1. Introduction
Nanotechnology finds diverse applications in agriculture, including nanosensors, nanopesticides, and nanofertilizers [
1,
2]. A nanoparticle is typically defined as a material with dimensions up to 100 nm in one of the three vector directions [
3].
Following the “essentiality” concept for plant nutrition proposed by Liu and Lal [
4], different NFs are categorized as (i) macro-NFs containing elements such as Ca, C, N, P, K; (ii) micro-NFs (e.g., Zn, Fe, Cu); (iii) nanomaterial-enhanced fertilizers (e.g., SiO
2 or silicates with nutrient-doped porous diatoms); and (iv) nanoparticulate plant-growth enhancers, such as carbon nanotubes, TiO
2-NPs, and Au-NPs, with undetermined modes of actions.
The ZnO-NP fertilizer shows promise as an alternative to conventional ZnSO
4 or macro-sized ZnO fertilizers due to its unique properties [
5,
6,
7]. Generally, Zn, an essential micronutrient, plays diverse roles in plant physiology, including chlorophyll synthesis, enzyme cofactor functions, stress tolerance, and the regulation of plant hormones like auxin and cytokinin [
8,
9,
10].
Iron is a crucial micronutrient for plants, indispensable for protein and enzyme formation, electron transport in cells, chlorophyll synthesis, nucleic acid production, ATP synthesis, atmospheric nitrogen fixation, and stress mitigation [
11]. Hence, Fe deficiency can result in leaf chlorosis [
12,
13]. In flowers, anthers serve as major Fe reservoirs [
14], and Fe is vital for proper pollen development. Deficiency during the reproductive stage can lead to distorted pollen due to tapetum dysfunction and ultimately, as a result of developmental disorders, pollen grain exine defects occur [
15].
TiO
2, originating from various industrial activities, including agriculture, has become a notable environmental factor [
16]. While not essential for plant growth, nano-sized TiO
2 positively influences plant physiology by enhancing enzyme activity [
17,
18], chlorophyll content, photosynthetic efficiency [
19], drought tolerance [
20], nutrient uptake and absorption [
21], plant morphology [
22], seedling development, pollen germination, DNA protection, and reducing heavy metal toxicity [
23]. Titanium-based organic complexes have shown positive effects on apple pollination [
24] and strawberry fruit development [
25]. On the other hand, several adverse impacts on plants have been described, such as a reduction in the initial germination stages of
Vicia narbonensis and
Zea mays at higher doses [
26]. Additionally, the application of TiO
2-NPs in soybean plants (
Glycine max) led to a significant reduction in the colonization of arbuscular mycorrhizal fungi, plant growth, and leaf phosphorus levels [
27].
Silica, although not traditionally considered essential, significantly influences plant growth and development. It strengthens cell walls, enhances stress resistance, stimulates the immune system, promotes antioxidant production, and supports photosynthesis by reducing transpiration [
28,
29,
30]. However, the role of SiO
2 mesoporous silica in anchoring Au-NPs remains unclear. Gold, typically insignificant to plant growth, has limited physiological or biochemical functions in NP form. Positive effects are observed at ultra-low concentrations during germination and early development [
31]. Gold NPs obtained by green synthesis induce cell division and increase pollen germination [
32,
33].
In the realm of applying NPs to plants from an ecotoxicological perspective, diverse tests evaluate their environmental effects, including assessments for acute toxicity, chronic toxicity, reproductive toxicity, genotoxicity, and more [
34].
Pollen, with its unique capacity to absorb and accumulate a diverse range of pollutants such as heavy metals, pesticides, toxins, and other contaminants, could serve as a valuable biomarker for assessing the impact of environmental pollution [
35,
36] or for environmental conditions biomonitoring [
37,
38]. Examination of the reproductive structures of a flower reveals that pollen, as the male gametophyte, is more sensitive compared to ovules of the female gametophyte [
39]. Pollen plays a critical role in fertilizing the female gamete to develop a plant embryo. Yet, this phase in the plant life cycle is notably vulnerable due to the heightened sensitivity of the male haploid phase to various environmental conditions, including the fertilization method [
40,
41].
In the current academic literature, the morphological alterations of pollen affected by diverse NFs are not comprehensively documented. Only a few studies have described variable negative effects of NPs at high concentrations. For example, the application of Ag-NPs led to decreased pollen viability in
Arabidopsis [
42], and a toxic impact on the pollen structure of kiwifruit was reported [
43]. Furthermore, Dutta Gupta et al. [
44] observed changes in exine structure and pollen grain morphology in
Peltophorum due to Ag-NPs, while Pd-NPs resulted in pore penetration and alterations in ultrastructure and pollen grain shape [
36]. Conversely, a few positive effects of Ag-NPs on the size and viability of peach pollen grains were demonstrated by Mosa et al. [
45]. In this specific instance, there is a notable absence of information regarding the influence on plant development and the pollen morphology associated with nano-domain effects of micronutrient-based Zn, Fe NFs, or growth stimulants such as Si and Au, as well as the potential consequences for the final crop yield.
Changes in the morphological attributes of pollen grains, including size, shape, exine structure, sculpture, polar axis length, equatorial diameter, colpi characteristics, and spines, can provide direct or indirect insights into various environmental and ecological changes [
46] or serve as significant diagnostic features for the
Asteraceae family [
47].
In general, the pollen grain coat plays a crucial role in protection, facilitating effective pollen transfer and enabling interactions with stigmas [
48].
Additionally, the size, structure, and density of spines influence the accessibility of pollen for pollinators and contribute to pollinator diversity [
49]. Moreover, the intricate ornamentation of the exine offers a greater surface area for interactions with the environment. Gametogenesis affects the resultant shape and size variations of pollen [
50], which in turn, determine pollen functionality [
51].
