
Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
- Dominant Poaceae species ie: Calamagrostis epigejos
- Subordinate Poaceae species important for grassland management
- Fabaceae species important for grassland management
- Dicotyledonous species neutral for grassland management
- Stinger plants
2.3. Data Analysis
3. Results
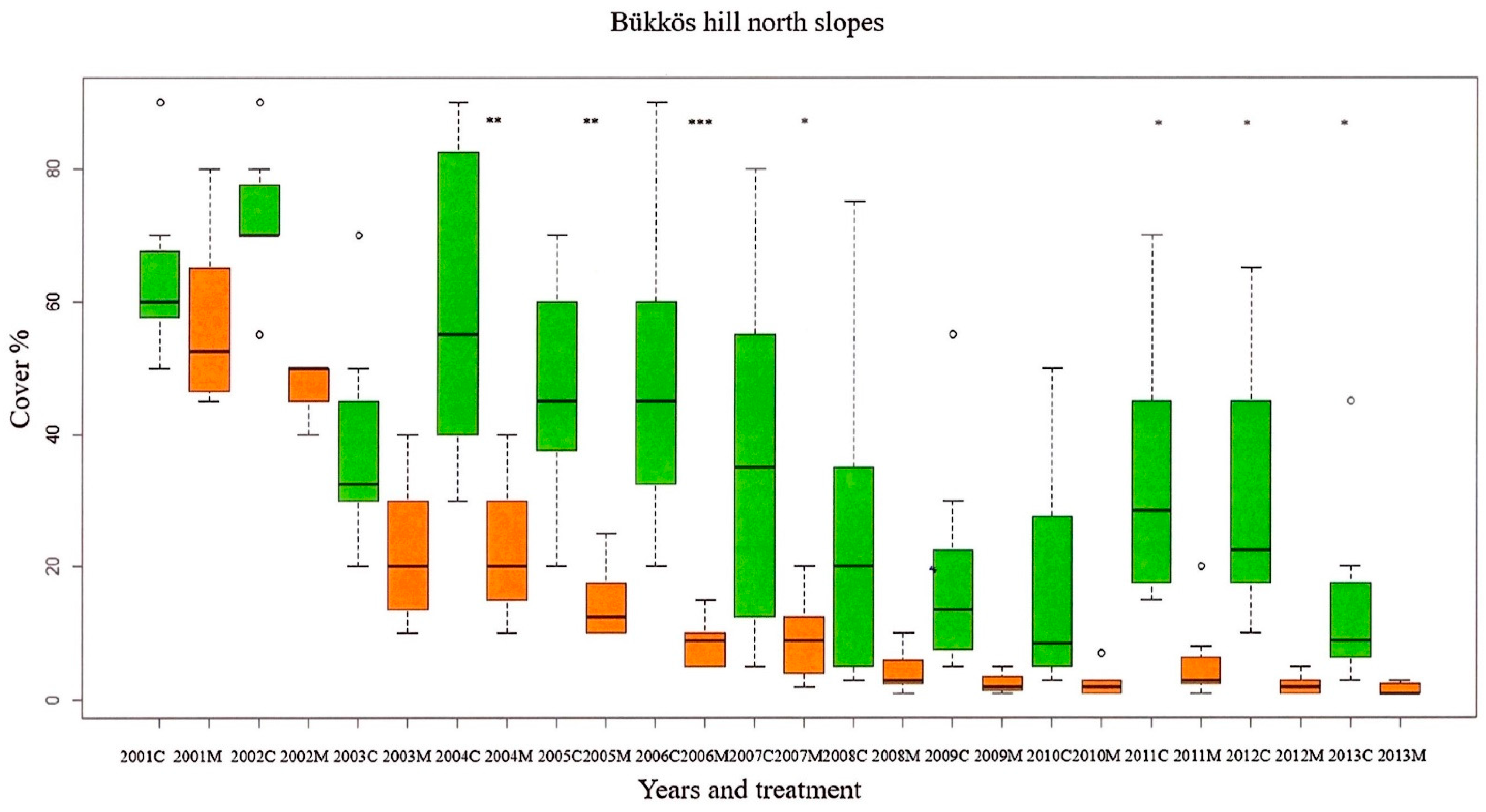
3.1. Cover of Calamagrostis epigejos
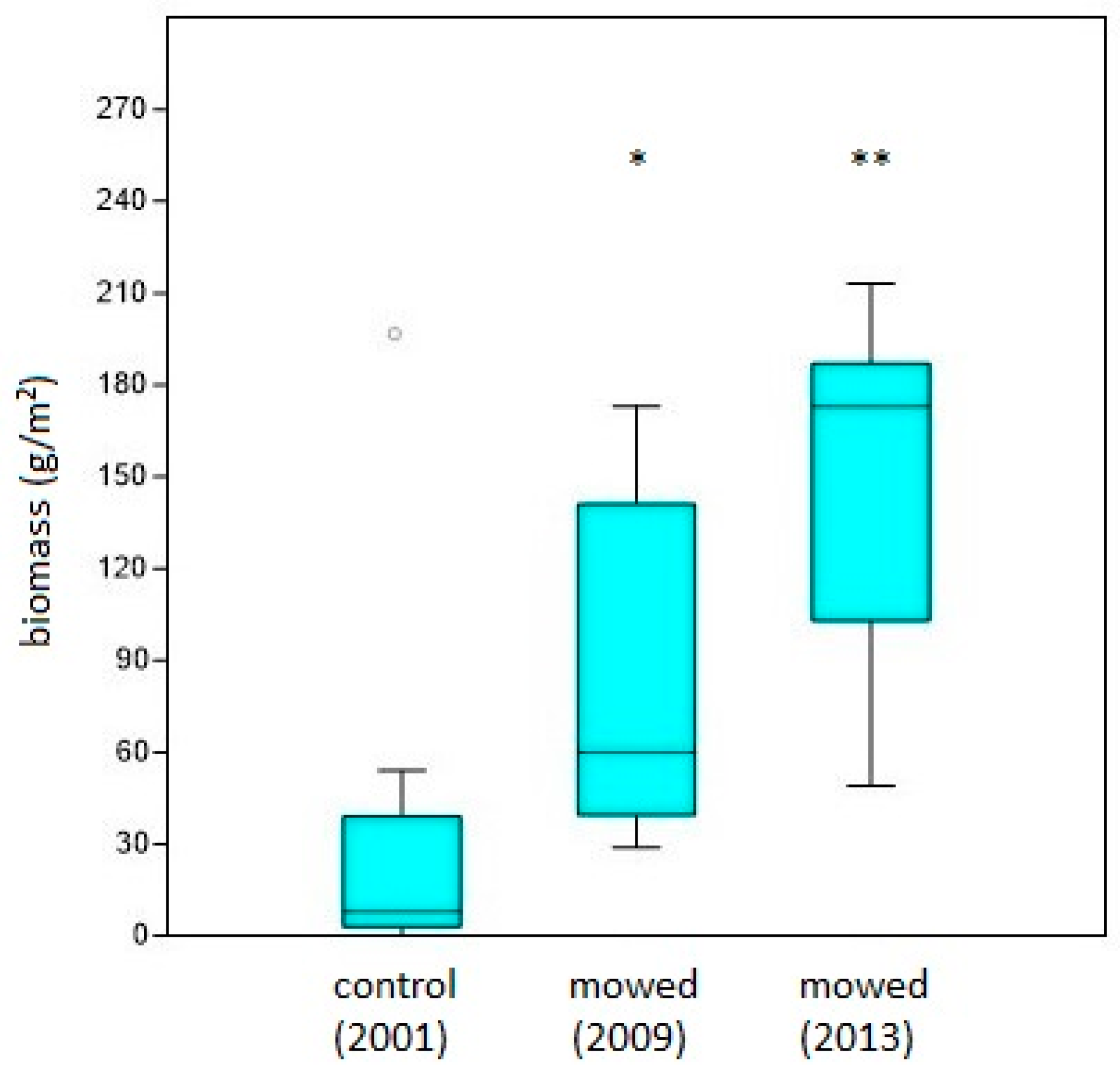
3.2. Cover of Other/Subordinate Species
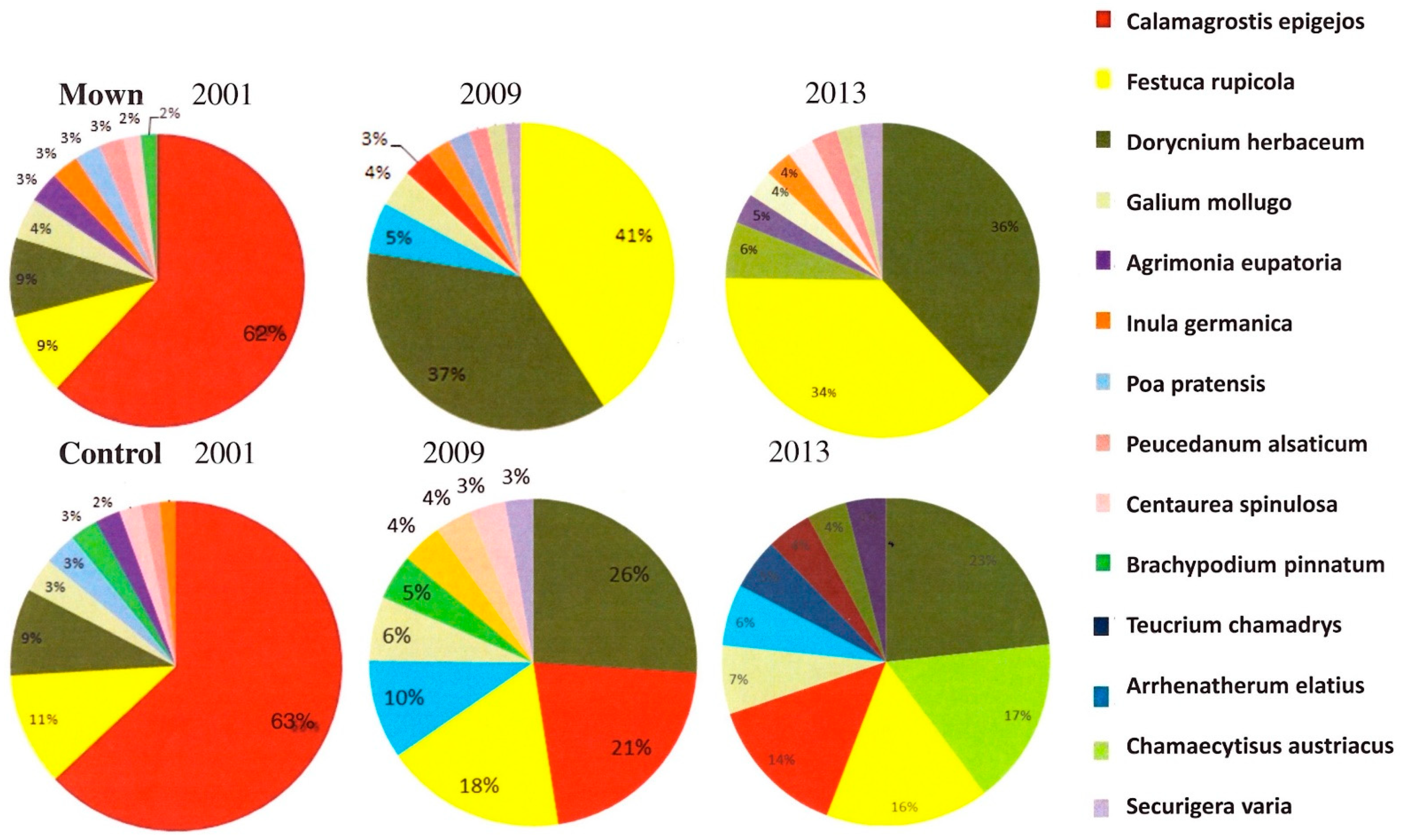
3.3. Species Rank Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef]
- Schuhmacher, O.; Dengler, J. Das Land-Reitgras als Problemart auf Trockenrasen Handlungsempfehlungen zur Reduktion von Calamagrostis epigejos; Ergebnisse aus einem Praxisversuch; NABU: Hamburg, Germany, 2013; pp. 10–15. [Google Scholar]
- Březina, S.; Koubek, T.; Münzbergová, Z.; Herbena, T. Ecological benefits of integration of Calamagrostis epigejos ramets under field conditions. Stanislav. Flora 2006, 201, 461–467. [Google Scholar] [CrossRef]
- Oborny, B.; Kun, A.; Czárán, T.; Bokros, S. The effect of clonal integration on plant competition for mosaic habitat space. Ecology 2000, 81, 3291–3304. [Google Scholar] [CrossRef]
- Zhe-fu, Y.; Shuang, S.; Xiao-lu, X.; Qing, M.; Yin, L. Sources, migration, accumulation and influence of microplastics in terrestrial plant communities. Environ. Exp. Bot. 2021, 192, 104635. [Google Scholar]
- Saul, W.C.; Roy, H.E.; Booy, O.; Carnevali, L.; Chen, H.J.; Genovesi, P.; Harrower, C.A.; Hulme, P.E.; Pagad, S.; Pergl, J.; et al. Assessing patterns in introduction pathways of alien species by linking major invasion databases. J. Appl. Ecol. 2017, 54, 657–669. [Google Scholar] [CrossRef]
