Abstract
Globally, grasslands have been heavily degraded, more so than any other biome. Grasslands of the eastern U.S. are no exception to this trend and, consequently, native biota associated with the region’s >20 million ha of agricultural grasslands are under considerable stress. For example, grassland associated breeding bird populations have declined precipitously in recent decades as have numerous species of pollinators. Although there is increasing awareness of the role grasslands can play in global carbon cycles and in providing high quality dietary proteins needed by an increasing global population, there is a lack of awareness of the alarming trends in the sustainability of the native biota of these ecosystems. Here, we present the status of this conservation challenge and offer prospective solutions through a working lands conservation approach. Such a strategy entails maintaining appropriate disturbances (i.e., grazing, fire, and their combination), improved grazing management, an increased reliance on native grasses and forbs, and improved plant diversity within pastures. Furthermore, we note some examples of opportunities to achieve these goals, offer suggestions for agricultural and conservation policy, and provide a framework for evaluating tradeoffs that are inevitably required when pursuing a multi-purpose grassland management framework.
1. Introduction
Increased demands for sustainability in beef and other ruminant production systems have placed greater emphasis on ecosystem services provided by grasslands agriculture. For instance, the U.S. Roundtable for Sustainable Beef (USRSB), an organization formed in 2015, developed their sustainability framework for the U.S. beef industry in 2019 [1] and more recently released indicators to document sustainability throughout the beef value chain [2]. Similarly, The Nature Conservancy (TNC) developed a strategy for sustainable beef production in 2021 [3]. Sustainability concerns relative to grasslands within these production systems include impacts on climate [4,5], soil health [6,7], ecosystem integrity [8,9,10], water conservation [11,12], and animal health [13].
Another critical issue for the sustainability of grasslands, and the agriculture systems that depend on those grasslands, is biodiversity. Although biodiversity has been the focus of some research within semi-arid grasslands of the U.S. [14,15,16], far less attention has been paid to grasslands of the humid eastern U.S. [17,18,19]. This is especially true as it relates to biodiversity associated with grasslands agriculture (i.e., working lands) within this region [20,21]. Given the scale of grasslands agriculture in this region (>20 million ha in pasture and hay production comprising >40% of all U.S. cow-calf operations [22]), addressing biodiversity within working grasslands should be an important priority [20].
Biodiversity is essential to sustainable grassland ecosystems and is, in and of itself, a clear imperative for land stewardship for all types of agricultural production systems. The long-term decline of grassland-associated bird populations [23] and the more recent reductions in insect communities [24], especially pollinators [25,26], provide two compelling examples of at-risk taxa that are critical components of healthy, resilient grassland ecosystems. Additionally, healthy grasslands require microbes that play critical roles in nutrient cycling and carbon sequestration. The types of microbial communities and their function are greatly affected by grassland management, particularly plant diversity, leading to corresponding, rapid changes in the microbial community [27,28].
In this paper, our objectives are to raise awareness of these issues and draw attention to the need to improve our understanding of how grasslands management practices impact key elements of biodiversity as the first steps in addressing these challenges. In addition, we note that practical management options that can reverse or, at a minimum, slow down these precipitous declines must be identified [29,30]. We assert that solutions based on a working lands conservation model—an approach that recognizes the importance of maintaining agricultural production and simultaneously enhancing biodiversity—are preferable to those focused simply on setting aside land for conservation purposes [17,20,31]. Indeed, the increasing global demand for high quality dietary proteins dictates that grassland productivity be maintained in the future. Furthermore, working lands approaches to these pressing conservation challenges can demonstrate to concerned consumers that production agriculture can be sustainable. Such an approach can go a long way towards showcasing the ability of agriculture to achieve societal demands for healthy ecosystems and the services that they can deliver; grassland agriculture can be the solution rather than the problem.
2. Critical Concerns for Grassland Biodiversity
2.1. The Plight of Grassland Birds
Avifauna serve as excellent environmental indicator for ecosystem health [32] and represent a substantial component of vertebrate diversity in grassland ecosystems. As such, grassland bird population trends and distributions are very sensitive to the extreme loss and fragmentation of native grasslands and to the degradation in grassland quality in North America, all of which have contributed to these declines. A recent evaluation of avian population trends among all breeding bird guilds in North America indicated that those species associated with grasslands have experienced by far the most severe declines (53% decline, on average, over the past 50 years [23]). Some species, notably northern bobwhite (Colinus virginianus; 79% decline), eastern meadowlark (Sturnella magna; 73% decline), grasshopper sparrow (Ammodramus savannarum; 72% decline), and Bachman’s Sparrow (Peucaea aestivalis; 70% decline), have experienced even greater declines over the past 50 years [33]. At current rates of decline (3.1%/year), the remaining northern bobwhite population may be reduced by 50% over the next decade.
Individual grassland bird species have unique habitat requirements typically associated with the structure of grassland vegetation [34] and with the structure of the landscape in which they are found in [35,36]. Different species occupy grasslands at differing seral stages and may respond differently to the type and extent of the associated disturbance regime [14,21]. Some grassland species are area sensitive, in that their occurrence and/or density is positively associated with grassland patch size [35]. Species which are extremely area sensitive may require grassland patches >100 ha (i.e., Henslow’s Sparrow) or even >1000 ha in extent (i.e., Greater Prairie-Chicken [Tympanuchus cupido], [34]).
2.2. The Plight of Native Pollinators
Declines in the abundance and diversity of insect pollinators have been widely documented in recent years [25,37]. Managed honey bees (Apis mellifera) in particular have received a great deal of attention, with an estimated 59% reduction in the number of colonies in North America alone [38]. Wild native pollinators, including bumble bees, solitary bees, butterflies, and moths, have also experienced substantial population losses [39,40,41]. In 2017, the rusty-patched bumble bee (Bombus affinis) became the first federally listed endangered bee under the U.S. Endangered Species Act (U.S. Fish & Wildlife Service, Federal Register 82 FR 3186). The monarch butterfly (Danaus plexippus) provides another well-known example of a species in decline, with the eastern U.S. migratory population reduced as much as 80% in its overwintering sites in Mexico [42]. Although multiple drivers likely contribute to on-going pollinator declines (e.g., habitat fragmentation, climate change, pesticide use, introduced species, and parasites), habitat loss has the most significant detrimental impact on the abundance and diversity of insect pollinators [25,39,43].
The loss of pollinators threatens wild and cultivated plants worldwide [25,26,44]. More than 80% the world’s flowering plants depend on animal pollination [45]. Of these, outcrossing and rare plants are particularly susceptible to pollinator declines [46,47], and may themselves decline as a result [40]. Crop production is also vulnerable to the loss of insect pollinators. Approximately 75% of crop species benefit from the pollination services of insects, primarily bees [48]. Globally, crop pollination services are valued at more than USD 200 billion per year [49]. In the U.S., wild native bees supply an estimated USD 3 billion annually in pollination services to crops [50].
Beyond pollinators, a global pattern of declining insect diversity and abundance more generally has begun to emerge [51,52] driven by multiple stressors, including land-use change, climate change, agricultural intensification, and introduced species [52]. Loss of insects can be detrimental to overall ecosystem function because of the critical role these organisms play in nutrient cycling, reducing pressure from economically important pests, and as the foundation for sustaining food webs [53]. The contributions of insects to these vital ecosystem processes have significant economic impacts as well. For example, naturally occurring insect predators and parasitoids are estimated to provide USD 4.5 billion annually to U.S. crop production based on the value of crop losses averted [50].
Managing grasslands to enhance plant biodiversity has the potential to benefit insect communities and the valuable ecosystem services they provide [54,55,56]. A growing body of research demonstrates that pollinators respond positively to habitat heterogeneity and floral resources [39]. Native wildflowers support insect pollinator diversity and abundance by providing nectar and pollen resources and nesting or reproduction sites [57,58]. Likewise, agricultural landscapes with greater plant diversity sustain larger populations of predators and parasitoids [59,60], which contribute to the natural control of economic pests [61]. Thus, diverse healthy grasslands can create multi-functional environments that generate positive insect conservation and economic outcomes.
