
Species Choice Influences Weed Suppression, N Sharing and Crop Productivity in Oilseed Rape–Legume Intercrops


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
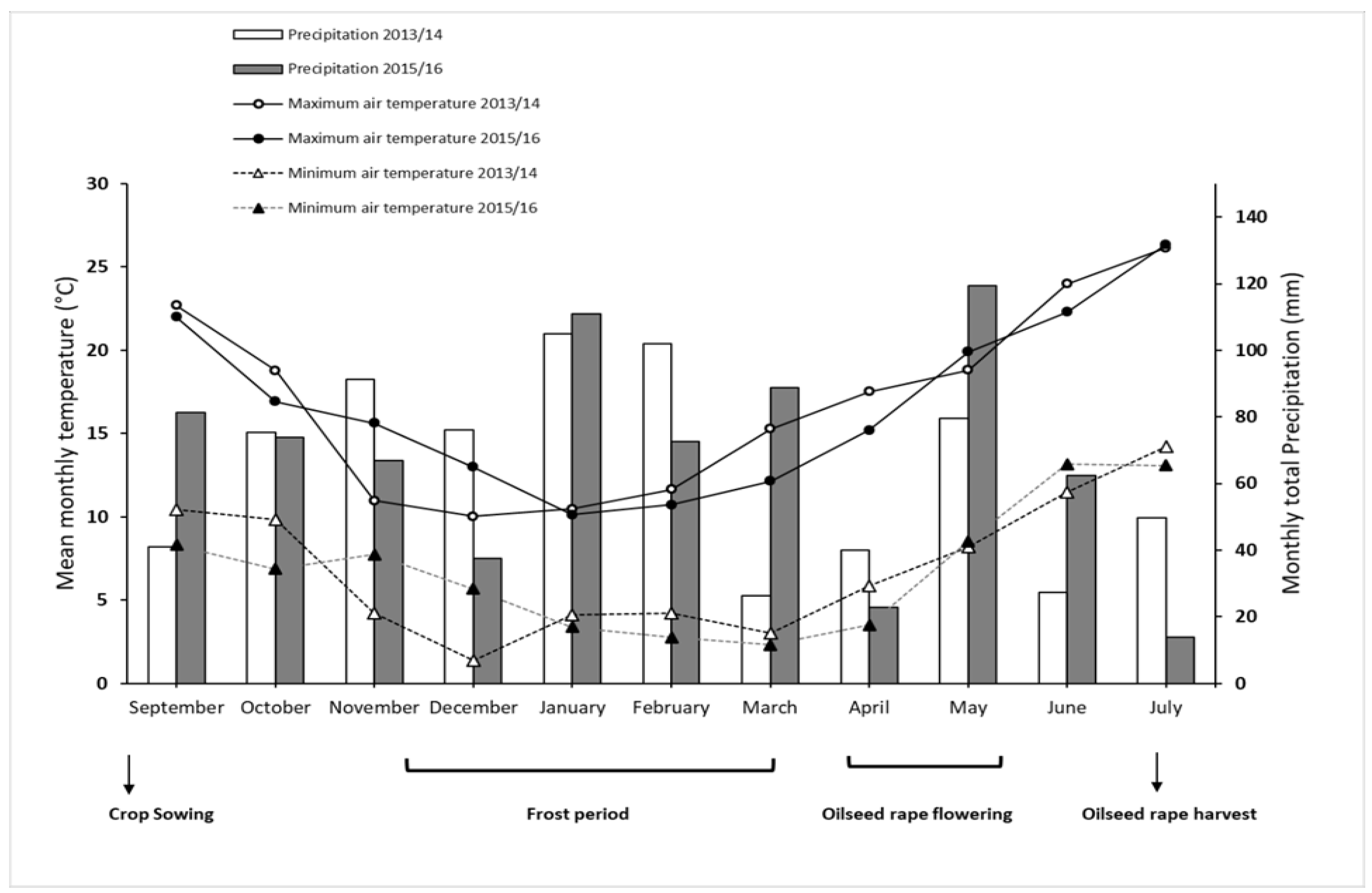
2.1. Site Characteristics
2.2. Experimental Design and Crop Management
2.3. Sampling Measurements and Analytical Methods
2.4. Calculations and Statistical Analysis
3. Results
3.1. Comparison between Three Sole Crops (Oilseed Rape and Legumes) Growth and Soil N Uptake Ability
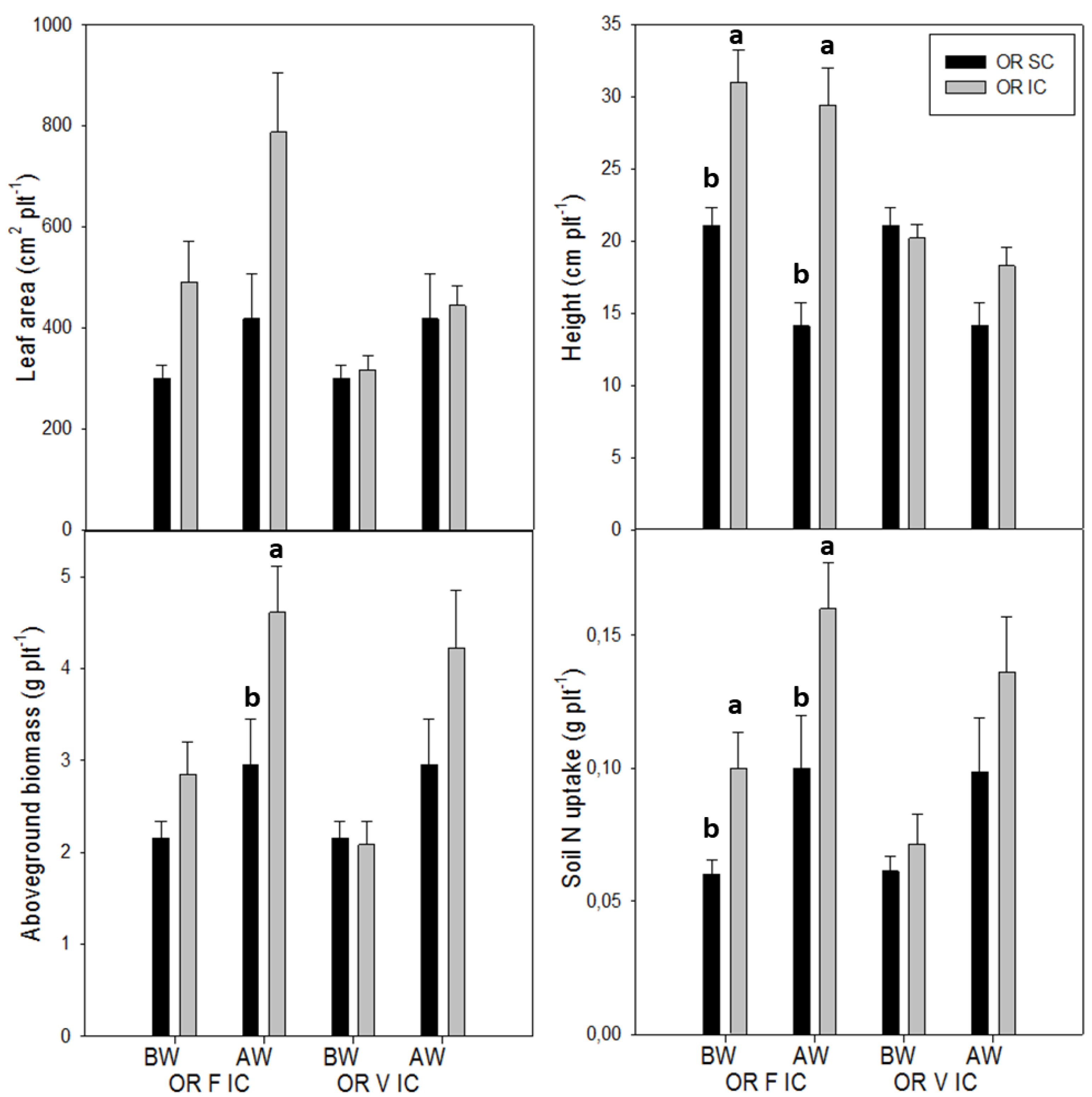
3.2. Effects of Intercropping on Oilseed Rape Growth and Productivity
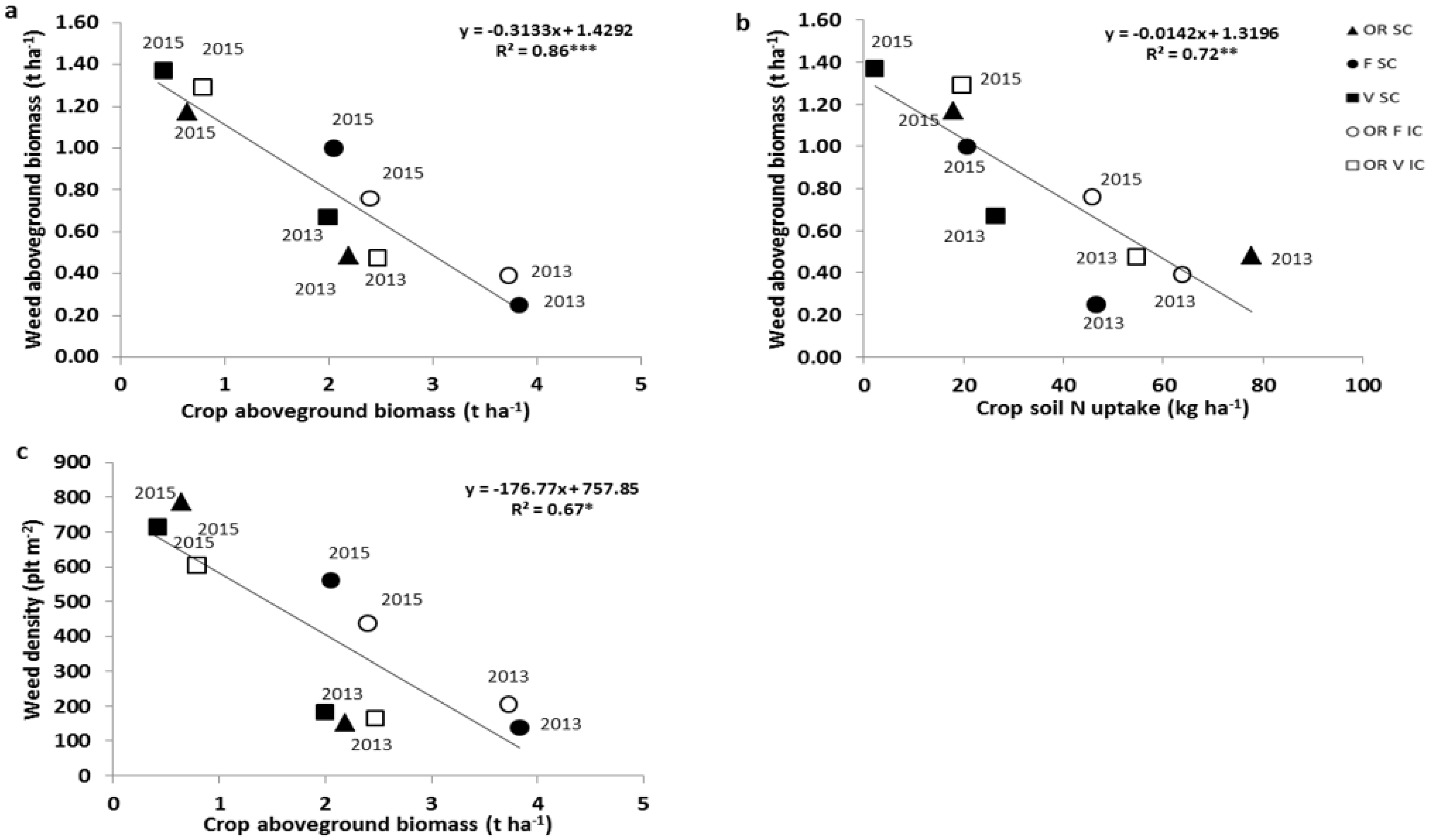
3.3. Effects of Intercropping on Weed Suppression
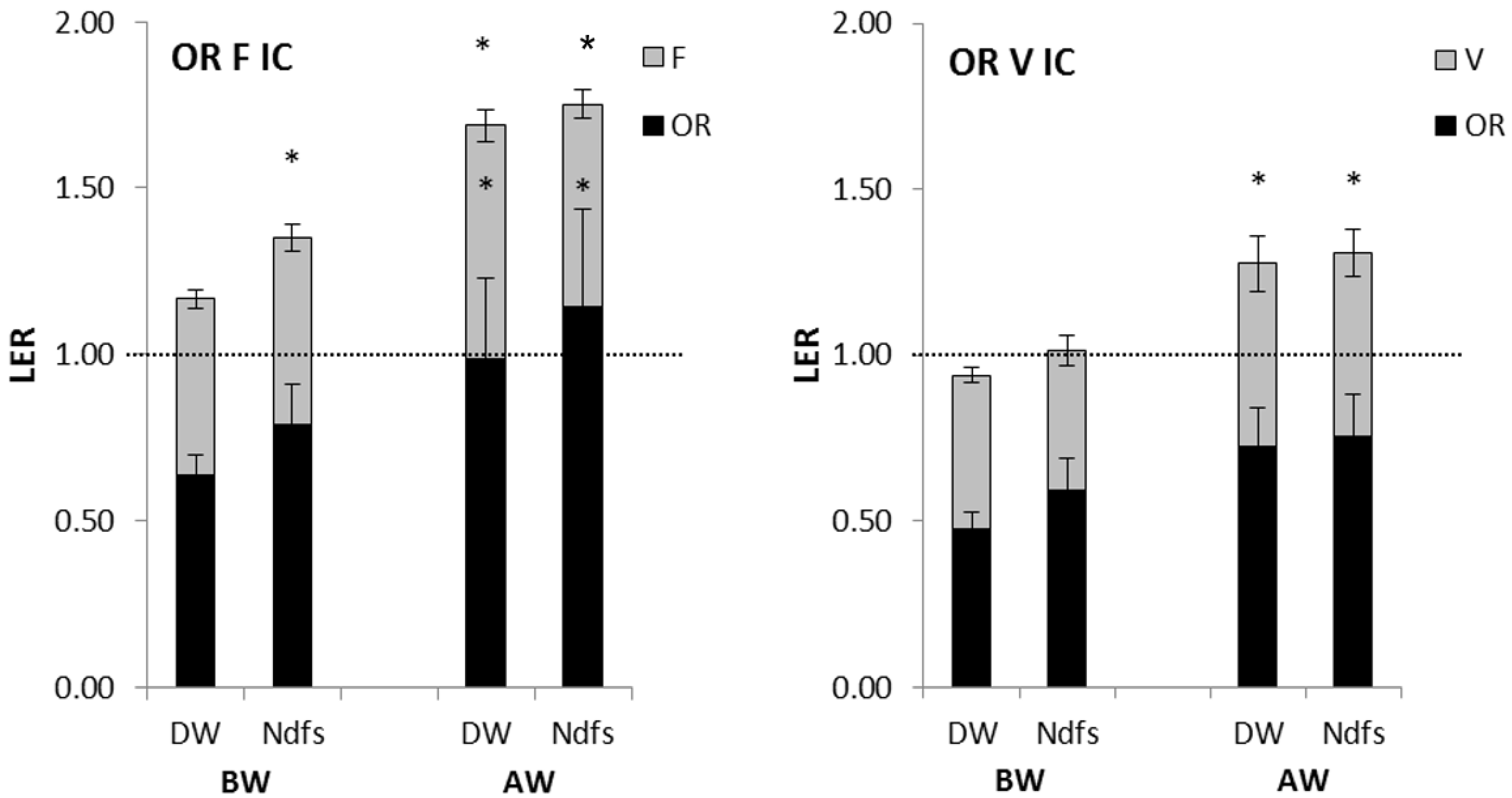
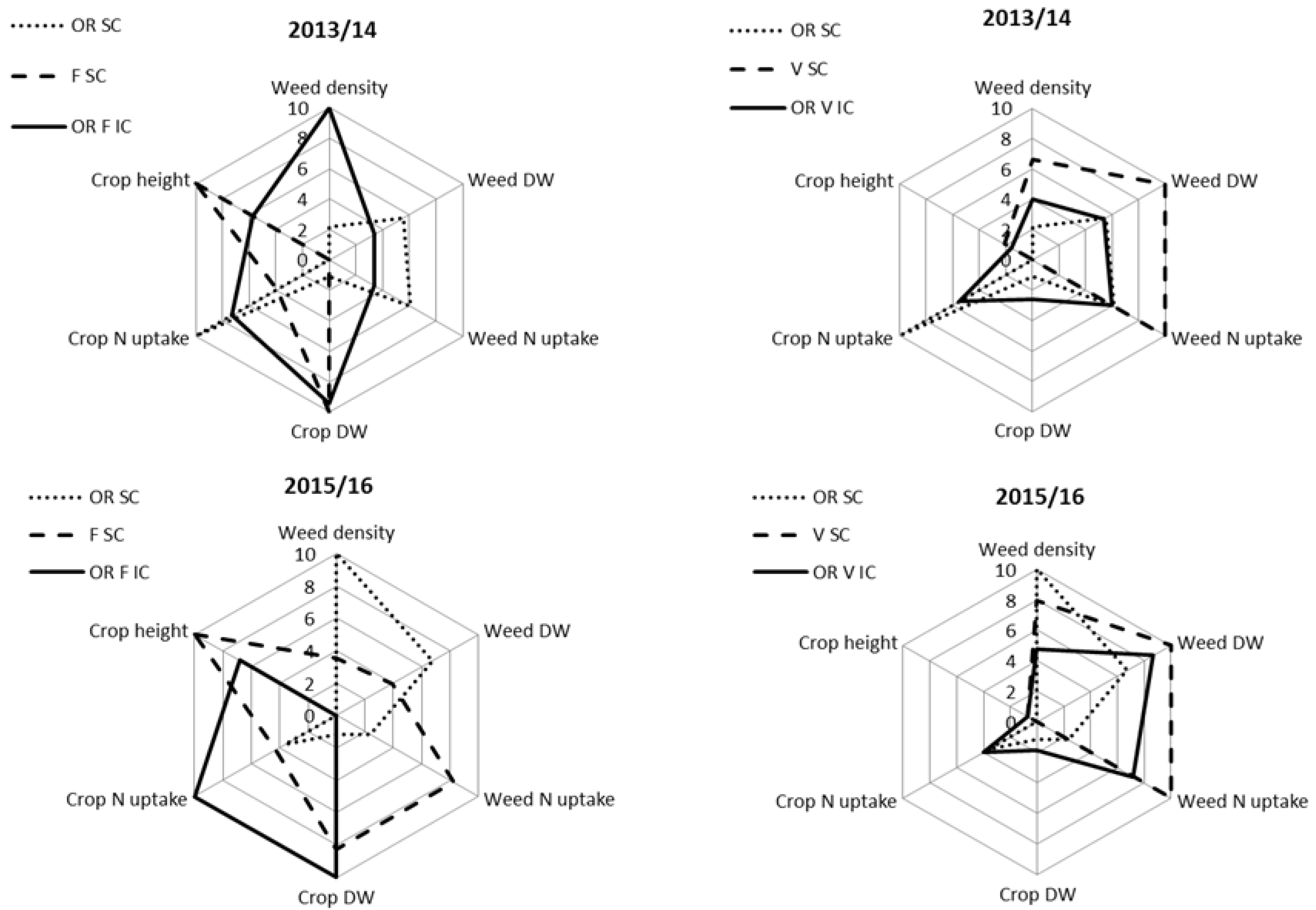
3.4. Competitive Abilities Profiles
3.5. Weed Diversity and Species Richness
3.6. Yield Components of Oilseed Rape
4. Discussion
4.1. Observed Complementarity between Intercropped Species
4.2. Contrasts between Species and Intercropping Effects on Weed Suppression
4.3. Competitiveness of the Intercrops against Weeds Compared to the Sole Crops
4.4. Effects of Intercropping on Crop Growth and Productivity: Which Benefits for Farmers?
4.5. Perspectives for Improving Design and Crop Management of Oilseed Rape–Legume Intercrops
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heap, I. Global Perspective of Herbicide-Resistant Weeds. Pest. Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Malagoli, P.; Laine, P.; Rossato, L.; Ourry, A. Dynamics of Nitrogen Uptake and Mobilization in Field-Grown Winter Oilseed Rape (Brassica napus) from Stem Extension to Harvest: I. Global N Flows between Vegetative and Reproductive Tissues in Relation to Leaf Fall and Their Residual N. Ann. Bot. 2005, 95, 853–861. [Google Scholar] [CrossRef]
- Lutman, P.J.W.; Dixon, F.L. The Competitive Effects of Volunteer Barley (Hordeum Vulgare) on the Growth of Oilseed Rape (Brassica napus). Ann. Appl. Biol. 1990, 117, 633–644. [Google Scholar] [CrossRef]
- Primot, S.; Valantin-Morison, M.; Makowski, D. Predicting the Risk of Weed Infestation in Winter Oilseed Rape Crops. Weed Res. 2006, 46, 22–33. [Google Scholar] [CrossRef]
- Valantin-Morison, M.; Meynard, J.M. Diagnosis of Limiting Factors of Organic Oilseed Rape Yield. A Survey of Farmers’ Fields. Agron. Sustain. Dev. 2008, 28, 527–539. [Google Scholar] [CrossRef]
- Beckie, H.J.; Johnson, E.N.; Blackshaw, R.E.; Gan, Y. Weed Suppression by Canola and Mustard Cultivars. Weed Technol. 2008, 22, 182–185. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Karley, A.J.; Newton, A.C.; Morcillo, L.; Brooker, R.W.; Schöb, C. A Trait-Based Approach to Crop-Weed Interactions. Eur. J. Agron. 2015, 70, 22–32. [Google Scholar] [CrossRef]
- Willey, R.W. Intercropping—Its Importance and Research Needs. 1.Competition and Yield Advantages. Field Crop Abstracts 1979, 32, 1–10. [Google Scholar]
- Gronle, A.; Böhm, H.; Heß, J. Effect of Intercropping Winter Peas of Differing Leaf Type and Time of Flowering on Annual Weed Infestation in Deep and Shallow Ploughed Soils and on Pea Pests. Landbauforschung-vTI Agric. For. Res. 2014, 64, 31–44. [Google Scholar] [CrossRef]
