
A Metallochaperone HIPP33 Is Required for Rice Zinc and Iron Homeostasis and Productivity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Metal Treatments
2.2. Analysis of Gene Structural Sequences
2.3. Transcript Analysis by Quantitative RT-PCR
2.4. Subcellular Localization of OsHIPP33
2.5. Yeast Complementation Assay
2.6. Generation and Analysis of RNAi and Mutant Lines
2.7. Determination of Metals
2.8. Statistical Analysis
3. Results
3.1. Analysis of OsHIPP33 Sequence and Structure
3.2. Expression Pattern of OsHIPP33
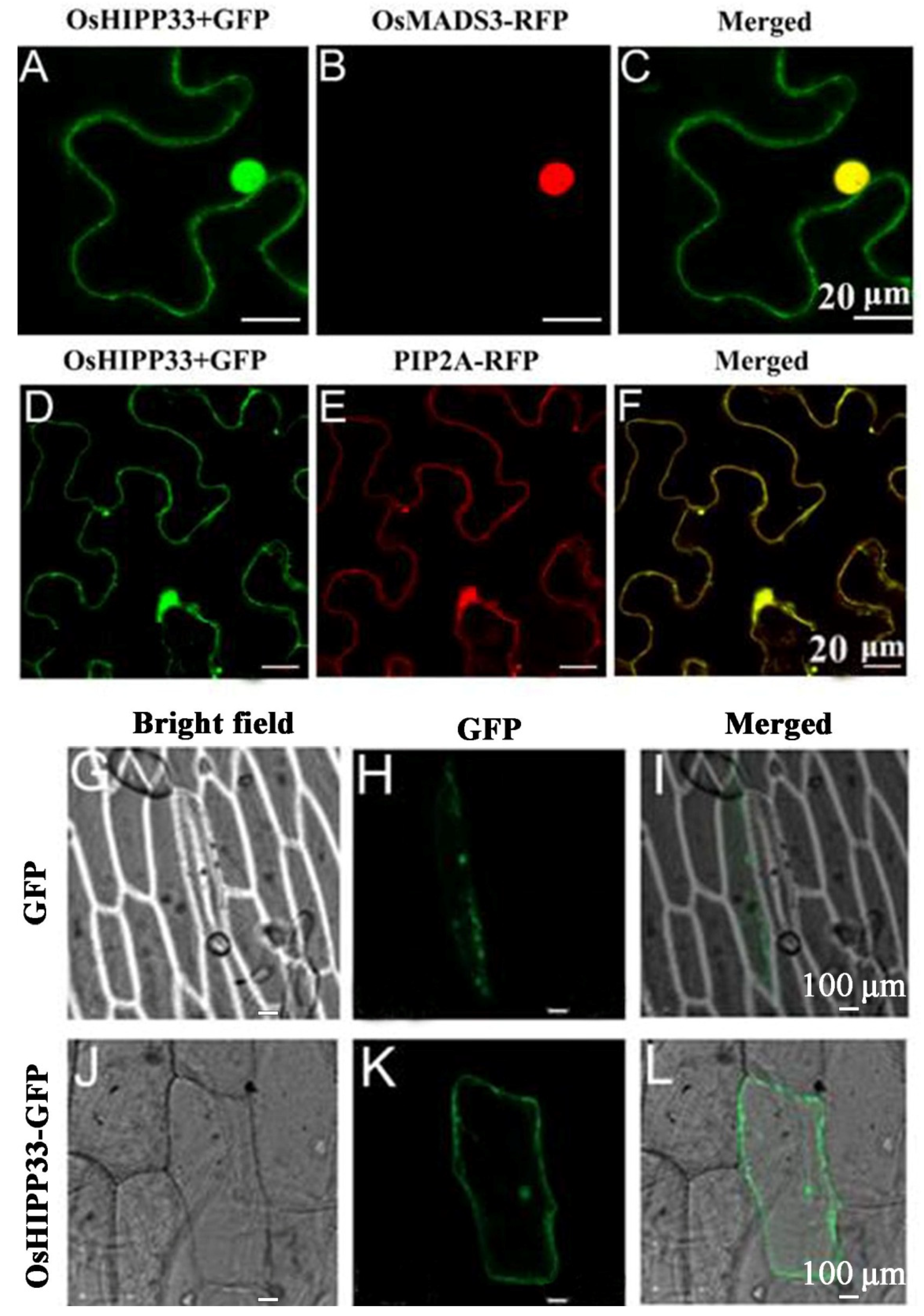
3.3. OsHIPP33 Is Localized to the Nucleus and Plasma Membrane
3.4. Expression of OsHIPP33 Led to Zn Accumulation in Yeast Cells
3.5. OsHIPP33 Did Not Functionally Regulate Rice Growth and Zn and Fe Accumulation during the Early Vegetative Stage
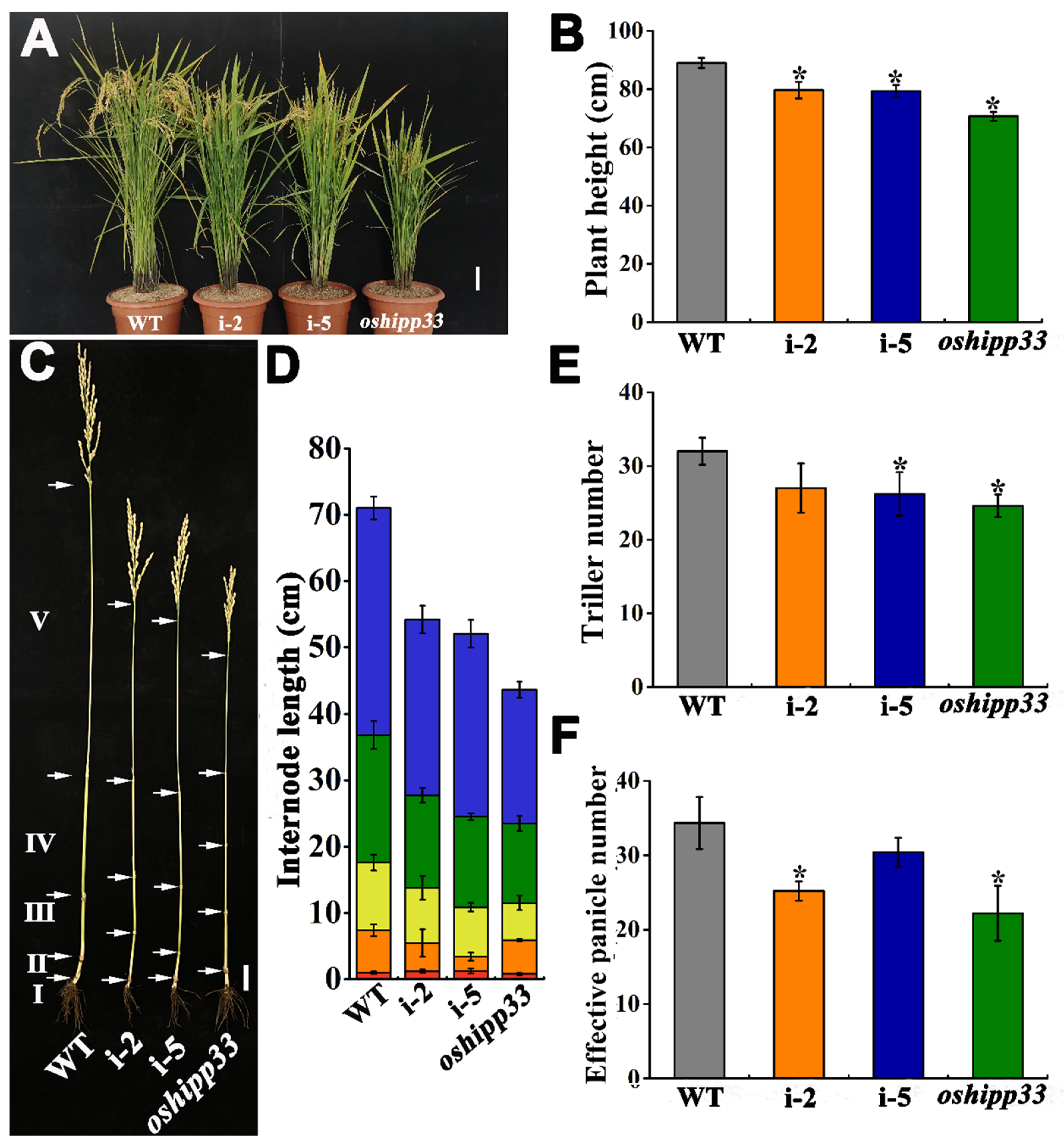
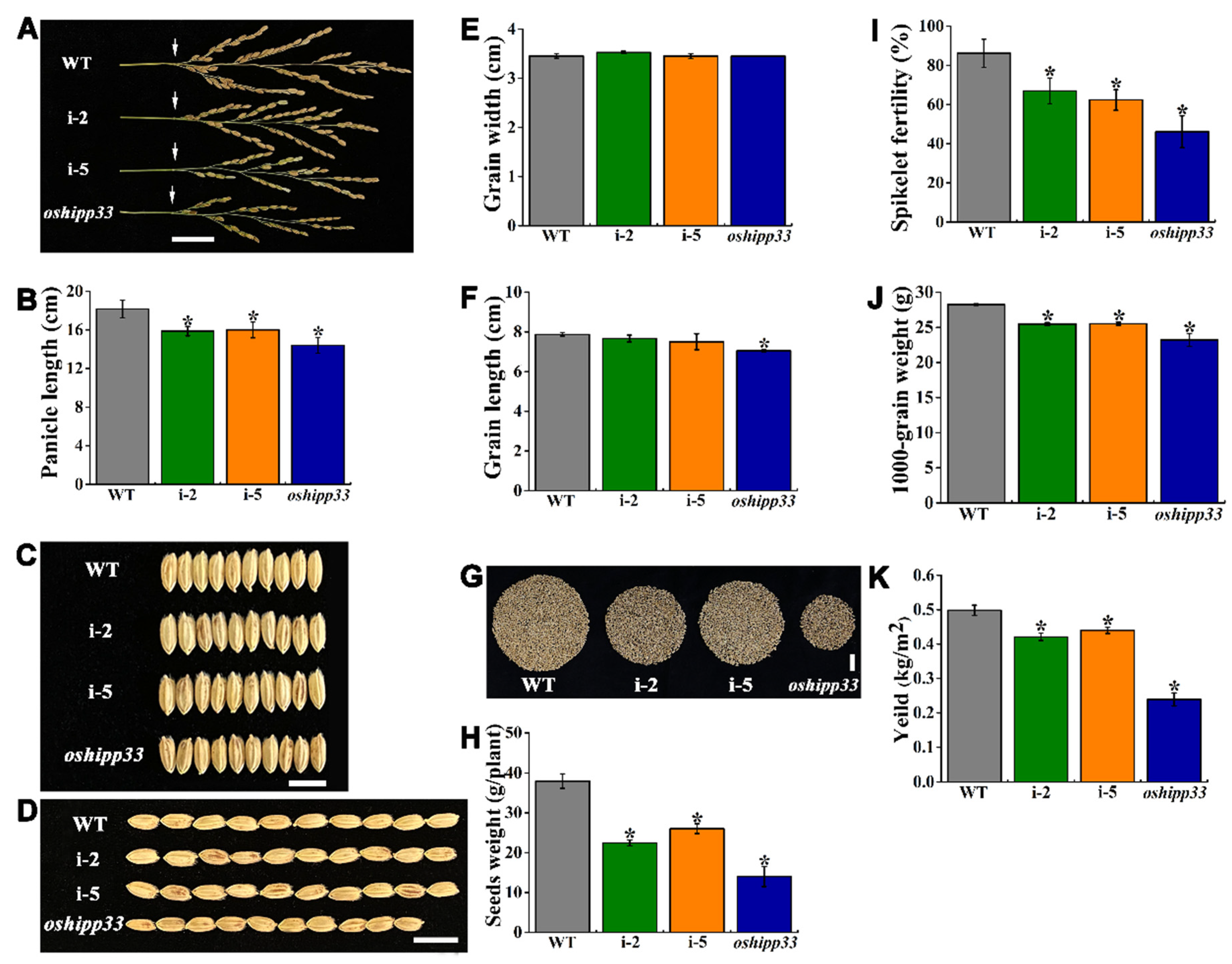
3.6. Mutation of OsHIPP33 Compromised Growth and Reproductivity of Rice
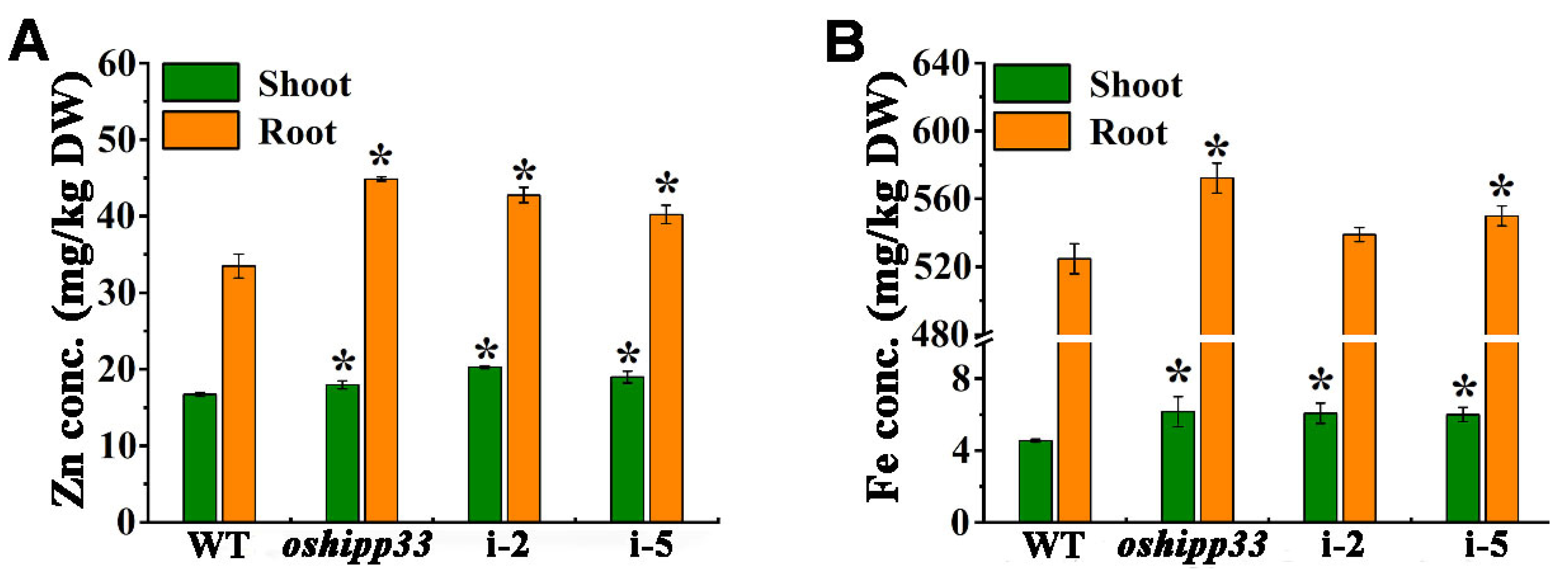
3.7. Mutation of OsHIPP33 Reduced Zn and Fe Concentrations in Rice Tissue
3.8. OsHIPP33 and RNAi Lines Are Sensitive to Excessive Zn and Fe Stress and Accumulated More Metals
4. Discussion
4.1. OsHIPP33-Mediated Zn and Fe Accumulation Is Responsible for Rice Grain Yield
4.2. OsHIPP33 Maintains Homeostasis under Excessive Zn/Fe Stress
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: New York, NY, USA, 2011. [Google Scholar]
- Wessells, K.R.; Brown, K.H. Estimating the global prevalence of zinc deficiency: Results based on zinc availability in national food supplies and the prevalence of stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [Green Version]
