
Better Land and Nitrogen Complementarity for Green Forage Than for Silage in Barley–Field Bean Intercropping



Abstract
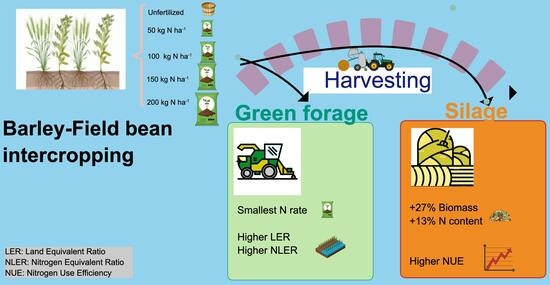
:
1. Introduction
2. Materials and Methods
2.1. Site Characteristics and Experimental Design
2.2. Crop Management
2.3. Sampling Procedures and Measurements
- LERB and LERF are the partial LERs of barley (B) and field bean (F);
- YBF is the dry matter production of the intercropped barley;
- YFB is the dry matter production of the intercropped field bean;
- YB and YF are the dry matter production of the barley and field bean sole crops, respectively.
- YBF, YFB, YB, and YF are the dry matter productions as defined above for the LER;
- ZBF and ZFB are the proportions of barley and field bean in the intercropping system, respectively;
- AB and AF are the barley’s and field bean’s aggressivity, respectively.
- CRB and CRF are the competitive ratios of barley (B) and field bean (F), respectively;
- ZBF and ZFB are the proportions of barley and field bean in the intercropping system, respectively.
- CbB and CbF are the barley and field bean’s competitive balance indices, respectively;
- NYBF, NYFB, NYB, and NYF have the same meaning as mentioned previously for the LER.
- NY is amount of N yield through forage (kg ha−1);
- FN is the rate of N fertilizer applied (kg ha−1).
2.4. Statistical Analysis
3. Results
3.1. Accumulation of Biomass
3.2. Nitrogen Concentration and Content
3.3. Intercropping Evaluation and Nitrogen Use Efficiency
4. Discussion
4.1. Accumulation of Biomass and Forage Quality
4.2. Accumulation of Nitrogen
4.3. Complementarity of Crops and Intercropping Performances
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Holman, J.D.; Obour, A.K.; Assefa, Y.; Johnson, S. Nutritive Value and Net Return of Forage Crop Rotations in the Central Great Plains. Agron. J. 2023, 115, 1399–1414. [Google Scholar] [CrossRef]
- Anil, L.; Park, J.; Phipps, R.H.; Miller, F.A. Temperate Intercropping of Cereals for Forage: A Review of the Potential for Growth and Utilization with Particular Reference to the UK. Grass Forage Sci. 1998, 53, 301–317. [Google Scholar] [CrossRef]
- Strydhorst, S.M.; King, J.R.; Lopetinsky, K.J.; Harker, K.N. Forage Potential of Intercropping Barley with Faba Bean, Lupin, or Field Pea. Agron. J. 2008, 100, 182–190. [Google Scholar] [CrossRef]
- Tabacco, E.; Comino, L.; Borreani, G. Production Efficiency, Costs and Environmental Impacts of Conventional and Dynamic Forage Systems for Dairy Farms in Italy. Eur. J. Agron. 2018, 99, 1–12. [Google Scholar] [CrossRef]
- Malézieux, E.; Beillouin, D.; Makowski, D. Feeding the World Better: Crop Diversification to Build Sustainable Food Systems. Perspective 2022, 58, 1–4. [Google Scholar] [CrossRef]
