
Low Plant Density Improves Fruit Quality without Affecting Yield of Cucumber in Different Cultivation Periods in Greenhouse

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Measurements of Cucumber Yield, Dry Weight of Leaves, Petioles, Stem and Fruits
2.2. Measurements of Outside Temperature, Radiation Sum and Greenhouse Temperature
2.3. Measurements of Leaf Gas Exchange Parameters
2.4. Measurements of Fruit Chlorophyll and Carotenoid Contents
2.5. Measurement of Soluble Sugar, Soluble Protein, Vitamin C, Total Phenolic and Flavonoid Contents
2.6. Measurements of Nitrate and Nitrite Contents
2.7. Statistical Analysis
3. Results
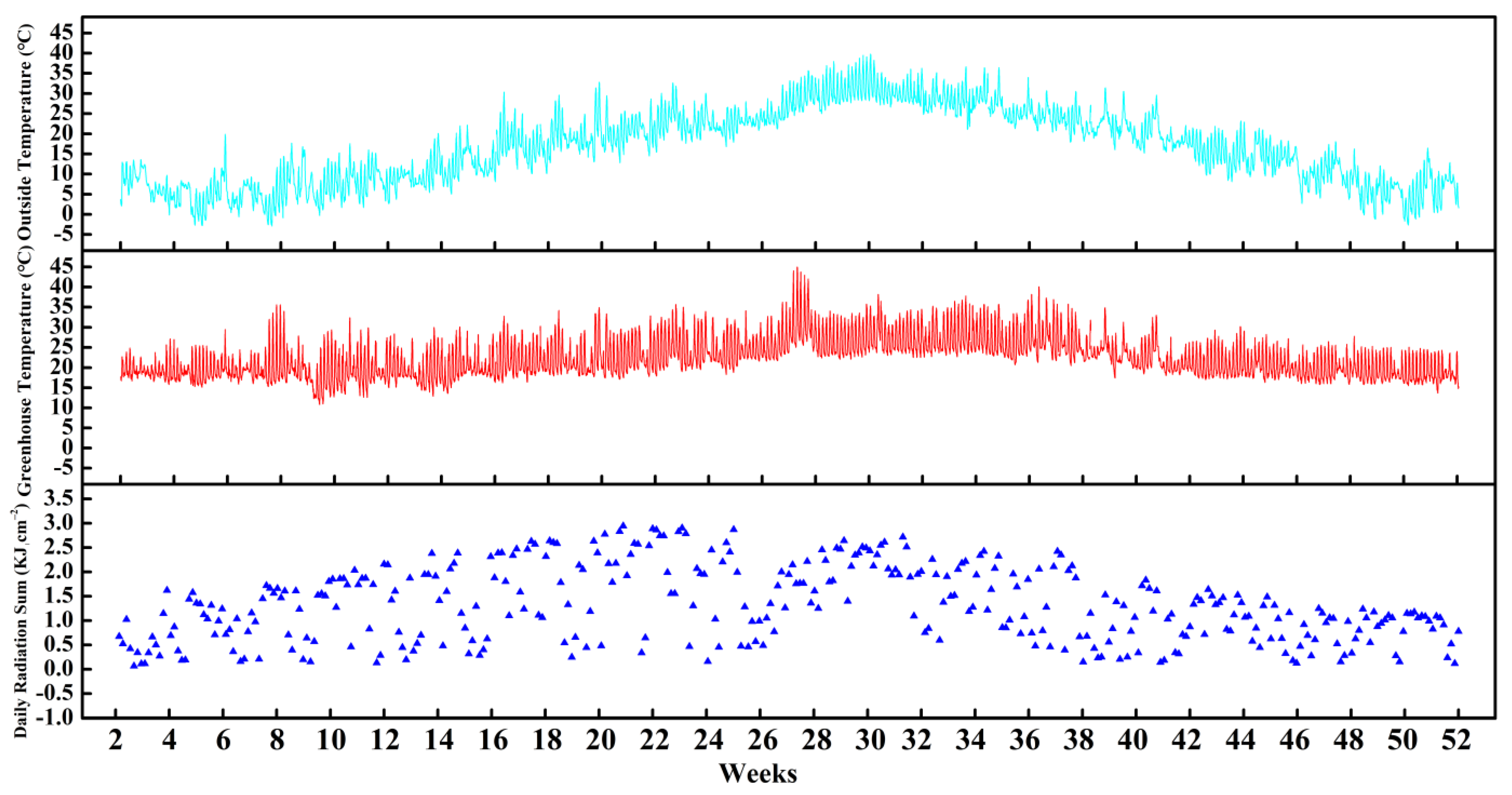
3.1. Environmental Changes in Greenhouse
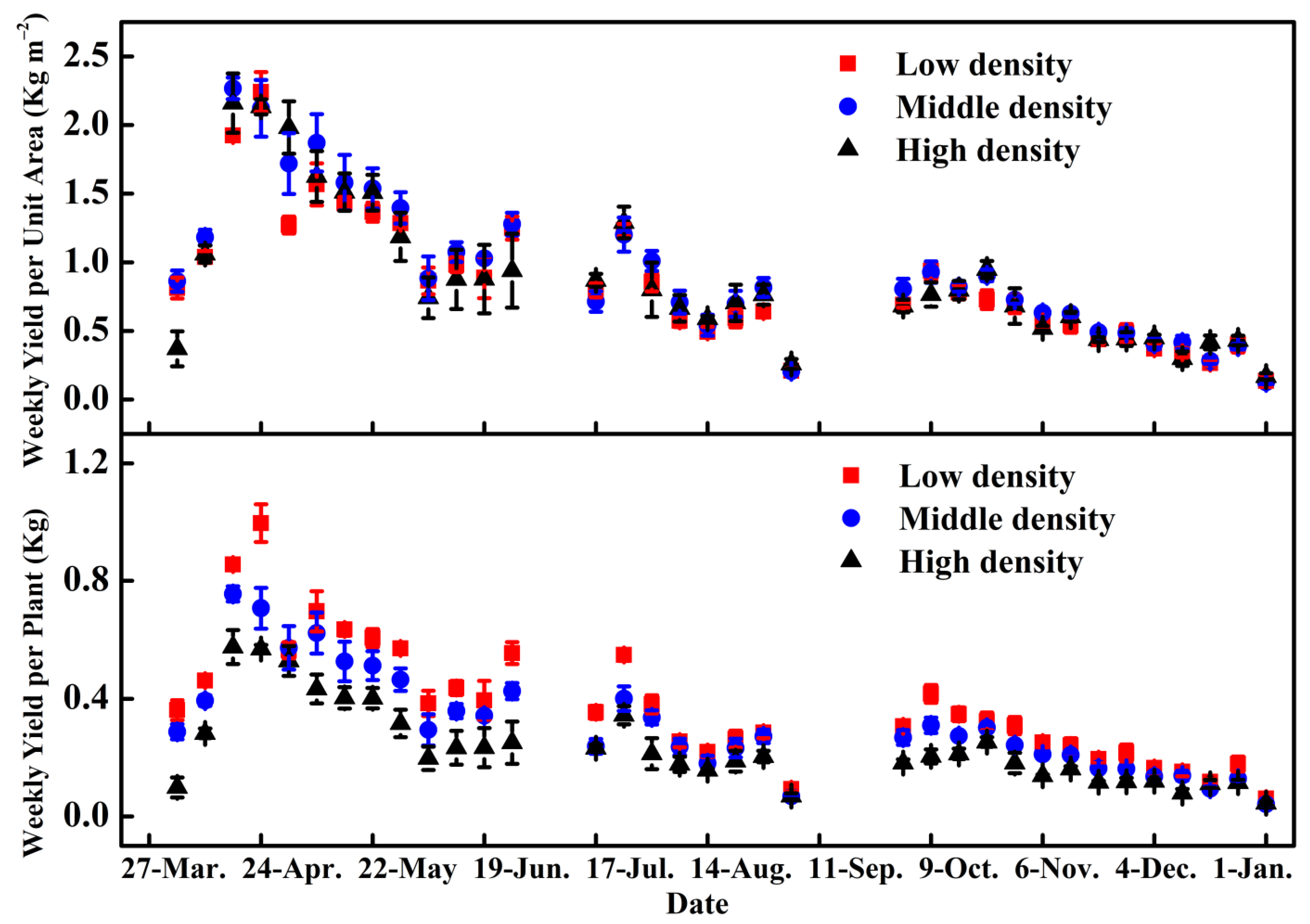
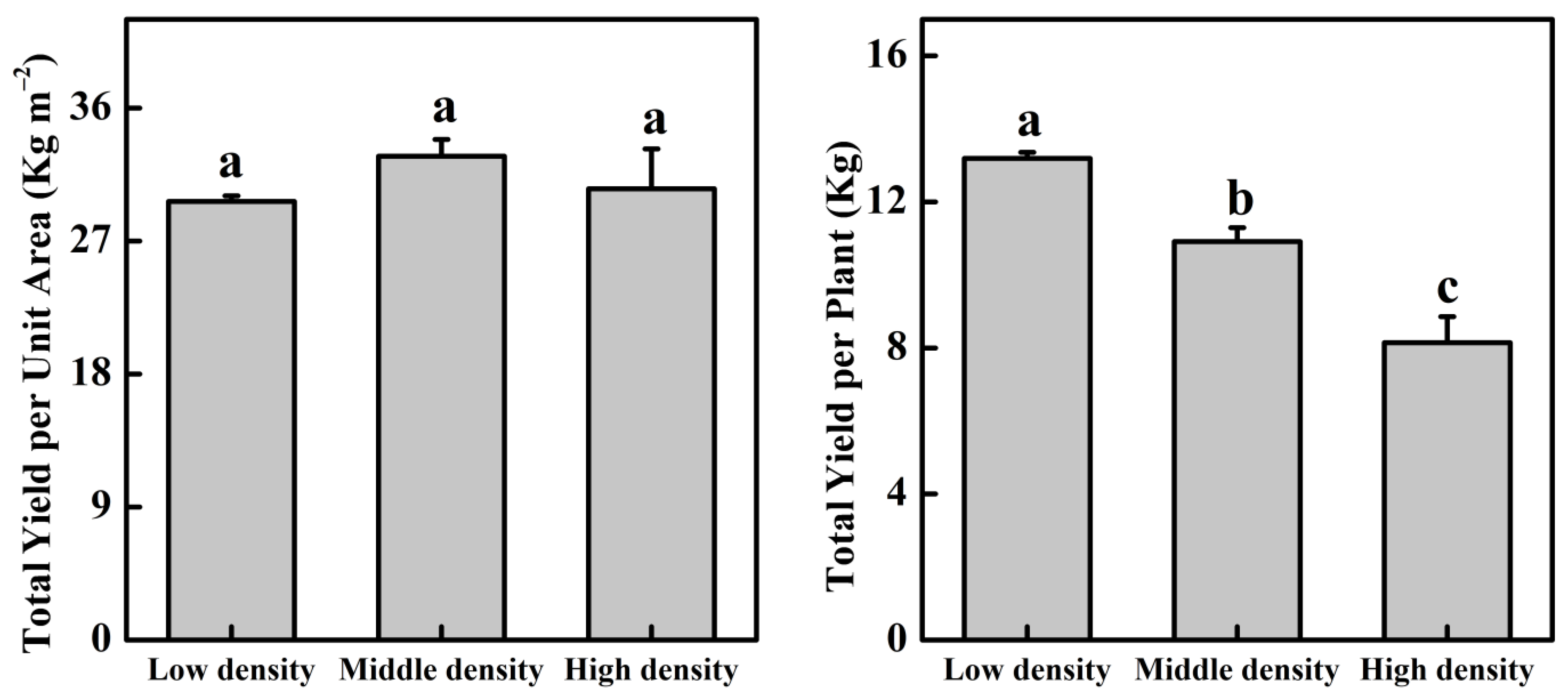
3.2. Cucumber Yield Changes in Three Plant Density Treatments
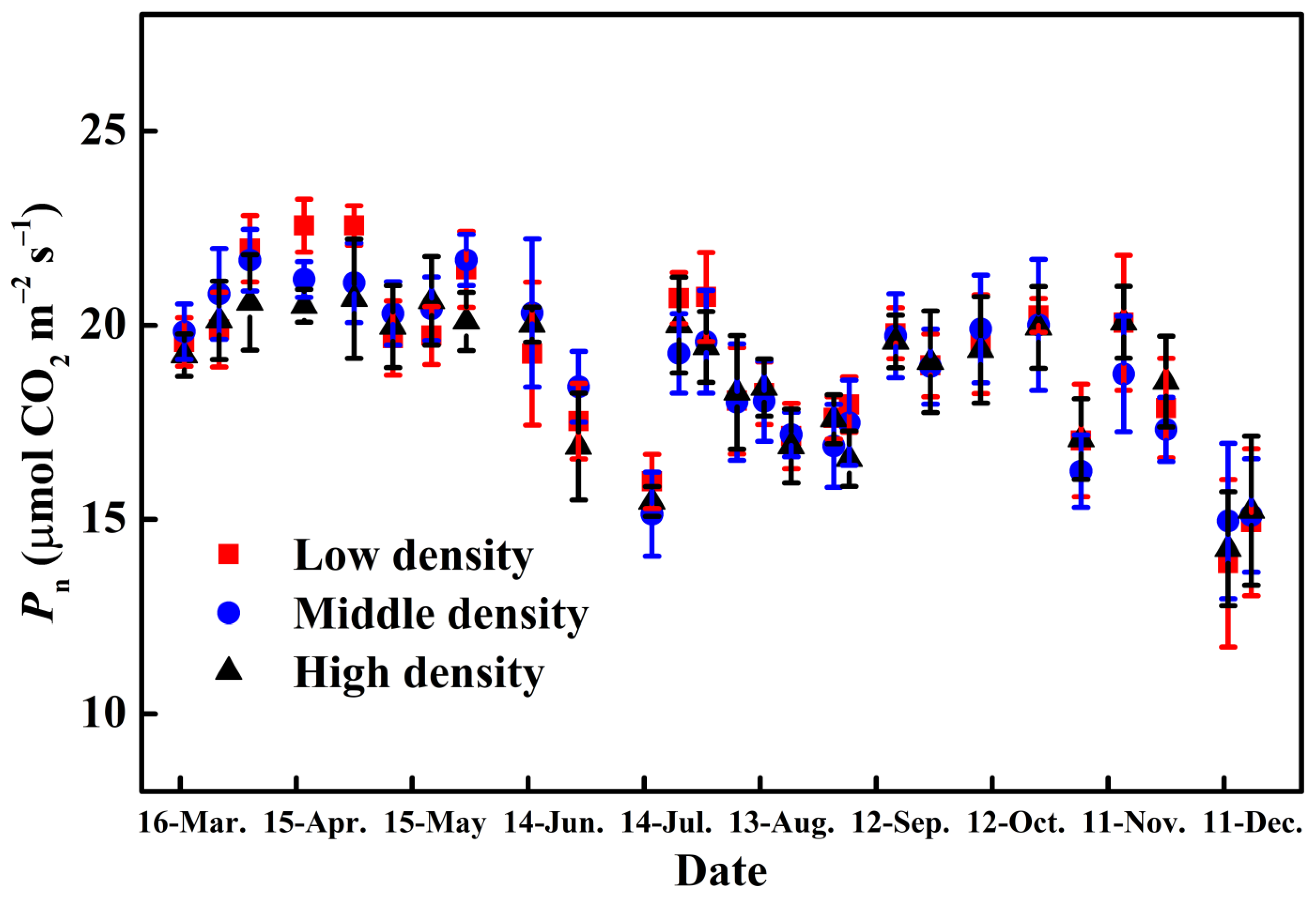
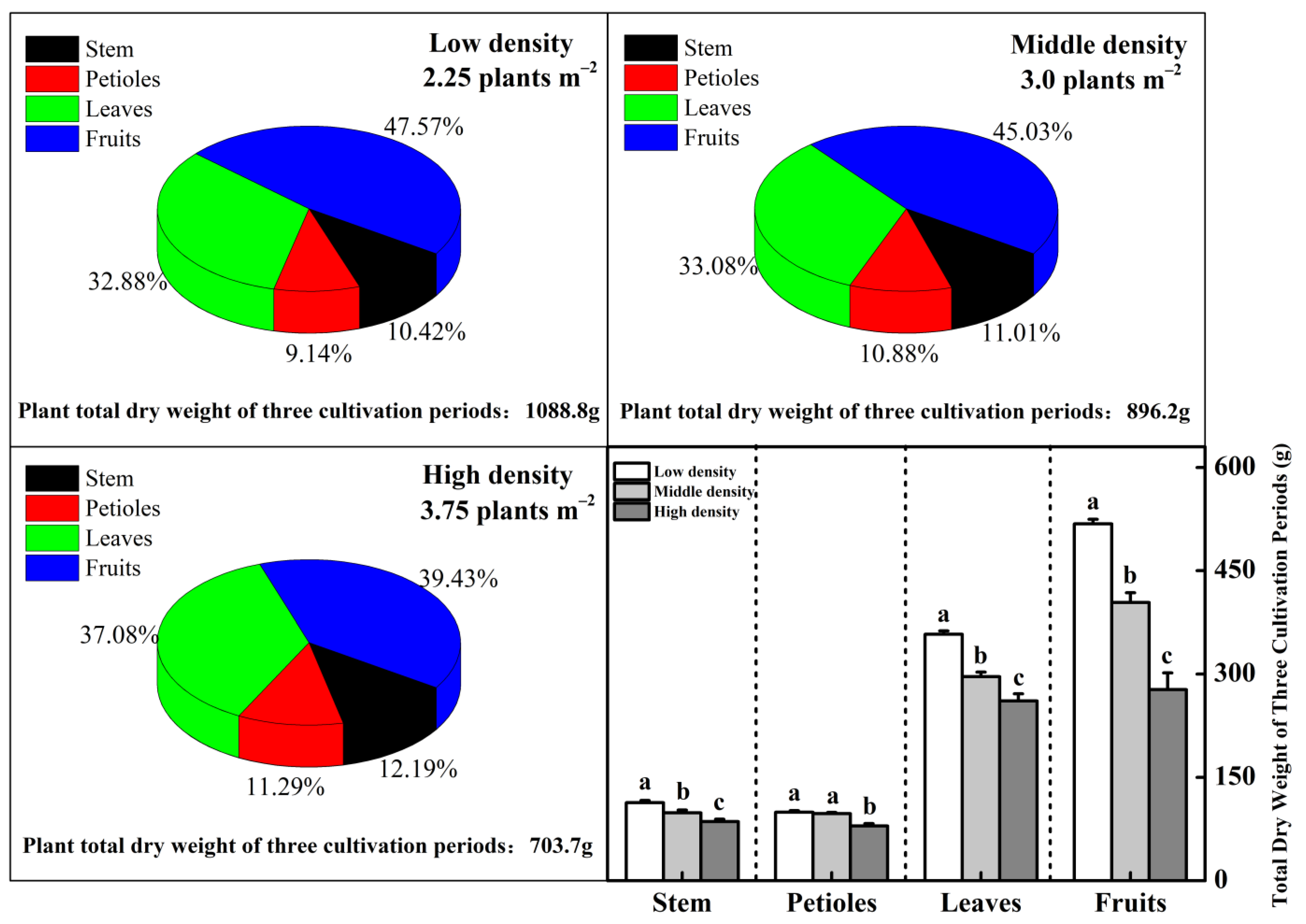
3.3. Leaf Photosynthesis and Dry Matter Distribution for Different Plant Density Treatments
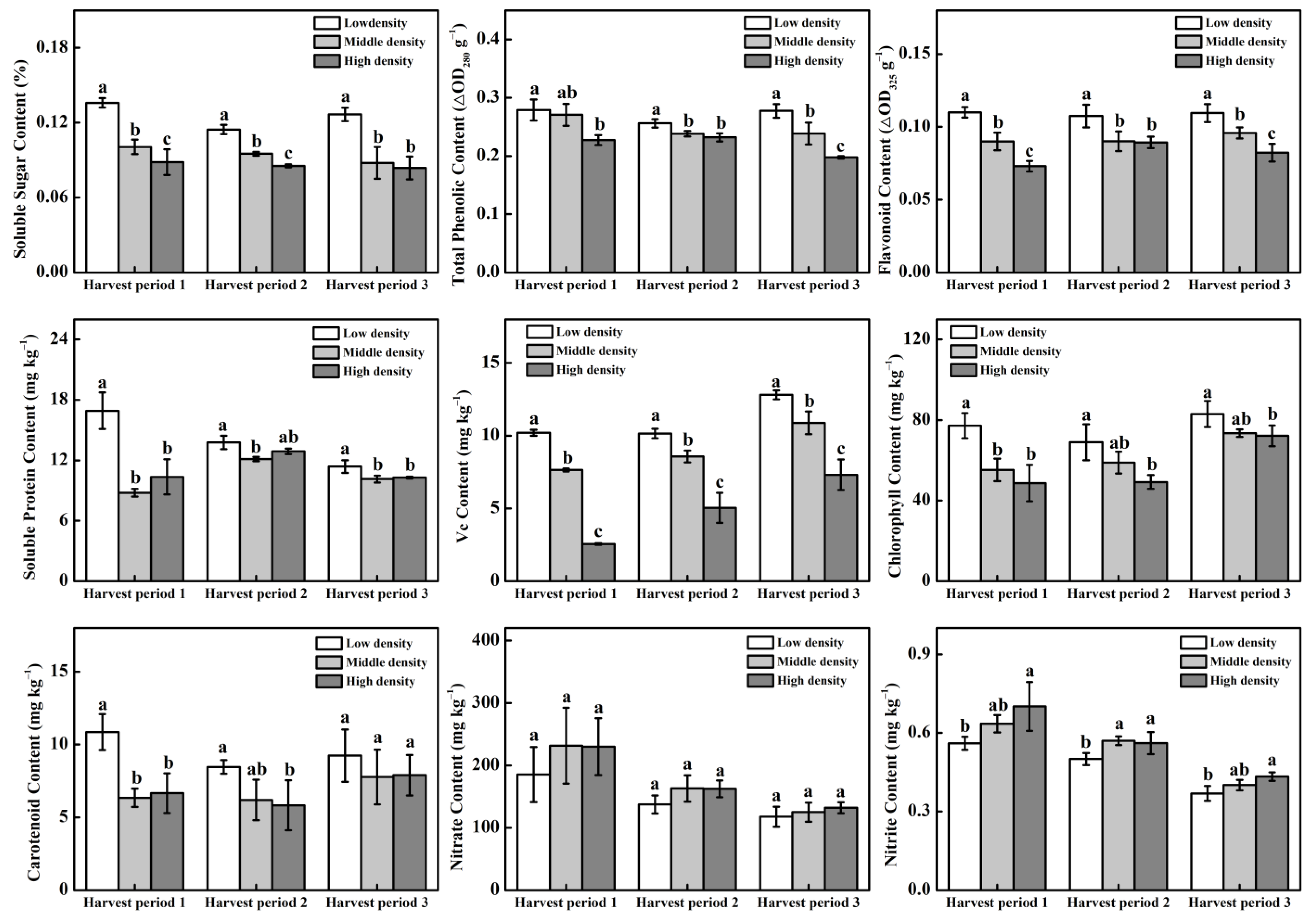
3.4. Cucumber Fruit Quality for Different Plant Density Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, W.M.; Chen, Y.Y.; Yang, S.J.; Wan, Y.H. Shanghai Greenhouse Vegetable Production and Agriculture Tour Experience. Acta Hortic. 2008, 797, 449–453. [Google Scholar]
- Ding, X.T.; Jiang, Y.P.; Hui, D.F.; He, L.Z.; Huang, D.F.; Yu, J.Z.; Zhou, Q. Model Simulation of Cucumber Yield and Microclimate Analysis in a Semi-Closed Greenhouse in China. HortScience 2019, 54, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Qian, T.T.; Zheng, X.G.; Yang, J.; Xu, Y.Y.; Wang, Y.; Zhou, Q.; Lu, S.L.; Ding, X.T. Optimal Utilization of Light Energy in Semi-Closed Greenhouse Using Three-Dimensional Cucumber Model. Sci. Program. 2020, 2020, 8855063. [Google Scholar]
