
Ethylenediurea Reduces Grain Nitrogen but Enhances Protein and Carbon Yield in Rice Cultivars

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
3. Results
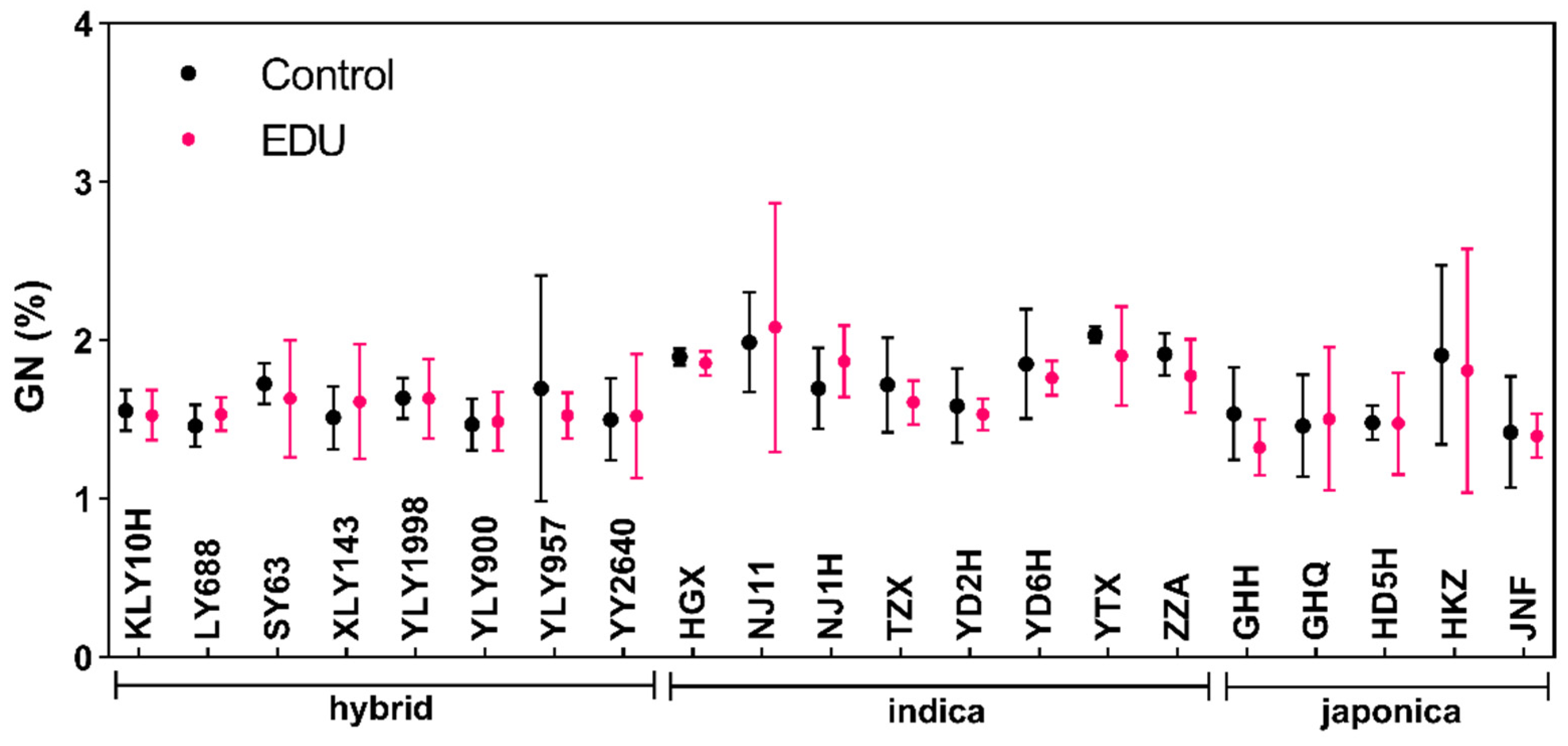
3.1. EDU Reduced Grain Nitrogen but Increased Grain Protein in Rice Cultivars
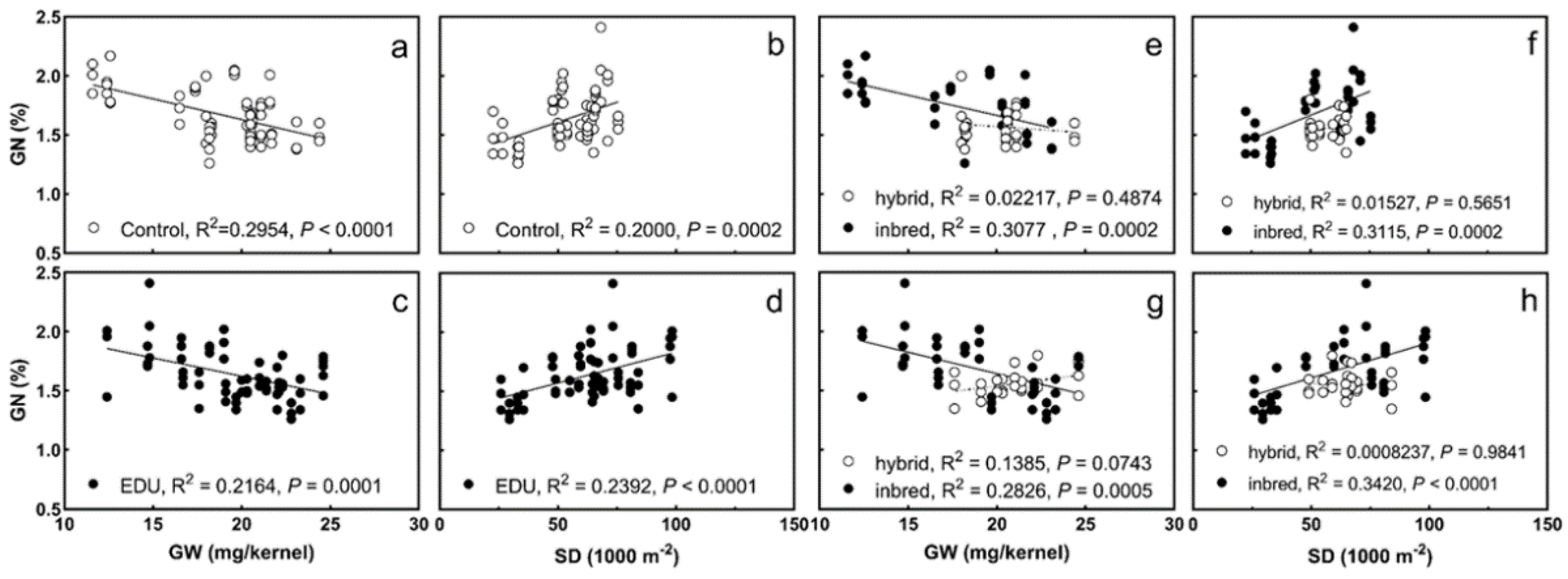
3.2. GN Significantly Correlates with Grain Number and Grain Weight
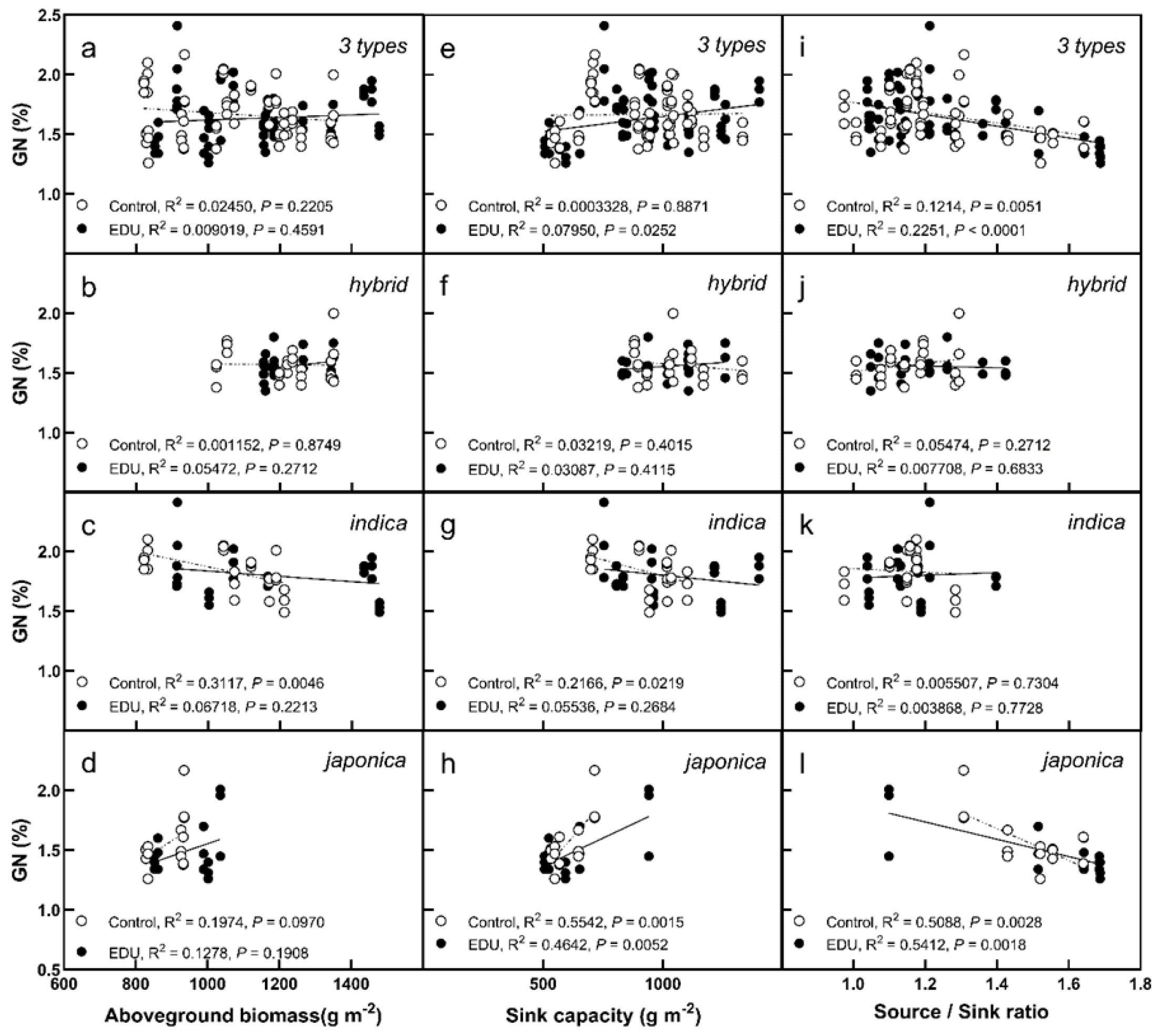
3.3. GN Changes with Sink and Source
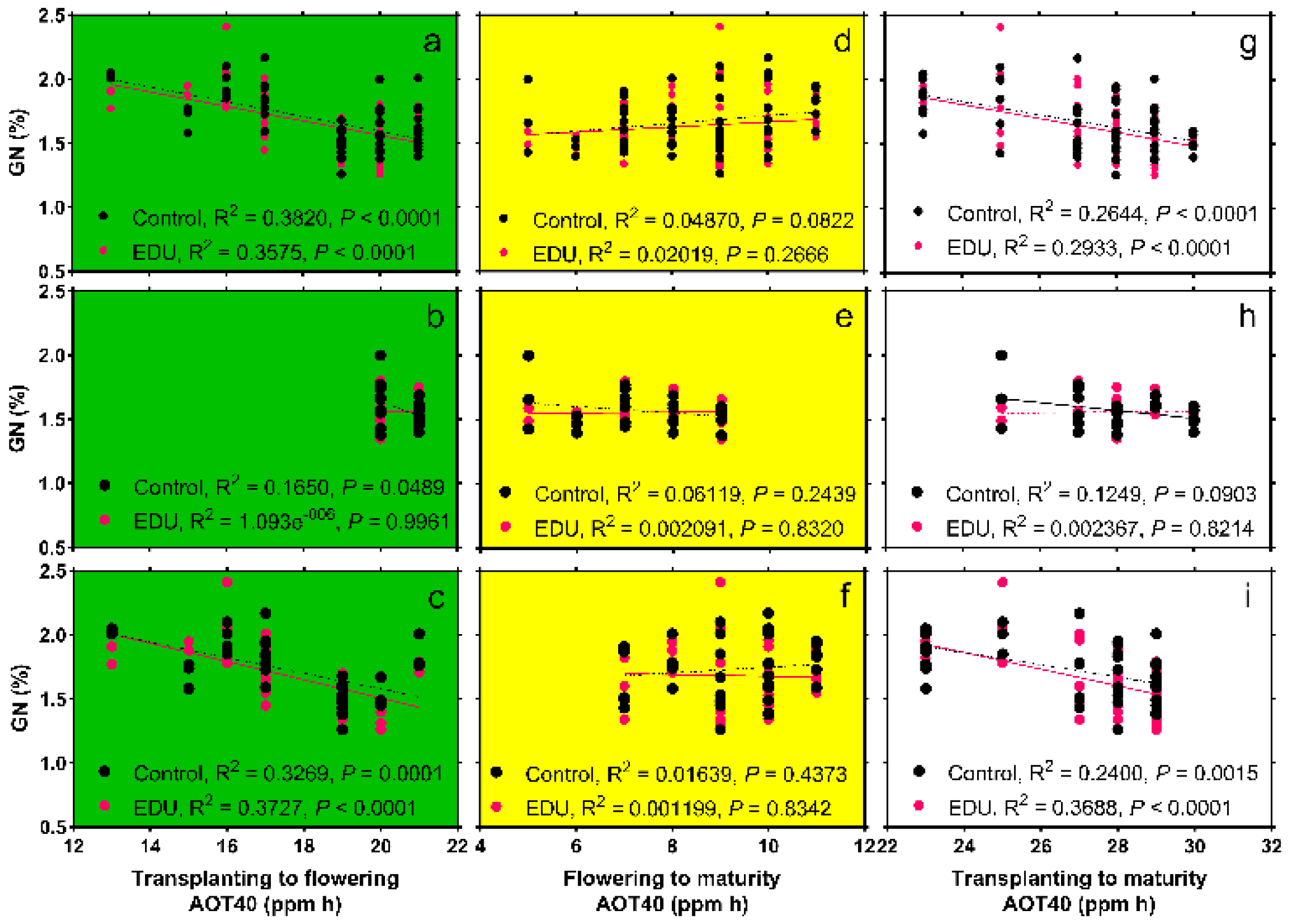
3.4. GN Is Negatively Correlated with the Concentration of Surface O3
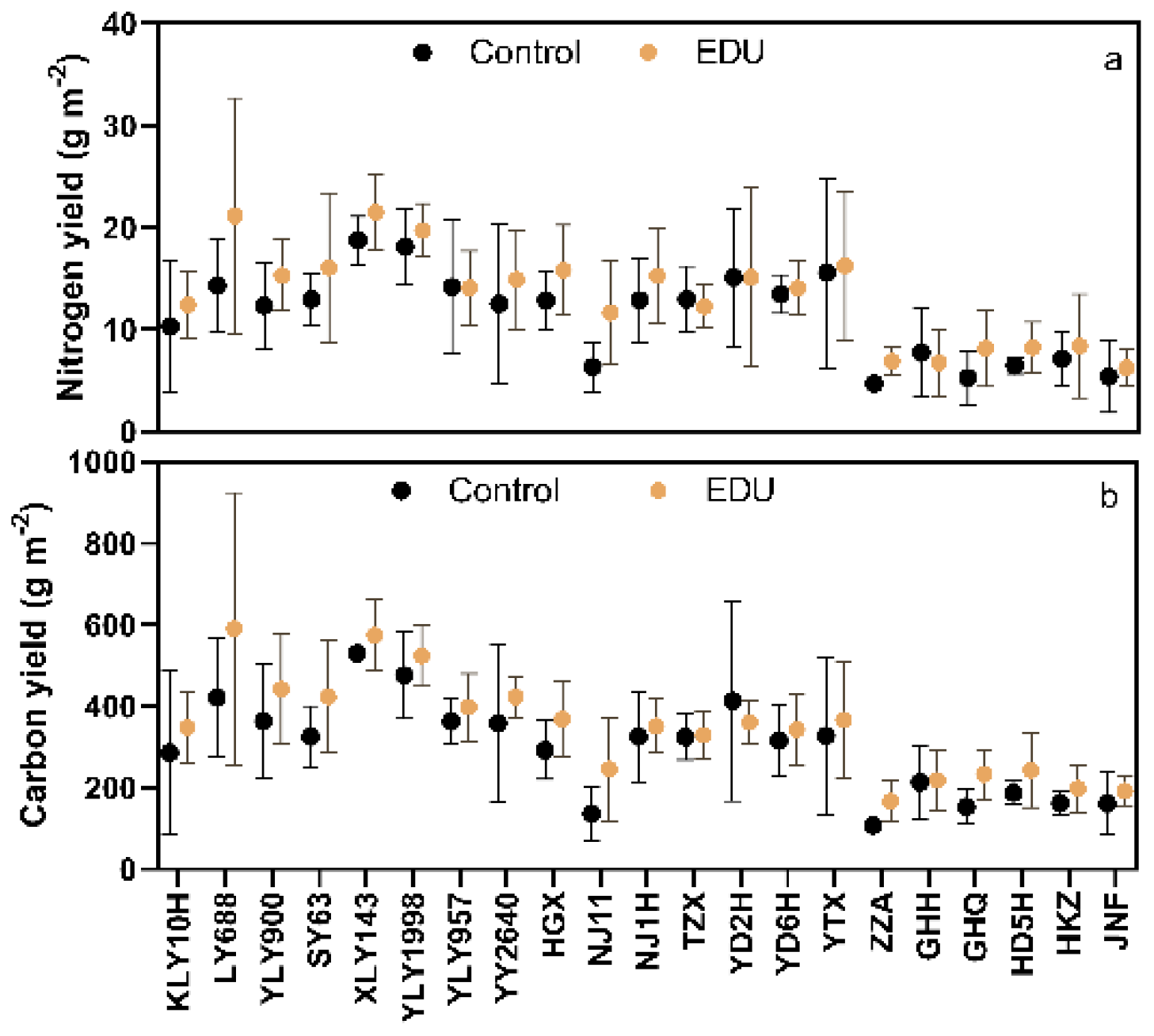
3.5. EDU Increased Nitrogen and Carbon Yield
4. Discussion
4.1. EDU Reduces Grain Nitrogen but Increases Grain Protein
4.2. Mechanisms for the Grain Nitrogen Change to EDU and Remaining Uncertainties
4.2.1. EDU Protection against O3-Induced Accelerated Senescence
4.2.2. Opposite to Concentration Effect Induced by O3 Phytotoxicity, EDU Reduces GN by Dilution
4.2.3. Source and Sink Related GN Changes
4.3. Imbalanced Increase in Nitrogen and Carbon Yield by EDU
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chuwah, C.; van Noije, T.; van Vuuren, D.P.; Stehfest, E.; Hazeleger, W. Global impacts of surface ozone changes on crop yields and land use. Atmos. Environ. 2015, 106, 11–23. [Google Scholar] [CrossRef]
- Feng, Z.; Xu, Y.; Kobayashi, K.; Dai, L.; Zhang, T.; Agathokleous, E.; Calatayud, V.; Paoletti, E.; Mukherjee, A.; Agrawal, M.; et al. Ozone pollution threatens the production of major staple crops in East Asia. Nat. Food 2022, 3, 47–56. [Google Scholar] [CrossRef]
- Li, X.B.; Wang, D.S.; Lu, Q.C.; Peng, Z.R.; Lu, S.J.; Li, B.; Li, C. Three-dimensional investigation of ozone pollution in the lower troposphere using an unmanned aerial vehicle platform. Environ. Pollut. 2017, 224, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, Y.; Li, X. Spatiotemporal distribution of ground-level ozone in China at a city level. Sci. Rep. 2020, 10, 7229. [Google Scholar] [CrossRef]