2.3. Soil Microbes
Diverse, healthy grasslands have vibrant microbiological communities that can be influenced by management practices. Although the relationship between grassland microbes, and other elements of biodiversity (e.g., grassland birds), is mostly indirect, the abundance and productivity of all these species likely reflects the nature of the grassland [62,63,64]. That is, in grasslands, the same features, habitat, structure, and food resources that diverse plant communities provide for birds have corresponding effects and benefits for soil microbial communities [65,66,67]. These responses are also associated with additional ecosystem services that can create feedback among system components. For example, soil water infiltration capacity is greater in systems with biodiverse groundcover [12,62,68], and reduced nutrient leaching losses also are observed with greater grassland diversity [28], a process mediated in part by the degree of legume presence [63].
Plant diversity can increase microbial community diversity (e.g., [28,69]). Not surprisingly, this may be correlated with presence of soil fungi, arbuscular mycorrhizae, and possibly with greater carbon storage at depth [65]. Moreover, from an agricultural management standpoint, lower-input systems foster greater soil faunal diversity [66]. At the very least, these findings reinforce a point of our thesis, that protecting and increasing use of native grasses—obligate mycotrophs—in forage livestock system can provide a suite of ecosystem services important not only for grassland birds but also for other organisms, including human beings.
3. Solutions for Grassland Biodiversity Conservation
3.1. Appropriate Disturbance
Persistence of grasslands is dependent upon climate, fire, and grazing [62,70]. Disturbances, such as fire and grazing, are critical, but the importance to conservation is dependent upon the intensity, frequency, and pattern of these disturbances directly; more important, though, is the interaction among these disturbances that leads to complex patterns [14,15,71]. Fire and grazing can have positive, negative, and neutral effects on ecosystem services depending how they are applied. In remaining, large intact grasslands, grazed areas and ungrazed areas are best suited to different species and can provide different ecosystem services [70]. Because of the tradeoffs, it is critical for conservation that landscapes remain large enough to include patterns of disturbance that include highly disturbed and relatively undisturbed areas. Traditional conservation focused on trying to promote uniform and moderate use by livestock while suppressing fire is incapable of maintaining diverse grassland landscape that support our desired ecosystem services [5,8]. The interaction of fire and grazing can result in patterns that can provide diverse ecosystem services, including livestock production, biodiversity, wildfire mitigation, water availability, and even human health [71].
Recently, there has been increased attention on carbon sequestration and storage on grassland landscapes because of their prospective roles under changing climate scenarios. Suggestions of massive tree planting efforts discount the importance of grasslands and promote climate solutions that have limited consideration of other ecosystem services. The focus on increased above-ground carbon from woodlands that were once grasslands is tempting from a carbon perspective. While this is debated and many argue that grasslands may be a more reliable carbon sink, tree planting proposals continue in spite of a long history of failed efforts to convert large regions from grassland to forests (e.g., tree planting in the Great Plains goes back over 100 years) [8,71].
3.2. Working Lands Approach
To address these challenges to sustaining biodiversity, the most practical solution is to pursue a working lands approach. That is, agricultural landscapes do not need to be managed solely for commodity production, but rather can be managed for biodiversity-and multiple other ecosystem services as well. A working lands approach can support both conservation needs and human livelihoods and recognizes humans not only as stewards, but as active participants within an ecosystem.
Much energy has been given to the scientific disputes regarding land sparing vs. land sharing management approaches, and it is not our intent to rehash that debate here. Indeed, both approaches likely have merit in a “both-and” framing of habitat conservation [72]. However, such arguments have merit depending on the conservation needs of the species and ecosystems of interest. For example, Wright et al. [73] describe agriculture as “a key element for conservation”, particularly where threatened species rely on the open habitat that low-impact agriculture can create. Indeed, there are several reasons why a working lands approach is the preferred path to achieving desired production and conservation goals in humid, temperate grasslands such as those of the eastern U.S.
First, it will be most readily acceptable to those who own and manage the extensive grasslands of this region to produce forage for cattle. This represents more than 300,000 farms, not including those that produce other classes of livestock [22]. Beef cattle producers have the lowest average farm income among all the major commodities, with an average annual net farm income from 2012–2020 of USD 32,866 [74]. Indeed, if the historical anomaly for returns to cow-calf operations in 2014 and 2015 are omitted, the average annual return between 2010 and 2020 was only USD 22 per head [75]. Clearly, the ability of individual farms to accept the burden of externalities such as supporting biodiversity enhancement is limited and practical, cost-effective solutions will be essential for achieving meaningful adoption of conservation practices.
Secondly, the scale required to address compromised biodiversity is such that conventional, government-subsidized practices will not be practical [20,36]. The amount of land enrolled in conservation practices under USDA’s Conservation Reserve Program (CRP) represents only 1.16% of the land area in forage production [20] across the eastern U.S., a level well below that required to create population effects for grassland birds [18,36]. Furthermore, there is substantial cost to implement and sustain government conservation programs; USD 518 million was spent in fiscal year 2017 alone on CRP, an amount that does not include administrative costs [76]. By contrast, working lands approaches could operate without the necessity of subsidies, or at least minimize the need for them, and could potentially impact millions of hectares of actively managed grasslands, a level capable of impacting biodiversity [20].
Lastly, working lands approaches that utilize management practices such as grazing and prescribed fire, impose essential—and ecologically appropriate—forms of disturbance [64,77]. Such disturbance is essential to maintaining the biodiversity of grasslands of North America [62,78]. Grazing fosters beneficial vegetative structure [70] and composition [79,80] within grasslands. Grazing has also been linked to improvements in avifauna and other elements of biodiversity [67,81,82]. Similarly, fire has long been recognized as a natural disturbance vector in North American grasslands [83,84,85] that positively influences vegetative communities and associated biodiversity [15,77,86].
Producer buy-in will be critical to these efforts and may not come easily, however. Such changes will require land managers both to re-envision what looks “good” on the landscape and to develop the skills and infrastructure needed to change. For many producers, livestock production is a lifestyle choice rooted in long-held traditions. Acting against perceived history, cultural benchmarks, or community norms can be significant barriers to change.
3.3. Improved Grassland Management
Good grassland management can be summarized simply as grazing to maintain both leaf area and the maintenance of carbohydrates reserve in stem bases, rhizomes/stolons, and roots [87,88]. Leaf area is maintained by leaving adequate residue after grazing and allowing a rest period and adequate vegetative regrowth before the next grazing cycle. Allowing adequate rest periods for regrowth also allows the production of carbohydrates in excess of those required for growth, which can then contribute to short- and long-term storage and be available during periods of stress. Therefore, overgrazing, which fails to promote positive energy balances for the grasses or proper nutrient recycling, leads to weakened stands, reduced biomass production, and increased cover of undesirable species [87,88]. Thus, the net effect is less quantity and quality of feed for livestock.
The priority for all livestock producers is the meat, milk, and wool production that they obtain from their grasslands, both those utilized for pasture and for stored feed. Good grassland management leads to improved livestock production and poor grassland management can lead to reduced livestock production. Although cattle can be raised on farms using poor grassland management practices, supplemental feed is often required and the net income from meat, milk, and wool are low. In short, the economic viability of these operations is compromised [89].
Researchers have long known that overgrazing, defined as close and frequent grazing, reduces both topgrowth and root growth [87]. Grazing below 5 cm will actually stop root growth and successive cycles of grazing decrease root biomass and lead to reduced nutrient uptake and lower drought resiliency [87]. Close and frequent grazing affects all species but has the most dramatic effect on tall and intermediate grasses, such as the tall native species, and on legumes with tap roots. For example, grazing alfalfa (Medicago sativa) without an approximately 30-day rest period will delete root carbohydrate reserves and lead to lower production and weakened plants that are more susceptible to disease [90]. Grazing tall native grasses to low stubble heights and with a short rest period will lead to slow regrowth, reduced tillering, and weed encroachment [88].
