
Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Crop Stem Characteristics and Lodging
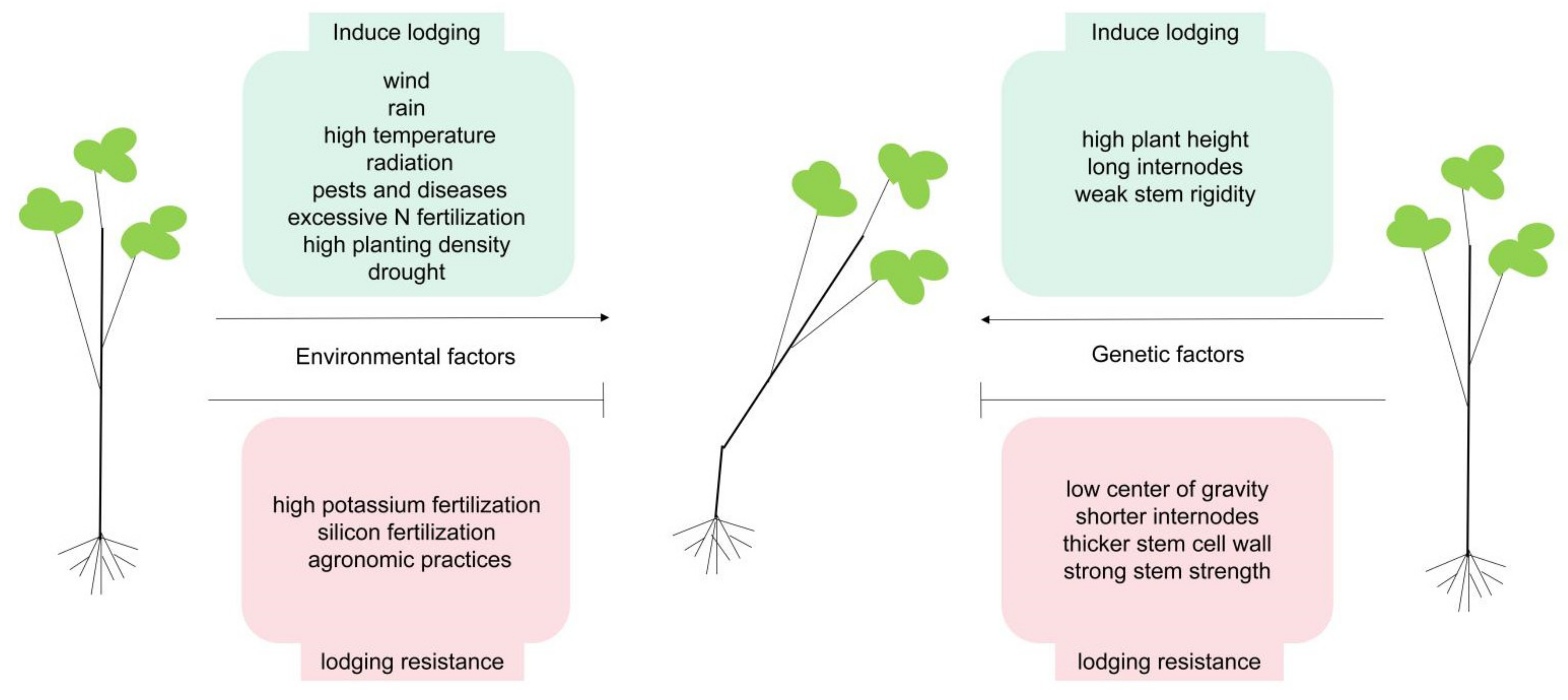
2.1. Environmental Factors Affecting Stem Lodging
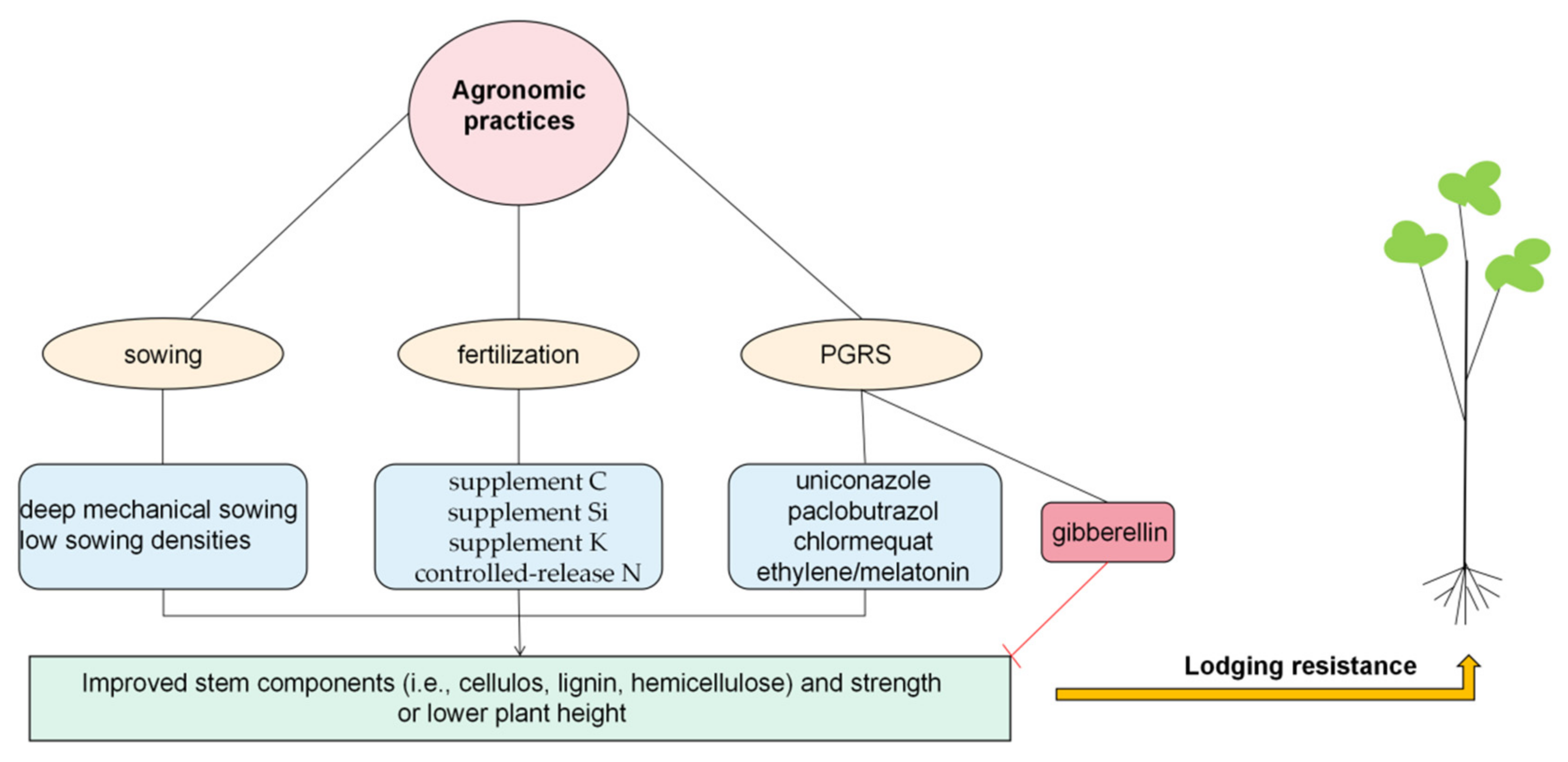
2.2. Agronomic Practices Affecting Stem Lodging
2.3. Genetic Factors Affecting Stem Lodging
2.4. Stem Anatomy and Lodging
2.5. Stem Components Affect Crop Lodging Resistance
2.5.1. Effects of Lignin on Lodging Resistance
2.5.2. Cellulose and Hemicellulose Influence Lodging Resistance
2.5.3. Effects of Carbon and Nitrogen Metabolism on Lodging Resistance
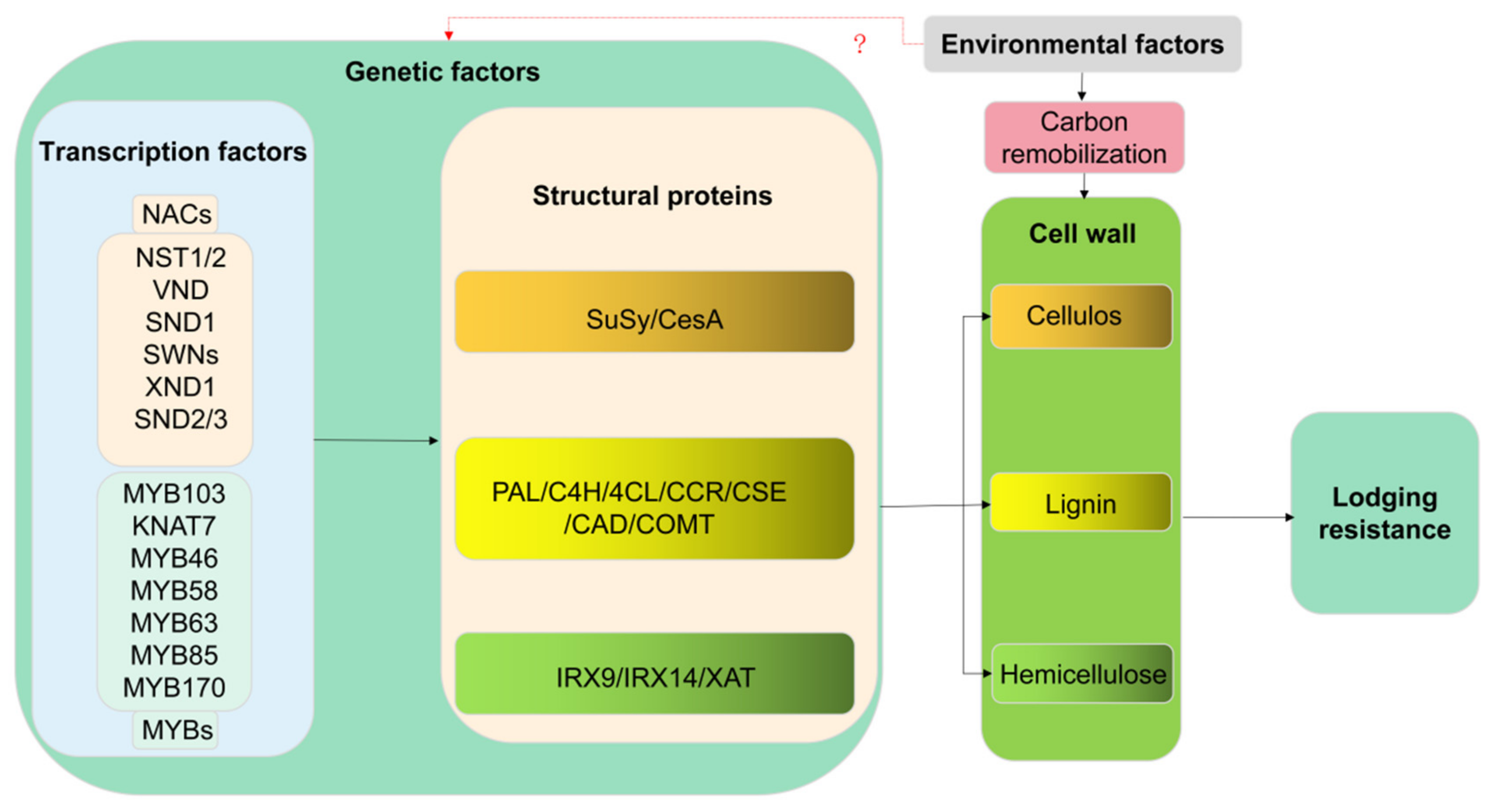
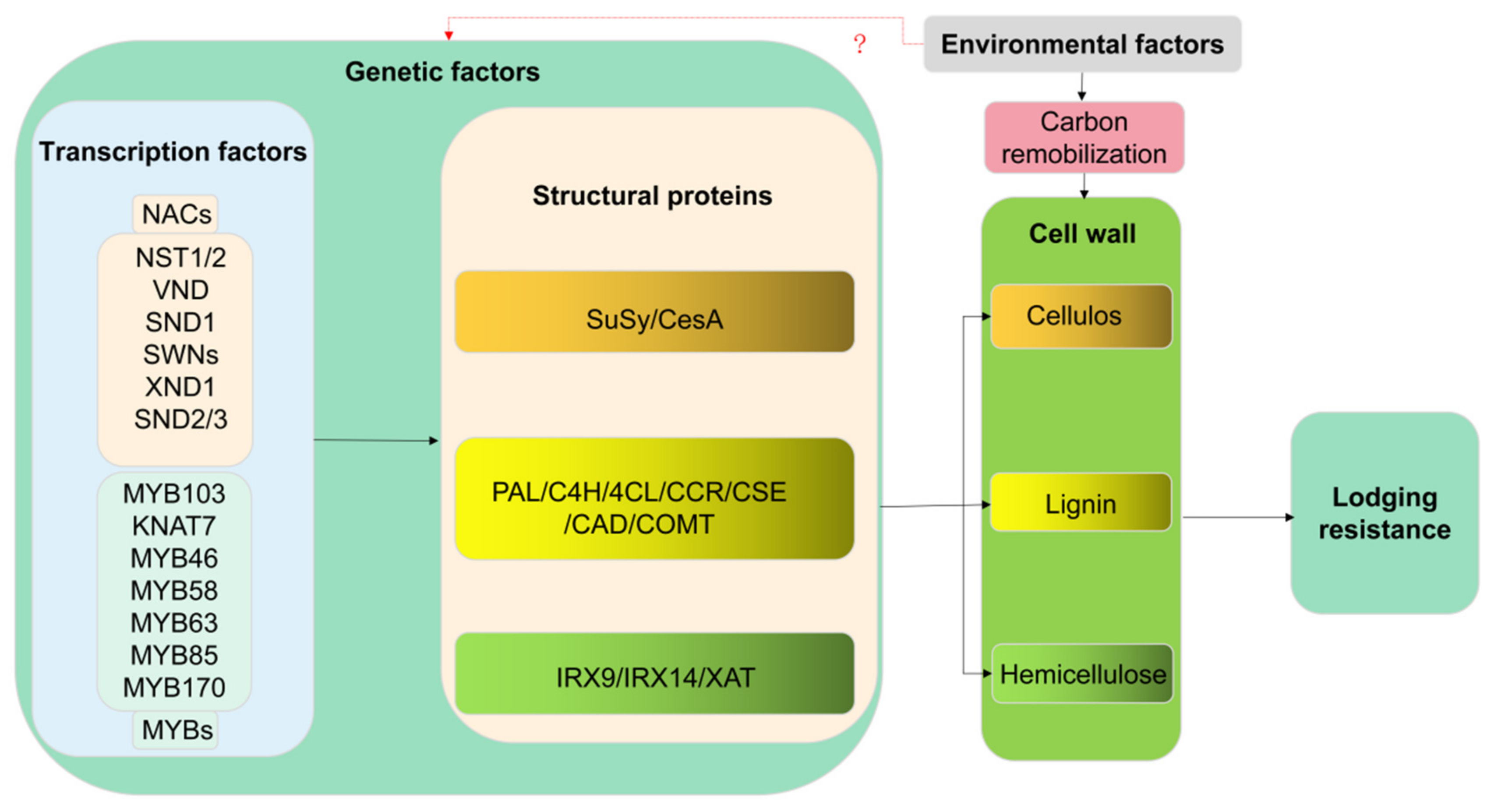
3. Anti-Lodging Molecular Regulatory Network Controlling Lignin, Cellulose and Hemicellulose
3.1. Transcriptional Regulation of Lignin, Cellulose, and Hemicellulose
3.2. Molecular Mechanisms of Lignin, Cellulose, and Hemicellulose Structural Genes in Resistance to Lodging
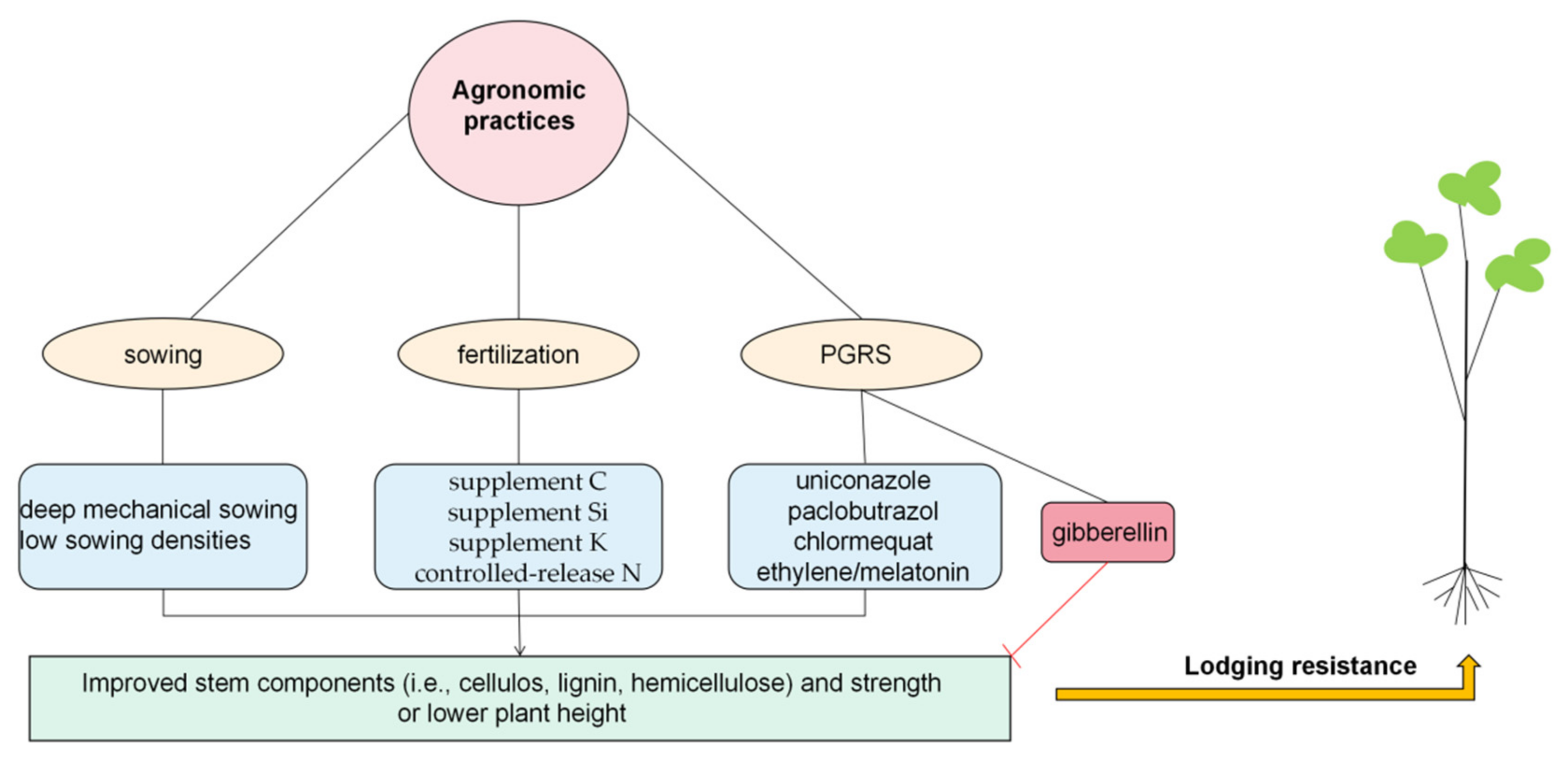
3.3. Agronomic Practices That Improve Levels of Lignin, Cellulose, and Hemicellulose
4. Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rajkumara, S. Lodging in cereals—A review. Agric. Rev. 2008, 29, 55. [Google Scholar]
- Berry, P.; Sterling, M.; Spink, J.; Baker, C.; Sylvester-Bradley, R.; Mooney, S.; Tams, A.; Ennos, A. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 215–269. [Google Scholar]
- Pu, D.F.; Zhou, J.R.; Li, B.F.; Li, Q.; Zhou, Q. Study on the evaluation method of root failure resistance of wheat. Acta Agric. Boreali-Sin. 2000, 9, 58–61. [Google Scholar] [CrossRef]
- Lindsey, A.J.; Carter, P.R.; Thomison, P.R. Impact of imposed root lodging on corn growth and yield. Agron. J. 2021, 113, 5054–5062. [Google Scholar] [CrossRef]
- Gu, H. Inheritance and QTL analysis of lodging resistance in Brassica napus L. Nanjing Agric. Univ. 2008. [Google Scholar] [CrossRef]
- Du, J.; Zhang, Y.; Guo, X.; Ma, L.; Shao, M.; Pan, X.; Zhao, C. Micron-scale phenotyping quantification and three-dimensional microstructure reconstruction of vascular bundles within maize stalks based on micro-CT scanning. Funct. Plant Biol. 2016, 44, 10–22. [Google Scholar] [CrossRef]
- Brulé, V.; Rafsanjani, A.; Pasini, D.; Western, T.L. Hierarchies of plant stiffness. Plant Sci. 2016, 250, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.U.; Reynolds, T.P.; Ramage, M.H. The strength of plants: Theory and experimental methods to measure the mechanical properties of stems. J. Exp. Bot. 2017, 68, 4497–4516. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, S.C.; Sayre, K.; Kaul, J.; Narang, R. Growth and morphology of spring wheat (Triticum aestivum L.) culms and their association with lodging: Effects of genotypes, N levels and ethephon. Field Crops Res. 2003, 84, 271–290. [Google Scholar] [CrossRef]
- Liu, W.; Deng, Y.; Hussain, S.; Zou, J.; Yuan, J.; Luo, L.; Yang, C.; Yuan, X.; Yang, W. Relationship between cellulose accumulation and lodging resistance in the stem of relay intercropped soybean (Glycine max (L.) Merr.). Field Crops Res. 2016, 196, 261–267. [Google Scholar] [CrossRef]
- Yang, D.G.; Ma, D.Z.; Yu, Q.Q.; Sun, Y.J.; Gu, W.R.; Chai, M.Z.; Zhang, Q. Progess of research on factors influencing corn overturning and resistance to overturning. J. China Agric. Univ. 2020, 25, 28–38. [Google Scholar]
- Li, W.B.; Wu, H.; Liu, J.; Zhang, X.C.; Guo, Z.W.; Zheng, L.N.; Zhao, X.; Han, Y.P.; Teng, W.L. Genome-wide association analysis of traits related to soybean resistance to overturning. Acta Agric. Boreali-Sin. 2021, 52, 497. [Google Scholar]
- Yang, J.; Liu, W.H.; Liang, G.L.; Jia, Z.F.; Liu, K.Q.; Zhang, Y.; Wu, R.; Yang, Y.J. Analysis of relevant traits of different oat lines in alpine regions for resistance to downfall. Acta Pratac. Sin. 2020, 29, 50–60. [Google Scholar]
- Wu, W.; Ma, B.L. Erect–leaf posture promotes lodging resistance in oat plants under high plant population. Eur. J. Agron. 2019, 103, 175–187. [Google Scholar] [CrossRef]
- Robertson, D.J.; Lee, S.Y.; Julias, M.; Cook, D.D. Maize stalk lodging: Flexural stiffness predicts strength. Crop Sci. 2016, 56, 1711–1718. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Feng, S.; Ru, Z.; Li, G.; Zhang, Z.; Wang, Z. Rapid determination of single-stalk and population lodging resistance strengths and an assessment of the stem lodging wind speeds for winter wheat. Field Crops Res. 2012, 139, 1–8. [Google Scholar] [CrossRef]
- Niu, L.; Feng, S.; Ding, W.; Li, G. Influence of speed and rainfall on large-scale wheat lodging from 2007 to 2014 in China. PLoS ONE 2016, 11, e0157677. [Google Scholar] [CrossRef]
- Weng, F.; Zhang, W.; Wu, X.; Xu, X.; Ding, Y.; Li, G.; Liu, Z.; Wang, S. Impact of low-temperature, overcast and rainy weather during the reproductive growth stage on lodging resistance of rice. Sci. Rep. 2017, 7, 46596. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Xiong, H.C.; Li, Y.T.; Guo, H.J.; Xie, Y.D.; Zhao, L.S.; Gu, J.Y.; Zhao, S.R.; Ding, Y.P.; Song, X.Y. Genetic analysis and QTL mapping of a novel reduced height gene in common wheat (Triticum aestivum L.). J. Integr. Agric. 2020, 19, 1721–1730. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, X.; Qian, Q.; Liu, L.; Dong, G.; Xiong, G.; Zeng, D.; Zhou, Y. Golgi nucleotide sugar transporter modulates cell wall biosynthesis and plant growth in rice. Proc. Natl. Acad. Sci. USA 2011, 108, 5110–5115. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Jia, Z.; Ma, X.; Ma, H.; Zhao, Y. Characterising the morphological characters and carbohydrate metabolism of oat culms and their association with lodging resistance. Plant Biol. 2020, 22, 267–276. [Google Scholar] [CrossRef]