- Lee, E.H.; Upadhyaya, A.; Agrawal, M.; Rowland, R.A. Mechanisms of ethylenediurea (EDU) induced ozone protection: Reexamination of free radical scavenger systems in snap bean exposed to O3. Environ. Exp. Bot. 1997, 38, 199–209. [Google Scholar] [CrossRef]
- Agathokleous, E.; Paoletti, E.; Saitanis, C.J.; Manning, W.J.; Shi, C.; Koike, T. High doses of ethylene diurea (EDU) are not toxic to willow and act as nitrogen fertilizer. Sci. Total Environ. 2016, 566–567, 841–850. [Google Scholar] [CrossRef]
- Deng, F.; Wang, L.; Mei, X.F.; Li, S.X.; Pu, S.L.; Li, Q.P.; Ren, W.J. Polyaspartic acid (PASP)-urea and optimised nitrogen management increase the grain nitrogen concentration of rice. Sci. Rep. 2019, 9, 313. [Google Scholar] [CrossRef]
- Zhang, G.; Kobayashi, K.; Wu, H.; Shang, B.; Wu, R.; Zhang, Z.; Feng, Z. Ethylenediurea (EDU) protects inbred but not hybrid cultivars of rice from yield losses due to surface ozone. Environ. Sci. Pollut. Res. Int. 2021, 28, 68946–68956. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Han, Y.; Zhu, J.; Kobayashi, K.; Tang, H.; Wang, Y. The impact of elevated tropospheric ozone on grain quality of hybrid rice: A free-air gas concentration enrichment (FACE) experiment. Field Crops Res. 2012, 129, 81–89. [Google Scholar] [CrossRef]
- Shang, B.; Fu, R.; Agathokleous, E.; Dai, L.; Zhang, G.; Wu, R.; Feng, Z. Ethylenediurea offers moderate protection against ozone-induced rice yield loss under high ozone pollution. Sci. Total Environ. 2022, 806, 151341. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, W.; Jing, Q.; Cao, W.; Horie, T. Modeling grain protein formation in relation to nitrogen uptake and remobilization in rice plant. Front. Agric. China 2007, 1, 8–16. [Google Scholar] [CrossRef]
