1. Introduction
Capsicum annuum L. is an economic important horticultural crop [
1]. Pepper fruit is an excellent source of antioxidants, vitamins, proteins, carbohydrates, fats and phenolic compounds [
2]. However, despite its economic importance, only limited published information exists in the literature for
Capsicum annuum L. Gisbert-Mullor et al. (2020) [
3] investigated the influence of grafting onto an appropriate rootstock on the control of deficit irrigation, while in the study of Karakurt et al. (2009) [
4] the influence of foliar and soil application of humic acids on pepper yields was studied, with promising results. The foliar application of Ca was concluded to be a good agronomic practice, in order to mitigate the negative effects of saline water supply in sweet pepper plants [
5].
Especially with regard to the fertilization needs of pepper crop, few data have been published on the effect of alternative sources of nutrients’ application on the nutrient uptake, physiological performance and fruit quality of pepper plants. Aminifard et al. (2012) [
6] studied the effect of N fertilization on pepper plants under field conditions and found that a fertilization rate of 50 g N ha
−1 led to the optimum yield and fruit quality at ripening. In a similar research work, the effect of different N forms and concentrations on sweet pepper flowering and fruit yield was studied [
7]. According to the same authors, nutrient uptake influenced flower abortion and fruit set in sweet pepper plants [
8], while in a third study of the same authors, they investigated the interaction between ammonium and K in sweet pepper plants [
9]. In another study by Nabi et al. (2006) [
10], the B nutrition of four sweet pepper cultivars grown in a B-deficient soil was investigated, while the effects of foliar spray of different Fe sources on pepper plants were also studied [
11]. Regarding the impact of organic fertilization on
Capsicum annuum L. crop, the influence of different compost rates (0, 5, 10 and 15 ton ha
−1) in sweet pepper plants was investigated, and it was found that compost had a strong impact on fruit quality and antioxidant components under field conditions [
12]. The study of Diaz-Perez et al. (2021) [
13] focused on the compost (made from a mix of chicken litter, yard trimmings, cull vegetables, peanut hay, cotton gin trash, clay and cured compost) effects at the transplant stage on the growth, nutrition and yields of bell pepper plants; they concluded that compost application at 10 t per ha before planting resulted in an increase in soil organic matter, as well as in foliar nutrient concentrations [
13]. Singh and Rathore (2021) [
14] studied the effects of fertilization with textile effluent on the germination, growth and some metabolites of
Capsicum annuum L. cultivars, and they concluded that it increased plant biomass, while it could also function as nutrient additive for chilli cultivars.
As it is clear from all the above-mentioned studies, no published comparative data exist between inorganic (conventional) fertilization and other sources of nutrient application (e.g., organic and/or inorganic soil amendments, such as manures, zeolite, etc.) in pepper crop; in addition, whether such kind of soil amendments could partially substitute chemical fertilizers’ application for decreasing high fertilization rates has not been investigated. The unique exception was the study of Assimakopoulou et al. (2020) [
15], who investigated the effects of combined olive mill wastewaters (three levels: 0, 2.5 and 5%) and natural zeolite (three levels: 0, 2.5 and 5%) application on the biomass, yields and nutrition of pepper plants; however, their study was not comparative (as our study was) between inorganic (conventional) fertilization and nutrient supply of alternative origin. Our research was based on the premise that alternative (nonconventional) sources of nutrient application (amendments, such as goat manure, zeolite, vermiculite, etc.) could sufficiently satisfy pepper nutritional needs (this was realized via a foliar analysis of nutrients) and could improve K uptake for plants and soil organic C. Furthermore, it was hypothesized that the soil application of suitable inorganic and organic amendments could significantly influence nutrient uptake, physiology and fruit quality.
Thus, the aims of our study were: (a) to compare the influence of inorganic (zeolite, vermiculite) and organic amendments (goat manure) with that of CRF application on the soil fertility, growth, nutrition, physiological performance and fruit quality of Capsicum annuum L. plants and (b) to evaluate the efficiency and suitability of alternative sources of nutrients (amendments) to satisfy the nutritional needs of pepper plants, via a comparative approach with a conventional inorganic CRF fertilizer. For the needs of the study, the cv. Bounty F1-Lenor was chosen, which is one of the most important and productive pepper hybrids, suitable either for field cultivation or for crop growth under greenhouse environment.
2. Materials and Methods
2.1. Plant Material, Soil Sampling and Treatments
Capsicum annuum L. plants of the cv. Bounty F1-Lenor, approximately 12–15 cm in height, were grown outdoors, in 5 L pots, for 140 days (from the 4th of May until the 21st of September), on a soil substrate from parent material gneiss. Pots were filled with soil collected from the region of Vasilika (40°28′48.0864″, 23°8′18.492″ N/E), close to the city of Thessaloniki, Macedonia, northern Greece. Most of this region is traditionally occupied with vegetable crops and the initial soil pH (6.52) was close to the optimum one for vegetable crops. However, organic matter, exchangeable K levels, as well NO3 and Olsen P concentrations were very low (0.8%, 65 mg kg−1, 2.16 mg kg−1 and 1.44 mg kg−1, respectively) compared to the optimum ones (they should be >2%, >100 mg kg−1, 20–40 mg kg−1 and 15–25 mg kg−1, respectively) for pepper plants’ growth; thus, the crucial matter for our study was to find out suitable soil amendments (i) firstly aiming to improve K and organic C levels, as well as (ii) to boost the low N and P concentrations in this vegetable growing zone. Samples were collected from the upper 30 cm of the soil profile, since most of the root system of vegetable crops grows in this layer. After soil samples were received (before filling the pots), they were carefully mixed, in order to achieve homogenization. The pepper plants were randomized (based on their initial height) and divided into 6 similar groups, corresponding to each one of the 6 treatments described below.
The treatments of the experiment were the following: (i) soil/ver (“Vermiterra medium” exfoliated vermiculite for horticultural uses, provided by the company NORDIA A.E., Greece) = 80:20; (ii) soil/zeo (“Zeoterra”, powder size 0.8–2.5 mm, provided by the company NORDIA A.E., Greece) = 80:20; (iii) soil: zeo/ver = 80:10:10; (iv) no mixture of soil with amendments, only the application of controlled release fertilizer (CRF); (v) soil/ man/ver = 80:5:15; (vi) soil/man/zeo = 80:5:15. The relations of mixtures between soil and amendments were expressed in % w/v. In the 4th treatment, a total quantity of 10 g of CRF per pot (during the whole experimental period) was applied, in two doses of 5 g each: the first was applied in June and the second in July.
The exact composition of the CRF was the following: 21% total N (11% NH
4-N and 10% NO
3-N), 5% P
2O
5, 10% K
2O, 3% MgO, 6% total S, 0.02% total B, 0.3% total Fe and 0.02% total Zn. The nutritional composition of the goat manure was the following: N—2.80%; P—0.13%; K—2.54%; Ca—1.15%; Mg—1.10%; Na—0.10%; Fe—1927 mg kg
−1; Mn—367 mg kg
−1; Zn—81 mg kg
−1; and Cu—15 mg kg
−1. The fertilizer was solid, and it was slightly incorporated into soil. The choice of the ratios 80:20 or 80:10:10 between the soil substrate and the inorganic amendments was based on previously published data [
16,
17]. The chemical properties of zeolite (“Zeoterra”, powder size 0.8–2.5 mm) and vermiculite (“Vermiterra medium”), respectively, were the following: pH, 7.28 and 8.83; organic matter, 0.00 and 0.18%; NO
3-N, 9.36 and 6.58 mg kg
−1; P, 1.87 and 2.53%; K, 14,984 and 445 mg kg
−1; Ca, 16,050 and 1727 mg kg
−1; Mg, 922 and 146 mg kg
−1; Fe, 0.57 and 14.44 mg kg
−1; Mn, 1.12 and 1.92 mg kg
−1; Zn, 0.24 and 0.33 mg kg
−1; Cu, 0.05 and 0.47 mg kg
−1.
In the present experiment, six replications were set up for each treatment, leading to a total number of 36 plants. During the experiment, all the plants were irrigated daily (with an automated spray irrigation system) with high-quality water, to achieve a soil moisture of approximately 70% of the water-holding capacity. In addition, during the whole experimental period, the plants were sprayed 5 times with Cu, in order to combat Alternaria solani and Alternaria alternata.