- Antichi, D.; Pampana, S.; Tramacere, L.G.; Biarnes, V.; Stute, I.; Kadžiulienė, Ž.; Howard, B.; Duarte, I.; Balodis, O.; Bertin, I.; et al. An Experimental Dataset on Yields of Pulses across Europe. Sci. Data 2023, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Willey, R.W. Intercropping—It’s Important and Research Needs. Part 1. Competition and Yield Advantages. Field Crop Abstr. 1979, 32, 1–10. [Google Scholar]
- Ghosh, P.K.; Manna, M.C.; Bandyopadhyay, K.K.; Ajay; Tripathi, A.K.; Wanjari, R.H.; Hati, K.M.; Misra, A.K.; Acharya, C.L.; Subba Rao, A. Interspecific Interaction and Nutrient Use in Soybean/Sorghum Intercropping System. Agron. J. 2006, 98, 1097–1108. [Google Scholar] [CrossRef]
- Xu, Z.; Li, C.; Zhang, C.; Yu, Y.; van der Werf, W.; Zhang, F. Intercropping Maize and Soybean Increases Efficiency of Land and Fertilizer Nitrogen Use; A Meta-Analysis. Field Crops Res. 2020, 246, 107661. [Google Scholar] [CrossRef]
- Monti, M.; Pellicanò, A.; Pristeri, A.; Badagliacca, G.; Preiti, G.; Gelsomino, A. Cereal/Grain Legume Intercropping in Rotation with Durum Wheat in Crop/Livestock Production Systems for Mediterranean Farming System. Field Crops Res. 2019, 240, 23–33. [Google Scholar] [CrossRef]
- Daryanto, S.; Fu, B.; Zhao, W.; Wang, S.; Jacinthe, P.-A.; Wang, L. Ecosystem Service Provision of Grain Legume and Cereal Intercropping in Africa. Agric. Syst. 2020, 178, 102761. [Google Scholar] [CrossRef]
- Stomph, T.; Dordas, C.; Baranger, A.; de Rijk, J.; Dong, B.; Evers, J.; Gu, C.; Li, L.; Simon, J.; Jensen, E.S. Designing Intercrops for High Yield, Yield Stability and Efficient Use of Resources: Are There Principles? Adv. Agron. 2020, 160, 1–50. [Google Scholar]
- Gu, C.; Bastiaans, L.; Anten, N.P.R.; Makowski, D.; van der Werf, W. Annual Intercropping Suppresses Weeds: A Meta-Analysis. Agric. Ecosyst. Environ. 2021, 322, 107658. [Google Scholar] [CrossRef]
- Pampana, S.; Arduini, I.; Andreuccetti, V.; Mariotti, M. Fine-Tuning N Fertilization for Forage and Grain Production of Barley–Field Bean Intercropping in Mediterranean Environments. Agronomy 2022, 12, 418. [Google Scholar] [CrossRef]
- Carr, P.M.; Martin, G.B.; Caton, J.S.; Poland, W.W. Forage and Nitrogen Yield of Barley—Pea and Oat—Pea Intercrops. Agron. J. 1998, 90, 79–84. [Google Scholar] [CrossRef]
- Carr, P.M.; Horsley, R.D.; Poland, W.W. Barley, Oat, and Cereal–Pea Mixtures as Dryland Forages in the Northern Great Plains. Agron. J. 2004, 96, 677–684. [Google Scholar] [CrossRef]
- Pappa, V.A.; Rees, R.M.; Walker, R.L.; Baddeley, J.A.; Watson, C.A. Legumes Intercropped with Spring Barley Contribute to Increased Biomass Production and Carry-over Effects. J. Agric. Sci. 2012, 150, 584–594. [Google Scholar] [CrossRef]
- Herbert, S.J.; Putnam, D.H.; Poos-Floyd, M.I.; Vargas, A.; Creighton, J.F. Forage Yield of Intercropped Corn and Soybean in Various Planting Patterns. Agron. J. 1984, 76, 507–510. [Google Scholar] [CrossRef]
- Sulc, R.M.; Tracy, B.F. Integrated Crop–Livestock Systems in the US Corn Belt. Agron. J. 2007, 99, 335–345. [Google Scholar] [CrossRef]