- Tasi, J. Diverse impacts of nature conservation grassland management. Cereal Res. Commun. 2007, 35, 1205–1209. [Google Scholar] [CrossRef]
- Poschlod, P.; Wallis de Vries, M.F. The historical and socioeconomic perspective of calcareous grasslands–lessons from the distant and recent past. Biol. Conserv. 2002, 104, 361–376. [Google Scholar] [CrossRef]
- Lindborg, R. Recreating grasslands in Swedish rural land scapes–Effects of seed sowing and management history. Biodiv. Conserv. 2006, 15, 957–969. [Google Scholar] [CrossRef]
- Dostatny, D.F.; Żurek, G.; Kapler, A.; Podyma, W. The Ex Situ Conservation and Potential Usage of Crop Wild Relatives in Poland on the Example of Grasses. Agronomy 2021, 11, 94. [Google Scholar] [CrossRef]
- Ryser, P.; Langenauer, R.; Gigon, A. Species richness and vegetation structure in a limestone grassland after 15 years management with six biomass removal regimes. Folia Geobot. Phytotax 1995, 30, 157–167. [Google Scholar] [CrossRef]
- Fiala, K.; Holub, P.; Sedláková, I.; Tůma, I.; Záhora, J.; Tesařová, M. Reasons and consequences of expansion of Calamagrostis epigejos in alluvial meadows of landscape affected by water control measures. Ekológia 2003, 22 (Suppl. S2), 242–252. [Google Scholar]
- Bartha, S. Composition, differentiation and dynamics in the forest steppe biome. In Slope Steppes, Loess Steppes and Forest Steppe Meadows in Hungary; Illyés, E., Bölöni, J., Eds.; MTA ÖBKI: Budapest, Hungary; ISBN 978-963-06-3673-5 2007. pp. 194–211.
- Rebele, F. Calamagrostis epigejos (L.) Roth auf anthro pogenen Standorten–ein Überblick. Verh. Ges. Okol. 1996, 26, 753–763. [Google Scholar]
- Sedláková, I.; Fiala, K. Ecological degradation of al luvial meadows due to expanding Calamagrostis epigejos. Ekológia 2001, 20 (Suppl. S3), 226–333. [Google Scholar]