One of the most overlooked but important considerations of grassland management is the effect of grazing management on species diversity. Maintaining rich species diversity contributes to improved grassland productivity and resiliency to climatic variability [13,89,91]. With increasing species diversity, however, increased management intensity will be needed with the primary criteria to graze according to the management requirements of the most desirable species (typically the highest quality). For example, in a heterogenous native grass/forb/legume planting containing species such as big bluestem (Andropogon gerardii), indiangrass (Sorghastrum nutans), little bluestem (Schizachyrium scoparium), purple coneflower (Echinacea purpurea), and white prairie clover (Dalea candida), it would be important to graze to favor the coneflower and clover, which would be more sensitive to early season overgrazing than the grass species. An increasingly common introduced species mixture in the lower southeastern U.S. is alfalfa/bermudagrass (Cynodon dactylon), which combines the high quality, yield, N-fixation, and season-long productivity of alfalfa and the strong sod and warm-season growth of bermudagrass [92]. An alfalfa/bermudagrass mixture should be grazed to favor the alfalfa with the first grazing at an early flowering stage of the alfalfa and subsequent grazings after 4 or more weeks to allow the alfalfa adequate time to replenish carbohydrate reserves [90]. This strategy also shades the bermudagrass in the late spring, limiting its aggressiveness in the mixture. Another option to maintain species diversity in the landscape is to graze more homogeneous stands in sequence based on their growth and quality attributes. An example of sequential grazing could be to graze fall rye (Secale cereale) late winter (interseeded into warm-season natives), tall fescue (Schedonorus arundinaceus) early spring (before toxicity increases), switchgrass (Panicum virgatum)/red clover (Trifolium pratense) late spring and early summer, big bluestem/indiangrass mid-to late-summer, and then rely on stockpiled fescue for fall and early winter grazing.
Grassland management can have a major effect on soil surface characteristics. When the majority of forage species are grazed close and frequently, the stand thins and the soil surface is exposed [88]. An exposed soil surface is more prone to wind and water erosion [88,93]. An additional negative consequence is that bare soil in summer months is warmer than shaded soil. Recent research in Kentucky [93] showed that bare soil exposed from frequent and close defoliation was 5–10 °C warmer than adjacent shaded soil.
One of the first topics that most producers consider with grassland management is the presence of broadleaf weeds in their pastures and, if present, how they can be eradicated. However, most forbs can be useful forages or at least their presence can be tolerated because of forage quality attributes. Many forbs rival the quality of the best forage legumes like alfalfa and clover. Examples are common ragweed (Ambrosia artemisiifolia), common lambsquarters (Chenopodium album), dandelion (Taraxacum officinale), and plantain (Plantago spp.) [94,95,96]. These and other forbs are not always palatable and are often lower yielding than common forages, but most livestock will consume them and show impressive gains. Some of these forbs even have medicinal compounds that improve rumen function and reduce parasite load (e.g., chicory [Cichorium intybus]). Therefore, good grassland management should tolerate some level of broadleaf plants.
Maximizing grassland stand health through management not only maximizes forage production and livestock production, but also enhances habitat for wildlife species, pollinators, and multiple other ecosystem benefits. Such management allows for greater plant diversity, leading to more heterogeneous canopy structure, more floral resources for pollinators, and more potential food resources for grassland associated birds. Furthermore, vigorous forage stands can provide greater soil organic matter, greater variability in root architecture, and longer periods with active root growth, all of which are beneficial for soil organisms.
3.4. Restoration of Native Plants Species within Forage Systems
Native grasslands are among the most imperiled ecosystems in North America [97,98]. Virtually all native grasslands in the eastern U.S. have been converted to other uses, such as row crop production, succession to closed-canopy forests, or conversion to introduced grasses, such as tall fescue, bermudagrass, or bahiagrass (Paspalum notatum) [99]. These introduced species negatively affect grassland birds, primarily because their morphology (and thus habitat structure) is quite different from that of native species, such as big bluestem, little bluestem, indiangrass, and switchgrass, that once dominated eastern grasslands [29,99]. These non-natives are typically managed with canopy heights below 25 cm, commonly to 5 or 10 cm, and thus, provide little cover [88,99,100]. Furthermore, they normally form dense sods that reduce vegetative diversity and alter successional trajectories [99,100,101]. By contrast, most native grasses suited for pasture systems are tall-growing species and out of necessity are managed at greater residual canopy heights (e.g., >30 cm) [88,102,103]. They are also bunch grasses that provide interstitial space between plants that foster greater plant diversity and enable easier movement of fledglings and enhanced ability to forage for insects [99,104]. Consequently, outcomes for at-risk avifauna within native grass pastures are more favorable than for introduced species [17,105,106]. There is also evidence that introduced species (e.g., tall fescue, but based on symbiotic endophytes) can have a detrimental impact on key soil biota [107,108].
4. Case Studies
4.1. Natives Complementing Tall Fescue
In recent years, numerous studies of native grasses have documented positive production [102,104,109] and economic [103,110,111,112] outcomes driven by high rates of gain in growing cattle (i.e., 0.8–1.1 kg/day) combined with high stocking rates and low input requirements. Where native grasses are used to complement tall fescue-based forage systems, an additional advantage is avoidance of fescue toxicity issues and associated negative impacts on animal health and productivity [104,113]. Recent research has also demonstrated that these grazed native forages can provide habitat for grassland birds [17,106,114] and substrate for pollinators [115], thus supporting a viable working lands conservation approach [20].
4.2. Patch-Burn Grazing
An alternative grazing system is patch-burn grazing (PBG), where grazing and rest periods are measured in years [15,70,71]. In this system, a “patch” within a pasture, typically one-third of the area for pastures in the eastern U.S., is burned each spring. Livestock are allowed to continuously graze the area once the burned area has initiated growth. In this scenario, livestock are attracted to the fresh growth in the burned area and graze it more frequently and closer that would normally be recommended. The area that was burned a year ago is not grazed as closely because of accumulated growth. Furthermore, the area burned two years previously has much less grazing intensity because of its more rank, mature growth. In essence, little to no rest occurs in year one, partial rest occurs in year two, and almost complete rest occurs in year three. The PBG approach simulates natural disturbance patterns typical of North American grasslands [14]. Consequently, overall heterogeneity is increased providing improved habitat for a variety of grassland wildlife [14,15].
4.3. Weediness and/or More Diverse Plantings
The negative perception of weediness in grasslands is one that must be overcome by the realization that diverse grasslands produce more forage and more seasonally consistent nutritive value and biomass relative to simple monoculture grasslands. This realization can be arrived at through better understanding forage production characteristics of diverse-planted grasslands. A case study developed to demonstrate biomass and nutritive value potential of grasslands planted to native warm-season grasses and interseeded with legumes and forbs in Missouri, suggests there is little, if any, difference in biomass production in grass–forb mixtures compared to grass alone (Table 1, [116]). There are likely differences in nutritive value between monocultures and grass-forb mixtures. Diversifying switchgrass-dominated grasslands with red clover alone may increase pounds of protein per ha on offer by as much as 24% compared to switchgrass monoculture [117]. Grasslands containing a diverse mix of forbs, such as ragweed, plantain, and dandelion, may contribute to greater nutritive value of available forage on offer when harvested at the vegetative stage of maturity [91].

Table 1.
A comparison of yields (Mg/ha) from 2010–2013 among mixtures of big bluestem-switchgrass pastures interseeded with various mixtures of forbs and legumes harvested at 15-cm residual plant height. Table adapted from [115].
5. Policy Solutions
The success of a working lands approach to enhance biodiversity within perennial grassland systems will depend on producer’s adoption of the essential elements of grassland biodiversity--native species, increased plant diversity, and appropriate disturbance regimes. Moreover, adoption of such practices will likely require new policy approaches, particularly those that remove barriers to adoption and/or incentivize such practices. Wright et al. [73] noted several potential policy measures including certification schemes, development opportunities tied to the support of ecotourism, payment for ecosystem services, and payment for conservation practices. Given the scale of need and costs of existing programs, however, many of these incentives may need new funding mechanisms (e.g., private philanthropy) or changes in policies connected to land management (e.g., land transfer taxation) that provide incentives for change. Others have noted that policy schemes (which are typically directed at farm-level outcomes), may have greater effect if farmer management could be coordinated to achieve landscape scale goals [118]. New approaches such as this hold promise, but they will require addressing issues, such as inefficiencies of landscape scale management and the cultural barriers to change. Regardless of the approach, policy measures that support livelihoods in rural communities will be essential for buy-in.