- Banik, P.; Midya, A.; Sarkar, B.K.; Ghose, S.S. Wheat and Chickpea Intercropping Systems in an Additive Series Experiment: Advantages and Weed Smothering. Eur. J. Agron. 2006, 24, 325–332. [Google Scholar] [CrossRef]
- Carr, P.M.; Gardner, J.C.; Schatz, B.G.; Zwinger, S.W.; Guldan, S.J. Grain Yield and Weed Biomass of a Wheat-Lentil Intercrop. Agron. J. 1995, 87, 574–579. [Google Scholar] [CrossRef]
- Carton, N.; Naudin, C.; Piva, G.; Corre-Hellou, G. Intercropping Winter Lupin and Triticale Increases Weed Suppression and Total Yield. Agriculture 2020, 10, 316. [Google Scholar] [CrossRef]
- Corre-Hellou, G.; Dibet, A.; Hauggaard-Nielsen, H.; Crozat, Y.; Gooding, M.; Ambus, P.; Dahlmann, C.; von Fragstein, P.; Pristeri, A.; Monti, M.; et al. The Competitive Ability of Pea-Barley Intercrops against Weeds and the Interactions with Crop Productivity and Soil N Availability. F. Crop. Res. 2011, 122, 264–272. [Google Scholar] [CrossRef]
- Liebman, M.; Dyck, E. Crop Rotation and Intercropping Strategies for Weed Management. Ecol. Appl. 1993, 3, 92–122. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Ambus, P.; Jensen, E.S. Interspecific Competition, N Use and Interference with Weeds in Pea-Barley Intercropping. Field Crop. Res. 2001, 70, 101–109. [Google Scholar] [CrossRef]
- Cadoux, S.; Sauzet, G.; Valantin-Morison, M.; Pontet, C.; Champolivier, L.; Robert, C.; Lieven, J.; Flénet, F.; Mangenot, O.; Fauvin, P.; et al. Intercropping Frost-Sensitive Legume Crops with Winter Oilseed Rape Reduces Weed Competition, Insect Damage, and Improves Nitrogen Use Efficiency. OCL Oilseeds Fats 2015, 22. [Google Scholar] [CrossRef]
- Lorin, M.; Jeuffroy, M.-H.; Butier, A.; Valantin-Morison, M. Undersowing Winter Oilseed Rape with Frost-Sensitive Legume Living Mulches to Improve Weed Control. Eur. J. Agron. 2015, 71, 96–105. [Google Scholar] [CrossRef]
- Gharineh, M.H.; Moradi-Telavat, M.R. Investigation of Ecological Relationship and Density Acceptance of Canola in Canola-Field Bean Intercropping. Asian J. Agric. Res. 2009, 1, 11–17. [Google Scholar] [CrossRef]
- Emery, S.E.; Anderson, P.; Carlsson, G.; Friberg, H.; Larsson, M.C.; Wallenhammar, A.-C.; Lundin, O. The Potential of Intercropping for Multifunctional Crop Protection in Oilseed Rape (Brassica napus L.). Front. Agron. 2021, 3, 782686. [Google Scholar] [CrossRef]
- Gewen, V. An Underground Revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef]
- Dowling, A.; Sadras, V.O.; Roberts, P.; Doolette, A.; Zhou, Y.; Denton, M.D. Legume-Oilseed Intercropping in Mechanised Broadacre Agriculture—A Review. Field Crop. Res. 2021, 260, 107980. [Google Scholar] [CrossRef]