- Mizuno, H.; Kasuga, S.; Kawahigashi, H. Root lodging is a physical stress that changes gene expression from sucrose accumulation to degradation in sorghum. BMC Plant Biol. 2018, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Iqbal, N.; Rahman, T.; Liu, T.; Brestic, M.; Safdar, M.E.; Asghar, M.A.; Farooq, M.U.; Shafiq, I.; Ali, A. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes. Environ. Exp. Bot. 2019, 162, 374–382. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Song, Y.; Liu, Z.; Yang, C.; Tang, S.; Zheng, C.; Wang, S.; Ding, Y. Lodging resistance characteristics of high-yielding rice populations. Field Crops Res. 2014, 161, 64–74. [Google Scholar] [CrossRef]
- Kumar, M.; Campbell, L.; Turner, S. Secondary cell walls: Biosynthesis and manipulation. J. Exp. Bot. 2016, 67, 515–531. [Google Scholar] [CrossRef]
- Ogden, M.; Hoefgen, R.; Roessner, U.; Persson, S.; Khan, G.A. Feeding the walls: How does nutrient availability regulate cell wall composition? Int. J. Mol. Sci. 2018, 19, 2691. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Fei, G.; Ren, B.; Dong, S.; Peng, L.; Bin, Z.; Zhang, J. Lignin metabolism regulates lodging resistance of maize hybrids under varying planting density. J. Integr. Agric. 2021, 20, 2077–2089. [Google Scholar] [CrossRef]
- Manga-Robles, A.; Santiago, R.; Malvar, R.A.; Moreno-González, V.; Fornalé, S.; López, I.; Centeno, M.L.; Acebes, J.L.; Álvarez, J.M.; Caparros-Ruiz, D. Elucidating compositional factors of maize cell walls contributing to stalk strength and lodging resistance. Plant Sci. 2021, 307, 110882. [Google Scholar] [CrossRef]
- Wu, W.; Shah, F.; Duncan, R.W.; Ma, B.L. Grain yield, root growth habit and lodging of eight oilseed rape genotypes in response to a short period of heat stress during flowering. Agric. For. Meteorol. 2020, 287, 107954. [Google Scholar] [CrossRef]
- Tu, B.; Tao, Z.; Wang, S.G.; Zhou, L.; Zheng, L.; Zhang, C.; Li, X.Z.; Zhang, X.Y.; Yin, J.J.; Zhu, X.; et al. Loss of Gn1a/OsCKX2 confers heavy-panicle rice with excellent lodging resistance. J. Integr. Plant Biol. 2022, 64, 23–38. [Google Scholar] [CrossRef]
- Xue, J.; Gou, L.; Zhao, Y.; Yao, M.; Yao, H.; Tian, J.; Zhang, W. Effects of light intensity within the canopy on maize lodging. Field Crops Res. 2016, 188, 133–141. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Q. Evaluation method of stem lodging resistance in wheat. Acta Agric. Boreali-Sin. 1995, 10, 84–88. [Google Scholar]
- Wang, X.Q.; Song, W.; Zhang, R.Y.; Chen, Y.N.; Sun, X.; Zhao, J.R. Research progress on the genetics of stalk overturning resistance in maize. J. China Agric. Univ. 2021, 54, 2261–2272. [Google Scholar]
- Hussain, S.; Liu, T.; Iqbal, N.; Brestic, M.; Pang, T.; Mumtaz, M.; Shafiq, I.; Li, L.S.; Gao, Y.; Khan, A.; et al. Effects of lignin, cellulose, hemicellulose, sucrose and monosaccharide carbohydrates on soybean physical stem strength and yield in intercropping. Photochem. Photobiol. Sci. 2020, 19, 462–472. [Google Scholar] [CrossRef]
- Feng, S.; Kong, D.; Ding, W.; Ru, Z.; Li, G.; Niu, L. A novel wheat lodging resistance evaluation method and device based on the thrust force of the stalks. PLoS ONE 2019, 14, e0224732. [Google Scholar] [CrossRef]
- Stubbs, C.J.; Oduntan, Y.A.; Keep, T.R.; Noble, S.D.; Robertson, D.J. The effect of plant weight on estimations of stalk lodging resistance. Plant Methods 2020, 16, 128. [Google Scholar] [CrossRef]
- Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ahmad, A.; Asif, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W. Improving lodging resistance: Using wheat and rice as classical examples. Int. J. Mol. Sci. 2019, 20, 4211. [Google Scholar] [CrossRef] [Green Version]
- Berry, P.; Spink, J. Predicting yield losses caused by lodging in wheat. Field Crops Res. 2012, 137, 19–26. [Google Scholar] [CrossRef]
- Chapman, N.H.; Burt, C.; Nicholson, P. The identification of candidate genes associated with Pch2 eyespot resistance in wheat using cDNA-AFLP. Theor. Appl. Genet. 2009, 118, 1045–1057. [Google Scholar] [CrossRef]
- Srinivasa Reddy, P.; Fakrudin, B.; Punnuri, S.; Arun, S.; Kuruvinashetti, M.; Das, I.; Seetharama, N. Molecular mapping of genomic regions harboring QTLs for stalk rot resistance in sorghum. Euphytica 2008, 159, 191–198. [Google Scholar] [CrossRef]
- Dobermann, A.; Witt, C.; Abdulrachman, S.; Gines, H.; Nagarajan, R.; Son, T.; Tan, P.; Wang, G.; Chien, N.; Thoa, V. Soil fertility and indigenous nutrient supply in irrigated rice domains of Asia. Agron. J. 2003, 95, 913–923. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain filling of cereals under soil drying. New Phytol. 2006, 169, 223–236. [Google Scholar] [CrossRef]
- Sterling, M.; Baker, C.; Berry, P.; Wade, A. An experimental investigation of the lodging of wheat. Agric. For. Meteorol. 2003, 119, 149–165. [Google Scholar] [CrossRef]
- Xue, J.; Gou, L.; Shi, Z.G.; Zhao, Y.S.; Zhang, W.F. Effect of leaf removal on photosynthetically active radiation distribution in maize canopy and stalk strength. J. Integr. Agric. 2017, 16, 85–96. [Google Scholar] [CrossRef]
- Lyu, X.; Cheng, Q.; Qin, C.; Li, Y.; Xu, X.; Ji, R.; Mu, R.; Li, H.; Zhao, T.; Liu, J. GmCRY1s modulate gibberellin metabolism to regulate soybean shade avoidance in response to reduced blue light. Mol. Plant 2021, 14, 298–314. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B.L. Assessment of canola crop lodging under elevated temperatures for adaptation to climate change. Agric. For. Meteorol. 2018, 248, 329–338. [Google Scholar] [CrossRef]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, D.; Berry, P.; King, M. Effects of shade on root characters associated with lodging in wheat (Triticum aestivum). Ann. Appl. Biol. 2008, 152, 389–395. [Google Scholar] [CrossRef]
- Zhang, M.H.; Mo, Z.W.; Liao, J.; Pan, S.G.; Chen, X.F.; Zheng, L.; Luo, X.W.; Wang, Z.M. Lodging resistance related to root traits for mechanized wet-seeding of two super rice cultivars. Rice Sci. 2021, 28, 200–208. [Google Scholar]