- Burchi, F.; Fanzo, J.; Frison, E. The role of food and nutrition system approaches in tackling hidden hunger. Int. J. Environ. Res. Public Health 2011, 8, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Amare, K.; Zeleke, H.; Bultosa, G. Variability for yield, yield related traits and association among traits of sorghum (Sorghum Bicolor (L.) Moench) varieties in Wollo, Ethiopia. J. Plant Breed. Crop Sci. 2015, 7, 125–133. [Google Scholar]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Ishimaru, Y.; Nishizawa, N.K. Molecular mechanisms of zinc uptake and translocation in rice. Plant Soil 2012, 361, 189–201. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Vatansever, R.; Filiz, E.; Eroglu, S. Genome-wide exploration of metal tolerance protein (MTP) genes in common wheat (Triticum aestivum): Insights into metal homeostasis and biofortification. Biometals 2017, 30, 217–235. [Google Scholar] [CrossRef]
- Eide, D.; Broderius, M.; Fett, J.; Guerinot, M.L. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 5624–5628. [Google Scholar] [CrossRef] [Green Version]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef] [Green Version]
- Kavitha, P.G.; Kruvilla, S.; Mathew, M.K. Functional characterization of a transition metal iron transporter OsZIP6 from rice Oryza sativa L. Plant Physiol. Biochem. 2015, 97, 165–174. [Google Scholar]