- Andersen, M.K.; Hauggaard-Nielsen, H.; Ambus, P.; Jensen, E.S. Biomass Production, Symbiotic Nitrogen Fixation and Inorganic N Use in Dual and Tri-Component Annual Intercrops. Plant Soil 2004, 266, 273–287. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. The Effect of Nitrogen Catch Crop Species on the Nitrogen Nutrition of Succeeding Crops. Fertil. Res. 1994, 37, 227–234. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. Are Differences in Root Growth of Nitrogen Catch Crops Important for Their Ability to Reduce Soil Nitrate-N Content, and How Can This Be Measured? Plant Soil 2001, 230, 185–195. [Google Scholar] [CrossRef]
- Gregory, P.J. Growth and Functioning of Plant Roots. Russell’s Soil Conditions and Plant Growth. Longman Scientific and Technical; Longman Group UK Limited: Haelow, Essex, UK, 1988. [Google Scholar]
- Schröder, D.; Köpke, U. Faba Bean (Vicia faba L.) Intercropped with Oil Crops—A Strategy to Enhance Rooting Density and to Optimize Nitrogen Use and Grain Production? Field Crop. Res. 2012, 135, 74–81. [Google Scholar] [CrossRef]
- Dayoub, E.; Naudin, C.; Piva, G.; Shirtliffe, S.J.; Fustec, J.; Corre-Hellou, G. Traits Affecting Early Season Nitrogen Uptake in Nine Legume Species. Heliyon 2017, 3, e00244. [Google Scholar] [CrossRef] [PubMed]
- Szumigalski, A.R.; Van Acker, R.C. Nitrogen Yield and Land Use Efficiency in Annual Sole Crops and Intercrops. Agron. J. 2006, 98, 1030–1040. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global Inputs of Biological Nitrogen Fixation in Agricultural Systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Hamblin, A.P.; Hamblin, J. Root Characteristics of Some Temperate Legume Species and Varieties on Deep, Free-Draining Entisols. Aust. J. Agric. Res. 1985, 36, 63–72. [Google Scholar] [CrossRef]
- Liu, L.; Gan, Y.; Bueckert, R.; Van Rees, K. Rooting Systems of Oilseed and Pulse Crops. II: Vertical Distribution Patterns across the Soil Profile. F. Crop. Res. 2011, 122, 248–255. [Google Scholar] [CrossRef]
- Corre-Hellou, G.; Fustec, J.; Crozat, Y. Interspecific Competition for Soil N and Its Interaction with N2 Fixation, Leaf Expansion and Crop Growth in Pea-Barley Intercrops. Plant Soil 2006, 282, 195–208. [Google Scholar] [CrossRef]
- Jensen, E.S. Grain Yield, Symbiotic N2 Fixation and Interspecific Competition for Inorganic N in Pea-Barley Intercrops. Plant Soil 1996, 182, 25–38. [Google Scholar] [CrossRef]
- Bedoussac, L.; Journet, E.P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological Principles Underlying the Increase of Productivity Achieved by Cereal-Grain Legume Intercrops in Organic Farming. A Review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Gooding, M.; Ambus, P.; Corre-Hellou, G.; Crozat, Y.; Dahlmann, C.; Dibet, A.; Von Fragstein, P.; Pristeri, A.; Monti, M. Pea–Barley Intercropping for Efficient Symbiotic N2-Fixation, Soil N Acquisition and Use of Other Nutrients in European Organic Cropping Systems. Field Crop. Res. 2009, 113, 64–71. [Google Scholar] [CrossRef]
- Naudin, C.; Corre-Hellou, G.; Pineau, S.; Crozat, Y.; Jeuffroy, M.H. The Effect of Various Dynamics of N Availability on Winter Pea-Wheat Intercrops: Crop Growth, N Partitioning and Symbiotic N2 Fixation. Field Crop. Res. 2010, 119, 2–11. [Google Scholar] [CrossRef]
- Szumigalski, A.; Van Acker, R. Weed Suppression and Crop Production in Annual Intercrops. Weed Sci. 2005, 53, 813–825. [Google Scholar] [CrossRef]