- Yue, H.W.; Xie, J.L.; Peng, H.C.; Bu, J.Z.; Gai, S.Q. Comparative analysis of filling progress and pest damage in inverted maize. Heilongjiang Agric. Sci. 2010, 10, 31–33. [Google Scholar] [CrossRef]
- Goodman, A.; Ennos, A. The effects of soil bulk density on the morphology and anchorage mechanics of the root systems of sunflower and maize. Ann. Bot. 1999, 83, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Sposaro, M.M.; Chimenti, C.A.; Hall, A.J. Root lodging in sunflower. Variations in anchorage strength across genotypes, soil types, crop population densities and crop developmental stages. Field Crops Res. 2008, 106, 179–186. [Google Scholar] [CrossRef]
- Zuo, Q.; Kuai, J.; Zhao, L.; Hu, Z.; Wu, J.; Zhou, G. The effect of sowing depth and soil compaction on the growth and yield of rapeseed in rice straw returning field. Field Crops Res. 2017, 203, 47–54. [Google Scholar] [CrossRef]
- Bian, D.; Jia, G.; Cai, L.; Ma, Z.; Eneji, A.E.; Cui, Y. Effects of tillage practices on root characteristics and root lodging resistance of maize. Field Crops Res. 2016, 185, 89–96. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Dong, X.; Qian, T.; Yin, L.; Dong, S.; Chu, J.; He, M. Delayed sowing can increase lodging resistance while maintaining grain yield and nitrogen use efficiency in winter wheat. J. Crop 2017, 5, 541–552. [Google Scholar] [CrossRef]
- Xiang, D.B.; Zhao, G.; Wan, Y.; Tan, M.L.; Song, C.; Song, Y. Effect of planting density on lodging-related morphology, lodging rate, and yield of tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2016, 19, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Chen, J.; Shi, Y.; Li, Y.; Yin, Y.; Yang, D.; Luo, Y.; Pang, D.; Xu, X.; Li, W. Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat. Sci. Rep. 2017, 7, 41805. [Google Scholar] [CrossRef]
- Zhang, P.; Yan, Y.; Gu, S.; Wang, Y.; Xu, C.; Sheng, D.; Li, Y.; Wang, P.; Huang, S. Lodging resistance in maize: A function of root–shoot interactions. Eur. J. Agron. 2022, 132, 126393. [Google Scholar] [CrossRef]
- Echezona, B. Corn-stalk lodging and borer damage as influenced by varying corn densities and planting geometry with soybean (Glycine max. L. Merrill). Int. Agrophys. 2007, 21, 133–143. [Google Scholar]
- Chen, X.; Sun, N.; Gu, Y.; Liu, Y.; Li, J.; Wu, C.; Wang, Z. Maize-soybean strip intercropping improved lodging resistance and productivity of maize. Int. J. Agric. Biol. 2020, 24, 1383–1392. [Google Scholar]
- Fang, X.; Li, Y.; Nie, J.; Wang, C.; Huang, K.; Zhang, Y.; Zhang, Y.; She, H.; Liu, X.; Ruan, R. Effects of nitrogen fertilizer and planting density on the leaf photosynthetic characteristics, agronomic traits and grain yield in common buckwheat (Fagopyrum esculentum M.). Field Crops Res. 2018, 219, 160–168. [Google Scholar] [CrossRef]
- Huang, G.; Liu, Y.; Guo, Y.; Peng, C.; Tan, W.; Zhang, M.; Li, Z.; Zhou, Y.; Duan, L. A novel plant growth regulator improves the grain yield of high-density maize crops by reducing stalk lodging and promoting a compact plant type. Field Crops Res. 2021, 260, 107982. [Google Scholar] [CrossRef]
- Shu, M.; Gu, X.; Lin, S.; Zhu, J.; Yang, G.; Wang, Y.; Qian, S.; Zhou, L. Structural characteristics change and spectral response analysis of maize canopy under lodging stress. Spectrosc. Spectr. Anal. 2019, 39, 3553–3559. [Google Scholar]
- Shao, H.; Shi, D.; Shi, W.; Ban, X.; Chen, Y.; Ren, W.; Chen, F.; Mi, G. The impact of high plant density on dry matter remobilization and stalk lodging in maize genotypes with a different stay-green degree. Arch. Agron. Soil Sci. 2021, 67, 504–518. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B.L.; Fan, J.J.; Sun, M.; Yi, Y.; Guo, W.S.; Voldeng, H.D. Management of nitrogen fertilization to balance reducing lodging risk and increasing yield and protein content in spring wheat. Field Crops Res. 2019, 241, 107584. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, L.; Ding, Y.; Yao, X.; Wu, X.; Weng, F.; Li, G.; Liu, Z.; Tang, S.; Ding, C.; et al. Nitrogen fertilizer application affects lodging resistance by altering secondary cell wall synthesis in japonica rice (Oryza sativa). J. Plant Res. 2017, 130, 859–871. [Google Scholar] [CrossRef]
- Zhai, J.; Zhang, Y.; Zhang, G.; Tian, M.; Xie, R.; Ming, B.; Hou, P.; Wang, K.; Xue, J.; Li, S. Effects of nitrogen fertilizer management on stalk lodging resistance traits in summer maize. Agriculture 2022, 12, 162. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, L.; Ding, Y.; Fei, W.; Wu, X.; Li, G.; Liu, Z.; She, T.; Ding, C.; Wang, S. Top-dressing nitrogen fertilizer rate contributes to decrease culm physical strength by reducing structural carbohydrate content in japonica rice. J. Integr. Agric. 2016, 15, 992–1004. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, Y.; Zhai, W.; Tong, Z.; Shen, T.; Li, Y.C.; Zhang, M.; Sigua, G.C.; Chen, J.; Ding, F. Controlled-release nitrogen fertilizer improved lodging resistance and potassium and silicon uptake of direct-seeded rice. Crop Sci. 2019, 59, 2733–2740. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J. Carbohydrate, hormone and enzyme regulations of rice grain filling under post-anthesis soil drying. Environ. Exp. Bot. 2020, 178, 104165. [Google Scholar] [CrossRef]
- Dorairaj, D.; Ismail, M.R.; Sinniah, U.R.; Kar Ban, T. Influence of silicon on growth, yield, and lodging resistance of MR219, a lowland rice of Malaysia. J. Plant Nutr. 2017, 40, 1111–1124. [Google Scholar] [CrossRef]
- Hu, Y.; Javed, H.H.; Asghar, M.A.; Peng, X.; Brestic, M.; Skalický, M.; Ghafoor, A.Z.; Cheema, H.N.; Zhang, F.F.; Wu, Y.C. Enhancement of lodging resistance and lignin content by application of organic carbon and silicon fertilization in Brassica napus L. Front. Plant Sci. 2022, 13, 807048. [Google Scholar] [CrossRef]