- De Vries, H. The rowwise correlation between two proximity matrices and the partial rowwise correlation. Psychometrika 1993, 58, 53–69. [Google Scholar] [CrossRef]
- Frei, M.; Kohno, Y.; Tietze, S.; Jekle, M.; Hussein, M.A.; Becker, T.; Becker, K. The response of rice grain quality to ozone exposure during growth depends on ozone level and genotype. Environ. Pollut. 2012, 163, 199–206. [Google Scholar] [CrossRef]
- Wang, Y.; Song, Q.; Frei, M.; Shao, Z.; Yang, L. Effects of elevated ozone, carbon dioxide, and the combination of both on the grain quality of Chinese hybrid rice. Environ. Pollut. 2014, 189, 9–17. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, J.; Wang, Y.; Peng, B.; Zhu, J.; Yang, L.; Wang, Y. Elevated tropospheric ozone increased grain protein and amino acid content of a hybrid rice without manipulation by planting density. J. Sci. Food Agric. 2015, 95, 72–78. [Google Scholar] [CrossRef]
- Shi, G.; Yang, L.; Wang, Y.; Kobayashi, K.; Zhu, J.; Tang, H.; Pan, S.; Chen, T.; Liu, G.; Wang, Y. Impact of elevated ozone concentration on yield of four Chinese rice cultivars under fully open-air field conditions. Agric. Ecosyst. Environ. 2009, 131, 178–184. [Google Scholar] [CrossRef]
- Pang, J.; Kobayashi, K.; Zhu, J. Yield and photosynthetic characteristics of flag leaves in Chinese rice (Oryza sativa L.) varieties subjected to free-air release of ozone. Agric. Ecosyst. Environ. 2009, 132, 203–211. [Google Scholar] [CrossRef]
- Gelang, J.; Selldén, G.; Younis, S.; Pleijel, H. Effects of ozone on biomass, non-structural carbohydrates and nitrogen in spring wheat with artificially manipulated source/sink ratio. Environ. Exp. Bot. 2001, 46, 155–169. [Google Scholar] [CrossRef]