- Banik, P.; Sasmal, T.; Ghosal, P.K.; Bagchi, D.K. Evaluation of Mustard (Brassica compestris var. Toria) and Legume Intercropping under 1: 1 and 2: 1 Row-replacement Series Systems. J. Agron. Crop Sci. 2000, 185, 9–14. [Google Scholar] [CrossRef]
- Blackshaw, R.E.; Brandt, R.N.; Janzen, H.H.; Entz, T.; Grant, C.A.; Derksen, D.A. Differential Response of Weed Species to Added Nitrogen. Weed Sci. 2003, 51, 532–539. [Google Scholar] [CrossRef]
- Moreau, D.; Milard, G.; Munier-Jolain, N. A Plant Nitrophily Index Based on Plant Leaf Area Response to Soil Nitrogen Availability. Agron. Sustain. Dev. 2013, 33, 809–815. [Google Scholar] [CrossRef]
- Poggio, S.L. Structure of Weed Communities Occurring in Monoculture and Intercropping of Field Pea and Barley. Agric. Ecosyst. Environ. 2005, 109, 48–58. [Google Scholar] [CrossRef]
- Saucke, H.; Ackermann, K. Weed Suppression in Mixed Cropped Grain Peas and False Flax (Camelina sativa). Weed Res. 2006, 46, 453–461. [Google Scholar] [CrossRef]
- Spitters, C.J.T.; Van Den Bergh, J.P. Competition between Crop and Weeds: A System Approach. In Biology and Ecology of Weeds; Springer: Dordrecht, The Netherlands, 1982; pp. 137–148. [Google Scholar] [CrossRef]
- Fried, G.; Cordeau, S.; Metay, A.; Kazakou, E. Relative Importance of Environmental Factors and Farming Practices in Shaping Weed Communities Structure and Composition in French Vineyards. Agric. Ecosyst. Environ. 2019, 275, 1–13. [Google Scholar] [CrossRef]
- Pyšek, P.; Lepš, J. Response of a Weed Community to Nitrogen Fertilization: A Multivariate Analysis. J. Veg. Sci. 1991, 2, 237–244. [Google Scholar] [CrossRef]
- Verret, V.; Gardarin, A.; Makowski, D.; Lorin, M.; Cadoux, S.; Butier, A.; Valantin-Morison, M. Assessment of the Benefits of Frost-Sensitive Companion Plants in Winter Rapeseed. Eur. J. Agron. 2017, 91, 93–103. [Google Scholar] [CrossRef]
- Gu, C.; Bastiaans, L.; Anten, N.P.R.; Makowski, D.; van der Werf, W. Annual Intercropping Suppresses Weeds: A Meta-Analysis. Agric. Ecosyst. Environ. 2021, 322, 107658. [Google Scholar] [CrossRef]
- Gu, C.; van der Werf, W.; Bastiaans, L. A Predictive Model for Weed Biomass in Annual Intercropping. Field Crop. Res. 2022, 277, 108388. [Google Scholar] [CrossRef]
- Li, C.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.; Li, H.; Zhang, F.; van der Werf, W. Syndromes of Production in Intercropping Impact Yield Gains. Nat. Plants 2020, 6, 653–660. [Google Scholar] [CrossRef]
- Cheriere, T.; Lorin, M.; Corre-Hellou, G. Species Choice and Spatial Arrangement in Soybean-Based Intercropping: Levers That Drive Yield and Weed Control. Field Crop. Res. 2020, 256, 107923. [Google Scholar] [CrossRef]
- Stomph, T.J.; Dordas, C.; Baranger, A.; de Rijk, J.; Dong, B.; Evers, J.; Gu, C.; Li, L.; Simon, J.; Jensen, E.S.; et al. Designing Intercrops for High Yield, Yield Stability and Efficient Use of Resources: Are There Principles? Adv. Agron. 2020, 160, 1–50. [Google Scholar] [CrossRef]