Traditionally, U.S. beef production systems have relied on significant exogenous inputs throughout the supply chain. From forage production in the cow-calf operation to grain grown for finishing cattle in feedlots, the system has been facilitated through the use of relatively cheap fossil fuel and its derivatives. As Smith et al. [119] noted, “[g]rain feeding livestock as presently practiced will be around only as long as there are grain surpluses and cheap energy.”
One of the prime arguments in support of the petroleum-subsidized production model is that finishing beef cattle in confinement with grain-based diets takes fewer ha and less time per unit output than finishing animals on pasture. Although the grain-finishing approach has historically been considered efficient, such accounting can be questioned for not considering all the costs and benefits of the different production models. For example, a recent life-cycle assessment comparing forage finished vs. feedlot finished beef cattle acknowledged that feedlot finishing may have reduced greenhouse gas emissions and beef per ha of pasture, but when one considers the potential value of well-managed pasture as a carbon sink, forage finishing would have less environmental impact per unit output than feedlot-based systems [120]. Those authors noted that perennial grasslands would yield provisioning services in conjunction with a more robust suite of ecosystem services—the key point we argue in this paper. Conversion of a part of a farm operation to native grasses—which require far fewer exogenous inputs and better resist weather extremes—would create more resistant and resilient rural landscapes with lower economic costs. Such systems can provide for critical biodiversity needs and may also prove to be a significant carbon sink. Moreover, if that sunk carbon has value in a marketplace, it may increase opportunities to reward producers for good grassland management.
The energy and effort needed to make systemic changes supporting a scale-up of grasslands supporting biodiversity (particularly those utilizing native species) must not be solely directed to (and expected from) producers and the conservation-minded. Rather, consumers must also be educated about the role of livestock in appropriately managed grazing systems as a means of sinking carbon and creating critical habitat for threatened species. Motivating consumers and creating opportunities for them to make economic “votes” in favor of these systems can be an important part of enlarging the economic pie for farms and rural communities engaged in these practices.
5.1. Encouraging Native Species
Encouraging the use of native grass forages as a component of the forage system can provide benefit to enhanced biodiversity within eastern grasslands and should be explored as a policy solution to address current declines. Concerns about climate change, including increased frequency and severity of droughts and/or floods and increased temperatures, all suggest a role for these well-adapted C4 species in regional forage production systems. In addition, many large processors within the beef value chain have made commitments to reduced greenhouse gas emissions and improved carbon sequestration goals. Because of their deep root systems and low input requirements, native forages can play a role in achieving these goals. Beyond simply educating producers regarding the option of using native grasses, it is also important to provide support for transitioning some portion of existing forage production into these species. The greatest barrier to adoption has been establishment, the cost, time required, and lost forage production during the seedling year and associated risks [121]. Programs in collaboration with USDA or beef industry partners could help mitigate risk and reduce the cost of transitioning to native forages.
5.2. Encouraging More Diverse Plant Communities
The concept of sustainable intensification of agroecosystems combines increased productivity with preservation of as well as positive impact on the environment [122]. One method of incorporating sustainable intensification practices into grasslands is to increase diversity of the plant community to mimic natural ecosystems. Natural ecosystems have greater species diversity, accumulate more biomass, demonstrate greater resiliency, and are lower input compared to either typical or sustainable agroecosystems [123]. From a forage grass standpoint alone, diverse mixtures of warm-season grasses accumulate more forage compared to their monoculture counterparts [124]. Greater plant community composition potentially results in greater functional group (C3 or C4 grasses, forbs, cool- and warm-legumes) diversity, which may address uneven forage production. When forages of adequate nutritive value are available throughout the entire grazing season, as opposed to only during peak times (as we have observed with C3 or C4 grass monocultures), we should expect an overall increase in animal production.
Management-intensive rotational grazing is another method of achieving sustainable intensification in grasslands, especially in temperate, humid, and sub-humid climates. The intensification is realized through increased output per unit area land. Management-intensive rotational grazing results in increased nutritive value and forage production [89], which contributes to increased carrying capacity and ultimately increased beef produced per ha. Managed grazing systems may better meet the growing demand for dietary protein per unit area compared to extensively managed beef production.
5.3. Implementation of Research
To date, research in eastern grasslands that evaluates trade-offs in terms of livestock production and enhanced biodiversity have been limited. Conducting replicated field experiments at a scale large enough to draw reliable conclusions is costly and logistically difficult. Additional investment in such research is urgently needed to better understand the range of outcomes under varying management scenarios to further inform large-scale conservation strategies. Within-field structural and compositional impacts and how these are influenced by management regimes need to be better understood. Trade-offs in productivity, profitability, and outcomes for improved soil health and carbon sequestration must also be documented. However, it is also important to document how landscape factors affect biodiversity outcomes as well as thresholds at which scale of adoption can bring positive benefits to at-risk elements of biodiversity.
5.4. Evaluation of Tradeoffs for Working Grasslands
It is beyond the scope of this paper to fully develop and describe a decision-making process or framework for assessing tradeoffs for working grasslands. However, we introduce a conceptual framework for making decisions with multiple objectives or attributes. We draw attention to a similar approach for Dutch dairy farming systems [125]. We rely heavily on this approach and nomenclature, as it is directly relevant to sustainable working grasslands in the U.S. However, van Calker et al.’s [126] purpose was to derive a sustainability function, while our purpose is to demonstrate the need to explicitly understand tradeoffs among multiple sustainability indicators for working grasslands
Management requires allocating resources that are typically finite to achieve a desired outcome. A single objective management problem is relatively simple to solve—find the action or set of actions that leads to the best desired outcome. Assuming a cattle producer only valued net revenue, each choice of action would be one that minimized costs and maximized yield. However, cattle producers are not single objective decision makers, as they value the environment, animal well-being, and numerous other outcomes [126]. Moving from a single resource objective problem to multiple objectives raises the complexity and requires tradeoffs. Using multi-attribute theory (MUAT) to arrive at optimal solutions in the presence of multiple objectives will facilitate an explicit and clear understanding of the tradeoffs for different potential management actions [127].
The USRSB identified six aspects or fundamental objectives that would achieve sustainable beef production: reduced air and greenhouse gas emissions, improved land resources, improved water quality, improved employee safety and well-being, improved animal health and well-being, and improved efficiency and yield to improve financial health [2]. Each of these aspects have their own performance metrics and targets that bring a needed specificity to decision making. Clearly, sustainable beef consists of multiple attributes and achieving all these simultaneously will be difficult. We argue that using MUAT will provide a framework to explicit evaluate a set of proposed actions or strategies (multi-actions) to achieve the ultimate goal of sustainability.
Let us create a fictious, but realistic decision-making scenario, using MUAT and the basic framework provided by USRSB such that we have clear objectives. For the sake of brevity and in the spirit of the paper, we will use an example with the land resources and finances aspects or objectives. We can express a general sustainability function (S) as Equation (1):
where Wj represents the weight for the jth aspect of sustainability, Uj is the utility function for the jth aspect, and N is the total number of aspects. Specifically, for the USRSB sustainability, the expression takes the form of Equation (2):
S(UAir, ULand, UWater, UEmployees, UAnimals, UFinances) = UAir * WAir + ULand * WLand + UWater * WWater + UEmployees * WEmployees + UAnimals * WAnimals + UFinances * WFinances
The theoretical goal is for S = 1, which would require Uj = 1; however, this is rarely possible in practice.