- Gao, L.Y.; Hao, N.; Wu, T.; Cao, J.J. Advances in Understanding and Harnessing the Molecular Regulatory Mechanisms of Vegetable Quality. Front. Plant Sci. 2022, 13, 836515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.P.; Feng, S.J.; Yuan, J.; Wang, C.; Lu, T.; Wang, H.S.; Yu, C. The Formation of Fruit Quality in Cucumis Sativus, L. Front. Plant Sci. 2021, 12, 729448. [Google Scholar] [CrossRef] [PubMed]
- Łaźny, R.; Mirgos, M.; Przybył, J.L.; Nowak, J.S.; Kunka, M.; Gajc-Wolska, J.; Kowalczyk, K. Effect of Re-Used Lignite and Mineral Wool Growing Mats on Plant Growth, Yield and Fruit Quality of Cucumber and Physical Parameters of Substrates in Hydroponic Cultivation. Agronomy 2021, 11, 998. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Leszczyńska, T.; Borczak, B. Influence of Packages on Nutritional Quality of Pickled Chilled Stored Cucumbers. Ecol. Chem. Eng. A 2016, 23, 357–371. [Google Scholar]
- Alsadon, A.; Al-Helal, I.; Ibrahim, A.; Abdel-Ghany, A.; Al-Zaharani, S.; Ashour, T. The Effects of Plastic Greenhouse Covering on Cucumber (Cucumis Sativus, L.) Growth. Ecol. Eng. 2016, 87, 305–312. [Google Scholar] [CrossRef]
- Ali, A.B.; Elshaikh, N.A.; Hussien, G.; Abdallah, F.E.; Hassan, S. Biochar Addition for Enhanced Cucumber Fruit Quality under Deficit Irrigation. Biosci. J. 2020, 36, 1930–1937. [Google Scholar] [CrossRef]
- Ali, A.; Ghani, M.I.; Ding, H.Y.; Fan, Y.; Cheng, Z.H.; Iqbal, M. Co-Amended Synergistic Interactions between Arbuscular Mycorrhizal Fungi and the Organic Substrate-Induced Cucumber Yield and Fruit Quality Associated with the Regulation of the Am-Fungal Community Structure under Anthropogenic Cultivated Soil. Int. J. Mol. Sci. 2019, 20, 1539. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.L.; Li, X.; Nazim, G.; Duan, Z.Q. Interactive Effects of Elevated Carbon Dioxide and Nitrogen Availability on Fruit Quality of Cucumber (Cucumis Sativus, L.). J. Integr. Agric. 2018, 17, 2438–2446. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Reyes-Pérez, J.J.; Ramírez-Rodríguez, S.C.; Salas-Pérez, L.; Fortis-Hernández, M.; Murillo-Amador, B.; Troyo-Diéguez, E. Foliar Aspersion of Salicylic Acid Improves Phenolic and Flavonoid Compounds, and Also the Fruit Yield in Cucumber (Cucumis Sativus, L.). Plants 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcelis, L.F.M.; Broekhuijsen, A.G.M.; Meinen, E.; Nijs, E.; Raaphorst, M.G.M. Quantification of the Growth Response to Light Quantity of Greenhouse Grown Crops. In Proceedings of the V International Symposium on Artificial Lighting in Horticulture, Lillehammer, Norway, 21–24 June 2005; Volume 711, pp. 97–104. [Google Scholar]
- Hao, X.; Zheng, J.M.; Little, C.; Khosla, S. LED Inter-Lighting in Year-Round Greenhouse Mini-Cucumber Production. In Proceedings of the VII International Symposium on Light in Horticultural Systems, Wageningen, The Netherlands, 15–18 October 2012; Volume 956, pp. 335–340. [Google Scholar]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for Energy Efficient Greenhouse Lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Jin, N.; Jin, L.; Xiao, X.M.; Hu, L.; Liu, Z.C.; Wu, Y.; Xie, Y.D.; Zhu, W.; Lyu, J.; et al. Response of Tomato Fruit Quality Depends on Period of LED Supplementary Light. Front. Nutr. 2022, 9, 833723. [Google Scholar] [CrossRef]