- Pandey, A.K.; Majumder, B.; Keski-Saari, S.; Kontunen-Soppela, S.; Mishra, A.; Sahu, N.; Pandey, V.; Oksanen, E. Searching for common responsive parameters for ozone tolerance in 18 rice cultivars in India: Results from ethylenediurea studies. Sci. Total Environ. 2015, 532, 230–238. [Google Scholar] [CrossRef]
- Eckardt, N.A.; Pell, E.J. Effects of ethylenediurea (EDU) on ozone-induced acceleration of foliar senescence in potato (Solanum tuberosum L.). Environ. Pollut. 1996, 92, 299–306. [Google Scholar] [CrossRef]
- Gupta, S.K.; Sharma, M.; Majumder, B.; Maurya, V.K.; Deeba, F.; Zhang, J.-L.; Pandey, V. Effects of ethylenediurea (EDU) on regulatory proteins in two maize (Zea mays L.) varieties under high tropospheric ozone phytotoxicity. Plant Physiol. Biochem. 2020, 154, 675–688. [Google Scholar] [CrossRef]
- Shao, Z.; Zhang, Y.; Mu, H.; Wang, Y.; Wang, Y.; Yang, L. Ozone-induced reduction in rice yield is closely related to the response of spikelet density under ozone stress. Sci. Total Environ. 2020, 712, 136560. [Google Scholar] [CrossRef]
- Huang, M.; Yang, C.; Ji, Q.; Jiang, L.; Tan, J.; Li, Y. Tillering responses of rice to plant density and nitrogen rate in a subtropical environment of southern China. Field Crops Res. 2013, 149, 187–192. [Google Scholar]
- Zhou, W.; Lv, T.; Yang, Z.; Wang, T.; Fu, Y.; Chen, Y.; Hu, B.; Ren, W. Morphophysiological mechanism of rice yield increases in response to optimized nitrogen management. Sci. Rep. 2017, 7, 17226. [Google Scholar]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2017, 217, 35–53. [Google Scholar] [CrossRef]
- He, H.; Wang, Q.; Yang, K.; Niu, M.; Wang, L.; Wu, H.; You, C.; Ke, J.; Zhu, D.; Wu, L. Optimizing nitrogen distribution in source-sink organs at heading for high grain yield and high nitrogen use efficiency in Japonica rice. Agron. J. 2021, 113, 4832–4849. [Google Scholar] [CrossRef]