- Yang, Z.; Yang, F.; Liu, J.L.; Wu, H.T.; Yang, H.; Shi, Y.; Liu, J.; Zhang, Y.F.; Luo, Y.R.; Chen, K.M. Heavy metal transporters: Functional mechanisms, regulation, and application in phytoremediation. Sci. Total Environ. 2021, 809, 151099. [Google Scholar] [CrossRef] [PubMed]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; de Oliveira, L.F.V.; Bodanese Zanettini, M.H.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Tehseen, M.; Cairns, N.; Sherson, S.; Cobbett, C.S. Metallochaperone-like genes in Arabidopsis thaliana. Metallomics 2010, 2, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Rono, J.K.; Zhang, B.Q.; Liu, X.S.; Wang, M.Q.; Wang, L.L.; Wu, X.C.; Chen, X.; Cao, H.W.; Yang, Z.M. Identification of novel rice (Oryza sativa) HPP and HIPP genes tolerant to heavy metal toxicity. Ecotoxicol. Environ. Saf. 2019, 175, 8–18. [Google Scholar] [CrossRef]
- Zhang, X.; Feng, H.; Feng, C.; Xu, H.; Huang, X.; Wang, Q.; Duan, X.; Wang, X.; Wei, G.; Huang, L.; et al. Isolation and characterization of cDNA encoding a wheat heavy metal-associated isoprenylated protein involved in stress responses. Plant Biol. 2010, 17, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.U.; Rono, J.K.; Liu, X.S.; Feng, S.J.; Li, H.; Chen, X.; Yang, Z.M. Functional characterization of a new metallochaperone for reducing cadmium concentration in rice crop. J. Clean. Prod. 2020, 272, 123152. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Liu, X.S.; Feng, S.J.; Zhao, Y.N.; Wang, L.L.; Rono, J.K.; Li, H.; Yang, Z.M. Developing a cadmium resistant rice genotype with OsHIPP29 locus for limiting cadmium accumulation in the paddy crop. Chemosphere 2020, 247, 125958. [Google Scholar] [CrossRef]
- Lin, S.J.; Pufahl, R.A.; Dancis, A.; O’Halloran, T.V.; Culotta, V.C. A role for the Saccharomyces cerevisiae ATX1 gene in copper trafficking and iron transport. Int. J. Biol. Chem. 1997, 272, 9215–9220. [Google Scholar] [CrossRef] [Green Version]