- Kuai, J.; Sun, Y.; Guo, C.; Zhao, L.; Zuo, Q.; Wu, J.; Zhou, G. Root-applied silicon in the early bud stage increases the rapeseed yield and optimizes the mechanical harvesting characteristics. Field Crops Res. 2017, 200, 88–97. [Google Scholar] [CrossRef]
- Olagunju, S.O.; Atayese, M.O.; Sakariyawo, O.S.; Dare, E.O.; Nassir, A.L. Culm morphological traits contributing to lodging resistance in first generation NERICA cultivars under foliar application of orthosilicic acid fertilizer. Silicon 2021, 13, 3059–3073. [Google Scholar] [CrossRef]
- Zhang, T.; He, X.; Chen, B.; He, L.; Tang, X. Effects of Different Potassium (K) Fertilizer Rates on Yield Formation and Lodging of Rice. Phyton 2021, 90, 815–826. [Google Scholar] [CrossRef]
- Zaman, U.; Ahmad, Z.; Farooq, M.; Saeed, S.; Ahmad, M.; Wakeel, A. Potassium fertilization may improve stem strength and yield of Basmati rice grown on nitrogen-fertilized soils. Pak. J. Agric. Sci. 2015, 52, 439–445. [Google Scholar]
- Martin, S.A.; Darrah, L.L.; Hibbard, B.E. Divergent selection for rind penetrometer resistance and its effects on European corn borer damage and stalk traits in corn. Crop Sci. 2004, 44, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Santiago, R.; Butrón, A.; Revilla, P.; Malvar, R.A. Is the basal area of maize internodes involved in borer resistance? BMC Plant Biol. 2011, 11, 137. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Gao, S.; Hou, L.; Li, L.; Ming, B.; Xie, R.; Wang, K.; Hou, P.; Li, S. Physiological Influence of Stalk Rot on Maize Lodging after Physiological Maturity. Agron. J. 2021, 11, 2271. [Google Scholar] [CrossRef]
- Wu, W.; Huang, J.; Cui, K.; Nie, L.; Wang, Q.; Yang, F.; Shah, F.; Yao, F.; Peng, S. Sheath blight reduces stem breaking resistance and increases lodging susceptibility of rice plants. Field Crops Res. 2012, 128, 101–108. [Google Scholar] [CrossRef]
- Das, I.; Indira, S. Role of stalk-anatomy and yield parameters in development of charcoal rot caused by macrophomina phaseolina in winter sorghum. Phytoparasitica 2008, 36, 199–208. [Google Scholar] [CrossRef]
- Showler, A.T.; Wilson, B.E.; Reagan, T.E. Mexican rice borer (Lepidoptera: Crambidae) injury to corn greater than to sorghum and sugarcane under field conditions. J. Econ. Entomol. 2012, 105, 1597–1602. [Google Scholar] [CrossRef] [Green Version]
- Khobra, R.; Sareen, S.; Meena, B.K.; Kumar, A.; Tiwari, V.; Singh, G. Exploring the traits for lodging tolerance in wheat genotypes: A review. Physiol. Mol. Biol. Plants 2019, 25, 589–600. [Google Scholar] [CrossRef]
- Guo, Y.; Hu, Y.M.; Chen, H.; Yan, P.S.; Du, Q.G.; Wang, Y.F.; Wang, H.Q.; Wang, Z.H.; Kang, D.M.; Li, W.X. Identification of traits and genes associated with lodging resistance in maize. J. Crop 2021, 9, 1408–1417. [Google Scholar] [CrossRef]
- Duan, C.R.; Wang, B.C.; Wang, P.Q. Correlation between the structure of rice stalks and their properties. J. Chongqing Univ. 2003, 26, 38–40. [Google Scholar] [CrossRef]
- Liu, W.X.; Wang, C.Y.; Wang, Q.; Yue, P.L.; Xie, X.D.; Liu, G.L.; Ma, K.; Lu, H.F. Stalk resistance characteristics of different maize varieties and their relationship with yield. Henan Agric. Sci. 2015, 44, 17–21. [Google Scholar]
- Hirano, K.; Okuno, A.; Hobo, T.; Ordonio, R.; Shinozaki, Y.; Asano, K.; Kitano, H.; Matsuoka, M. Utilization of stiff culm trait of rice smos1 mutant for increased lodging resistance. PLoS ONE 2014, 9, e96009. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, S.; Gui, J.; Fu, C.; Yu, H.; Song, D.; Shen, J.; Qin, P.; Liu, X.; Han, B.; et al. Shortened Basal Internodes Encodes a Gibberellin 2-Oxidase and Contributes to Lodging Resistance in Rice. Mol. Plant 2018, 11, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Gaur, V.S.; Channappa, G.; Chakraborti, M.; Sharma, T.R.; Mondal, T.K. ‘Green revolution’ dwarf gene sd1 of rice has gigantic impact. Brief. Funct. Genom. 2020, 19, 390–409. [Google Scholar] [CrossRef]
- Jia, Q.J.; Zhang, X.Q.; Westcott, S.; Broughton, S.; Cakir, M.; Yang, J.; Lance, R.; Li, C. Expression level of a gibberellin 20-oxidase gene is associated with multiple agronomic and quality traits in barley. Theor. Appl. Genet. 2011, 122, 1451–1460. [Google Scholar] [CrossRef]
- Jia, Q.J.; Zhang, J.J.; Westcott, S.; Zhang, X.Q.; Bellgard, M.; Lance, R.; Li, C.D. GA-20 oxidase as a candidate for the semidwarf gene sdw1/denso in barley. Funct. Integr. Genom. 2009, 9, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, Z.; Shamsuddin, A.; Rahman, L.; Ara, R. Genotypic and phenotypic correlation and path analysis for lodging resistance traits in bread wheat (Triticum aestivum L.). J. Plant Breed. Genet. 2007, 20, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Sameri, M.; Nakamura, S.; Nair, S.; Takeda, K.; Komatsuda, T. A quantitative trait locus for reduced culm internode length in barley segregates as a Mendelian gene. Theor. Appl. Genet. 2009, 118, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Huang, Y.; Xu, H.; Zhao, M.; Xu, Q.; Li, F. Genetic enhancement of lodging resistance in rice due to the key cell wall polymer lignin, which affects stem characteristics. Breed. Sci. 2018, 68, 5. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Mace, E.; Tao, Y.; Cruickshank, A.; Hunt, C.; Hammer, G.; Jordan, D. Large-scale genome-wide association study reveals that drought-induced lodging in grain sorghum is associated with plant height and traits linked to carbon remobilisation. Theor. Appl. Genet. 2020, 133, 3201–3215. [Google Scholar] [CrossRef]
- Ookawa, T.; Inoue, K.; Matsuoka, M.; Ebitani, T.; Takarada, T.; Yamamoto, T.; Ueda, T.; Yokoyama, T.; Sugiyama, C.; Nakaba, S. Increased lodging resistance in long-culm, low-lignin gh2 rice for improved feed and bioenergy production. Sci. Rep. 2014, 4, 6567. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Liu, W.; Zhou, F.; Peng, Y.; Wang, N. Mechanical properties of maize fibre bundles and their contribution to lodging resistance. Biosyst. Eng. 2016, 151, 298–307. [Google Scholar] [CrossRef]