- Wei, H.; Meng, T.; Li, X.; Dai, Q.; Zhang, H.; Yin, X. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration. Field Crops Res. 2018, 215, 23–38. [Google Scholar]
- Pan, Y.; Guo, J.; Fan, L.; Ji, Y.; Liu, Z.; Wang, F.; Pu, Z.; Ling, N.; Shen, Q.; Guo, S. The source sink balance during the grain filling period facilitates rice production under organic fertilizer substitution. Eur. J. Agron. 2022, 134, 126468. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, J.; Cao, X. Grain sink strength may be related to the poor grain filling of indica-japonica rice (Oryza sativa) hybrids. Physiol. Plant. 2001, 112, 470–477. [Google Scholar] [CrossRef]
- Chen, X.; Chen, M.; Lin, G.; Yang, Y.; Yu, X.; Wu, Y.; Xiong, F. Structural development and physicochemical properties of starch in caryopsis of super rice with different types of panicle. BMC Plant Biol. 2019, 19, 482. [Google Scholar] [CrossRef]
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Heuvelink, E.; Marcelis, L.F. Quantifying the source-sink balance and carbohydrate content in three tomato cultivars. Front Plant Sci. 2015, 6, 416. [Google Scholar] [PubMed]
- Liu, E.; Zeng, S.; Zhu, S.; Liu, Y.; Wu, G.; Zhao, K.; Liu, X.; Liu, Q.; Dong, Z.; Dang, X.; et al. Favorable alleles of grain-filling rate1 increase the grain-filling rate and yield of rice. Plant Physiol. 2019, 181, 1207–1222. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Tu, B.; Yang, W.; Yuan, H.; Li, J.; Guo, L.; Zheng, L.; Chen, W.; Zhu, X.; Wang, Y.; et al. Mitochondria-associated pyruvate kinase complexes regulate grain filling in rice. Plant Physiol. 2020, 183, 1073–1087. [Google Scholar] [CrossRef]