- Bousselin, X.; Cassagne, N.; Baux, A.; Valantin-Morison, M.; Herrera, J.M.; Lorin, M.; Hédan, M.; Fustec, J. Interactions between Plants and Plant-Soil in Functionally Complex Mixtures Including Grass Pea, Faba Bean and Niger, Intercropped with Oilseed Rape. Agronomy 2021, 11, 1493. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Kuai, J.; Ullah, S.; Fahad, S.; Zhou, G. Optimization of Nitrogen Rate and Planting Density for Improving Yield, Nitrogen Use Efficiency, and Lodging Resistance in Oilseed Rape. Front. Plant Sci. 2017, 8, 532. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Hauggaard-Nielsen, H. Faba Bean in Cropping Systems. Field Crop. Res. 2010, 115, 203–216. [Google Scholar] [CrossRef]
- Lagarde, F.; Champolivier, L. Le Raisonnement et Les Avancées Techniques Permettent de Réduire La Fertilisation Azotée: Le Cas de Farmstar-Colza®. Oléagineux Corps Gras Lipides 2006, 13, 384–387. [Google Scholar] [CrossRef]
- Parnaudeau, V.; Jeuffroy, M.H.; Machet, J.M.; Reau, R.; Bissuel, C.; Eveillard, P. Methods for Determining the Nitrogen Fertiliser Requirements of Some Major Arable Crops in France. In Proceedings of the Conference of the International Fertiliser Society, Cambridge, UK, 10 December 2009; pp. 1–26. [Google Scholar]
- JKI, O. Growth Stages of Mono-and Dicotyledonous Plants; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany; Braunschweig, Germany, 2010. [Google Scholar]
- Amarger, N.; Mariotti, A.; Mariotti, F.; Durr, J.C.; Bourguignon, C.; Lagacherie, B. Estimate of Symbiotically Fixed Nitrogen in Field Grown Soybeans Using Variations In15N Natural Abundance. Plant Soil 1979, 52, 269–280. [Google Scholar] [CrossRef]
- López-Bellido, F.J.; López-Bellido, R.J.; Redondo, R.; López-Bellido, L. B Value and Isotopic Fractionation in N2 Fixation by Chickpea (Cicer arietinum L.) and Faba Bean (Vicia faba L.). Plant Soil 2010, 337, 425–434. [Google Scholar] [CrossRef]
- Yoneyama, T.; Fujita, K.; Yoshida, T.; Matsumoto, T.; Kambayashi, I.; Yazaki, J. Variation in Natural Abundance of 15N among Plant Parts and in 15N/14N Fractionation during N2 Fixation in the Legume-Rhizobia Symbiotic System. Plant Cell Physiol. 1986, 27, 791–799. [Google Scholar] [CrossRef]
- De Wit, C.T.; Van den Bergh, J.P. Competition between Herbage Plants. J. Agric. Sci. 1965, 13, 212–221. [Google Scholar] [CrossRef]
- Willey, R.W.; Osiru, D.S.O. Studies on Mixtures of Dwarf Sorghum and Beans (Phaseolus vulgaris) with Particular Reference to Plant Population. J. Agric. Sci. 1972, 79, 531–540. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar] [CrossRef]
- RCore. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Fisher, R.A.; Yates, F. Statistical Tables for Biological, Agricultural Aad Medical Research; Oliver and Boyd: Edinburgh, UK, 1938; p. 90. [Google Scholar]
- Jamont, M.; Piva, G.; Fustec, J. Sharing N Resources in the Early Growth of Rapeseed Intercropped with Faba Bean: Does N Transfer Matter? Plant Soil 2013, 371, 641–653. [Google Scholar] [CrossRef]
- Ajal, J.; Kiær, L.P.; Pakeman, R.J.; Scherber, C.; Weih, M. Intercropping Drives Plant Phenotypic Plasticity and Changes in Functional Trait Space. Basic Appl. Ecol. 2022, 61, 41–52. [Google Scholar] [CrossRef]