- Wang, T.J.; Zhang, L.; Han, Q.; Zheng, F.X.; Wang, T.Q.; Feng, N.N.; Wang, T.X. Effect of cell wall and tissue construction on compressive strength of maize stalks. J. Plant Sci. 2015, 33, 109–115. [Google Scholar]
- Muhammad, A.; Hao, H.; Xue, Y.; Alam, A.; Bai, S.; Hu, W.; Sajid, M.; Hu, Z.; Samad, R.A.; Li, Z. Survey of wheat straw stem characteristics for enhanced resistance to lodging. Cellulose 2020, 27, 2469–2484. [Google Scholar] [CrossRef]
- Li, J.B.; Qi, H.X. Progress of genetic research on rice resistance to overturning. Hubei Agric. Sci. 2017, 56, 4450–4453. [Google Scholar]
- Nan, M. Physiological mechanism of oat resistance to downfall and expression of stem lignin synthesis gene. J. Gansu Agric. Univ. 2021. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Hu, C.H. Anatomical studies on stalk resistance of corn. Crop J. 1991, 17, 70–75. [Google Scholar]
- Yang, Y.H.; Zhu, Z.; Zhang, Y.D.; Chen, T.; Zhao, Q.Y.; Zhou, L.H.; Yao, S.; Zhang, Y.H.; Dong, S.L.; Wang, C.L. Relationship between different rice varieties (lines) and stalk morphological traits in terms of resistance to overturning. J. Jiangsu Agric. Sci. 2011, 27, 231–235. [Google Scholar]
- Hu, D.; Liu, X.; She, H.; Gao, Z.; Ruan, R.; Wu, D.; Yi, Z. The lignin synthesis related genes and lodging resistance of Fagopyrum esculentum. Biol. Plant 2017, 61, 138–146. [Google Scholar] [CrossRef]
- Ahmad, I.; Meng, X.P.; Kamran, M.; Ali, S.; Ahmad, S.; Liu, T.N.; Cai, T.; Han, Q.F. Effects of uniconazole with or without micronutrient on the lignin biosynthesis, lodging resistance, and winter wheat production in semiarid regions. J. Integr. Agric. 2020, 19, 62–77. [Google Scholar] [CrossRef]
- Wang, D.; Liu, Y.; Peng, X.L.; Liu, Z.L.; Song, W.B. Effect of optimal fertilizer and water management on the performance of cold rice against overturning. J. Nucl. Agric. 2012, 26, 352–357. [Google Scholar]
- Sun, Q.; Liu, X.G.; Yang, J.; Liu, W.W.; Du, Q.G.; Wang, H.Q.; Fu, C.X.; Li, W.X. MicroRNA528 affects lodging resistance of maize by regulating lignin biosynthesis under nitrogen-luxury conditions. Mol. Plant 2018, 11, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Liu, L.; Zhang, L.; Song, S.; Wang, J.; Wu, L.; Li, H. Characterization of culm morphology, anatomy and chemical composition of foxtail millet cultivars differing in lodging resistance. J. Agric. Sci. 2015, 153, 1437–1448. [Google Scholar] [CrossRef]
- Liu, X.; Gu, W.; Li, C. Effects of nitrogen fertilizer and chemical regulation on spring maize lodging characteristics, grain filling and yield formation under high planting density in Heilongjiang Province, China. J. Integr. Agric. 2021, 20, 511–526. [Google Scholar] [CrossRef]
- Rao, Y.C.; Li, Y.; Dong, G.J.; Zeng, D.L.; Qian, Q. Research progress on rice resistance to overwhelm. China Rice 2009, 6, 15–19. [Google Scholar] [CrossRef]
- Burgert, I. Exploring the micromechanical design of plant cell walls. Am. J. Bot. 2006, 93, 1391–1401. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Luan, Y.T.; Xia, X.; Shi, W.B.; Tang, Y.H.; Tao, J. Lignin provides mechanical support to herbaceous peony (Paeonia lactiflora Pall.) stems. Hortic. Res. 2020, 7, 213. [Google Scholar] [CrossRef]
- Ma, Q.H. Functional analysis of a cinnamyl alcohol dehydrogenase involved in lignin biosynthesis in wheat. J. Exp. Bot. 2010, 61, 2735–2744. [Google Scholar] [CrossRef] [Green Version]
- Bonawitz, N.D.; Chapple, C. The genetics of lignin biosynthesis: Connecting genotype to phenotype. Annu. Rev. Genet. 2010, 44, 337–363. [Google Scholar] [CrossRef]
- Wang, K.; Zhao, S.H.; Yao, X.H.; Yao, Y.H.; Bai, E.X.; Wu, K.L. Stem characteristics and lignin synthesis in relation to Barley’s resistance to lodging. J. Crop Sci. 2019, 45, 621–627. [Google Scholar]
- Liu, X.Y.; Jin, J.Y.; He, P.; Gao, W.; Li, W.J. Effect of potassium chloride on lignin metabolism in maize and its relationship with stem rot resistance. China Agric. Sci. 2007, 40, 2780–2787. [Google Scholar] [CrossRef]
- He, Y.; Mouthier, T.M.; Kabel, M.A.; Dijkstra, J.; Hendriks, W.H.; Struik, P.C.; Cone, J.W. Lignin composition is more important than content for maize stem cell wall degradation. Sci. Food Agric. 2018, 98, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Chen, X.; Yin, Y.; Lu, K.; Yang, W.; Tang, Y.; Wang, Z. Lodging resistance of winter wheat (Triticum aestivum L.): Lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid. Field Crops Res. 2014, 157, 1–7. [Google Scholar] [CrossRef]
- Liu, S.; Tang, Y.; Ruan, N.; Dang, Z.; Huang, Y.; Miao, W.; Xu, Z.; Li, F. The rice BZ1 locus is required for glycosylation of arabinogalactan proteins and galactolipid and plays a role in both mechanical strength and leaf color. Rice 2020, 13, 41. [Google Scholar] [CrossRef]
- Wang, X.; Shi, Z.; Zhang, R.; Sun, X.; Wang, J.; Wang, S.; Zhang, Y.; Zhao, Y.; Su, A.; Li, C. Stalk architecture, cell wall composition, and QTL underlying high stalk flexibility for improved lodging resistance in maize. BMC Plant Biol. 2020, 20, 515. [Google Scholar] [CrossRef]
- Jiang, J.; Liao, X.; Jin, X.; Tan, L.; Lu, Q.; Yuan, C.; Xue, Y.; Yin, N.; Lin, N.; Chai, Y. MYB43 in oilseed rape (Brassica napus) positively regulates vascular lignification, plant morphology and yield potential but negatively affects resistance to Sclerotinia sclerotiorum. Genes 2020, 11, 581. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Raza, A.; Wang, L.; Xu, M.; Lu, J.; Gao, Y.; Qin, S.; Zhang, Y.; Ahmad, I.; Zhou, T. Effects of multiple planting densities on lignin metabolism and lodging resistance of the strip intercropped soybean stem. Agron. J. 2020, 10, 1177. [Google Scholar] [CrossRef]