- Catorci, A.; Cesaretti, S.; Gatti, R. Effect of long-term abandonment and spring grazing on floristic and functional composition of dry grasslands in a central Apennine farmland. Pol. J. Ecol. 2013, 61, 505–518. [Google Scholar]
- Prach, K.; Pyšek, P. Clonal plants–What is their role in succession? Folia Geobot. Phytotax 1994, 29, 307–320. [Google Scholar] [CrossRef]
- Prach, K.; Pyšek, P. Using spontaneous succession for restoration of human-disturbed habitats: Experience from Central Europe. Ecol. Eng. 2001, 17, 55–62. [Google Scholar] [CrossRef]
- Somodi, I.; Virágh, K.; Podani, J. The effect of the expansion of the clonal grass Calamagrostis epigejos on the species turnover of a semi-arid grassland. Appl. Veg. Sci. 2008, 11, 187–194. [Google Scholar] [CrossRef]
- Zhukovskaya, O.; Ulanova, N.G. Influence of brushing frequency on birch population structure after felling. Ecoscience 2006, 13, 219–225. [Google Scholar] [CrossRef]
- Csontos, P. Light ecology and regeneration on clearings of sessile oak-turkey oak forests in the Visegrád mountains, Hungary. Acta Bot. Hung. 2010, 52, 265–286. [Google Scholar] [CrossRef]
- Gergely, A.; Hahn, I.; Mészáros-Draskovits, R.; Simon, T.; Szabó, M.; Barabás, S. Vegetation succession in a newly exposed Danube riverbed. Appl. Veg. Sci. 2001, 4, 35–40. [Google Scholar] [CrossRef]
- Prach, K. Succession of vegetation on dumps from strip coal mining, N.W.Bohemia, Czechoslovakia. Folia Geobot. Phyto tax 1987, 22, 339–354. [Google Scholar] [CrossRef]
- Bartha, S. Preliminary scaling for multi-species coalitions in primary succession. Abstr. Bot. 1992, 16, 31–41. [Google Scholar]
- Rebele, F. Competition and coexistence of rhizomatous perennial plants along a nutrient gradient. Plant Ecol. 2000, 147, 77–94. [Google Scholar] [CrossRef]