2.2. Chemical Analyses of Soil Samples and Mixtures
After representative soil samples were received from the field, they were mixed with the amendments (as described above, in the 6 treatments), and a quantity of approximately 1–1.5 kg from each mixture treatment (used as plant substrate) was transferred to the laboratory for chemical analyses, to determine the fertility of the soil mixtures. Before analyses, soil mixtures were dried at room temperature, their stones were removed, and afterwards, they were sieved to pass a 10-mesh screen. Chemical analyses included: pH, particle size analysis, organic matter, NO
3-N, NH
4-N, available P, exchangeable cations (K, Ca and Mg) and extractable micronutrients (Fe, Mn, Zn and Cu). These parameters were determined as follows: pH in a soil-distilled water paste (1:1) [
18], the particle size analysis according to the Bouyoucos method [
19] and organic matter with the potassium dichromate method [
20]; the NO
3-N concentration was determined according the VCl
3/Griess method [
21], while extractable NH
4-N was measured after extracting 2.5 g of soil with 15 mL of 2 M KCl for 1 h [
22]. The available P was determined according to the Olsen method [
23], the exchangeable cations (Ca, Mg and K) after extraction with ammonium acetate [
24], and finally the extractable micronutrients (Fe, Mn, Zn and Cu) were determined with the DTPA (pH 7.3) method [
25]. The concentrations of K, Ca, Mg, Fe, Mn, Zn and Cu were measured by ICP (OPTIMA 2100 DV optical emission spectrometer, Perkin Elmer, Waltham, MA, USA) [
26].
2.3. Plant and Fruit Growth
At the end of the experiment, the length of the plants’ main shoot was measured; in addition, the elongation rate (cm/day) of the main shoot of pepper plants during the experimentation period was also calculated. After all the plants had been harvested, the leaves were separated from the stem, and the root system from the other plant parts. Before being washed, the fresh weights (F.W.) of all the plant tissues were measured. By the addition of the F.W. of all the plant parts, the total plant fresh weight was calculated. After all the plant tissues had been carefully washed (once with tap and twice with distilled water), they were dried at 75 °C for 48 h. Then, the dry weights (D.W.) of the root, stem and leaves were determined. By the addition of the D.W. of all the plant tissues, total plant dry weight was calculated. Furthermore, the ratio (leaves + stem)/root was also determined. Finally, the mean fresh fruit weights among the treatments were determined at the end of the experiment.
2.4. Tissue Nutrient Analyses, Total Plant Nutrient Content and Nutrient Use Efficiency
At the end of the experiment, after the plants had been harvested and the tissues separated, washed and dried, they were ground to a fine powder to pass a 30-mesh screen. A portion of 0.5 g of fine powder of each sample was dry-ashed in a muffle furnace, at 515 °C, for 5 h. Then, the ash was dissolved with 3 mL of 6 N HCl and diluted with double-distilled water up to 50 mL. The concentrations of P, K, Ca, Mg, Fe, Mn, Zn and Cu were determined by ICP (OPTIMA 2100 DV optical emission spectrometer, Perkin Elmer, Waltham, MA, USA) [
26]. Nitrogen was determined by the Kjeldahl method [
27]. Macronutrient concentrations were expressed in % D.W., while those of micronutrients (Fe, Mn, Zn and Cu) in mg kg
−1. Multiplying the concentration of each nutrient in each plant tissue by the corresponding D.W., the content (absolute quantity) of each nutrient in each plant part was determined at the end of the experiment. By adding the nutrient contents of the different plant parts, the total plant nutrient content (i.e., the total nutrient uptake) was calculated. With regard to the determination of the nutrient concentrations in fruits, they were first crushed and dried at 75 °C for 24 h, then they were dry-ashed at 515 °C for 5 h, and the concentrations were calculated according to the methodology described above. Finally, the nutrient use efficiency was calculated as the ratio of total plant biomass (D.W.)/mg of macronutrient (or μg of micronutrient), taken up by the plants, according to Chapin and Van Cleve (1991) [
28].
2.5. Chlorophyll Fluorescence and Gas Exchange Measurements
At the end of the experiment, the following chlorophyll fluorescence parameters were determined: F
v/F
m, the maximum quantum yield of primary photochemistry; F
0, the minimum fluorescence; F
m, the maximum fluorescence; F
v = F
m − F
0, the variable fluorescence; and the performance index (PI), which reflects the functionality of both photosystems I and II and provides a quantitative information on the current state of plant performance under stress conditions [
29]. All these parameters were determined on the youngest, fully expanded, mature leaves (from the upper part of the main shoot) of pepper plants, by the PAM-2000 fluorometer (Heinz Walz GmbH, Effeltrich, Germany), after leaf preconditioning under dark conditions, for 20 min [
30].
For the gas exchange measurements (i.e., net photosynthetic rate, transpiration rate, stomatal conductance, intercellular CO2 concentration), the LC PRO portable gas exchange measuring system (ADC Bioscientific Ltd., Hoddesdon, UK) was used. The measurements were carried out on the youngest, mature, fully expanded leaves. Measurements were performed in the time period from 10:00 to 12:00 a.m., at natural full light intensity. Finally, the intrinsic water use efficiency (WUEi) was determined as the net photosynthetic rate per stomatal conductance, since it provided information on how efficiently water was used at the plant level.
2.6. Total Soluble Solids, Total Phenolics and FRAP
At harvest, the fruits of each replication were macerated in a blender for the compositional analysis. The soluble solids content (SSC) was measured in the juice of the blended material, using an Atago PR-1 refractometer (Atago Co., Ltd., Tokyo, Japan). The total soluble phenols were determined with the Folin–Ciocalteu reagent [
31]. The total antioxidant activity was determined by a FRAP assay [
32].
2.7. qPCR Expression Analysis
Total RNA was isolated from each sample using the RNeasy extraction kit (Qiagen, Hilden Germany). The quantity and quality of total RNA were assessed by spectrophotometric and electrophoretic analyses, measuring both the absorbance at 260 nm and the absorbance ratio of 260/280 nm in Nanodrop (Thermo, Wilmington, NC, USA), as well as, by 1.5%
w/v agarose-gel electrophoresis. The elimination of total DNA and the cDNA synthesis were performed with RNAse free DNaseI (Takara, Otsu Shiga, Japan) and AffinityScript™ Multi Temperature Reverse Transcriptase (Stratagene, Santa Clara, CA, USA), respectively, according to the manufacturer’s instructions. The complete DNA removal was confirmed with primers designed against Ubiquitin (UBQ—NCBI accession number: AY486137.1), while
Capsicum annuum genomic DNA was used as a positive control. UBQ was also used for the normalization of the first-strand cDNA. Gene specific primers for paper genes coding for sugar transporters, UGP-ase (UDP-glucose pyrophosphorylase) and SPS (Sucrose phosphate synthase), and UBQ genes were designed using Beacon designer v 7.01. Quantitative real-time PCR reactions were performed on a MX-3005P system (Stratagene, Santa Clara, CA, USA) using Kapa Fast Universal 2X qPCR Master Mix (Kapa, Woburn, MA, USA). The expression levels of
Capsicum annuum UBQ were used for normalization. The relative quantification of gene expression was performed, as previously described by Pappi et al. (2021) [
33]. For all samples, qPCR reactions were performed in triplicates. The UBQ gene was used as a reference gene, and the actin gene was used for the verification of the results.
2.8. Statistical Analysis
The experimental design consisted of a 6 × 1 completely randomized factorial, including 6 treatments and one pepper cultivar (Bounty F1-Lenor). Six replicates per treatment were included, leading to 36 experimental plants in total. The data were analyzed by the SPSS statistical program (ONE-WAY ANOVA); for the comparison of the means among the treatments, the Duncan’s multiple range test, for p ≤ 0.05, was used.