- Alsina, I.; Erdberga, I.; Duma, M.; Alksnis, R.; Dubova, L. Changes in Greenhouse Grown Tomatoes Metabolite Content Depending on Supplemental Light Quality. Front. Nutr. 2022, 9, 830186. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Guo, X.; Chen, X.; Khosla, S. Inter-Lighting in Mini-Cucumbers: Interactions with Overhead Lighting and Plant Density. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC 2014), Brisbane, Australia, 17–22 August 2014; Volume 1107, pp. 291–296. [Google Scholar]
- Russo, V.M. Plant Density and Nitrogen Fertilizer Rate on Yield and Nutrient Content of Onion Developed from Greenhouse-Grown Transplants. HortScience 2008, 43, 1759–1764. [Google Scholar] [CrossRef] [Green Version]
- Calori, A.H.; Factor, T.L.; Feltran, J.C.; Watanabe, E.Y.; de Moraes, C.C.; Purquerio, L.F.V. Electrical Conductivity of the Nutrient Solution and Plant Density in Aeroponic Production of Seed Potato under Tropical Conditions (Winter/Spring). Bragantia 2017, 76, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Jovicich, E.; Cantliffe, D.J.; Stoffella, P.J. Fruit Yield and Quality of Greenhouse-Grown Bell Pepper as Influenced by Density, Container, and Trellis System. HortTechnology 2004, 14, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Obaid, A.A.; Khalil, N.H.; Al-Alawy, H.H.; Fahmi, A.H. Effect of Planting Density, Foliar Spraying and Overlapping System on the Growth and Productivity Using Soilless Culture System. J. Saudi Soc. Agric. Sci. 2022, in press. [Google Scholar]
- Logendra, L.S.; Gianfagna, T.J.; Specca, D.R.; Janes, H.W. Greenhouse Tomato Limited Cluster Production Systems: Crop Management Practices Affect Yield. HortScience 2001, 36, 893–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.T.; Zhang, H.M.; Qian, T.T.; He, L.Z.; Jin, H.J.; Zhou, Q.; Yu, J.Z. Nutrient Concentrations Induced Abiotic Stresses to Sweet Pepper Seedlings in Hydroponic Culture. Plants 2022, 11, 1098. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugar and related substance. Anal. Chem. 1956, 3, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Qi, Z. Instruction in Plant Physiological Experiment; China Agricultural Press: Beijing, China, 2003. [Google Scholar]
- Dewanto, V.; Wu, X.Z.; Adom, K.K.; Liu, R.H. Thermal Processing Enhances the Nutritional Value of Tomatoes by Increasing Total Antioxidant Activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Kaur, R.; Gupta, A.K.; Taggar, G.K. Nitrate Reductase and Nitrite as Additional Components of Defense System in Pigeonpea (Cajanus Cajan, L.) against Helicoverpa Armigera Herbivory. Pestic. Biochem. Physiol. 2014, 115, 39–47. [Google Scholar] [CrossRef]
- Papadopoulos, A.P.; Ormrod, D.P. Plant Spacing Effects on Yield of the Greenhouse Tomato. Can. J. Plant Sci. 1990, 70, 565–573. [Google Scholar] [CrossRef]
- Monge-Pérez, J.E. Effect of Pruning and Plant Density on Yield and Quality of Bell Pepper (Capsicum Annuum, L.) Grown under Greenhouse Conditions in Costa Rica. Rev. Tecnol. Marcha 2016, 29, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Marcelis, L.F.M. Fruit Growth and Biomass Allocation to the Fruits in Cucumber. 1. Effect of Fruit Load and Temperature. Sci. Hortic. 1993, 54, 107–121. [Google Scholar] [CrossRef]
- Marcelis, L.F.M. Fruit Growth and Biomass Allocation to the Fruits in Cucumber. 2. Effect of Irradiance. Sci. Hortic. 1993, 54, 123–130. [Google Scholar] [CrossRef]
- Qiu, R.J.; Song, J.J.; Du, T.S.; Kang, S.Z.; Tong, L.; Chen, R.Q.; Wu, L.S. Response of Evapotranspiration and Yield to Planting Density of Solar Greenhouse Grown Tomato in Northwest China. Agric. Water Manag. 2013, 130, 44–51. [Google Scholar] [CrossRef]