- Huhta, A.; Pasi, R.; Tuomi, J.; Laine, K. Restorative mowing on an abandoned semi-natural meadow: Short-term and predicted long-term effects. J. Veg. Sci. 2001, 12, 677–686. [Google Scholar] [CrossRef]
- Deák, B.; Tóthmérész, B. A kaszálás hatása a hortobányi Nyírőlapos csetkákás társulásában (Effect of cutting on a Bolboschoenetum maritimi eleochariosum association in the Nyirőlapos Hortobágy). Természetvédelmi Közlemények 2007, 13, 179–186. [Google Scholar]
- Kramberger, B.; Kaligaric, M. Semi-natural grasslands: The effect of cutting frequency on long-term changes of floristic composition. Polish J. Ecol. 2008, 56, 33–43. [Google Scholar]
- Rebele, F.; Lehmann, C. Biological Flora of Central Europe: Calamagrostis epigejos (L.). Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Valkó, O.; Deák, B. Increasing the potential of prescribed burning for the biodiversity conservation of European grasslands. Curr. Opin. Environ. Sci. Health 2021, 22, 100268. [Google Scholar] [CrossRef]
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural grassland management by long-term mowing of Calamagrostis epigejos in western Cserhát, Hungary (Management sekundärer Trockenrasen durch Langzeit-Mahd von Calamagrostis epigejos im westlichen Cserhát, Ungarn). In Steppenlebensräume Europas–Gefährdung, Erhaltungsmassnahmen und Schutz; Baumbach, H., Pfützenreuter, S., Eds.; Thüringer Ministerium für Landwirtschaft, Forsten, Umwelt und Naturschutz (TMLFUN): Erfurt, Germany, 2013; pp. 331–340. ISBN 978-3-00-044248-3. [Google Scholar]