- Zhang, G.; Sakai, H.; Tokida, T.; Usui, Y.; Zhu, C.; Nakamura, H.; Yoshimoto, M.; Fukuoka, M.; Kobayashi, K.; Hasegawa, T. The effects of free-air CO₂ enrichment (FACE) on carbon and nitrogen accumulation in grains of rice (Oryza sativa L.). J. Exp. Bot. 2013, 64, 3179–3188. [Google Scholar] [CrossRef]
- Zhang, G.; Sakai, H.; Usui, Y.; Tokida, T.; Nakamura, H.; Zhu, C.; Fukuoka, M.; Kobayashi, K.; Hasegawa, T. Grain growth of different rice cultivars under elevated CO2 concentrations affects yield and quality. Field Crops Res. 2015, 179, 72–80. [Google Scholar] [CrossRef]
- Chen, T.; Zhao, Y.; Zhang, W.; Yang, S.; Ye, Z.; Zhang, G. Variation of the light stable isotopes in the superior and inferior grains of rice (Oryza sativa L.) with different geographical origins. Food Chem. 2016, 209, 95–98. [Google Scholar] [CrossRef]
- Zhang, G. Elevated CO2 and positional variation in cereal grains. Crop Sci. 2021, 61, 3859–3860. [Google Scholar] [CrossRef]
- Ren, D.; Li, Y.; He, G.; Qian, Q. Multifloret spikelet improves rice yield. New Phytol. 2020, 225, 2301–2306. [Google Scholar] [CrossRef]
- Kim, D.M.; Lee, H.S.; Kwon, S.J.; Fabreag, M.E.; Kang, J.W.; Yun, Y.T.; Chung, C.T.; Ahn, S.N. High-density mapping of quantitative trait loci for grain-weight and spikelet number in rice. Rice 2014, 7, 14. [Google Scholar] [CrossRef]
- Ding, C.; You, J.; Chen, L.; Wang, S.; Ding, Y. Nitrogen fertilizer increases spikelet number per panicle by enhancing cytokinin synthesis in rice. Plant Cell Rep. 2014, 33, 363–371. [Google Scholar] [CrossRef]
- Chaiwong, N.; Rerkasem, B.; Pusadee, T.; Prom-U-Thai, C. Silicon application improves caryopsis development and yield in rice. J. Sci. Food Agric. 2021, 101, 220–228. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Full Name | Type |
|---|---|---|
| GHH | Guihua Huang | japonica |
| GHQ | Guihua Qiu | japonica |