- Russelle, M.P.; Entz, M.H.; Franzluebbers, A.J. Reconsidering Integrated Crop–Livestock Systems in North America. Agron. J. 2007, 99, 325–334. [Google Scholar] [CrossRef]
- Hadjichristodoulou, A. Effect of Genotype and Rainfall on Yield and Quality of Forage Barley and Wheat Varieties in a Semi-Arid Region. J. Agric. Sci. 1976, 87, 489–497. [Google Scholar] [CrossRef]
- Sleugh, B.; Moore, K.J.; George, J.R.; Brummer, E.C. Binary Legume–Grass Mixtures Improve Forage Yield, Quality, and Seasonal Distribution. Agron. J. 2000, 92, 24–29. [Google Scholar] [CrossRef]
- Pirhofer-Walzl, K.; Rasmussen, J.; Jensen, H.H.; Eriksen, J.; Søegaard, K.; Rasmussen, J. Nitrogen transfer from forage legumes to nine neighbouring plants in a multi-species grassland. Plant Soil 2012, 350, 71–84. [Google Scholar] [CrossRef]
- Ghaley, B.B.; Hauggaard-Nielsen, H.; Høgh-Jensen, H.; Jensen, E.S. Intercropping of Wheat and Pea as Influenced by Nitrogen Fertilization. Nutr. Cycl. Agroecosyst. 2005, 73, 201–212. [Google Scholar] [CrossRef]
- Kalembasa, D.; Szukała, J.; Symanowicz, B.; Kalembasa, S.; Faligowska, A.; Becher, M. Amount of biologically nitrogen fixed by faba bean and its uptake by winter wheat determined by 15N ID method. Arch. Agron. Soil Sci. 2020, 67, 1875–1888. [Google Scholar] [CrossRef]
- van Kessel, C.; Hartley, C. Agricultural Management of Grain Legumes: Has It Led to an Increase in Nitrogen Fixation? Field Crops Res. 2000, 65, 165–181. [Google Scholar] [CrossRef]
- Peoples, M.B.; Bowman, A.M.; Gault, R.R.; Herridge, D.F.; McCallum, M.H.; McCormick, K.M.; Norton, R.M.; Rochester, I.J.; Scammell, G.J.; Schwenke, G.D. Factors Regulating the Contributions of Fixed Nitrogen by Pasture and Crop Legumes to Different Farming Systems of Eastern Australia. Plant Soil 2001, 228, 29–41. [Google Scholar] [CrossRef]
- Pampana, S.; Masoni, A.; Mariotti, M.; Ercoli, L.; Arduini, I. Nitrogen Fixation of Grain Legumes Differs in Response to Nitrogen Fertilisation. Exp. Agric. 2018, 54, 66–82. [Google Scholar] [CrossRef]
- Meyer, U. Growth Stages of Mono-and Dicotyledonous Plants—BBCH Monograph; Federal Biology Research Centre Agriculture Forestry: Berlin/Braunschweig, Germany, 2001. [Google Scholar]
- McGilchrist, C.A. Analysis of competition experiments. Biometrics 1965, 21, 975–985. [Google Scholar] [CrossRef]
- Willey, R.W.; Rao, M.R. A Competitive Ratio for Quantifying Competition between Intercrops. Exp. Agric. 1980, 16, 117–125. [Google Scholar] [CrossRef]
- Wilson, J.B. Shoot Competition and Root Competition. J. Appl. Ecol. 1988, 25, 279–296. [Google Scholar] [CrossRef]
- Martinez-Feria, R.A.; Castellano, M.J.; Dietzel, R.N.; Helmers, M.J.; Liebman, M.; Huber, I.; Archontoulis, S.V. Linking Crop- and Soil-Based Approaches to Evaluate System Nitrogen-Use Efficiency and Tradeoffs. Agric. Ecosyst. Environ. 2018, 256, 131–143. [Google Scholar] [CrossRef]