- Carton, N.; Naudin, C.; Piva, G.; Baccar, R.; Corre-Hellou, G. Differences for Traits Associated with Early N Acquisition in a Grain Legume and Early Complementarity in Grain Legume-Triticale Mixtures. AoB Plants 2018, 10, ply001. [Google Scholar] [CrossRef]
- Waterer, J.G.; Vessey, J.K.; Stobbe, E.H.; Soper, R.J. Yield and Symbiotic Nitrogen Fixation in a Pea-Mustard Intercrop as Influenced by N Fertilizer Addition. Soil Biol. Biochem. 1994, 26, 447–453. [Google Scholar] [CrossRef]
- Raseduzzaman, M.; Jensen, E.S. Does Intercropping Enhance Yield Stability in Arable Crop Production? A Meta-Analysis. Eur. J. Agron. 2017, 91, 25–33. [Google Scholar] [CrossRef]
- Breitenmoser, S.; Steinger, T.; Baux, A.; Hiltpold, I. Intercropping Winter Oilseed Rape (Brassica napus L.) Has the Potential to Lessen the Impact of the Insect Pest Complex. Agronomy 2022, 12, 723. [Google Scholar] [CrossRef]
- Gardarin, A.; Celette, F.; Naudin, C.; Piva, G.; Valantin-Morison, M.; Vrignon-Brenas, S.; Verret, V.; Médiène, S. Intercropping with Service Crops Provides Multiple Services in Temperate Arable Systems: A Review. Agron. Sustain. Dev. 2022, 42, 39. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2013/14 | 2015/16 | Norms | Analysis Method * | ||
|---|---|---|---|---|---|
| Granulometric analysis | % Clay | 14.7 | 16.2 | Granulometric analysis after decarbonation (×31.107) | |
| % Silt | 22 | 20.3 | |||
| % Sand | 61.1 | 38.5 | |||
| Chemical analysis | pH (water) | 7.3 | 8.2 | Water extraction, “acidity active” (NF ISO 10390) | |
| CaCO3 Total (%) | <0.1 | 22.5 | (NF ISO 10693) | ||
| P2O5 (mg kg−1) | 176 | 60 | 20 to 70 | Olsen (NF ISO 11263) | |
| K2O (mg kg−1) | 466 | 392 | 80 to 150 | Extraction with ammonium acetate (NF × 31.108) | |
| MgO (mg kg−1) | 316 | 250 | 100 to 140 | Extraction with ammonium acetate (NF × 31.108) | |
| % Organic matter | 2.2 | 2.6 | 2.20 | Organic carbon × 1.72 (NF ISO 14235) | |
| % Carbon | 1.26 | 1.51 | 1.3 | Oxidizable soil organic matter determination | |
| % Total N | 0.17 | 0.17 | 0.15 | DUMAS (NF ISO 13878) |
| 2013/14 | 2015/16 | |||||||
|---|---|---|---|---|---|---|---|---|
| Days after Sowing | Growth Stage 1 | Days After Sowing | Growth Stage 1 | |||||
| Oilseed Rape | Faba Bean | Common Vetch | Oilseed Rape | Faba Bean | Common Vetch | |||
| 2013 | 2015 | |||||||
| Crop and weed biomass | 74 | Leaf development (16) | Formation of side shoots (21) | Stem elongation (34) | 85 | Leaf development (16) | Formation of side shoots (21) | Stem elongation (34) |
| 2014 | 2016 | |||||||
| Crop and weed biomass | 151 | Stem elongation (32) | Inflorescence emergence (55) | Inflorescence emergence (51) | 168 | Inflorescence emergence (51) | _ | _ |
| Oilseed rape yield | 286 | Ripening (81) | _ | _ | 294 | Ripening (81) | _ | _ |
| Before Winter | ||||
|---|---|---|---|---|
| OR SC | F SC | V SC | p-Value 1 | |
| Density (plt m−2) | 45.48 ± 1.57 b | 36.90 ± 1.80 c | 70.83 ± 4.69 a | 1.39 × 10−6 *** |