Before we can parameterize this function, we need to develop utility functions and objective weights. Again, for simplicity, we will only demonstrate this with two aspects: land resources and finances. Each aspect can have multiple attributes. Utility values are scaled between 0 and 1, with greater values corresponding to a greater degree of satisfaction. For finances, the attributes could be kg/ha of beef produced (i.e., yield) and USD/ha (i.e., input costs). For land resources, it could be grassland bird species richness/ha. We have provided example utility functions for demonstration purposes (Figure 1). For measuring a single aspect of sustainability, the multi-attribute function is used, i.e., Equation (3):
where Uj is the utility function for the jth aspect as defined above, ui(xi) is the utility corresponding to the ith attribute, and wi represent the weight for the ith attribute. For the financial aspect of the USRSB, the utility function would simply be Equation (4):
UFinance(x1,x2) = wYield * uYield + wInputs * uInputs
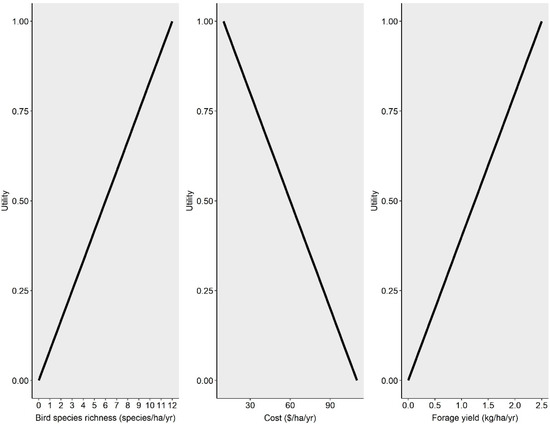
Figure 1.
Hypothetical utility functions demonstrating relationships between utility and bird species richness (l), production costs (center), and forage yield (r) for prioritizing decision making under multiple scenarios for sustainable beef production.
Thus, if we weight these attributes equally (w = 0.5) and the uYield = 0.20 (a yield of 0.5 kg/ha/year) and uinputs = 0.60 (input costs of USD 50/ha/year), the UFinance would equal 0.40 (Figure 1). Similarly, if we expect to only measure three species of birds, ULand would equal 0.25 (Figure 1). These two utility values would then be used in eqn. 2 to determine the overall utility or sustainability once the other aspects are enumerated. For this single-strategy scenario, the land aspect conflicts with the finance aspect, as evidenced by the lower utility (0.40 vs. 0.25); thus, we are trading off satisfaction for the land aspect for gain in the financial aspect.
Exploring tradeoffs with a single strategy to choose from is not all that useful because if you only have one option to choose from then the decision is easy. Thus, this approach is much more useful with two or more strategies to choose from. Let us consider two different strategies for managing a working grassland: Strategy 1 is continuous grazing and Strategy 2 is intensive rotational grazing. If we assume that we value each aspect equally, Strategy 2 provides the greatest overall satisfaction (S = 0.57; calculated using Equation (2)). Notice that we are trading off finances (a 39% decline in UFinance from Strategy 1 to 2; 0.55 vs. 0.37, respectively) but gaining in land resources (77% increase in ULand; 0.31 vs. 0.70, respectively). However, we rarely value each aspect equally, so the weights need to reflect what we are willing to trade off based on the options we have at hand. A common technique for weighting aspects or attributes is called swing weighting. A detailed description of swing weighting can be found at [128] (p. 220). We have provided the outputs of a swing weighting exercise in Table 2. Notice that we now weight finances the most, followed by land, and then the rest equally. Using the new weights, S = 0.53 for Strategy 1 and S = 0.51 for Strategy 2. The decision maker was not willing to trade off finances for land resources but was willing to do the opposite. Thus, Strategy 1 is the optimal choice given the decision maker’s objectives and willingness to make certain tradeoffs. As a society, if we wanted to pay this producer to select Strategy 2 because we collectively value land resources more, then we would have to offset the producer’s finances to eliminate that tradeoff. Using this approach, an explicit accounting of such is possible.
Table 2.
Hypothetical utility values and swing weighing outputs for a simple, two-strategy decision problem. We used the USRSB’s aspects for context but make no claim these values reflect theirs.
6. Conclusions
It has become increasingly clear that grasslands are a critical biome, one that makes a tremendous contribution to the production of essential agricultural products, those that ensure adequate nutrition to a growing global population. It has also become apparent that as a major carbon sink, grasslands can and will play an important role in global C-cycles with all of the attendant implications for future sustainability. It is our hope, though, that this paper will also bring to light the crisis in biodiversity associated with degraded grasslands. We have focused on grasslands of the eastern U.S., where avifauna, pollinators, and perhaps soil biota are at risk and must be conserved. An approach based on a working lands conservation model will ensure that appropriate disturbances, particularly grazing and fire, can be implemented along with an appreciation for enhanced plant diversity and an increased reliance on native plants. Just as importantly, a working lands approach can contribute to farms that are economically viable and can, therefore, remain productive grasslands.
Author Contributions
Conceptualization, P.D.K., D.A.B., J.H.F., D.L.F., S.D.F., J.A.M., H.D.N., S.R.S.; writing—original draft, review, editing, P.D.K., D.A.B., J.H.F., D.L.F., S.D.F., J.A.M., H.D.N., S.R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This paper was not supported by external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- USRSB. U.S. Beef Industry Sustainability Framework, U.S. Roundtable for Sustainable Beef. 2019. Available online: https://www.beefsustainability.us/ (accessed on 13 May 2022).
- USRSB. U.S. Roundtable for Sustainable Beef, High-Priority Indicator Goals & Sector Targets. 2022. Available online: https://www.usrsb.org/goals (accessed on 13 May 2022).
- TNC. A Roadmap to a Sustainable Beef System. The Nature Conservancy. 2021. Available online: https://www.nature.org/content/dam/tnc/nature/en/documents/TNCBeefRoadmap_FINAL_April152020.pdf (accessed on 13 May 2022).
- Rotz, C.A.; Asem-Hiablie, S.; Place, S.; Thoma, G. Environmental footprints of beef cattle production in the United States. Agric. Syst. 2019, 169, 1–13. [Google Scholar] [CrossRef]
- Sanderson, J.S.; Beutler, C.; Brown, J.R.; Burke, I.; Chapman, T.; Conant, R.T.; Derner, J.D.; Easter, M.; Fuhlendorf, S.D.; Grissom, G.; et al. Cattle, conservation, and carbon in the western Great Plains. J. Soil Water Conserv. 2020, 75, 5A–12A. [Google Scholar] [CrossRef]
- Sprunger, C.D.; Martin, T.; Mann, M. Systems with greater perenniality and crop diversity enhance soil biological health. Agric. Environ. Lett. 2020, 5, e20030. [Google Scholar] [CrossRef]
- Hu, J.; Richwine, J.D.; Keyser, P.D.; Li, L.; Yao, F.; Jagadamma, S.; DeBruyn, J.M. Nitrogen Fertilization and native C4 grass species alter abundance, activity, and diversity of soil diazotrophic communities. Front. Microbiol. 2021, 12, 675693. [Google Scholar] [CrossRef]
- Engle, D.M.; Coppedge, B.R.; Fuhlendorf, S.D. From the Dust Bowl to the Green Glacier: Human activity and environmental change in Great Plains grasslands. In Western North American Juniperus Communities; Van Auken, O.W., Ed.; Ecological Studies; Springer: New York, NY, USA, 2008; p. 196. [Google Scholar] [CrossRef]
- Lark, T.J. Protecting our prairies: Research and policy actions for conserving America’s grasslands. Land Use Policy 2020, 97, 104727. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Teague, W.R.; Dowhower, S.L.; Baker, S.A.; Haile, N.; DeLaune, P.B.; Conover, D.M. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 2011, 141, 3–4. [Google Scholar] [CrossRef]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Hildebrandt, A.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Distel, R.A.; Arroquy, J.I.; Lagrange, S.; Villalba, J.J. Designing diverse agricultural pastures for improving ruminant production systems. Front. Sustain. Food Syst. 2020, 4, 596869. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Harrell, W.C.; Engle, D.M.; Hamilton, R.G.; Davis, C.A.; Leslie, D.M., Jr. Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol. Appl. 2006, 16, 1706–1716. [Google Scholar] [CrossRef]
- Powell, A.F.L.A. Responses of breeding birds in tallgrass prairie to fire and cattle grazing. J. Field Ornith. 2008, 79, 41–52. [Google Scholar] [CrossRef]
- Johnson, T.N.; Kennedy, P.L.; Etterson, M.A. Nest success and cause-specific nest failure of grassland passerines breeding in prairie grazed by livestock. J. Wildl. Manag. 2012, 76, 1607–1616. [Google Scholar] [CrossRef]
- Monroe, A.P.; Chandler, R.B.; Burger Jr., L.W.; Martin, J.A. Converting exotic forages to native warm-season grass can increase avian productivity in beef production systems. Agric. Ecosyst. Environ. 2016, 233, 85–93. [Google Scholar] [CrossRef]
- Yeiser, J.M.; Morgan, J.J.; Baxley, D.L.; Chandler, R.B.; Martin, J.A. Private land conservation has landscape-scale benefits for wildlife in agroecosystems. J. Appl. Ecol. 2018, 55, 1930–1939. [Google Scholar] [CrossRef]
- Keyser, P.D.; West, A.S.; Buehler, D.A.; Lituma, C.M.; Morgan, J.J.; Applegate, R.D. Breeding bird use of production stands of native grasses—A working lands conservation approach. Rangel. Ecol. Manag. 2020, 73, 827–837. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Hedges, K.; Hodges, J.; Lituma, C.M.; Loncarich, F.; Martin, J.A. Eastern grasslands: Conservation challenges and opportunities on private lands. Wildl. Soc. Bull. 2019, 43, 382–390. [Google Scholar] [CrossRef]
- Lituma, C.M.; Buckley, B.R.; Keyser, P.D.; Holcomb, E.; Smith, R.; Morgan, J.; Applegate, R.D. Effects of patch-burn grazing and rotational grazing on grassland bird abundance, species richness, and diversity in native grassland pastures of the Midsouth USA. Agric. Ecosyst. Environ. 2021, 324, 107710. [Google Scholar] [CrossRef]
- USDA. United States Department of Agriculture National Agricultural Statistic Service. 2017. US Census of Agriculture. USDA—National Agricultural Statistics Service—2017 Census of Agriculture—Volume 1, Chapter 2: State Level Data. 2017. Available online: https://www.nass.usda.gov/Publications/AgCensus/2017/Full_Report/Census_by_State/index.php (accessed on 28 April 2022).