- Little, T.M.; Hills, F.J. Agricultural Experimentation Design and Analysis; Wiley: New York, NY, USA, 1978. [Google Scholar]
- Lithourgidis, A.S.; Dordas, C.A. Forage Yield, Growth Rate, and Nitrogen Uptake of Faba Bean Intercrops with Wheat, Barley, and Rye in Three Seeding Ratios. Crop Sci. 2010, 50, 2148–2158. [Google Scholar] [CrossRef]
- Bo, P.T.; Bai, Y.; Dong, Y.; Shi, H.; Soe Htet, M.N.; Samoon, H.A.; Zhang, R.; Tanveer, S.K.; Hai, J. Influence of Different Harvesting Stages and Cereals–Legume Mixture on Forage Biomass Yield, Nutritional Compositions, and Quality under Loess Plateau Region. Plants 2022, 11, 2801. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Andreuccetti, V.; Arduini, I.; Minieri, S.; Pampana, S. Field Bean for Forage and Grain in Short-Season Rainfed Mediterranean Conditions. Ital. J. Agron. 2018, 11, 1112. [Google Scholar] [CrossRef]
- Villalba, J.J.; Ates, S.; MacAdam, J.W. Non-Fiber Carbohydrates in Forages and Their Influence on Beef Production Systems. Front. Sustain. Food Syst. 2021, 5, 566338. [Google Scholar] [CrossRef]
- Luo, C.; Zhu, J.; Ma, L.; Guo, Z.; Dong, K.; Dong, Y. Effects of Nitrogen Regulation and Strip Intercropping on Faba Bean Biomass, Nitrogen Accumulation and Distribution, and Interspecific Interactions. Crop Sci. 2021, 61, 4325–4343. [Google Scholar] [CrossRef]
- Greenwood, D.J.; Lemaire, G.; Gosse, G.; Cruz, P.; Draycott, A.; Neeteson, J.J. Decline in Percentage N of C3 and C4 Crops with Increasing Plant Mass. Ann. Bot. 1990, 66, 425–436. [Google Scholar] [CrossRef]
- Capstaff, N.M.; Miller, A.J. Improving the Yield and Nutritional Quality of Forage Crops. Front. Plant Sci. 2018, 9, 535. [Google Scholar] [CrossRef]
- Urbatzka, P.; Graß, R.; Haase, T.; Schüler, C.; Heß, J. Fate of Legume-Derived Nitrogen in Monocultures and Mixtures with Cereals. Agric. Ecosyst. Environ. 2009, 132, 116–125. [Google Scholar] [CrossRef]
- Bannink, A.; France, J.; Lopez, S.; Gerrits, W.J.J.; Kebreab, E.; Tamminga, S.; Dijkstra, J. Modelling the Implications of Feeding Strategy on Rumen Fermentation and Functioning of the Rumen Wall. Anim. Feed. Sci. Technol. 2008, 143, 3–26. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Davies, T.E.; Edwards, J.E.; Theodorou, M.K. From Plants to Animals; the Role of Plant Cell Death in Ruminant Herbivores. J. Exp. Bot. 2008, 59, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Arisnabarreta, S.; Miralles, D.J. Nitrogen and Radiation Effects during the Active Spike-Growth Phase on Floret Development and Biomass Partitioning in 2-and 6-Rowed Barley Isolines. Crop Pasture Sci. 2010, 61, 578–587. [Google Scholar] [CrossRef]
- Dordas, C. Variation in Dry Matter and Nitrogen Accumulation and Remobilization in Barley as Affected by Fertilization, Cultivar, and Source–Sink Relations. Eur. J. Agron. 2012, 37, 31–42. [Google Scholar] [CrossRef]