| Leaf area (cm2 plt−1) | 299.83 ± 25.95 b | 884.19 ± 37.92 a | 157.66 ± 16.32 c | 8.71 × 10−12 *** |
| Aboveground biomass (g plt−1) | 2.15 ± 0.19 b | 5.44 ± 0.16 a | 1.13 ± 0.07 c | 2.42 × 10−12 *** |
| Soil mineral N uptake (g plt−1) | 0.06 ± 0.01 a | 0.05 ± 0.00 a | 0.01 ± 0.00 b | 1.48 × 10−9 *** |
| Canopy height (cm) | 21.10 ± 1.25 b | 70.90 ± 4.41 a | 18.20 ± 0.97 b | 3.07 × 10−11 *** |
| After Winter | ||||
| Density (plt m−2) | 45.83 ± 3.10 ab | 35.71 ± 1.37 b | 48.57 ± 4.27 a | 3.58 × 10−2 * |
| Leaf area (cm2 plt−1) | 418.00 ± 90.08 | 739.30 ± 125.89 | 548.36 ± 18.07 | NS |
| Aboveground biomass (g plt−1) | 2.95 ± 0.50 b | 8.52 ± 1.27 a | 2.26 ± 0.41 b | 1.80 × 10−4 *** |
| Soil mineral N uptake (g plt−1) | 0.10 ± 0.02 a | 0.10 ± 0.02 a | 0.03 ± 0.01 b | 2 × 10−2 * |
| Canopy height (cm) | 14.13 ± 1.61 b | 66.73 ± 4.28 a | 21.60 ± 2.95 b | 5.65 × 10−9 *** |
| Weed Aboveground Biomass (t ha−1) | Weed Biomass in Total Aboveground Biomass (%) | Weed N (kg ha−1) | Weed N in Total Aboveground N (%) | Weed Density (plt m−2) | Weed Leaf Area Index (m2 m−2) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BW | AW | BW | AW | BW | AW | BW | AW | BW | AW | BW | AW | |
| 2013/14 | ||||||||||||
| OR SC | 0.20 ± 0.03 | 0.48 ± 0.08 ab | 13.80 ± 1.85 ab | 18.13 ± 3.03 ab | 6.28 ± 1.24 | 15.24 ± 2.65 ab | 14.62 ± 1.8 b | 16.26 ± 2.34 b | 224.29 ± 55.28 | 153.10 ± 43.92 | 0.31 ± 0.07 | 0.67 ± 0.15 |
| F SC | 0.17 ± 0.07 | 0.25 ± 0.08 b | 8.08 ± 3.79 b | 6.27 ± 2.19 c | 3.51 ± 1.27 | 6.97 ± 1.76 b | 16.82 ± 5.25 b | 13.99 ± 4.29 b | 228.57 ± 79.13 | 138.33 ± 34.79 | 0.40 ± 0.17 | 0.50 ± 0.15 |
| V SC | 0.30 ± 0.09 | 0.67 ± 0.11 a | 28.15 ± 6.67 a | 24.73 ± 3.16 a | 7.72 ± 1.93 | 20.52 ± 3.33 a | 49.97 ± 9.47 a | 42.69 ± 3.52 a | 182.38 ± 72.73 | 183.10 ± 43.02 | 0.47 ± 0.11 | 1.10 ± 0.21 |
| OR F IC | 0.23 ± 0.05 | 0.39 ± 0.06 ab | 12.32 ± 3.07 ab | 9.62 ± 1.76 bc | 6.17 ± 1.15 | 11.58 ± 1.68 ab | 16.72 ± 3.97 b | 15.11 ± 1.62 b | 168.33 ± 31.91 | 205.95 ± 40.28 | 0.33 ± 0.05 | 0.76 ± 0.13 |
| OR V IC | 0.31 ± 0.09 | 0.48 ± 0.06 ab | 24.94 ± 6.26 a | 16.01 ± 1.43 ab | 9.68 ± 3.16 | 15.11 ± 2.04 ab | 31.78 ± 9.36 ab | 21.32 ± 1.66 b | 204.76 ± 94.03 | 165.48 ± 33.16 | 0.71 ± 0.25 | 0.74 ± 0.11 |
| p-value 1 | NS | 4.33 × 10−2 * | 8.32 × 10−3 ** | 1.97 × 10−3 ** | NS | 3.70 × 10−2 * | 7.55 × 10−3 ** | 1.61 × 10−4 *** | NS | NS | NS | NS |
| 2015/16 | ||||||||||||
| OR SC | 0.77 ± 0.10 | 1.17 ± 0.10 ab | 51.16 ± 4.43 a | 64.41 ± 4.89 a | 22.05 ± 1.42 | 26.03 ± 2.41 ab | 51.58 ± 3.92 | 59.33 ± 5.47 b | 906.67 ± 185.63 | 786.90 ± 148.59 | 1.28 ± 0.14 | _ |
| F SC | 0.53 ± 0.04 | 1.00 ± 0.12 bc | 20.66 ± 1.66 c | 33.21 ± 4.24 b | 22.11 ± 2.36 | 33.37 ± 3.99 ab | 50.20 ± 2.76 | 61.03 ± 5.27 b | 585.00 ± 64.45 | 561.19 ± 177.17 | 1.34 ± 1.80 | _ |
| V SC | 0.57 ± 0.11 | 1.37 ± 0.10 a | 38.61 ± 7.28 abc | 76.30 ± 2.07 a | 20.85 ± 3.37 | 35.52 ± 2.00 a | 66.73 ± 6.48 | 94.04 ± 1.03 a | 719.76 ± 246.65 | 715.48 ± 235.56 | 1.19 ± 0.26 | _ |
| OR F IC | 0.62 ± 0.12 | 0.76 ± 0.06 c | 27.36 ± 4.49 bc | 25.46 ± 4.53 b | 24.77 ± 5.23 | 23.11 ± 0.96 b | 42.75 ± 5.4 | 35.83 ± 5.56 c | 695.48 ± 198.57 | 438.10 ± 111.11 | 1.53 ± 0.24 | _ |