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of pollinators and natural enemies: A meta-analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Grab, H.; Branstetter, M.G.; Amon, N.; Urban-Mead, K.R.; Park, M.G.; Gibbs, J.; Blitzer, E.J.; Poveda, K.; Loeb, G.; Danforth, B.N. Agriculturally dominated landscapes reduce bee phylogenetic diversity and pollination services. Science 2019, 363, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Loranger-Merciris, G.; Barthes, L.; Gastine, A.; Leadley, P. Rapid effects of plant species diversity and identity on soil microbial communities in experimental grassland ecosystems. Soil Biol. Biochem. 2006, 38, 2336–2343. [Google Scholar] [CrossRef]
- Thompson, G.L.; Kao-Kniffin, J. Diversity enhances NPP, N retention, and soil microbial diversity in experimental urban grassland assemblages. PLoS ONE 2016, 11, e0155986. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.A.; Kuvlesky, W.P. Invited paper: North American grassland birds: 642 an unfolding conservation crisis. J. Wildl. Manag. 2005, 69, 1–13. [Google Scholar] [CrossRef]
- West, A.S.; Keyser, P.D.; Lituma, C.M.; Buehler, D.A.; Applegate, R.D.; Morgan, J. Grasslands bird occupancy of native warm-season grass. J. Wildl. Manag. 2016, 80, 1081–1090. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E. Reconciling food production and biodiversity conservation: Land sharing and land sparing compared. Science 2011, 333, 1289–1291. [Google Scholar] [CrossRef]
- Morrison, M.L. Bird populations as indicators of environmental change. In Current Ornithology; Johnston, R.F., Ed.; Springer: Boston, MA, USA, 1986; pp. 251–429. [Google Scholar]
- Sauer, J.R.; Link, W.A.; Hines, J.E. The North American Breeding Bird Survey. Anal. Results 1966–2019; U.S. Geological Survey Data Release Laurel: Laurel, MD, USA, 2020. [Google Scholar]
- Herkert, J.R. The effects of habitat fragmentation on Midwestern grassland bird communities. Ecol. Appl. 1994, 4, 461–471. [Google Scholar] [CrossRef]
- Ribic, C.A.; Koford, R.R.; Herkert, J.R.; Johnson, D.H.; Niemuth, N.D.; Naugle, D.E.; Bakker, K.K.; Sample, D.W.; Renfrew, R.B. Area sensitivity in North American grassland birds: Patterns and processes. Auk 2009, 126, 233–244. [Google Scholar] [CrossRef]
- Lituma, C.M.; Buehler, D.B. Cost-share conservation practices have mixed effects on priority grassland and shrubland breeding bird occupancy in the Central Hardwoods Bird Conservation Region, USA. Biol. Conserv. 2020, 244, 108510. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1435–1444. [Google Scholar] [CrossRef]
- Van Engelsdorp, D.; Hayes, J.; Underwood, R.M.; Pettis, J. A survey of honey bee colony losses in the U.S., Fall 2007 to Spring 2008. PLoS ONE 2008, 3, e4071. [Google Scholar] [CrossRef]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native pollinators in anthropogenic habitats. Ann. Rev. Ecol. Evol. Syst. 2011, 42, 1–22. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Forister, M.L.; Halsch, C.A.; Nice, C.C.; Fordyce, J.A.; Dilts, T.E.; Oliver, J.C.; Prudic, K.L.; Shapiro, A.M.; Wilson, J.K.; Glassberg, J. Fewer butterflies seen by community scientists across the warming and drying landscapes of the American West. Science 2021, 371, 1042–1045. [Google Scholar] [CrossRef]
- Vidal, O.; Rendon-Salinas, E. Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biol. Conserv. 2014, 180, 165–175. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vazquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R. The conservation and restoration of wild bees. Ann. N.Y. Acad. Sci. 2010, 1195, 169–197. [Google Scholar] [CrossRef] [PubMed]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Gibson, R.H.; Nelson, I.L.; Hopkins, G.W.; Hamlett, B.J.; Memmott, J. Pollinator webs, plant communities and the conservation of rare plants: Arable weeds as a case study. J. Appl. Ecol. 2006, 43, 246–257. [Google Scholar] [CrossRef]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Losey, J.; Vaughan, M. The economic value of ecological services provided by insects. BioScience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.E. The insect apocalypse, and why it matters. Curr. Biol. 2019, 29, R942–R995. [Google Scholar] [CrossRef]
- Liere, H.; Jha, S.; Philpott, S.M. Intersection between biodiversity conservation, agroecology, and ecosystem services. Agroecol. Sustain. Food Sys. 2018, 41, 723–760. [Google Scholar] [CrossRef]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Sanchez, M.E.N.; Arango, J.A.C.; Burkart, S. Promoting forage legume-pollinator interactions: Integrating crop pollination management, native beekeeping and silvopastoral systems in tropical Latin America. Front. Sustain. Food Syst. 2021, 5, 725981. [Google Scholar] [CrossRef]
- Lagerhöf, J.; Stark, J.; Svensson, B. Margins of agricultural field as habitats for pollinating insects. Agric. Ecosyst. Environ. 1992, 40, 117–124. [Google Scholar] [CrossRef]
- Croxton, P.J.; Hann, J.P.; Greatorex-Davies, J.N.; Sparks, T.H. Linear hotspots? The floral and butterfly diversity of green lanes. Biol. Conserv. 2005, 121, 579–584. [Google Scholar] [CrossRef]
- Thies, C.; Tscharntke, T. Landscape structure and biological control in agroecosystems. Science 1999, 285, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entom. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Lavoir, A.-V.; Rodriguez-Saona, C.; Desneux, N. Bottom-up forces in agroecosystems and their potential impact on arthropod pest management. Ann. Rev. Entom. 2022, 67, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Blair, J.M.; Briggs, J.M.; Collins, S.L.; Hartnett, D.C.; Johnson, L.C.; Towne, E.G. The Keystone role of bison in American Tallgrass Prairie: Bison increase habitat heterogeneity and alter a broad array of plant, community, and ecosystem processes. BioScience 1999, 49, 39–50. [Google Scholar] [CrossRef]
- Palmberg, C.; Scherer-Lorenzen, M.; Jumpponen, M.; Carlsson, G.; Huss-Danell, K.; Högberg, P. Inorganic soil nitrogen under grassland plant communities of different species composition and diversity. Oikos 2005, 110, 271–282. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Herbivore-mediated linkages between aboveground and belowground communities. Ecology 2003, 84, 2258–2268. [Google Scholar] [CrossRef]