- Bulman, P.; Smith, D.L. Post-heading Nitrogen Uptake, Retranslocation, and Partitioning in Spring Barley. Crop Sci. 1994, 34, 977–984. [Google Scholar] [CrossRef]
- Corre-Hellou, G.; Brisson, N.; Launay, M.; Fustec, J.; Crozat, Y. Effect of Root Depth Penetration on Soil Nitrogen Competitive Interactions and Dry Matter Production in Pea-Barley Intercrops given Different Soil Nitrogen Supplies. Field Crops Res. 2007, 103, 76–85. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Gooding, M.; Ambus, P.; Corre-Hellou, G.; Crozat, Y.; Dahlmann, C.; Dibet, A.; von Fragstein, P.; Pristeri, A.; Monti, M.; et al. Pea-Barley Intercropping for Efficient Symbiotic N2-Fixation, Soil N Acquisition and Use of Other Nutrients in European Organic Cropping Systems. Field Crops Res. 2009, 113, 64–71. [Google Scholar] [CrossRef]
- Dordas, C.A.; Lithourgidis, A.S. Growth, Yield and Nitrogen Performance of Faba Bean Intercrops with Oat and Triticale at Varying Seeding Ratios. Grass Forage Sci. 2011, 66, 569–577. [Google Scholar] [CrossRef]
- Herridge, D.F.; Pate, J.S. Utilization of Net Photosynthate for Nitrogen Fixation and Protein Production in an Annual Legume. Plant Physiol. 1977, 60, 759–764. [Google Scholar] [CrossRef]
- Pampana, S.; Masoni, A.; Arduini, I. Grain Legumes Differ in Nitrogen Accumulation and Remobilisation during Seed Filling. Acta Agric. Scand. B Soil Plant Sci. 2016, 66, 127–132. [Google Scholar] [CrossRef]
- Ofori, F.; Stern, W.R. Cereal-Legume Intercropping Systems. Adv. Agron. 1987, 41, 41–90. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Units | Method |
|---|---|---|---|
| Sand | 45.2 | % | Hydrometer (0.05 mm < Ø < 2 mm) |
| Silt | 40.6 | % | Hydrometer (0.002 mm < Ø < 0.05 mm) |
| Clay | 14.2 | % | Hydrometer (Ø < 0.002 mm) |
| pH | 8.5 | - | 1:2.5 (m:v) soil/water suspension |
| Organic matter | 2.1 | % | Walkley and Black |
| Total nitrogen | 1.1 | g N kg−1 | Kjeldahl |
| Available phosphorus | 20.1 | mg kg−1 P2O5 | Olsen |
| Exchangeable potassium | 141.1 | mg kg−1 K2O | Dirks–Sheffer |
| Period | Rainfall | T Max | T Min | GDD * Barley | GDD Field Bean |
|---|---|---|---|---|---|
| mm | °C | °C | °C day−1 | °C day−1 | |
| I Year | |||||
| Sowing–heading ** | 432.2 | 14.5 | 4.2 | 1452.9 | 1189.4 |
| Heading–early dough | 25.6 | 22.0 | 10.4 | 308.4 | 276.1 |
| Growth cycle | 457.8 | 18.3 | 7.3 | 1761.3 | 1465.5 |
| II Year | |||||
| Sowing–heading ** | 434.8 | 14.8 | 5.4 | 1624.7 | 1351.0 |
| Heading–early dough | 47.2 | 18.9 | 8.3 | 190.1 | 166.3 |
| Growth cycle | 482.0 | 16.8 | 6.8 | 1814.7 | 1517.2 |
| Stage of Harvesting | Growth Stage | Code of Stage | Date |
|---|---|---|---|
| Green forage | Barley: beginning of heading * Field bean: first pods visible | BBCH 70 BBCH 51 | 21 April 2015 20 April 2016 |
| Silage | Barley: early dough Field bean: final pods’ development | BBCH 83 BCCH 79 | 10 May 2015 4 May 2016 |
| Stage of Harvesting | Biomass | |||
|---|---|---|---|---|