- Pao, Y.-C.; Kahlen, K.; Chen, T.-W.; Wiechers, D.; Stützel, H. How Does Structure Matter? Comparison of Canopy Photosynthesis Using One-and Three-Dimensional Light Models: A Case Study Using Greenhouse Cucumber Canopies. Silico Plants 2021, 3, diab031. [Google Scholar] [CrossRef]
- Alba, R.; Cordonnier-Pratt, M.-M.; Pratt, L.H. Fruit-Localized Phytochromes Regulate Lycopene Accumulation Independently of Ethylene Production in Tomato. Plant Physiol. 2000, 123, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Adams-Phillips, L.; Barry, C.; Giovannoni, J. Signal Transduction Systems Regulating Fruit Ripening. Trends Plant Sci. 2004, 9, 331–338. [Google Scholar] [CrossRef]
- Goularte, A.C.; Capello, C.; Valencia, G.A. Recovery of Chlorophylls from Cucumber (Cucumis Sativus, L.) Peel Using a Synthetic Layered Silicate. Children 2020, 5, 6. [Google Scholar]
- Wang, M.; Chen, L.; Liang, Z.J.; He, X.M.; Liu, W.R.; Jiang, B.; Yan, J.Q.; Sun, P.Y.; Cao, Z.Q.; Peng, Q.W.; et al. Metabolome and Transcriptome Analyses Reveal Chlorophyll and Anthocyanin Metabolism Pathway Associated with Cucumber Fruit Skin Color. BMC Plant Biol. 2020, 20, 386. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.E.; Son, S.Y.; Lee, C.H. Comparison of Metabolome and Functional Properties of Three Korean Cucumber Cultivars. Front. Plant Sci. 2022, 13, 882120. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H. Role of Antioxidants in the Skin: Anti-Aging Effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.T.; Jiang, Y.P.; Zhao, H.; Guo, D.D.; He, L.Z.; Liu, F.G.; Zhou, Q.; Nandwani, D.; Hui, D.F.; Yu, J.Z. Electrical Conductivity of Nutrient Solution Influenced Photosynthesis, Quality, and Antioxidant Enzyme Activity of Pakchoi (Brassica Campestris, L. ssp. Chinensis) in a Hydroponic System. PLoS ONE 2018, 13, e0202090. [Google Scholar] [CrossRef]