- Liang, C.; Kao-Kniffin, J.; Sanford, G.R.; Wickings, K.; Balser, T.C.; Jackson, R.D. Microorganisms and their residues under restored perennial grassland communities of varying diversity. Soil Biol. Biochem. 2016, 103, 192–200. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Cook, R. Functional aspects of soil animal diversity in agricultural grasslands. Appl. Soil Ecol. 1998, 10, 263–276. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Gregg, A.L.; Simpson, R.J.; Li, G.D.; Riley, I.T.; McKay, A.C. Pasture management clearly affects soil microbial community structure and N-cycling bacteria. Pedobiologia 2009, 52, 237–251. [Google Scholar] [CrossRef]
- Liu, Y.; Miao, H.-T.; Chang, X.; Wu, G.-L. Higher species diversity improves soil water infiltration capacity by increasing soil organic matter content in semiarid grasslands. Land Degrad. Dev. 2019, 30, 1599–1606. [Google Scholar] [CrossRef]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suprressiveness. Annu. Rev. Phytopath. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Fuhlendorf, S.D.; Engle, D.M. Restoring heterogeneity on rangelands: Ecosystem management based on evolutionary grazing patterns. BioScience 2001, 51, 625. [Google Scholar] [CrossRef]
- Wilcox, B.P.; Fuhlendorf, S.D.; Walker, J.W.; Twidwell, D.; Wu, X.B.; Goodman, L.E.; Treadwell, M.; Birt, A. Saving imperiled grassland biomes by recoupling fire and grazing: A case study from the Great Plains. Front. Ecol. Environ. 2021, 20, 179–186. [Google Scholar] [CrossRef]
- Kremen, C. Reframing the land-sparing/land-sharing debate for biodiversity conservation. Ann. N.Y. Acad. Sci. 2015, 1355, 52–76. [Google Scholar] [CrossRef]
- Wright, H.L.; Lake, I.R.; Dolman, P.M. Agriculture—a key element for conservation in the developing world. Conserv. Lett. 2012, 5, 11–19. [Google Scholar] [CrossRef]
- USDA ERS. Farm Household and Income Characteristics. 2020. Available online: https://www.ers.usda.gov/data-products/farm-household-income-and-characteristics/ (accessed on 17 December 2020).
- USDA ERS. Cash Receipts. 2020. Available online: https://data.ers.usda.gov/reports.aspx?ID=17843 (accessed on 17 December 2020).
- USDA-FSA. U.S. Department of Agriculture-Farm Service Agency, Conservation Reserve Program Statistics. 2018. Available online: https://www.fsa.usda.gov/programs-and-services/conservation-programs/reports-and-statistics/conservation-reserve-program-statistics/index (accessed on 11 April 2019).
- Askins, R.A.; Chvez-Ramrez, F.; Dale, B.C.; Haas, C.A.; Herkert, J.R.; Knopf, F.L.; Vickery, P.D. Conservation of grassland birds in North America: Understanding ecological processes in different regions. Ornithol. Mono. 2007, 64, iii-46. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Sala, O.E.; Lauenroth, W.K. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 1988, 132, 87–106. [Google Scholar] [CrossRef]
- Pykälä, J. Cattle grazing increases plant species richness of most species trait groups in mesic semi-natural grasslands. Plant Ecol. 2005, 175, 217–226. [Google Scholar] [CrossRef]
- Raffaele, E.; Veblen, T.T.; Blackhall, M.; Tercero-Bucardo, N. Synergistic influences of introduced herbivores and fire on vegetation change in northern Patagonia, Argentina. J. Veg. Sci. 2011, 22, 59–71. [Google Scholar] [CrossRef]
- Derner, J.D.; Lauenroth, W.K.; Stapp, P.; Augustine, D.J. Livestock as ecosystem engineers for grassland bird habitat in the Western Great Plains of North America. Rangel. Ecol. Manag. 2009, 62, 111–118. [Google Scholar] [CrossRef]
- Harper, C.A.; Birckhead, J.L.; Keyser, P.D.; Waller, J.C.; Backus, W.M.; Bates, G.E.; Holcomb, E.D.; Brooke, J.M. Avian habitat following grazing native warm-season forages in the mid-South United States. Rangel. Ecol. Manag. 2015, 68, 166–172. [Google Scholar] [CrossRef]
- Denevan, W.M. The Pristine Myth: The landscape of the Americas in 1492. Ann. Assoc. Am. Geogr. 1992, 82, 369–385. [Google Scholar] [CrossRef]
- Delcourt, H.R.; Delcourt, P.A. Pre-Columbian native American use of fire, on southern Appalachian landscapes. Conserv. Biol. 1997, 11, 1010–1014. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.A.; Meunier, J.; Holoubek, N.S. Disentangling effects of fire, habitat, and climate on an endangered prairie specialist butterfly. Biol. Conserv. 2018, 218, 41–48. [Google Scholar] [CrossRef]
- Crider, F.J. Root-Growth Stoppage Resulting from Defoliation of Grass; No. 1102; USDA Tech. Bull.: Washington, D.C., USA, 1955. [Google Scholar]
- Keyser, P.D.; Smith, S.R. Grazing management. In Native Grass Forages for the Eastern United States; Keyser, P.D., Ed.; PB 1893; University of Tennessee Extension: Knoxville, TN, USA, 2021; pp. 189–222. [Google Scholar]
- Oates, L.G.; Undersander, D.J.; Gratton, C.; Bell, M.M.; Jackson, R.D. Management-intensive rotational grazing enhances forage production and quality of subhumid cool-season pastures. Crop Sci. 2011, 51, 892–901. [Google Scholar] [CrossRef]
- Smith, S.R.; Bouton, J.H.; Singh, A.; McCaughey, W.P. Development and evaluation of grazing-tolerant alfalfa cultivars: A review. Can. J. Plant Sci. 2000, 80, 503–512. [Google Scholar] [CrossRef]
- Bunton, G.; Trower, Z.; Roberts, C.; Bradley, K.W. Seasonal changes in forage nutritive value of common weeds encountered in Missouri pastures. Weed Technol. 2020, 34, 164–171. [Google Scholar] [CrossRef]
- Hendricks, T.J.; Tucker, J.J.; Hancock, D.W.; Mullenix, M.K.; Baxter, L.L.; Stewart, R.L., Jr.; Segers, J.R.; Bernard, J.K. Forage accumulation and nutritive value of bermudagrass and alfalfa-bermudagrass mixtures when harvested for baleage. Crop Sci. 2020, 60, 2792–2801. [Google Scholar] [CrossRef]
- Buckman, J.; Hatfield, G.; Teutsch, C. Impact of defoliation height on crown and soil temperature. In Proceedings of the American Forage and Grasslands Council’s Annual Meeting, Greenville, SC, USA, 5–8 January 2020. [Google Scholar]
- Elgersma, A.; Soegaard, K.; Jensen, S.K. Herbage dry-matter production and forage quality of three legumes and four non-leguminous forbs grown in single-species stands. Grass Forage Sci. 2014, 69, 705–716. [Google Scholar] [CrossRef]
- Marten, G.C.; Andersen, R.N. Forage nutritive value and palatability of 12 common annual weeds. Crop Sci. 1975, 15, 821–827. [Google Scholar] [CrossRef]
- Rosenbaun, K.K.; Bradley, K.W.; Roberts, C.A. Influence of increasing common ragweed (Ambrosia artemisiifolia) or common cocklebur (Xanthium strumarium) densities on forage nutritive value and yield in tall fescue pastures and hay fields. Weed Technol. 2011, 25, 222–229. [Google Scholar] [CrossRef]
- Samson, F.; Knopf, F. Prairie conservation in North America. Bioscience 1994, 44, 418–421. [Google Scholar] [CrossRef]
- Noss, R.F.; LaRoe, E.T.; Scott, J.M. Endangered Ecosystems of the United States: A Preliminary Assessment of Loss and Degradation; Biological Report 28; U.S. Deptartment Interior, Fish and Wildlife Service: Washington, DC, USA, 1995. [Google Scholar]