- Robinson, N.J.; Winge, D.R. Copper metallochaperones. Annu. Rev. Biochem. 2010, 79, 537–562. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Zhao, F.J.; Sun, C.; Jin, C.; Shi, Y.; Sun, Y.; Li, Y.; Yang, M.; Jing, X.; et al. OsATX1 interacts with heavy metal P1B-type ATPases and affects copper transport and distribution. Plant Physiol. 2018, 178, 329–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.D.; Wang, Y.; Li, H.B.; Yang, Z.M. Isolation and identification of rapeseed (Brassica napus) cultivars for potential higher and lower Cd accumulation. J. Plant Nutr. Soil Sci. 2018, 181, 479–487. [Google Scholar] [CrossRef]
- Feng, S.J.; Liu, X.S.; Ma, L.Y.; Khan, I.U.; Rono, J.K.; Yang, Z.M. Identification of epigenetic mechanisms in paddy crop associated with lowering environmentally related cadmium risks to food safety. Environ. Pollut. 2020, 256, 113464. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Song, J.B.; Zhao, W.T.; Yang, Z.M. AtHO1 is involved in iron homeostasis in a NO-dependent manner. Plant Cell Physiol. 2013, 54, 1105–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.J.; Liu, X.S.; Cao, H.W.; Yang, Z.M. Identification of a rice metallochaperone for cadmium tolerance by an epigenetic mechanism and potential use for clean up in wetland. Environ. Pollut. 2021, 288, 117837. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.N.; Wang, M.Q.; Li, C.; Cao, H.W.; Rono, J.K.; Yang, Z.M. The metallochaperone OsHIPP56 gene is required for cadmium detoxification in rice crops. Environ. Exp. Bot. 2022, 193, 104680. [Google Scholar] [CrossRef]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Clemens, S.; Deinlein, U.; Ahmadi, H.; Höreth, S.; Uraguchi, S. Nicotianamine is a major player in plant Zn homeostasis. Biometals 2013, 26, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Rono, J.K.; Wang, L.L.; Wu, X.C.; Cao, H.W.; Zhao, Y.N.; Khan, I.U.; Yang, Z.M. Identification of a new metallothionein-like gene OsMT1e for cadmium detoxification and potential phytoremediation. Chemosphere 2021, 265, 129136. [Google Scholar] [CrossRef] [PubMed]
- Bocanski, J.; Sreckov, Z.; Nastasic, A. Genetic and phenotypic relationship between grain yield and components of grain yield of maize (Zea mays L.). Genetika 2009, 41, 145–154. [Google Scholar] [CrossRef]
- Li, L.; Tong, H.; Xiao, Y.; Che, R.; Xu, F.; Hu, B.; Liang, C.; Chu, J.; Li, J.; Chu, C. Activation of Big Grain1 significantly improves grain size by regulating auxi transport in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11102–11107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, R.; Xing, Y.; Xu, Y.; Xiong, D.; Wang, Y.; Yao, S. Sepqrable regulation of POW1 in grain size and leaf angle development in rice. Plant Biotech. J. 2021, 19, 2517–2531. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–545. [Google Scholar] [CrossRef]
- Mu, S.; Yamaji, N.; Sasaki, A.; Luo, L.; Du, B.; Che, J.; Shi, H.; Zhao, H.; Huang, S.; Deng, F.; et al. A transporter for delivering zinc to the developing tiller bud and panicle in rice. Plant J. 2020, 105, 786–799. [Google Scholar] [CrossRef]