4. Discussion
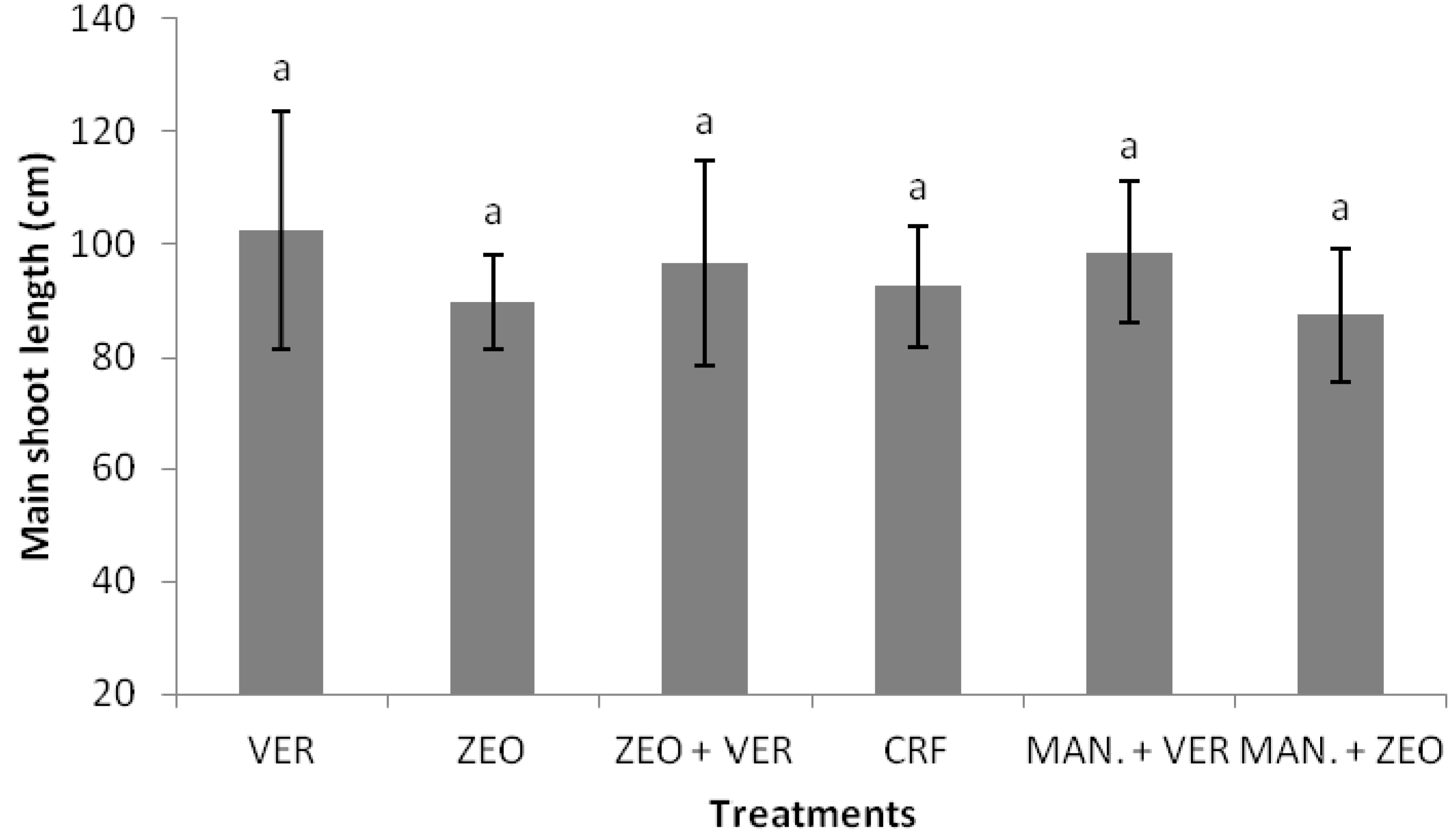
Plant growth (as indicated by total plant biomass and main shoot length) did not significantly differ among CRF and the other treatments (
Table 1;
Figure 1). This in contrast to the results of Chatzistathis et al. (2020) [
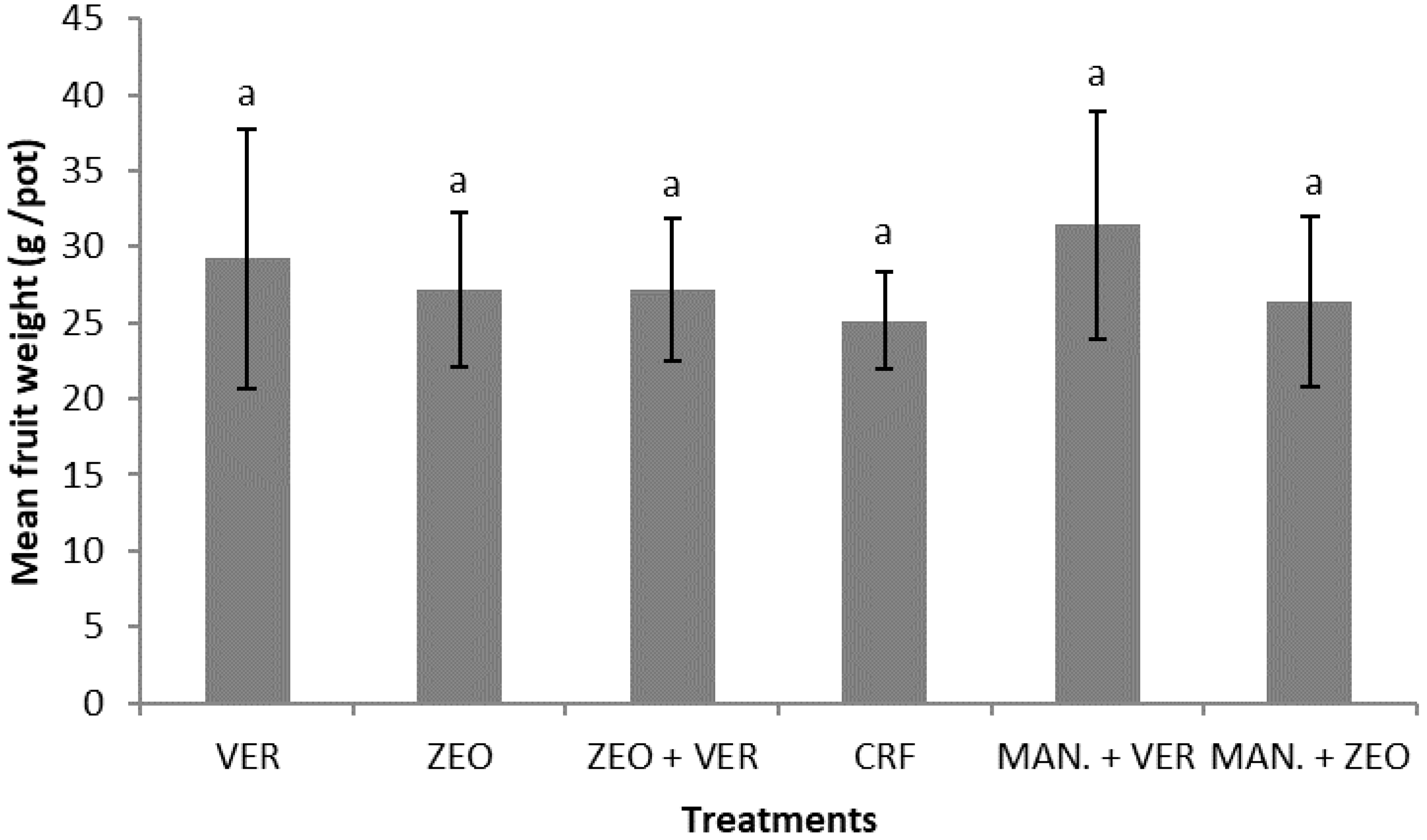
16] for tomato plants, who found that the highest total biomass was recorded in the inorganic fertilization (CRF) compared to the other treatments. The possible differences between the data of this study and those of Chatzistathis et al. (2020) should be mainly focused on the differential response between the two vegetable crop species to the fertilization treatments. With regard to the mean fruit weight, insignificant differences were also found among the treatments (
Figure 2); our results are partially in disagreement with the data of Assimakopoulou et al. (2020) [
15], who found a higher pepper fruit biomass under the treatment with 5% zeolite and 2.5% olive mill wastewater (OMW), compared to the plants that received 2.5% zeolite and 2.5% OMW. The differences in fruit biomass between our data and those of Assimakopoulou et al. (2020) [
15] could be possibly ascribed to the differences in the organic amendment (goat manure vs. OMW) and the other materials used as nutrient suppliers (vermiculite, vermiculite + zeolite, etc.). In the study of Chatzistathis et al. (2020) [
16] it was found that the mean weight of tomato fruits was significantly higher in the inorganic fertilization (CRF) treatment; the differences between the data of the present study and those of Chatzistathis et al. (2020) for tomato crop should be ascribed again to the differential response of the two vegetable crop species.