- Tesitel, J.; Mládek, J.; Fajmon, K.; Blazek, P.; Mudrák, O. Reversing expansion of Calamagrostis epigejos in a grassland biodiversity hotspot: Hemiparasitic Rhinanthus major does a better job than increased mowing intensity. Appl. Veg. Sci. 2018, 21, 104–112. [Google Scholar] [CrossRef]
- Tesitel, J.; Mládek, J.; Horník, J.; Tesitelová, T.; Adamec, V.; Tichy, L. Suppressing competitive dominants and community restoration with native parasitic plants using the hemiparasitic Rhinanthus alectorolophus and the dominant grass Calamagrostis epigejos. J. Appl. Ecol. 2017, 54, 1487–1495. [Google Scholar] [CrossRef] [Green Version]
- Pruchniewicz, D.; Żołnierz, I. The effect of different restoration treatments on the vegetation of the mesic meadow degraded by the expansion of Calamagrostis epigejos. Int. J. Agric. Biol. 2019, 22, 347–354. [Google Scholar]
- Marosi, S.; Somogyi, S. (Eds.) Magyarország Kistájainak Katasztere (Cadastral of Microregions of Hungary); MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 1991; pp. 379–388. [Google Scholar]
- Klapp, E.; Boeker, P.; König, F.; Stählin, A. Wertzahlen der Grünlandpflanzen. Grünland 1953, 2, 38–40. [Google Scholar]
- Briemle, G.; Nitsche, S.; Nitsche, L. Nutzungswertzahlen für Gefäßpflanzen des Grünlandes. Schriftenreihe für Vegetationskunde; Bundesamt für Naturschutz: Bonn, Germany, 2005; pp. 203–225. [Google Scholar]
- Balázs, F. A Gyepek Botanikai és Gazdasági Értékelése; Mezőgazdasági kiadó: Budapest, Hungary, 1960. [Google Scholar]
- Tasi, J.; Bajnok, M.; Sutyinszki, Z.; Szentes, S. Assesing the quality and quantity of green forage with the help of a three-dimensional method. In Proceedings of the 19th International Scientific Symposium on Nutrition of Farm Animals “Zadravec-erjavec Days”, Radenci, Slovenia, 28–29 October 2010; pp. 152–160. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 20 February 2018).