- Chen, G.; Xiong, S. OsHIPP24 is a copper metallochaperone which affects rice growth. J. Plant Biol. 2021, 64, 145–153. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Rahman, M.A.; Inabangan-Asilo, M.A.; Amparado, A.; Manito, C.; Chadha-Mohanty, P.; Reinke, R.; Slamet-Loedin, I.H. Advances in breeding for high grain zinc in rice. Rice 2016, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Inoue, A.; Takasaki, H.; Kaku, H.; Akao, S.; Komatsu, S. A proteomic approach to analyze auxin- and zinc-responsive protein in rice. J. Proteome Res. 2005, 4, 456–463. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, T.; Wang, Z.; Yang, J.; Zhang, J. Involvement of cytokinins in the grain filling of rice under alternate wetting and drying irrigation. J. Exp. Bot. 2010, 61, 3719–3733. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Robe, K.; Gaymard, F.; Izquierdo, E.; Dubos, C. The transcriptional control of iron homeostasis in plants: A tale of bHLH transcription factors? Front. Plant Sci. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Xiao, Y.; Xu, F.; Gao, X.; Cao, S.; Zhang, F.; Wang, G.; Sanders, D.; Chu, C. Cytokinin-dependent regulatory module underlies the maintenance of zinc mutrition in rice. New Phytol. 2019, 224, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Gruissem, W.; Bhullar, N.K. Targeting intracellular transport combined with efficient uptake and storage significantly increases grain iron and zinc levels in rice. Plant Biotechnol. J. 2019, 17, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, H.X.; Wang, Z.F.; Wang, Q.H.; Wang, M.M.; Bao, Y.M.; Huang, J.; Zhang, H.S. Characterization of a vacuolar zinc transporter OZT1 in rice (Oryza sativa