A significantly higher foliar N concentration (3.84% D.W.) was recorded in the CRF compared to all the other treatments, while the leaf’s K was significantly higher with the VER + ZEO (5.10% D.W.) and ZEO (4.51% D.W.) treatments (
Table 3), which shows the ability of zeolite to enhance soil exchangeable K and its uptake by plants, since it has a high K concentration (14,984 mg kg
−1 vs. 445 mg kg
−1 for vermiculite). Indeed, other studies have also shown the beneficial effects of zeolite application on soil fertility and K nutrition for plants [
15,
34]. In contrast to K, where the independent application of zeolite, or in combination with vermiculite, seems to be a good agronomic practice in order to satisfy K nutritional needs of pepper plants, N nutritional needs were not sufficiently satisfied by inorganic and organic amendments (zeolite, vermiculite and goat manure), since in all these cases, foliar N varied from 1.43% to 2.73% D.W. (
Table 3). According to Bergmann (1986), the optimum range for foliar N, P and K concentrations in pepper plants should be 3.0–4.5% D.W., 0.3–0.6% D.W. and 4.0–5.4% D.W., respectively [
35]. Based on this categorization, only foliar N in CRF was sufficient (3.84% D.W.) and within the optimum range of sufficiency, while in all the other treatments, the leaf’s N concentrations were insufficient (
Table 3). Foliar P levels with the VER, VER + ZEO and MAN + ZEO treatments were marginal to slightly deficient (0.28% D.W., 0.30% D.W. and 0.29% D.W., respectively) (
Table 3), while only the leaf’s K concentrations recorded in the ZEO and VER + ZEO treatments were within the optimum levels of sufficiency (4.51% D.W. and 5.10% D.W., respectively); in all the other treatments, foliar K was deficient (<3.0% D.W.) (
Table 3). From these data, it can be concluded that ZEO application or ZEO coapplication with VER was beneficial in satisfying the K nutritional needs of pepper plants; thus, their use as soil amendments to enhance K uptake by pepper plants is suggested. However, further multiyear additional research, under field conditions, is required to verify these results.
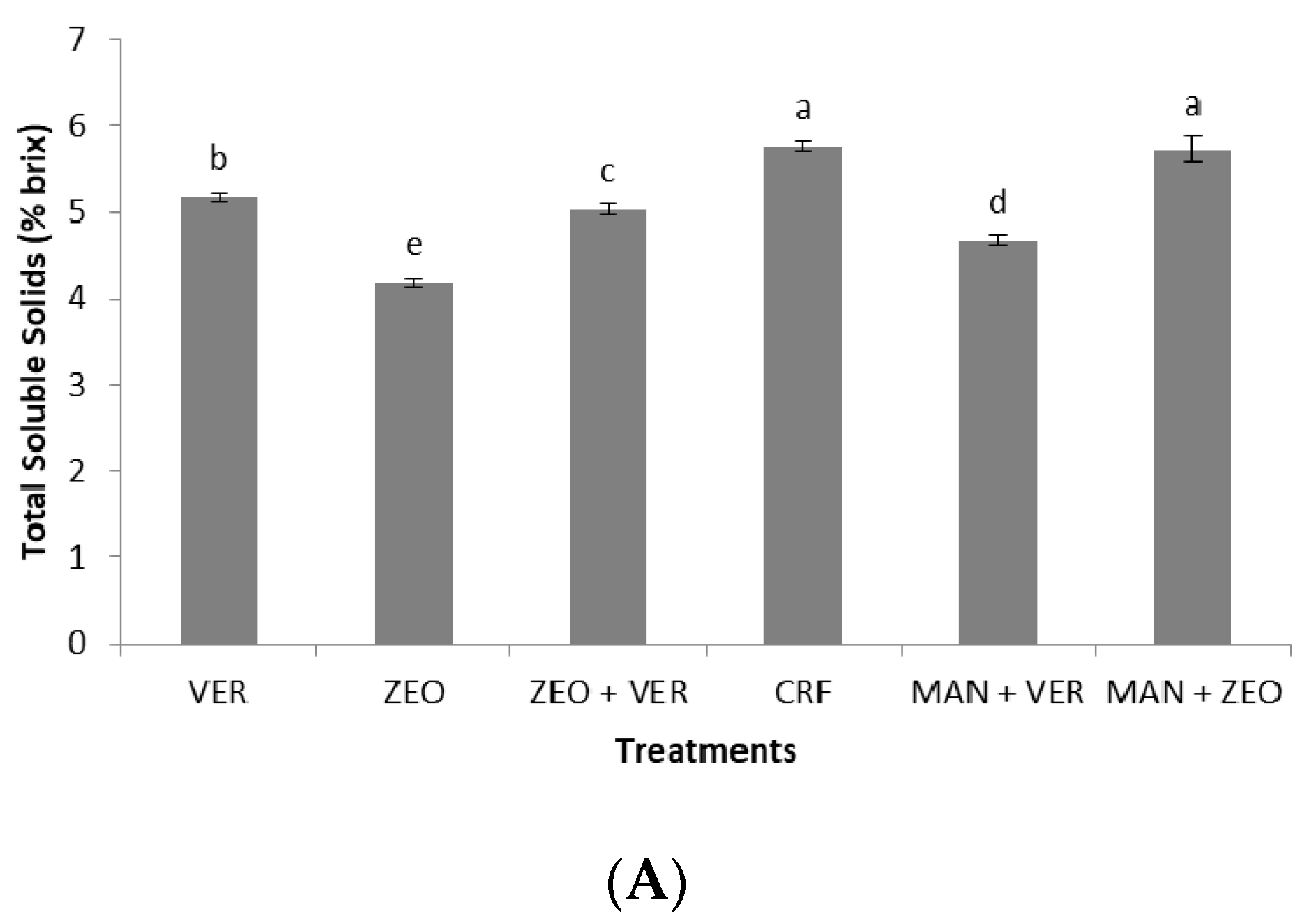