| Leaves | Stems | Aerial | Leaves | |
| kg ha−1 | % on Total Aerial Biomass | |||
| Heading | 1884 b | 3898 b | 5930 b | 31.8 a |
| Early dough | 2077 a | 4373 a | 7380 a | 28.1 b |
| Stage of Harvesting | Nitrogen Concentration | Nitrogen Content | |||
|---|---|---|---|---|---|
| Leaves | Inflorescences | Aerial Biomass | Leaves | Aerial Biomass | |
| % | kg ha−1 | ||||
| Heading | 2.0 a | 1.7 a | 1.3 a | 28.6 a | 67.3 b |
| Early dough | 1.8 b | 1.5 b | 1.2 b | 23.8 b | 81.4 a |
| Stage of Harvesting | Nitrogen Concentration | Nitrogen Content | |||||
|---|---|---|---|---|---|---|---|
| Leaves | Stems | Inflorescences | Aerial Biomass | Leaves | Stems | Aerial Biomass | |
| % | kg ha−1 | ||||||
| Heading | 4.2 a | 1.2 a | 5.4 a | 2.2 a | 79.7 a | 45.7 a | 133.3 b |
| Early dough | 3.4 b | 0.9 b | 4.1 b | 2.0 b | 71.8 b | 38.2 b | 147.6 a |
| Stage of Harvesting | Nitrogen Concentration | Nitrogen Content | ||||
|---|---|---|---|---|---|---|
| Leaves | Stems | Aerial Biomass | Leaves | Stems | Aerial Biomass | |
| % | kg ha−1 | |||||
| Heading | 3.3 a | 1.0 a | 1.8 a | 77.6 a | 50.90 a | 143.6 b |
| Early dough | 2.9 b | 0.8 b | 1.6 b | 67.7 b | 47.13 a | 161.8 a |
| Stage of Harvesting | Barley | Field Bean | Total LER | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Partial LER | A | CR | Cb | Partial LER | A | CR | Cb | ||
| Heading | 0. 77 a | 0.07 a | 1.17 a | 0.08 a | 0.70 a | −0.07 b | 1.00 b | −0.08 b | 1.47 a |
| Early dough | 0.67 b | −0.04 b | 0.96 b | −0.07 b | 0.72 a | 0.04 a | 1.11 a | 0.07 a | 1.39 b |
| Stage of Harvesting | Barley | Field Bean | Total N-LER | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Partial N-LER | N-A | N-CR | N-Cb | Partial N-LER | N-A | N-CR | N-Cb | ||
| Heading | 1.18 a | 0.51 a | 1.86 a | 0.55 a | 0.66 a | −0.51 a | 0.62 a | −0.55 a | 1.84 a |
| Early dough | 1.04 b | 0.40 a | 1.64 a | 0.44 a | 0.64 a | −0.40 a | 0.67 a | −0.44 a | 1.69 b |
| Stage of Harvesting | Nitrogen Use Efficiency | ||
|---|---|---|---|
| Barley | Field Bean | Combined Species | |
| kg kg−1 | |||
| Heading | 0.7 b | 1.4 b | 1.5 b |
| Early dough | 0.8 a | 1.6 a | 1.7 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeletti, F.G.S.; Pampana, S.; Saia, S.; Mariotti, M. Better Land and Nitrogen Complementarity for Green Forage Than for Silage in Barley–Field Bean Intercropping. Agronomy 2023, 13, 2886. https://doi.org/10.3390/agronomy13122886
Angeletti FGS, Pampana S, Saia S, Mariotti M. Better Land and Nitrogen Complementarity for Green Forage Than for Silage in Barley–Field Bean Intercropping. Agronomy. 2023; 13(12):2886. https://doi.org/10.3390/agronomy13122886
Chicago/Turabian StyleAngeletti, Francesco Giovanni Salvo, Silvia Pampana, Sergio Saia, and Marco Mariotti. 2023. "Better Land and Nitrogen Complementarity for Green Forage Than for Silage in Barley–Field Bean Intercropping" Agronomy 13, no. 12: 2886. https://doi.org/10.3390/agronomy13122886