| HD5H | Huaidao 5 Hao | japonica |
| HKZ | Huangke Zao | japonica |
| JNF | Jinnan Feng | japonica |
| HGX | Huanggua Xian | indica |
| NJ11 | Nanjing 11 | indica |
| NJ1H | Nanjing 1 Hao | indica |
| TZX | Taizhong Xian | indica |
| YD2H | Yangdao 2 Hao | indica |
| YD6H | Yangdao 6 Hao | indica |
| YTX | Yintiao Xian | indica |
| ZZA | Zhenzhu Ai | indica |
| KLY10H | Keliangyou 10 Hao | two-line indica hybrid |
| LY688 | Liangyou 688 | two-line indica hybrid |
| SY63 | Shanyou 63 | three-line indica hybrid |
| XLY143 | Xiangliangyou 143 | two-line indica hybrid |
| YLY1998 | Yliangyou 1998 | two-line indica hybrid |
| YLY900 | Yliangyou 900 | two-line indica hybrid |
| YLY975 | Yliangyou 957 | two-line indica hybrid |
| YY2640 | Yongyou 2640 | three-line indica-japonica hybrid |
| Grain Nitrogen | Grain Carbon | Grain Protein | Nitrogen Yield | Carbon Yield | Protein Yield | ||
|---|---|---|---|---|---|---|---|
| Variable | p values | ||||||
| Treatment | 0.0028 | 0.6564 | 0.0093 | <0.0001 | <0.0001 | <0.0001 | |
| Type | <0.0001 | 0.8223 | 0.0553 | 0.0003 | 0.0003 | 0.0003 | |
| Cultivar | <0.0001 | 0.8572 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Treatment × Type | 0.0948 | 0.1428 | 0.9924 | 0.3585 | 0.4642 | 0.3585 | |
| Least square means by treatment | |||||||
| Treatment | % | % | g | g m−2 | g m−2 | g m−2 | |
| Control | 1.65 | 42.99 | 1.82 | 10.81 | 282.90 | 64.32 | |
| EDU | 1.59 | 43.01 | 1.91 | 12.74 | 339.25 | 75.83 | |
| EDU effect (%) | −3.81 | 0.06 | 4.90 | 17.89 | 19.92 | 17.89 | |
| Least square means by rice type | |||||||
| Type | % | % | g | g m−2 | g m−2 | g m−2 | |
| hybrid | 1.56 | 43.02 | 1.92 | 15.52 | 427.36 | 92.36 | |
| indica | 1.82 | 42.98 | 1.93 | 12.85 | 310.04 | 76.45 | |
| japonica | 1.53 | 43.00 | 1.76 | 6.96 | 195.82 | 41.41 | |