Fruit K, which influences the qualitative fruit characteristics [
36], was significantly higher with the ZEO treatment (4.27% D.W.), compared to all the other treatments (
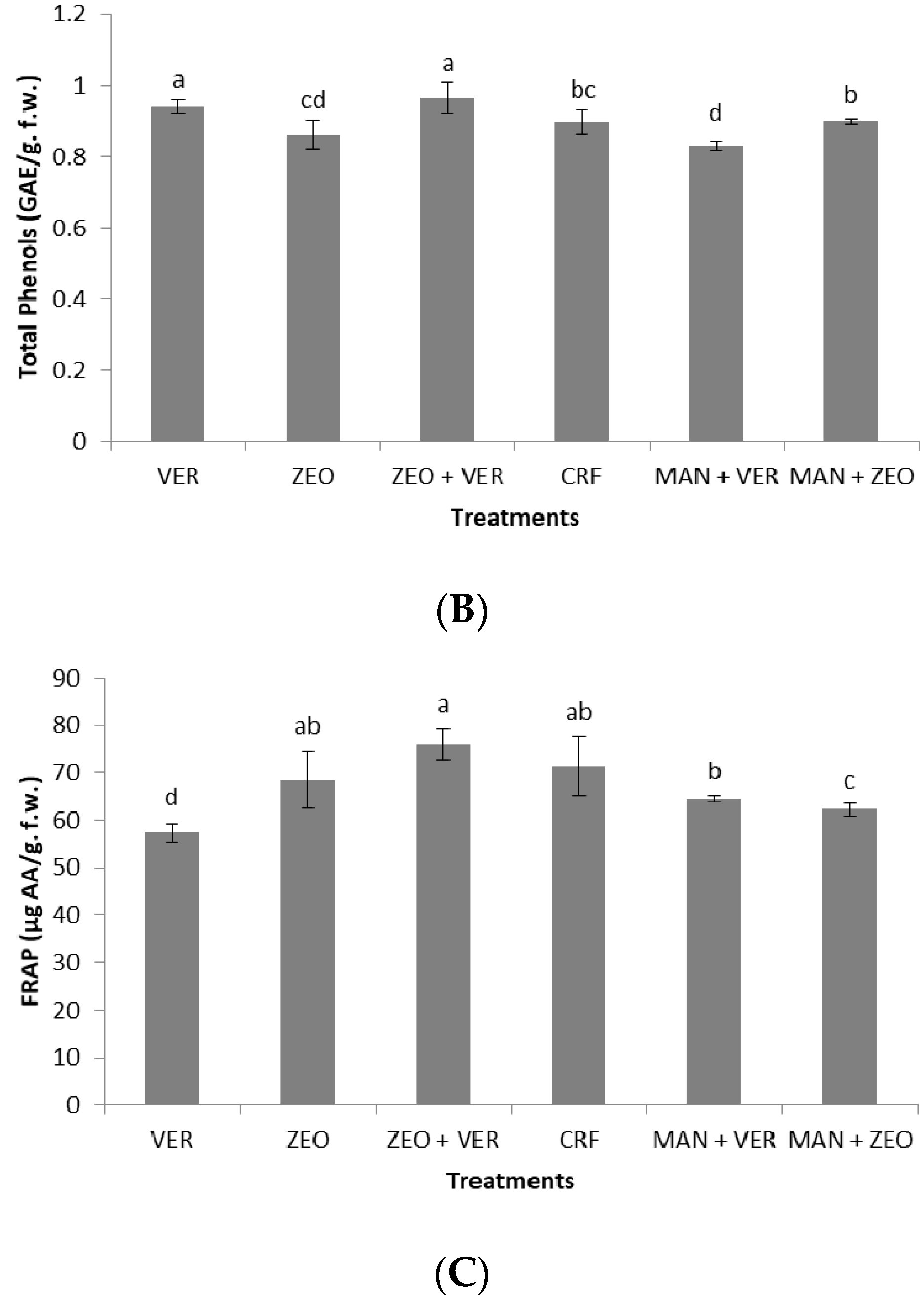
Table 5). Nevertheless, the total soluble solids and total phenols were not positively influenced by the high fruit K levels under ZEO application (
Figure 5A,B); in contrast, the optimum fruit quality was achieved either in the ZEO + VER, CRF, MAN + ZEO, or in the VER treatments (
Figure 5A–C). It seems possible that the influence of K on fruit quality is more profound for other plant species (e.g., tomato, fruits of perennial tree species), rather than for
Capsicum annuum L. This is also supported by the fact that insignificant differences among the treatments were found for the mean fruit weight of pepper plants (
Figure 2). Thus, it is clear from all the above-mentioned results, that further studies, in the near future, are required to draw stable conclusions about the effects of the optimum K nutrition on pepper fruit quality. Glisic et al. (2009) [
37] found that the combinational supply of zeolite with manure provided similar fruit quality characteristics (soluble solids, total acids) with the coapplication of the inorganic fertilizer 15-15-15 and manure, which partially confirms our data for total soluble solids (
Figure 5A). Finally, the coapplication of the inorganic fertilizer 15-15-15, calcium ammonium nitrate and natural zeolite induced the highest levels of total phenolics, flavonoid content and total antioxidant capacity in apricot fruits [
38].
The glucose formation via photosynthesis is the first step of the processes taking place towards the maintenance of the cellular homeostasis of all plant organs [
39]. These processes involve di- and polysaccharide formation, transport via the cellular membranes and the tonoplast as well as phloem loading and translocation to the sink organs [
39,
40]. Rather recently, a group of specific sugar transporters with discrete roles was identified as a critical regulator of these processes that function in parallel to the well-established biosynthetic routes involving the formation of simple and more complex sugars as well as a plethora of secondary metabolites [