- Rezaei, M.; Fani, A.; Moini, A.L.; Mirzajani, P.; Malekirad, A.A.; Rafiei, M. Determining Nitrate and Nitrite Content in Beverages, Fruits, Vegetables, and Stews Marketed in Arak, Iran. Int. Sch. Res. Not. 2014, 2014, 439702. [Google Scholar] [CrossRef] [PubMed]
- Omer, C.; Nisan, Z.; Rav-David, D.; Elad, Y. Effects of Agronomic Practices on the Severity of Sweet Basil Downy Mildew (Peronospora Belbahrii). Plants 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Andrieu, B.; Vos, J.; van der Werf, W.; Fournier, C.; Evers, J.B. Towards Modelling the Flexible Timing of Shoot Development: Simulation of Maize Organogenesis Based on Coordination within and between Phytomers. Ann. Bot. 2014, 114, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Savvides, A.; van Ieperen, W.; Dieleman, J.A.; Marcelis, L.F.M. Phenotypic Plasticity to Altered Apical Bud Temperature in Cucumis Sativus: More Leaves-smaller Leaves and Vice Versa. Plant. Cell Environ. 2017, 40, 69–79. [Google Scholar] [CrossRef]
- Yu, B.W.; Yan, S.W.; Zhou, H.Y.; Dong, R.Y.; Lei, J.J.; Chen, C.M.; Cao, B.H. Overexpression of CsCaM3 Improves High Temperature Tolerance in Cucumber. Front. Plant Sci. 2018, 9, 797. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | kg/1000 L | A | g/1000 L | B | kg/1000 L |
|---|---|---|---|---|---|
| Ca(NO3)2·4H2O | 175 | MnSO4·H2O | 180 | KNO3 | 50 |
| EDTA-Fe (13%Fe) | 1.5 | ZnSO4·7H2O | 125 | KH2PO4 | 25 |
| Na2B4O7·10H2O | 280 | K2SO4 | 10 | ||
| CuSO4·5H2O | 25 | MgSO4·7H2O | 60 | ||
| Na2MoO4·2H2O | 15 |
