google Scholar] [CrossRef]
- Chung, Y.; Zhu, Y.; Wu, M.F.; Simonini, S.; Kuhn, A.; Armenta-Medina, A.; Jin, R.; Østergaard, L.; Gillmor, C.S.; Wagner, D. Auxin Response Factors promote organogenesis by chromatin-mediated repression of the pluripotency gene SHOOTMERISTEMLESS. Nat. Commu. 2019, 10, 886. [Google Scholar] [CrossRef]
- Jung, J.H.; Lee, S.; Yun, J.; Lee, M.; Park, C.M. The miR172 target TOE3 represses AGAMOUS expression during Arabidopsis floral patterning. Plant Sci. 2014, 215, 29–38. [Google Scholar] [CrossRef]
- Kinoshita, A.; Vayssières, A.; Richter, R.; Sang, Q.; Roggen, A.; van Driel, A.D.; Richard, S.S.; Coupland, G. Regulation of shoot meristem shape by photoperiodic signaling and phytohormones during floral induction of Arabidopsis. Elife 2020, 9, e60661. [Google Scholar] [CrossRef]
- Wang, H.; Caruso, L.V.; Downie, A.B.; Perry, S.E. The embryo MADS domain protein AGAMOUS-Like 15 directly regulated expression of a gene encoding an enzymes involved in gibberellin metabolism. Plant Cell 2004, 166, 1206–1219. [Google Scholar] [CrossRef]
- Li, Q.F.; Wang, C.; Jiang, L.; Li, S.; Sun, S.S.M.; He, J.X. An interaction between BZR1 and DELLAs mediates direct signaling crosstalk between brassinosteroids and gibberellins in Arabidopsis. Sci. Signal. 2012, 5, ra72. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate passes puster: A receptor and targets for the defense hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizerd, F.; Godoy, M.; Franco-Zorrillae, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Pan, J.; Lou, D.; Hu, Y.; Yu, D. The bHLH transcription factors MYC2, MYC3, and MYC4 are required for jasmonate-mediated inhibition of flowering in Arabidopsis. Mol. Plant 2017, 10, 1461–1464. [Google Scholar] [CrossRef]
- Wu, K.; Zhang, L.; Zhou, C.; Yu, C.W.; Chaikam, V. HDA6 is required for jasmonate response, senescence and flowering in Arabidopsis. J Exp. Bot. 2008, 59, 225–234. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, L.; Duan, J.; Miki, B.; Wu, K. HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant. Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Ye, T.; Lu, Y.; Chen, X.; Wu, Y. The inhibitory effect of ABA on floral transition is mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Riboni, M.; Robustelli Test, A.; Galbiati, M.; Tonelli, C.; Conti, L. ABA-dependent control of GIGANTEA signaling enables drought escape via up-regulation of FLOWERING LOCUS T in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6309–6632. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Schippers, J.H.M. ROS-mediated redox signaling during cell differentiation in plants. Biochim. Biophys. Acta-Gen. Subj. 2015, 1850, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Phy. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox regulation of plant stem cell fate. EMBO J. 2017, 36, 2844–2855. [Google Scholar] [CrossRef]
- Wang, Y.; Shirakawa, M.; Ito, T. Dynamic changes in reactive oxygen species in the shoot apex contribute to stem cell death in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 2022, 3864. [Google Scholar] [CrossRef]
- Berrios, L.; Rentsch, J.D. Linking reactive oxygen species (ROS) to abiotic and biotic feedbacks in plant microbiomes: The dose makes the poison. Int. J. Mol. Sci. 2022, 23, 4402. [Google Scholar] [CrossRef]
- Leijten, W.; Koes, R.; Roobeek, I.; Frugis, G. Translating flowering time from Arabidopsis thaliana to Brassicaceae and Asteraceae crop species. Plants 2018, 7, 111. [Google Scholar] [CrossRef]
- Sheldon, C.C.; Rouse, D.T.; Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. The molecular basis of vernalization: The central role of FLOWERING LOCUS C (FLC). Proc. Natl. Acad. Sci. USA 2000, 97, 3753–3758. [Google Scholar] [CrossRef]
- Shindo, C.; Aranzana, M.J.; Lister, C.; Baxter, C.; Nicholls, C.; Nordborg, M.; Dean, C. Role of FRIGIDA and FLOWERING LOCUS C in determining variation in flowering time of Arabidopsis. Plant Physiol. 2005, 138, 1163–1173. [Google Scholar] [CrossRef]
- Choi, K.; Kim, J.; Hwang, H.-J.; Kim, S.; Park, C.; Kim, S.Y.; Lee, I. The FRIGIDA complex activates transcription of FLC, a strong flowering repressor in Arabidopsis by recruiting chromatin modification factors. Plant Cell 2011, 23, 289–303. [Google Scholar] [CrossRef]
- Kim, D.H.; Doyle, M.R.; Sung, S.; Amasino, R.M. Vernalization: Winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol. 2009, 25, 277–299. [Google Scholar] [CrossRef]
- Bastow, R.; Mylne, J.; Lister, C.; Lippman, Z.; Martienssen, R.; Dean, C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004, 427, 164–167. [Google Scholar] [CrossRef]
- Amasino, R.M.; Sung, S. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 2004, 427, 159–164. [Google Scholar] [CrossRef]
- Yang, H.; Howard, M.; Dean, C. Antagonistic roles for H3K36me3 and H3K27me3 in the cold-induced epigenetic switch at Arabidopsis FLC. Curr. Biol. 2014, 24, 1793–1797. [Google Scholar] [CrossRef]
- Blazquez, M.A.; Green, R.; Nilsson, O.; Sussman, M.R.; Weigel, D. Gibberelllins promote flowering Arabidopsis by activating the LEAFY promoter. Plant Cell 1998, 10, 791–800. [Google Scholar] [CrossRef]
- Liu, C.; Chen, H.; Er, H.L.; Soo, H.M.; Kumar, P.; Han, J.H.; Liou, Y.C.; Yu, H. Direct interaction of AGL24 and SOC1 integrates flowering signals in Arabidopsis. Development 2008, 135, 1481–1491. [Google Scholar] [CrossRef]
- Sheldon, C.C.; Hills, M.J.; Lister, C.; Dean, C.; Dennis, E.S.; Peacock, W.J. Resetting of FLOWERING LOCUS C expression after epigenetic repression by vernalization. Proc. Natl. Acad. Sci. USA 2008, 105, 2214–2219. [Google Scholar] [CrossRef]
- Choi, J.; Hyun, Y.; Kang, M.J.; Yun, H.I.; Yun, J.Y.; Lister, C.; Dean, C.; Amasino, R.M.; Noh, B.; Noh, Y.S.; et al. Resetting and regulation of FLOWERING LOCUS C expression during Arabidopsis reproductive development. Plant J. 2009, 57, 918–931. [Google Scholar] [CrossRef]
- Angel, A.; Song, J.; Dean, C.; Howard, M. A Polycomb-based switch underlying quantitative epigenetic memory. Nature 2011, 476, 105–108. [Google Scholar] [CrossRef]
- Yun, H.; Hyun, Y.; Kang, M.J.; Noh, Y.S.; Noh, B.; Choi, Y. Identification of regulators required for the reactivation of FLOWERING LOCUS C during Arabidopsis reproduction. Planta 2011, 234, 1237–1250. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).