41,
42]. The importance of the Sweet 2 (SW2) transporter for the phloem loading was identified [
43], while the expression of the N3 gene was correlated to the soluble solids’ content in figs [
44]. Our results suggest that the substrate may substantially influence the expression of sugar transporters, as well as the expression of genes coding for enzymes critical for the sucrose synthesis, the disaccharide that is synthesized mainly in the leaves and then transferred via the phloem to support the non-photosynthetic organs [
40,
45] in a comparable way to the expression of a specific potassium transporter that was found to be closely associated to the fruit ripening and accumulation of total soluble solids in strawberry [
46]. The coexpression of a Sweet transporter and SPS was also observed in
Vitis vinifera L berries [
47]. Since the K levels are closely associated with the phloem’s ability to transfer several metabolites [
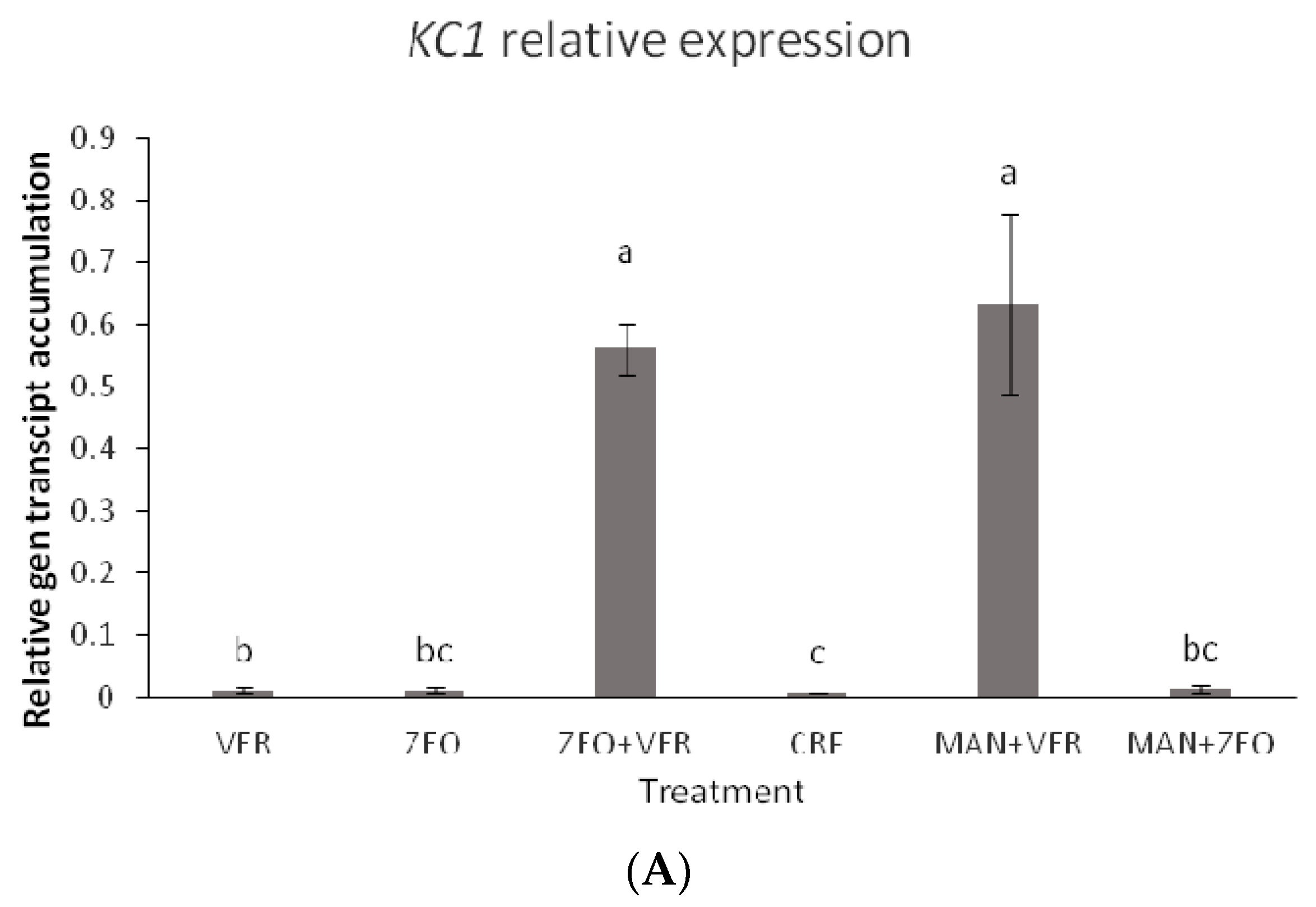
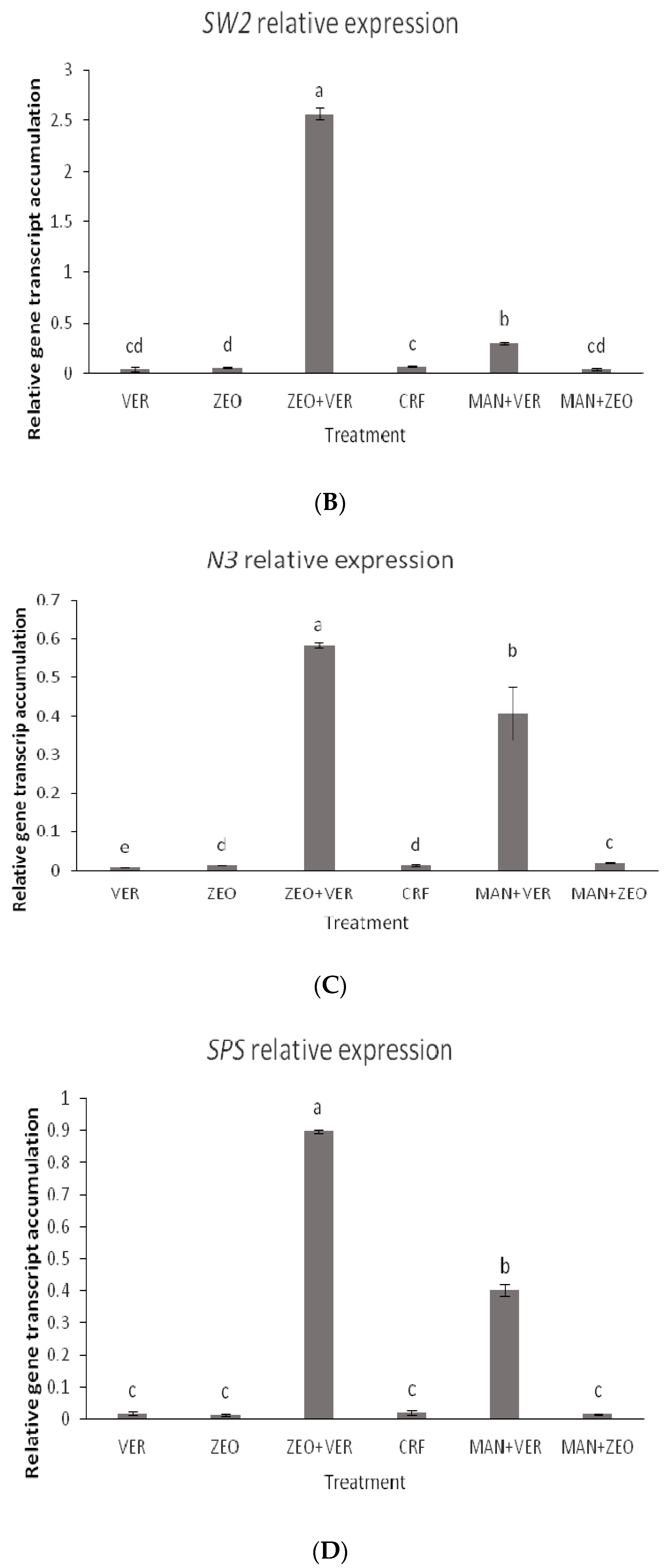
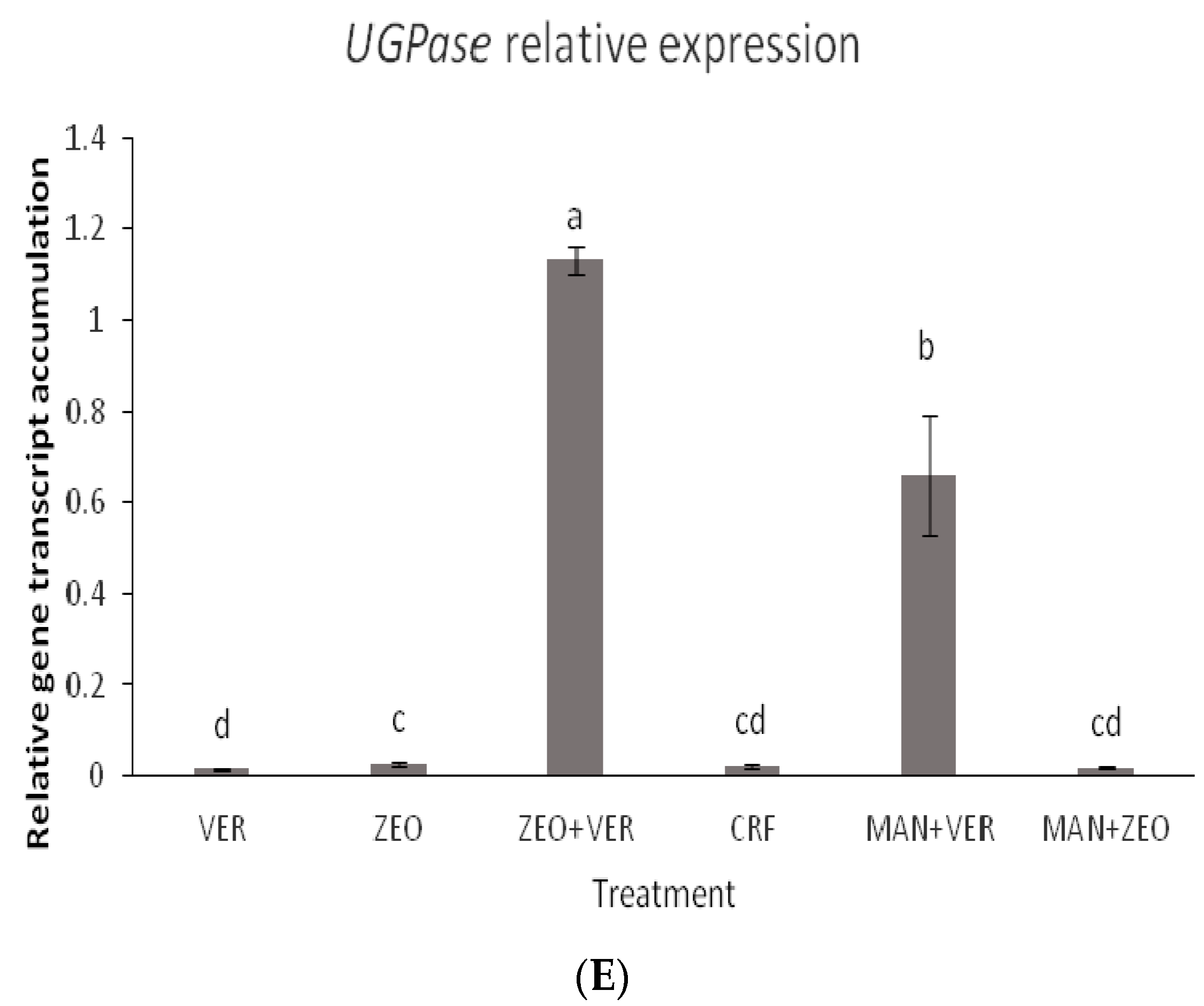
48], via a mechanism that is not yet fully elucidated, the coregulation of the expression of the sugar transport, the sucrose synthesis and the K transporter is not surprising. In our study, the transcript accumulation of all studied genes peaked in the ZEO + VER and MAN + ZEO treatments, while minimal differences were observed between the other treatments (
Figure 6A–E). Intriguingly, while a specific clear pattern between the K levels of leaves and fruits and the total soluble solids in fruits is absent, we suggest that the dynamics of the foliar K and sugar levels along with the ones of the sink organs (such as the fruits) may influence gene expression. The ZEO + VER treatment showed high foliar and low fruit K levels (
Table 3 and
Table 5, respectively), while the MAN + ZEO treatment exhibited rather low total K levels (
Table 3 and
Table 5) and high soluble sugar levels of fruits (
Figure 5A). The exact mechanism, however, triggering gene expression towards the facilitation of the sugar and K transport as well as the sucrose synthesis should be further investigated.