- Keyser, P.D. Wildlife conservation. In Native Grass Forages for the Eastern United States; Keyser, P.D., Ed.; PB 1893; University of Tennessee Extension: Knoxville, TN, USA, 2021; pp. 377–397. [Google Scholar]
- Barnes, T.G.; Madison, L.A.; Sole, J.D.; Lacki, M.J. An assessment of habitat quality for northern bobwhite in tall fescue-dominated fields. Wildl. Soc. Bull. 1995, 23, 231–237. [Google Scholar]
- Barnes, T.G.; DeMaso, S.J.; Bahm, M.A. The impact of 3 exotic, invasive grasses in the southeastern United States on wildlife. Wildl. Soc. Bull. 2013, 37, 497–502. [Google Scholar] [CrossRef]
- Backus, W.M.; Waller, J.C.; Bates, G.E.; Harper, C.A.; Saxton, A.; McIntosh, D.W.; Birckhead, J.; Keyser, P.D. Management of native warm-season grasses for beef cattle and biomass production in the mid-South USA. J. Anim. Sci. 2017, 95, 3143–3153. [Google Scholar] [CrossRef]
- Keyser, P.D.; Lituma, C.M.; Bates, G.E.; Holcomb, E.D.; Waller, J.C.; Griffith, A.P. Evaluation of eastern gamagrass and a sorghum × sudangrass for summer pasture. Agron. J. 2020, 112, 1702–1712. [Google Scholar] [CrossRef]
- Brazil, K.A.; Keyser, P.D.; Bates, G.E.; Saxton, A.M.; Holcomb, E.D. Continuous grazing of mixed native warm-season grass in the fescue belt. Agron. J. 2020, 112, 5067–5080. [Google Scholar] [CrossRef]
- Walk, J.W.; Warner, R.E. Grassland management for the conservation of songbirds in the midwestern USA. Biol. Conserv. 2000, 94, 165–172. [Google Scholar] [CrossRef]
- West, A.S.; Keyser, P.; Morgan, J. Northern bobwhite survival, nest success, and habitat use in Kentucky during the breeding season. In Proceedings of the National Quail Symposium VII, Tucson, AZ, USA, 10 January 2012; pp. 217–222. [Google Scholar]
- Antunes, P.M.; Miller, J.; Carvalho, L.M.; Klironomos, J.N.; Newman, J.A. Even after death the endophytic fungus of Schedonorus phoenix reduces the arbuscular mycorrhizas of other plants. Funct. Ecol. 2008, 22, 912–918. [Google Scholar] [CrossRef]
- Mack, M.L.; Rudgers, J.A. Balancing multiple mutualists: Asymmetric interactions among plants, arbuscular mycorrhizal fungi, and fungal endophytes. Oikos 2008, 117, 310–320. [Google Scholar] [CrossRef]
- Burns, J.C.; Fisher, D.S. Steer performance and pasture productivity among five perennial warm-season grasses. Agron. J. 2013, 105, 113–123. [Google Scholar] [CrossRef]
- Lowe, J.K., II; Boyer, C.N.; Griffith, A.P.; Waller, J.C.; Bates, G.E.; Keyser, P.D.; Larson, J.A.; Holcomb, E.D. The cost of feeding bred dairy heifers on native warm-season grasses and harvested feedstuffs. J. Dairy Sci. 2016, 99, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Boyer, C.N.; Zechiel, K.; Keyser, P.D.; Rhinehart, J.; Bates, G.E. Risk and returns from grazing beef cattle on warm-season grasses in Tennessee. Agron. J. 2020, 112, 301–308. [Google Scholar] [CrossRef]
- Monroe, A.P.; Burger, L.W.; Boland, H.T.; Martin, J.A. Economic and conservation implications of converting exotic forages to native warm-Season grass. Glob. Ecol. Conserv. 2017, 11, 23–32. [Google Scholar] [CrossRef]
- Keyser, P.D. Roles for native grasses. In Native Grass Forages for the Eastern United States.; Keyser, P.D., Ed.; PB 1893; University of Tennessee Extension: Knoxville, TN, USA, 2021; pp. 65–78. [Google Scholar]
- Brazil, K. Avian Density and Nest Survival and Beef Production on Continuously-Grazed Native Warm-Season Grass Pastures. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, August 2019. [Google Scholar]
- Richwine, J.D. Interseeding Native Warm-Season Grass Pastures. Ph.D. Thesis, University of Tennessee, Knoxville, TN, USA, August 2021. [Google Scholar]
- Pierce, R.A.; Wright, R.L.; Gantzer, C.J.; Reinbott, T.M.; Udawatta, R.P.; Kallenbach, R.L. Using Native Warm Season Grass, Forb and Legume Mixtures for Biomass, Livestock Forage and Wildlife Benefits: A Case Study; G9424; University of Missouri Extension: Columbia, MO, USA, 2017. [Google Scholar]
- Ashworth, A.; Keyser, P.; Allen, F.; Bates, G.; Harper, C. Intercropping Legumes with Native Warm Season Grasses for Livestock Forage Production in the Mid-South; SP 731-G; University of Tennessee Extension: Knoxville, TN, USA, 2012. [Google Scholar]
- Huber, R.; Le’Clec’h, S.; Buchmann, N.; Finger, R. Economic value of three grassland ecosystem services when managed at the regional and farm scale. Sci. Rep. 2022, 12, 4194. [Google Scholar] [CrossRef]
- Smith, B.; Leung, P.; Love, G. Intensive Grazing Management: Forage, Animals, Men, Profits; The Graziers Hui.: Kamuela, HI, USA, 1986; 350p. [Google Scholar]
- Stanley, P.L.; Rowntree, J.E.; Beede, D.K.; DeLonge, M.S.; Hamm, M.W. Impacts of soil carbon sequestration on life cycle greenhouse gas emissions in Midwestern USA beef finishing systems. Agric. Syst. 2018, 162, 249–258. [Google Scholar] [CrossRef]
- Keyser, P.; Schexnayder, S.; Willcox, A.; Bates, G.; Boyer, C. Identifying barriers to forage innovation: Native grasses and producer knowledge. J. Ext. 2019, 57, 6RIB4. [Google Scholar]
- Pretty, J.; Bharucha, Z.P. Sustainable intensification of agricultural systems. Ann. Botany 2014, 114, 1571–1596. [Google Scholar] [CrossRef]
- Gliessman, S.R. Agroecology and agroecosystems. In The Earthscan Reader in Sustainable Agriculture; Pretty, J., Ed.; Earthscan: London, UK, 2005; pp. 104–114. [Google Scholar]
- Paredes, S.S.; Stritzler, N.P.; Bono, A.; Distel, R.A. Perennial warm-season grass monocultures and mixtures: Biomass production and soil improvement in semiarid and shallow soil conditions. J. Arid. Environ. 2018, 154, 82–88. [Google Scholar] [CrossRef]
- van Calker, K.J.; Berentsen, P.B.M.; Romero, C.; Giesen, G.W.J.; Huirne, R.B.M. Development and application of a multi-attribute sustainability function for dutch dairy farming systems. Ecol. Econ. 2006, 57, 640–658. [Google Scholar] [CrossRef]
- Sitienei, I.; Gillespie, J.; Scaglia, G.U.S. Grass-fed beef producers: Goal structure and reasons for enterprise selection. J. Agric. Appl. Econ. 2020, 52, 78–95. [Google Scholar] [CrossRef]
- Romero, C.; Rehman, T. Multiple Criteria Analysis for Agricultural Decisions; Elsevier: Amsterdam, the Netherlands, 2003; Volume 11, ISBN 0444503439. [Google Scholar]
- Gregory, R.; Failing, L.; Harstone, M.; Long, G.; McDaniels, T.; Ohlson, D. Structured Decision Making: A Practical Guide to Environmental Management Choices; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 1444398539. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).