L.). Mol. Biol. Rep. 2013, 40, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Menguer, P.K.; Farthing, E.; Peaston, K.A.; Ricachenevsky, F.K.; Fett, J.P.; Williams, L.E. Functional analysis of the rice vacuolar zinc transporter OsMTP1. J. Exp. Bot. 2013, 64, 2871–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briat, J.F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and iron storage in plants. Biochim. Biophys. Acta 2010, 1800, 806–814. [Google Scholar] [CrossRef]
- Silveira, V.C.D.; Fadanelli, C.; Sperotto, R.A.; Stein, R.J.; Basso, L.A.; Santos, D.S.; Vaz Junior, I.D.S.; Dias, J.F.; Fett, J.P. Role of ferritin in the rice tolerance to iron overload. Sci. Agric. 2009, 66, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ye, L.; Kong, Q.; Shou, H. A vacuolar membrane ferric-chelate reductase, OsFRO1, alleviates Fe toxicity in rice (Oryza sativa L.). Front. Plant Sci. 2019, 10, 700. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, H.W.; Li, C.; Zhang, B.Q.; Rono, J.K.; Yang, Z.M. A Metallochaperone HIPP33 Is Required for Rice Zinc and Iron Homeostasis and Productivity. Agronomy 2022, 12, 488. https://doi.org/10.3390/agronomy12020488
Cao HW, Li C, Zhang BQ, Rono JK, Yang ZM. A Metallochaperone HIPP33 Is Required for Rice Zinc and Iron Homeostasis and Productivity. Agronomy. 2022; 12(2):488. https://doi.org/10.3390/agronomy12020488
Chicago/Turabian StyleCao, Hong Wei, Cao Li, Bai Qing Zhang, Justice Kipkorir Rono, and Zhi Min Yang. 2022. "A Metallochaperone HIPP33 Is Required for Rice Zinc and Iron Homeostasis and Productivity" Agronomy 12, no. 2: 488. https://doi.org/10.3390/agronomy12020488