The total plant nutrient content showed significant differences among the CRF and other treatments (
Table 4). Since plant biomass did not significantly differ among the treatments (
Table 1), the differences determined in nutrient content should be ascribed to differences in nutrient uptake, which were quite distinct among the soil amendments (
Table 3). This is in contrast to the results of Chatzistathis et al. (2020) [
49], who studied the comparative effects of inorganic fertilization and manure application on olive plants’ response and concluded that the higher nutrient accumulation in the inorganic fertilization treatment should be ascribed to the higher plant biomass, rather than to a higher nutrient uptake. In another study with zeolite (ZEO), vermiculite (VER), combined zeolite and vermiculite (ZEO + VER) applications and fertilization effects with a controlled-release fertilizer (CRF) on
Castanea sativa L. plants [
17], it was found that the total per-plant uptake for most nutrients was significantly higher in the CRF- and ZEO-treated chestnut plants; these results of Chatzistathis et al. (2021) [
17] are partially in contrast to our current data for pepper plants (
Table 4). Τhe differences between the two studies may be possibly ascribed to the differential response of the two plant species used for the experimentation (a perennial plant species,
Castanea sativa Mill, and an annual one,
Capsicum annuum L.).
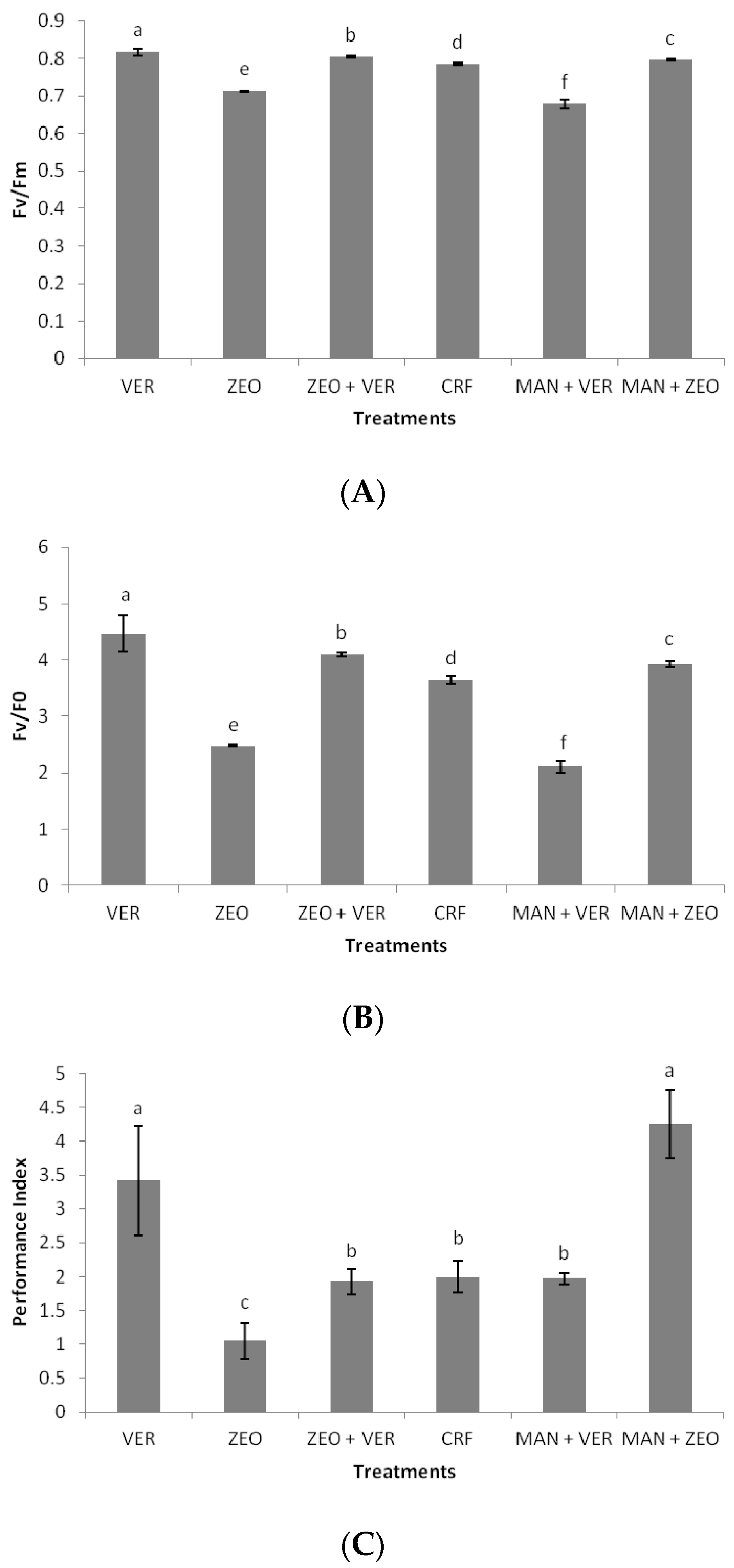
The maximum efficiency of PSII photochemistry (Fv/Fm), as well as the maximum efficiency of the water-splitting complex on the donor side of PSII (Fv/Fo), was recorded in the VER-treated plants, which was significantly higher than in all the other treatments (
Figure 3A,B); similar conclusions for the function of PSII were obtained in the studies of Chatzistathis et al. (2020) [
16] and Chatzistathis et al. (2021) [
17] for
Solanum lycopersicon L. and
Castanea sativa Mill plants, respectively. Although insignificant differences in photosynthetic rates were determined between CRF- and VER-treated plants (
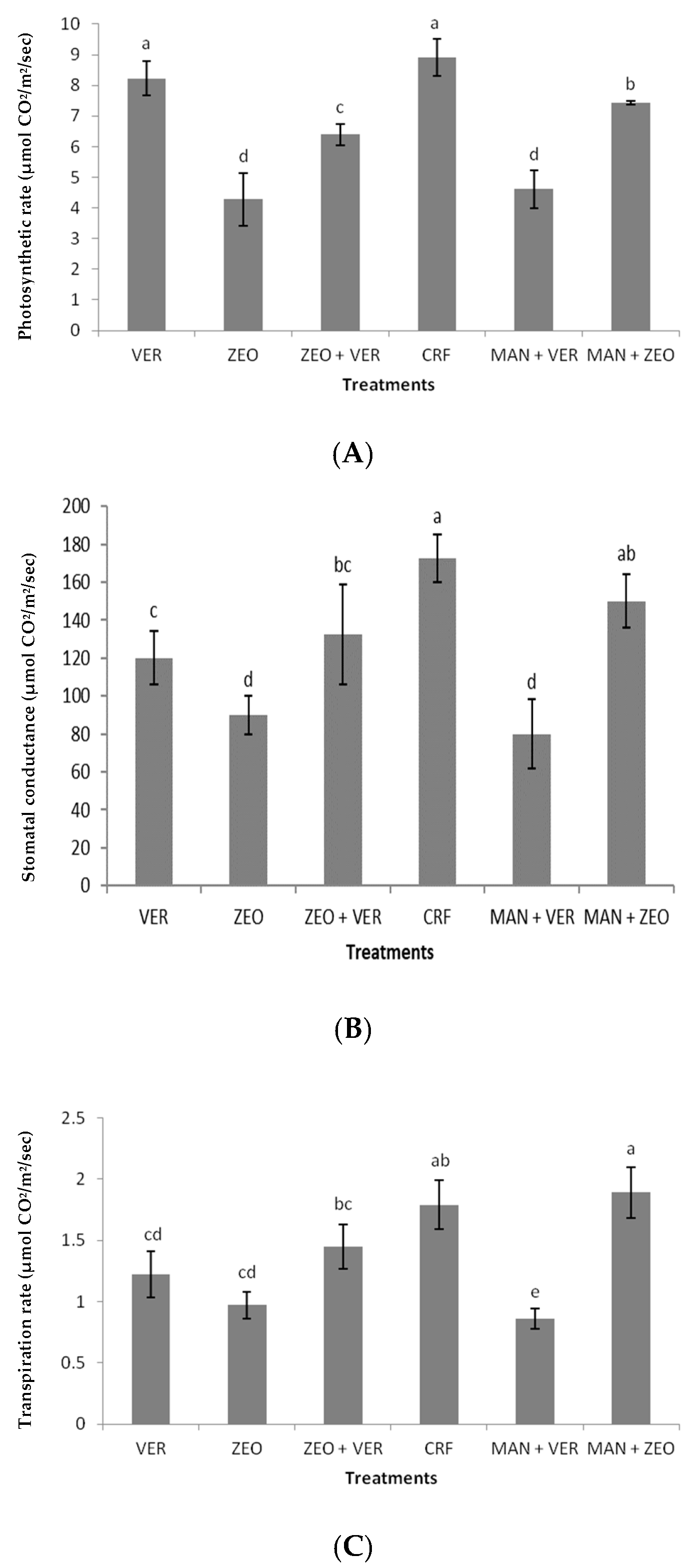
Figure 4A), a significant difference in stomata opening was found between the same treatments (
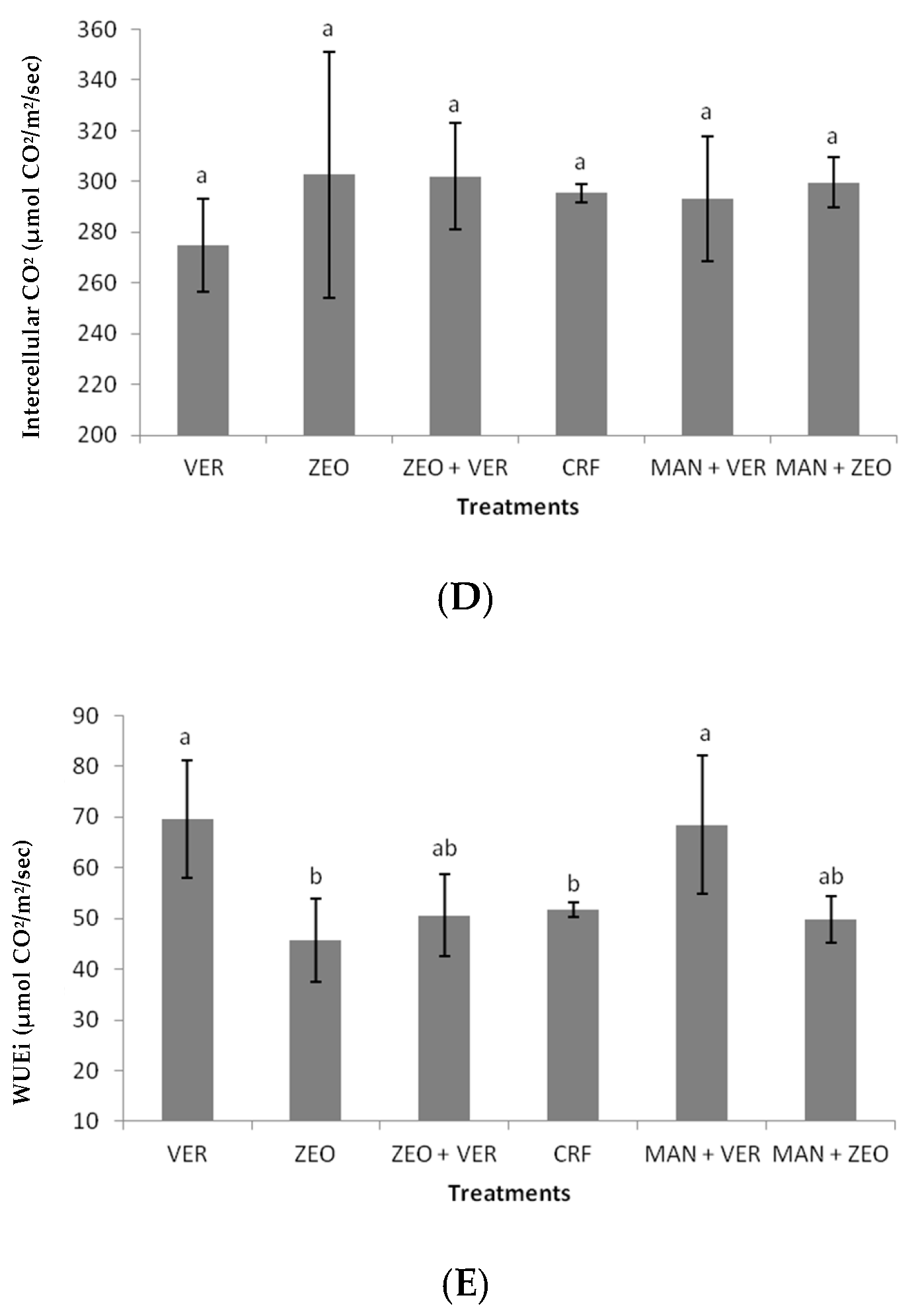
Figure 4B). This probably means that stomatal factors were responsible for the photosynthetic rate of CRF-treated plants, while nonstomatal factors (e.g., an increased activity of photosynthetic enzymes) were responsible for the photosynthetic rate of VER-treated plants. This is also confirmed by the statistically insignificant differences in intercellular CO
2 concentrations among the treatments (
Figure 4D). Additionally, the ZEO-treated plants exhibited a lower efficiency of the water-splitting complex on the donor side of PSII, as indicated by the decreased F
v/F
0 ratio. In our experiment, the decreased F
v/F