- Pauly, M.; Gille, S.; Liu, L.; Mansoori, N.; de Souza, A.; Schultink, A.; Xiong, G. Hemicellulose biosynthesis. Planta 2013, 238, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.L.; Wang, Z.; Yang, W.Y. Seedling shading affects morphogenesis and substance accumulation of stem in soybean. Soybean Sci. 2008, 26, 868. [Google Scholar]
- Samadi, A.F.; Suzuki, H.; Ueda, T.; Yamamoto, T.; Adachi, S.; Ookawa, T. Identification of quantitative trait loci for breaking and bending types lodging resistance in rice, using recombinant inbred lines derived from Koshihikari and a strong culm variety, Leaf Star. Plant Growth Regul. 2019, 89, 83–98. [Google Scholar] [CrossRef]
- Zhao, X.H.; Bai, Y.X.; Wang, K.; Yao, Y.H.; Yao, X.H.; Wu, K.L. Effects of planting density on lodging resistance and straw forage characteristics in two hulless barley varieties. Acta Agron. Sin. 2019, 46, 585–586. [Google Scholar] [CrossRef]
- Wei, F.Z.; Li, J.C.; Wang, C.Y.; Qu, H.J.; Shen, X.S. Effect of nitrogen fertilizer transport pattern on wheat stalk lodging resistance. J. Crop Sci. 2008, 34, 1080–1085. [Google Scholar]
- Khan, A.; Ahmad, A.; Ali, W.; Hussain, S.; Ajayo, B.S.; Raza, M.A.; Kamran, M.; Te, X.; al Amin, N.; Ali, S. Optimization of plant density and nitrogen regimes to mitigate lodging risk in wheat. Agron. J. 2020, 112, 2535–2551. [Google Scholar] [CrossRef]
- Wei, F.Z.; Li, J.C.; Qu, H.J.; Shen, X.S. Effect of nitrogen application pattern on lodging frost damage and stalk resistance performance of winter wheat. J. Jiangsu Agric. Sci. 2010, 26, 696–699. [Google Scholar]
- Du, Q.; Avci, U.; Li, S.; Gallego-Giraldo, L.; Pattathil, S.; Qi, L.; Hahn, M.G.; Wang, H. Activation of miR165b represses AtHB15 expression and induces pith secondary wall development in Arabidopsis. Plant J. 2015, 83, 388–400. [Google Scholar] [CrossRef]
- Hossain, Z.; Pillai, B.V.; Gruber, M.Y.; Yu, M.; Amyot, L.; Hannoufa, A. Transcriptome profiling of Brassica napus stem sections in relation to differences in lignin content. BMC Genom. 2018, 19, 255. [Google Scholar] [CrossRef] [Green Version]
- McCahill, I.W.; Hazen, S.P. Regulation of cell wall thickening by a medley of mechanisms. Trends Plant Sci. 2019, 24, 853–866. [Google Scholar] [CrossRef]
- Xiao, R.; Zhang, C.; Guo, X.; Li, H.; Lu, H. MYB transcription factors and its regulation in secondary cell wall formation and lignin biosynthesis during xylem development. Int. J. Mol. Sci. 2021, 22, 3560. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, C.; Zhang, J.; Liu, Y.; Liu, X.; Guo, X.; Li, H.; Liu, D. Integrated transcriptomic and proteomic analysis the roadmap of the xylem development stage in Populus tomentosa. Front. Plant Sci. 2021, 12, 724559. [Google Scholar] [CrossRef]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Z.; Dixon, R.A. On–off switches for secondary cell wall biosynthesis. Mol. Plant 2012, 5, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yu, H.; Rao, X.; Li, L.; Dixon, R.A. Abscisic acid regulates secondary cell-wall formation and lignin deposition in Arabidopsis thaliana through phosphorylation of NST1. Proc. Natl. Acad. Sci. USA 2021, 118, e2010911118. [Google Scholar] [CrossRef]
- Zhong, R.Q.; Kandasamy, M.K.; Ye, Z.H. XND1 regulates secondary wall deposition in xylem vessels through the inhibition of VND functions. Plant Cell Physiol. 2021, 62, 53–65. [Google Scholar] [CrossRef]
- Zhong, R.Q.; Lee, C.H.; McCarthy, R.L.; Reeves, C.K.; Jones, E.G.; Ye, Z.H. Transcriptional activation of secondary wall biosynthesis by rice and maize NAC and MYB transcription factors. Plant Cell Physiol. 2011, 52, 1856–1871. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.P.; Jeong, H.Y.; Jeon, S.H.; Kim, D.; Lee, C. Rice pectin methylesterase inhibitor28 (OsPMEI28) encodes a functional PMEI and its overexpression results in a dwarf phenotype through increased pectin methylesterification levels. J. Plant Physiol. 2017, 208, 17–25. [Google Scholar] [CrossRef]
- Zhong, R.Q.; Lee, C.H.; Zhou, J.L.; McCarthy, R.L.; Ye, Z.H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 2008, 20, 2763–2782. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.Z.; Fu, X.K.; Liu, R.; Guo, L.; Ran, L.Y.; Li, C.F.; Tian, Q.Y.; Jiao, B.; Wang, B.J.; Luo, K.M. PtoMYB170 positively regulates lignin deposition during wood formation in poplar and confers drought tolerance in transgenic Arabidopsis. Tree Physiol. 2017, 37, 1713–1726. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, J.M.; Chapple, C. Rewriting the lignin roadmap. Curr. Opin. Plant Biol. 2002, 5, 224–229. [Google Scholar] [CrossRef]
- Nakahama, K.; Urata, N.; Shinya, T.; Hayashi, K.; Nanto, K.; Rosa, A.C.; Kawaoka, A. RNA-seq analysis of lignocellulose-related genes in hybrid Eucalyptus with contrasting wood basic density. BMC Plant Biol. 2018, 18, 156. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.A.; Cass, C.L.; Petrik, D.L.; Padmakshan, D.; Ralph, J.; Sedbrook, J.C.; Karlen, S.D. Stacking AsFMT overexpression with BdPMT loss of function enhances monolignol ferulate production in Brachypodium distachyon. Plant Biotechnol. J. 2021, 19, 1878. [Google Scholar] [CrossRef]
- Ma, Q.H. The expression of caffeic acid 3-O-methyltransferase in two wheat genotypes differing in lodging resistance. J. Exp. Bot. 2009, 60, 2763–2771. [Google Scholar] [CrossRef] [Green Version]