| OR V IC | 0.71 ± 0.09 | 1.29 ± 0.08 a | 46.83 ± 4.07 ab | 62.50 ± 3.99 a | 23.09 ± 3.04 | 32.02 ± 3.04 ab | 54.83 ± 5.76 | 62.42 ± 3.07 b | 712.14 ± 98.53 | 603.81 ± 159.35 | 1.36 ± 0.17 | _ |
| p-value 1 | NS | 3.49 × 10−05 *** | 2.49 × 10−3 ** | 4.53 × 10−06 *** | NS | 1.35 × 10−2 * | NS | 5.55 × 10−05 *** | NS | NS | NS | |
| Before Winter | After Winter | |||||
|---|---|---|---|---|---|---|
| Treatment | H | S | D | H | S | D |
| OR SC | 1.52 ± 0.13 | 16.25 ± 0.93 | 0.65 ± 0.05 | 1.55 ± 0.19 | 14.25 ± 1.06 | 0.65 ± 0.06 |
| F SC | 1.70 ± 0.18 | 17.50 ± 0.56 | 0.68 ± 0.05 | 1.51 ± 0.12 | 13.25 ± 0.82 | 0.66 ± 0.05 |
| V SC | 1.66 ± 0.20 | 16.63 ± 1.13 | 0.68 ± 0.07 | 1.92 ± 0.46 | 15.13 ± 1.08 | 0.65 ± 0.06 |
| OR F IC | 1.64 ± 0.19 | 16.38 ± 0.73 | 0.65 ± 0.06 | 1.38 ± 0.15 | 14.13 ± 0.91 | 0.64 ± 0.05 |
| OR V IC | 1.66 ± 0.19 | 15.13 ± 1.10 | 0.69 ± 0.07 | 2.19 ± 0.52 | 14.63 ± 0.88 | 0.64 ± 0.06 |
| Treatment | Plant m−2 | Grain Yield (t ha−1) | Grain Yield (g plt−1) | Weight per 1000 Seed (g) | Number of Grains m−2 | Number of Grains per Plant |
|---|---|---|---|---|---|---|
| 2013/14 | ||||||
| OR SC | 37 ± 0.4 a | 1.82 ± 0.19 | 4.89 ± 0.51 | 4.40 ± 0.05 | 41187 ± 3938 | 1109 ± 106 |
| OR F IC | 15 ± 1.6 b | 1.17 ± 0.33 | 8.08 ± 2.82 | 4.66 ± 0.10 | 25371 ± 7567 | 1771 ± 654 |
| OR V IC | 21 ± 1.4 b | 1.51 ± 0.18 | 7.6 ± 1.41 | 4.58 ± 0.16 | 32747 ± 2960 | 1632 ± 246 |
| p-value 1 | 2.76 × 10−3 ** | NS | NS | NS | NS | NS |
| 2015/16 | ||||||
| OR SC | 49 ± 1.6 a | 1.57 ± 0.45 | 3.24 ± 0.99 b | 3.41 ± 0.06 | 45649 ± 12620 | 940 ± 275 b |
| OR F IC | 21 ± 0.8 b | 1.97 ± 0.16 | 9.39 ± 0.54 a | 3.42 ± 0.05 | 57515 ± 3999 | 2741 ± 138 a |
| OR V IC | 23 ± 1.1 b | 1.47 ± 0.23 | 6.28 ± 0.81 ab | 3.34 ± 0.05 | 43,865 ± 6345 | 1874 ± 225 ab |
| p-value 1 | 1.76 × 10−4 *** | NS | 4.92 × 10−2 * | NS | NS | 3.96 × 10−2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dayoub, E.; Piva, G.; Shirtliffe, S.J.; Fustec, J.; Corre-Hellou, G.; Naudin, C. Species Choice Influences Weed Suppression, N Sharing and Crop Productivity in Oilseed Rape–Legume Intercrops. Agronomy 2022, 12, 2187. https://doi.org/10.3390/agronomy12092187
Dayoub E, Piva G, Shirtliffe SJ, Fustec J, Corre-Hellou G, Naudin C. Species Choice Influences Weed Suppression, N Sharing and Crop Productivity in Oilseed Rape–Legume Intercrops. Agronomy. 2022; 12(9):2187. https://doi.org/10.3390/agronomy12092187
Chicago/Turabian StyleDayoub, Elana, Guillaume Piva, Steven J. Shirtliffe, Joëlle Fustec, Guénaëlle Corre-Hellou, and Christophe Naudin. 2022. "Species Choice Influences Weed Suppression, N Sharing and Crop Productivity in Oilseed Rape–Legume Intercrops" Agronomy 12, no. 9: 2187. https://doi.org/10.3390/agronomy12092187
APA StyleDayoub, E., Piva, G., Shirtliffe, S. J., Fustec, J., Corre-Hellou, G., & Naudin, C. (2022). Species Choice Influences Weed Suppression, N Sharing and Crop Productivity in Oilseed Rape–Legume Intercrops. Agronomy, 12(9), 2187. https://doi.org/10.3390/agronomy12092187