0 ratio in the ZEO-treated plants revealed a lower efficiency of the water-splitting complex on the donor side of PSII. The donor side photoinhibition mechanism elucidated photoinhibition by the malfunction of the water-splitting complex. If the water-splitting complex does not properly reduce the primary electron donor P680+, then P680+ might cause harmful oxidations in PSII. Finally, intrinsic water use efficiency (WUEi), which provides information on how efficiently water is used by plants, was significantly higher in the VER- and (MAN + VER)-treated plants, compared to ZEO- and CRF-treated plants. This proves that inorganic (ZEO, VER) and organic (MAN) soil amendments affected WUEi. Other researchers found that WUEi was influenced by K application, N fertilization, arbuscular mycorrhiza fungi (AMF) inoculation and this kind of organic fertilization [
49,
50,
51,
52].
5. Conclusions
Zeolite, vermiculite and goat manure application influenced soil fertility. In addition, significant differences in nutrient uptake among CRF and the other treatments were determined, since significant differences in foliar nutrient concentrations were recorded. Although for N, only inorganic fertilization (CRF) satisfied the N nutritional needs of pepper plants (leaf N: 3.84% D.W.), for the K uptake, zeolite (in its independent application, or in coapplication with vermiculite) showed very promising results (foliar K concentrations varied between 4.51% D.W. and 5.10% D.W.). A similar image was also obtained for fruit K concentrations, since the maximum K concentration was determined in the ZEO treatment (4.27% D.W.). Although K influences the qualitative characteristics of fruits, no clear conclusions were obtained in our study from the total soluble solids, total phenols and antioxidant activity (FRAP) of pepper fruits; thus, further research studies will be needed in the near future, under field conditions, to investigate the effect of K nutrition on pepper fruit quality. However, the application of soil amendments affected fruit quality characteristics, which is also supported by the results of other researchers. Plant biomass did not significantly differ among CRF and the other treatments; the same also happened for mean fruit weight. The optimum photosystem II (PSII) activity was recorded in the VER-treated plants or in the MAN + ZEO treatment, while the maximum photosynthetic rates were recorded both in VER- and CRF-treated plants. In the first case, it seemed that nonstomatal factors (e.g., an increased activity of photosynthetic enzymes) affected the photosynthetic activity of pepper plants, while in the second case, the CRF caused higher stomata opening. Finally, the intercellular CO2 concentration remained statistically unaffected among the treatments, while WUEi was affected by amendments’ application. It is believed that these data will constitute the first step towards revealing the effects of alternative sources of nutrients’ application on pepper plants’ nutrition, productivity and fruit quality. In addition, it is expected that our results will contribute towards a more sustainable approach of nutrient management for the Capsicum annuum L. crop. Of course, more research, under field conditions, will be necessary to draw more stable conclusions in comparative studies between inorganic fertilization and soil amendments’ application.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}