- Gui, J.; Shen, J.; Li, L. Functional characterization of evolutionarily divergent 4-coumarate: Coenzyme A ligases in rice. Plant Physiol. 2011, 157, 574–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhu, X.; Wang, K.; Lu, C.; Luo, M.; Shan, T.; Zhang, Z. A wheat caffeic acid 3-O-methyltransferase TaCOMT-3D positively contributes to both resistance to sharp eyespot disease and stem mechanical strength. Sci. Rep. 2018, 8, 6543. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chang, J.; Wang, S.; Lu, J.; Liu, Y.; Si, H.; Sun, G.; Ma, C. Cloning, expression analysis and molecular marker development of cinnamyl alcohol dehydrogenase gene in common wheat. Protoplasma 2021, 258, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, Q.; Zhou, Y.; Yan, M.; Sun, L.; Zhang, M.; Fu, Z.; Wang, Y.; Han, B.; Pang, X. BRITTLE CULM1, which encodes a COBRA-like protein, affects the mechanical properties of rice plants. Plant Cell 2003, 15, 2020–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, G.Y.; Li, R.; Qian, Q.; Song, X.Q.; Liu, X.L.; Yu, Y.C.; Zeng, D.L.; Wan, J.M.; Li, J.Y.; Zhou, Y.H. The rice dynamin-related protein DRP2B mediates membrane trafficking, and thereby plays a critical role in secondary cell wall cellulose biosynthesis. Plant J. 2010, 64, 56–70. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Li, S.B.; Qian, Q.; Zeng, D.L.; Zhang, M.; Guo, L.B.; Liu, X.L.; Zhang, B.C.; Deng, L.W.; Liu, X.F. BC10, a DUF266-containing and Golgi-located type II membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.). Plant J. 2009, 57, 446–462. [Google Scholar] [CrossRef]
- Song, X.Q.; Zhang, B.C.; Zhou, Y.H. Golgi-localized UDP-glucose transporter is required for cell wall integrity in rice. Plant Signal. Behav. 2011, 6, 1097–1100. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, Y.; Tan, W.; Chen, J.; Zhu, M.; Lv, Y.; Liu, Y.; Yu, S.; Zhang, W.; Cai, H. Brittle Culm 1 encodes a COBRA-like protein involved in secondary cell wall cellulose biosynthesis in sorghum. Plant Cell Physiol. 2019, 60, 788–801. [Google Scholar] [CrossRef]
- Sato, K.; Suzuki, R.; Nishikubo, N.; Takenouchi, S.; Ito, S.; Nakano, Y.; Nakaba, S.; Sano, Y.; Funada, R.; Kajita, S. Isolation of a novel cell wall architecture mutant of rice with defective Arabidopsis COBL4 ortholog BC1 required for regulated deposition of secondary cell wall components. Planta 2010, 232, 257–270. [Google Scholar] [CrossRef]
- Sindhu, A.; Langewisch, T.; Olek, A.; Multani, D.S.; McCann, M.C.; Vermerris, W.; Carpita, N.C.; Johal, G. Maize Brittle stalk2 encodes a COBRA-like protein expressed in early organ development but required for tissue flexibility at maturity. Plant Physiol. 2007, 145, 1444–1459. [Google Scholar] [CrossRef] [Green Version]
- Persson, S.; Wei, H.; Milne, J.; Page, G.P.; Somerville, C.R. Identification of genes required for cellulose synthesis by regression analysis of public microarray data sets. Proc. Natl. Acad. Sci. USA 2005, 102, 8633–8638. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.M.; Zeef, L.A.; Ellis, J.; Goodacre, R.; Turner, S.R. Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. Plant Cell 2005, 17, 2281–2295. [Google Scholar] [CrossRef]
- Tanaka, K.; Murata, K.; Yamazaki, M.; Onosato, K.; Miyao, A.; Hirochika, H. Three distinct rice cellulose synthase catalytic subunit genes required for cellulose synthesis in the secondary wall. Plant Physiol. 2003, 133, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Atanassov, I.I.; Pittman, J.K.; Turner, S.R. Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J. Biol. Chem. 2009, 284, 3833–3841. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.C.; Ko, J.H.; Kim, J.Y.; Kim, J.; Bae, H.J.; Han, K.H. MYB 46 directly regulates the gene expression of secondary wall-associated cellulose synthases in Arabidopsis. Plant J. 2013, 73, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Atanassov, I.; Turner, S. Functional analysis of cellulose synthase (CESA) protein class specificity. Plant Physiol. 2017, 173, 970–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Ding, S.Y. The N-terminal zinc finger of CELLULOSE SYNTHASE6 is critical in defining its functional properties by determining the level of homodimerization in Arabidopsis. Plant J. 2020, 103, 1826–1838. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Li, Y.; Hu, Z.; Hu, H.; Wang, G.; Li, A.; Wang, Y.; Tu, Y.; Xia, T.; Peng, L. Ectopic expression of a novel OsExtensin-like gene consistently enhances plant lodging resistance by regulating cell elongation and cell wall thickening in rice. Plant Biotechnol. J. 2018, 16, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Liu, S.; Xu, H.; Xu, Q. A novel FC17/CESA4 mutation causes increased biomass saccharification and lodging resistance by remodeling cell wall in rice. Biotechnol. Biofuels 2018, 11, 298. [Google Scholar] [CrossRef]
- Li, F.; Zhang, M.; Guo, K.; Hu, Z.; Zhang, R.; Feng, Y.; Yi, X.; Zou, W.; Wang, L.; Wu, C. High-level hemicellulosic arabinose predominately affects lignocellulose crystallinity for genetically enhancing both plant lodging resistance and biomass enzymatic digestibility in rice mutants. Plant Biotechnol. J. 2015, 13, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Wu, P.; Zhu, M.; Qian, H.; Cao, W.; Hu, Y.; Guo, B.; Wei, H.; Xu, K.; Dai, Q. Effect of mechanized planting methods on plant type and lodging resistance of different rice varieties. Trans. Chin. Soc. Agric. Eng. 2017, 33, 52–62. [Google Scholar]
- Berry, P.; Sylvester-Bradley, R.; Berry, S. Ideotype design for lodging-resistant wheat. Euphytica 2007, 154, 165–179. [Google Scholar] [CrossRef]
- Alam, M.; Sultana, M.; Hossain, M.; Salahin, M.; Roy, U. Effect of sowing depth on the yield of spring wheat. J. Environ. Sci. Nat. Res. 2014, 7, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Song, F.; Liu, F.; Zhu, X.; Xu, H. Effect of planting density on root lodging resistance and its relationship to nodal root growth characteristics in maize (Zea mays L.). J. Agric. Sci. 2012, 4, 182. [Google Scholar] [CrossRef] [Green Version]
- Easson, D.; White, E.; Pickles, S. The effects of weather, seed rate and cultivar on lodging and yield in winter wheat. J. Agric. Sci. 1993, 121, 145–156. [Google Scholar] [CrossRef]