| Correlation R | GC | PD | SPP | SD | RS | GW | GY | SC | |
|---|---|---|---|---|---|---|---|---|---|
| Control | 3 types | 0.0278 | 0.5285 * | 0.1665 | 0.5183 * | −0.6641 ** | −0.6068 * | −0.2663 | 0.0743 |
| hybrid | 0.5312 | 0.3303 | −0.3083 | −0.0333 | −0.5221 | −0.2005 | −0.2948 | −0.1624 | |
| inbred | 0.0082 | 0.4305 | 0.6266 * | 0.7247 ** | −0.5807 * | −0.6097 * | 0.0952 | 0.4586 | |
| EDU | 3 types | 0.1082 | 0.4877 * | 0.2551 | 0.5721 ** | −0.6725 *** | −0.5422 * | −0.1090 | 0.3220 |
| hybrid | 0.5230 | 0.4840 | −0.4065 | −0.0733 | 0.2699 | 0.7339 * | 0.4957 | 0.4217 | |
| Inbred | −0.0379 | 0.4072 | 0.6995 ** | 0.6738 * | −0.7037 ** | −0.6109 * | 0.1104 | 0.4938 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Cao, R.; Risalat, H.; Hu, Q.; Pan, X.; Hu, Y.; Shang, B.; Wu, H.; Zhang, Z.; Feng, Z. Ethylenediurea Reduces Grain Nitrogen but Enhances Protein and Carbon Yield in Rice Cultivars. Agronomy 2022, 12, 1988. https://doi.org/10.3390/agronomy12091988
Zhang G, Cao R, Risalat H, Hu Q, Pan X, Hu Y, Shang B, Wu H, Zhang Z, Feng Z. Ethylenediurea Reduces Grain Nitrogen but Enhances Protein and Carbon Yield in Rice Cultivars. Agronomy. 2022; 12(9):1988. https://doi.org/10.3390/agronomy12091988
Chicago/Turabian StyleZhang, Guoyou, Rong Cao, Hamdulla Risalat, Qinan Hu, Xiaoya Pan, Yaxin Hu, Bo Shang, Hengchao Wu, Zujian Zhang, and Zhaozhong Feng. 2022. "Ethylenediurea Reduces Grain Nitrogen but Enhances Protein and Carbon Yield in Rice Cultivars" Agronomy 12, no. 9: 1988. https://doi.org/10.3390/agronomy12091988
APA StyleZhang, G., Cao, R., Risalat, H., Hu, Q., Pan, X., Hu, Y., Shang, B., Wu, H., Zhang, Z., & Feng, Z. (2022). Ethylenediurea Reduces Grain Nitrogen but Enhances Protein and Carbon Yield in Rice Cultivars. Agronomy, 12(9), 1988. https://doi.org/10.3390/agronomy12091988