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural grassland management by mowing of Calamagrostis epigejos in Hungary. Plant Biosyst. 2011, 145, 699–707. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Laurenroth, W.K. A generalized model of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Klimes, L.; Klimesova, J. The effects of mowing and fertilisation on carbohydrate reserves and regrowth of grasses: Do they promote plant coexistence in species-rich meadows? Evol. Ecol. 2002, 15, 363–382. [Google Scholar] [CrossRef]
- Kavanová, M.; Gloser, V. The use of internal nitrogen stores in the rhizomatous grass Calamagrostis epigejos during regrowth after defoliation. Ann. Bot. 2005, 85, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; During, H.; Schreurs, J.; Willems, J.; Zielman, R. Effects of selective clipping and mowing time on species diversity in chalk grassland. Folia Geobot. Phytotax 1987, 22, 363–376. [Google Scholar] [CrossRef]
- Tasi, J.; Szemán, L. Landbewirtschaftung in Ungarn. Multifunktionale Landnutzung und Perspektiven für Extensive Weidesysteme; Fachverlag Köhler: Giessen, Germany, 2007; pp. 45–57. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MOWN | CONTROL | |||||
|---|---|---|---|---|---|---|
| 2001 | 2009 | 2013 | 2001 | 2009 | 2013 | |
| Total cover | 103.38 | 102.66 | 121.15 | 115.70 | 103.18 | 139.26 |
| Cover of Calamagrostis epigejos absolute | 56.63 | 2.50 | 1.62 | 63.75 | 18.37 | 14.25 |
| Cover of Calamagrostis epigejos relative | 0.55 | 0.02 | 0.01 | 0.55 | 0.18 | 0.10 |
| Cover of subordinated species absolute | 46.75 | 100.16 | 119.52 | 51.95 | 84.81 | 125.01 |
| Cover of subordinated species relative | 0.45 | 0.95 | 0.98 | 0.45 | 0.81 | 0.89 |
| Number of species | 14.88 | 29.37 | 32.62 | 18.25 | 24.25 | 27.62 |
| Shannon Diversity | 1.62 | 2.07 | 2.34 | 1.66 | 2.07 | 2.53 |
| Plots | 01C | 01M | 02C | 03M | 03C | 03M | 04C | 04M | 05C | 05M | 06C | 06M | 07C | 07M |
| Klapp average values | 1.7 | 1.8 | 1.8 | 1.9 | 1.8 | 2.4 | 1.8 | 2.9 | 2.4 | 1.9 | 2.3 | 2.9 | 2.1 | 3.2 |
| Grass yield (t/ha) | 7.1 | 10.4 | 7.1 | 10.4 | 7.1 | 10.4 | 7.1 | 12.5 | 7.4 | 14.1 | 7.1 | 14.8 | 7.1 | 14.4 |
| Plots | 08C | 08M | 09C | 09M | 10C | 10M | 11C | 11M | 12C | 12M | 13C | 13M | ||
| Klapp average values | 2.1 | 2.8 | 3.4 | 3.8 | 3.3 | 3.9 | 2.5 | 3.2 | 2.1 | 3.8 | 2.1 | 3.9 | ||
| Grass yield (t/ha) | 7.1 | 13.6 | 8.8 | 12.4 | 8.1 | 14.8 | 7.6 | 14.84 | 7.5 | 13.6 | 7.4 | 14.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Házi, J.; Penksza, K.; Barczi, A.; Szentes, S.; Pápay, G. Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands. Agronomy 2022, 12, 1107. https://doi.org/10.3390/agronomy12051107
Házi J, Penksza K, Barczi A, Szentes S, Pápay G. Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands. Agronomy. 2022; 12(5):1107. https://doi.org/10.3390/agronomy12051107
Chicago/Turabian StyleHázi, Judit, Károly Penksza, András Barczi, Szilárd Szentes, and Gergely Pápay. 2022. "Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands" Agronomy 12, no. 5: 1107. https://doi.org/10.3390/agronomy12051107
APA StyleHázi, J., Penksza, K., Barczi, A., Szentes, S., & Pápay, G. (2022). Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands. Agronomy, 12(5), 1107. https://doi.org/10.3390/agronomy12051107