- Zhu, C.; Ziska, L.H.; Sakai, H.; Zhu, J.; Hasegawa, T. Vulnerability of lodging risk to elevated CO2 and increased soil temperature differs between rice cultivars. Eur. J. Agron. 2013, 46, 20–24. [Google Scholar] [CrossRef]
- Zhu, C.; Cheng, W.; Sakai, H.; Oikawa, S.; Laza, R.C.; Usui, Y.; Hasegawa, T. Effects of elevated (CO2) on stem and root lodging among rice cultivars. Chin. Sci. Bull. 2013, 58, 1787–1794. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhou, N.; Lai, S.; Frei, M.; Wang, Y.; Yang, L. Elevated CO2 improves lodging resistance of rice by changing physicochemical properties of the basal internodes. Sci. Total Environ. 2019, 647, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, Y.; Ogata, T.; Sato, H.; Matsue, Y. Effects of silicate application on lodging, yield and palatability of rice grown by direct sowing culture. Jpn. J. Crop Sci. 2000, 69, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Gong, D.; Zhang, X.; Yao, J.; Dai, G.; Yu, G.; Zhu, Q.; Gao, Q.; Zheng, W. Synergistic effects of bast fiber seedling film and nano-silicon fertilizer to increase the lodging resistance and yield of rice. Sci. Rep. 2021, 11, 12788. [Google Scholar] [CrossRef]
- Dorairaj, D.; Ismail, M.R.; Sinniah, U.R.; Tan, K.B. Silicon mediated improvement in agronomic traits, physiological parameters and fiber content in Oryza sativa. Acta Physiol. Plant 2020, 42, 38. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Shuxian, L.; Mumtaz, M.; Shafiq, I.; Iqbal, N.; Brestic, M.; Shoaib, M.; Sisi, Q.; Li, W.; Mei, X.; et al. Foliar application of silicon improves stem strength under low light stress by regulating lignin biosynthesis genes in soybean (Glycine max (L.) Merr.). J. Hazard. Mater. 2021, 401, 123256. [Google Scholar] [CrossRef]
- Xu, Z.; Lai, T.; Li, S.; Si, D.; Zhang, C.; Cui, Z.; Chen, X. Promoting potassium allocation to stalk enhances stalk bending resistance of maize (Zea mays L.). Field Crops Res. 2018, 215, 200–206. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, H.; Yi, Y.; Ding, J.; Zhu, M.; Li, C.; Guo, W.; Feng, C.; Zhu, X. Effect of nitrogen levels and nitrogen ratios on lodging resistance and yield potential of winter wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0187543. [Google Scholar] [CrossRef]
- Chen, X.; Wang, J.; Wang, Z.; Li, W.; Wang, C.; Yan, S.; Li, H.; Zhang, A.; Tang, Z.; Wei, M. Optimized nitrogen fertilizer application mode increased culms lignin accumulation and lodging resistance in culms of winter wheat. Field Crops Res. 2018, 228, 31–38. [Google Scholar] [CrossRef]
- Wang, C.; Wu Ruan, R.; Hui Yuan, X.; Hu, D.; Yang, H.; Li, Y.; Lin Yi, Z. Effects of nitrogen fertilizer and planting density on the lignin synthesis in the culm in relation to lodging resistance of buckwheat. Plant Prod. Sci. 2015, 18, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Bitarafan, Z.; Rasmussen, J.; Westergaard, J.C.; Andreasen, C. Seed Yield and Lodging Assessment in Red Fescue (Festuca rubra L.) Sprayed with Trinexapac-Ethyl. Agron. J. 2019, 9, 617. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Chen, T.; Zhao, C.; Zhou, M. Improving Crop Lodging Resistance by Adjusting Plant Height and Stem Strength. Agron. J. 2021, 11, 2421. [Google Scholar] [CrossRef]
- Rolston, P.; Trethewey, J.; Chynoweth, R.; McCloy, B. Trinexapac-ethyl delays lodging and increases seed yield in perennial ryegrass seed crops. N. Z. J. Agric. Res. 2010, 53, 403–406. [Google Scholar] [CrossRef]
- Asahina, M.; Iwai, H.; Kikuchi, A.; Yamaguchi, S.; Kamiya, Y.; Kamada, H.; Satoh, S. Gibberellin produced in the cotyledon is required for cell division during tissue reunion in the cortex of cut cucumber and tomato hypocotyls. Plant Physiol. 2002, 129, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.; Fedak, G. Effects of chlormequat on plant height, disease development and chemical constituents of cultivars of barley, oats, and wheat. Can. J. Plant Sci. 1977, 57, 31–36. [Google Scholar] [CrossRef]
- Puran, B.; Ronell, S.B. Evaluation of anti-lodging plant growth regulators on the growth and development of rice (Oryza sativa). J. Cereals Oilseeds 2014, 5, 12–16. [Google Scholar]
- Shah, A.N.; Tanveer, M.; Anjum, S.A.; Iqbal, J.; Ahmad, R. Lodging stress in cereal—Effects and management: An overview. Environ. Sci. Pollut. Res. 2017, 24, 5222–5237. [Google Scholar] [CrossRef]
- Han, L.P.; Wang, X.; Guo, X.; Rao, M.S.; Steinberger, Y.; Cheng, X.; Xie, G.H. Effects of plant growth regulators on growth, yield and lodging of sweet sorghum. Res. Crops 2011, 12, 372–382. [Google Scholar]
- Zhao, D.; Luan, Y.; Shi, W.; Tang, Y.; Huang, X.; Tao, J. Melatonin enhances stem strength by increasing the lignin content and secondary cell wall thickness in herbaceous peony. J. Exp. Bot. 2022, erac165. [Google Scholar] [CrossRef]
- Arai-Sanoh, Y.; Ida, M.; Zhao, R.; Yoshinaga, S.; Takai, T.; Ishimaru, T.; Maeda, H.; Nishitani, K.; Terashima, Y.; Gau, M. Genotypic variations in non-structural carbohydrate and cell-wall components of the stem in rice, sorghum, and sugar vane. Biosci. Biotechnol. Biochem. 2011, 75, 1104. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Fu, C.; Liang, C.; Ni, X.; Zhao, X.; Chen, M.; Ou, L. Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy 2022, 12, 1795. https://doi.org/10.3390/agronomy12081795
Li Q, Fu C, Liang C, Ni X, Zhao X, Chen M, Ou L. Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy. 2022; 12(8):1795. https://doi.org/10.3390/agronomy12081795
Chicago/Turabian StyleLi, Qing, Canfang Fu, Chengliang Liang, Xiangjiang Ni, Xuanhua Zhao, Meng Chen, and Lijun Ou. 2022. "Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance" Agronomy 12, no. 8: 1795. https://doi.org/10.3390/agronomy12081795
APA StyleLi, Q., Fu, C., Liang, C., Ni, X., Zhao, X., Chen, M., & Ou, L. (2022). Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy, 12(8), 1795. https://doi.org/10.3390/agronomy12081795