| Climate Parameters | The First Harvest Period (4 April 2017–2 July 2017) | The Second Harvest Period (19 July 2017–7 September 2017) | The Third Harvest Period (3 October 2017–2 January 2018) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Max | Min | AV | Max | Min | AV | Max | Min | AV | |
| Outside temperature (°C) | 33.19 | 8.1 | 20.26 | 39.79 | 21.09 | 28.82 | 29.6 | −2.68 | 12.03 |
| Greenhouse temperature (°C) | 36.27 | 16.2 | 23.11 | 38.2 | 20.7 | 27.32 | 33 | 13.68 | 20.12 |
| Daily radiation sum (KJ cm−2) | 2.945 | 0.160 | 1.696 | 2.715 | 0.480 | 1.782 | 1.833 | 0.119 | 0.906 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Nie, W.; Qian, T.; He, L.; Zhang, H.; Jin, H.; Cui, J.; Wang, H.; Zhou, Q.; Yu, J. Low Plant Density Improves Fruit Quality without Affecting Yield of Cucumber in Different Cultivation Periods in Greenhouse. Agronomy 2022, 12, 1441. https://doi.org/10.3390/agronomy12061441
Ding X, Nie W, Qian T, He L, Zhang H, Jin H, Cui J, Wang H, Zhou Q, Yu J. Low Plant Density Improves Fruit Quality without Affecting Yield of Cucumber in Different Cultivation Periods in Greenhouse. Agronomy. 2022; 12(6):1441. https://doi.org/10.3390/agronomy12061441
Chicago/Turabian StyleDing, Xiaotao, Wenfeng Nie, Tingting Qian, Lizhong He, Hongmei Zhang, Haijun Jin, Jiawei Cui, Hong Wang, Qiang Zhou, and Jizhu Yu. 2022. "Low Plant Density Improves Fruit Quality without Affecting Yield of Cucumber in Different Cultivation Periods in Greenhouse" Agronomy 12, no. 6: 1441. https://doi.org/10.3390/agronomy12061441
APA StyleDing, X., Nie, W., Qian, T., He, L., Zhang, H., Jin, H., Cui, J., Wang, H., Zhou, Q., & Yu, J. (2022). Low Plant Density Improves Fruit Quality without Affecting Yield of Cucumber in Different Cultivation Periods in Greenhouse. Agronomy, 12(6), 1441. https://doi.org/10.3390/agronomy12061441