In this context, sunflower (
Helianthus annuus L.) is one of the most widely cultivated oil-bearing crops worldwide with significant agronomic perspective [
52,
53]. Sunflower pollen holds promise as a valuable nutritional resource, being rich in lipids, antioxidants, and essential nutrients such as Na and K [
54]. These attributes make it a vital source for bees and other pollinators that play crucial roles in supporting ecological communities [
55]. Additionally, sunflower pollen exhibits noteworthy antiseptic effects on pollinators [
55,
56].
Current studies on changes in pollen morphology primarily focus on ecotoxicological
in vitro investigations conducted under strict laboratory conditions, usually using higher concentrations of NPs [
36,
43]. Surprisingly, academic outcomes lack real-world agronomic research addressing the implications of environmentally sustainable NP concentrations compared to conventional fertilizers.
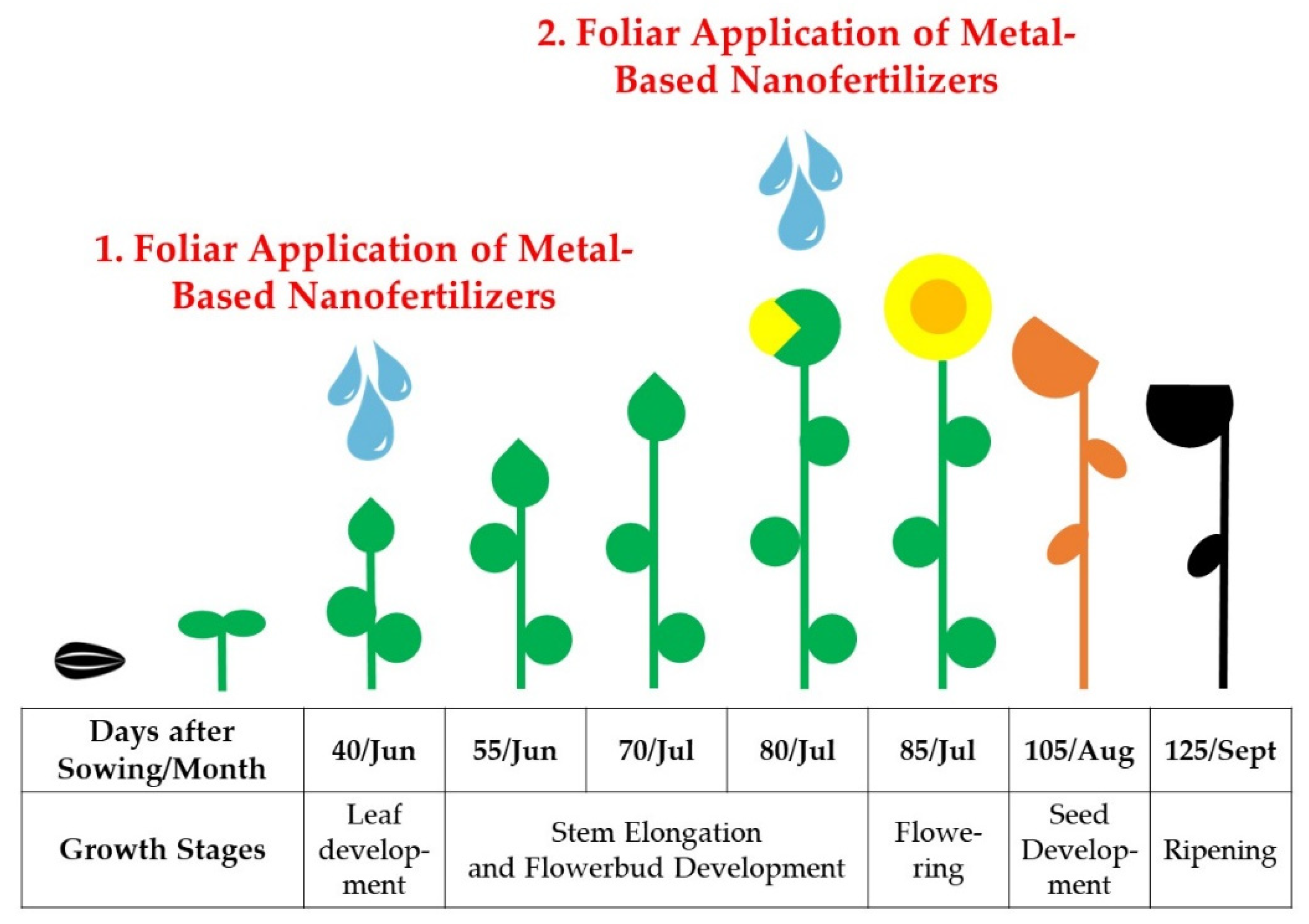
The aim of the current study was to evaluate selected micro-based and plant-enhanced NPs at eco-friendly concentrations. This involved examining their chemical reactivity, physical stability, as well as their influence on pollen morphology in association with their agronomical yield. The research was conducted in field conditions over three growing seasons, employing two sunflower hybrids.
4. Discussion
Metal-based NPs enter the environment through various anthropogenic pathways, including agricultural activities [
1]. In the form of soluble ion-metals, they naturally contribute to plant metabolism, growth, and development [
8]. However, at higher concentrations, NPs can often have negative or toxic effects on plants [
72]. The effects of metallic-based NPs and ionic components in plants have been extensively studied, particularly concerning physiological and biochemical aspects such as enzymatic activity [
73], photosynthesis and transpiration [
74], metal accumulation affecting plant tissue [
75], or the development of reproductive organs [
76]. However, only a few studies have investigated the direct impact of NPs on pollen or pollen morphology in real agronomic conditions. Most research has focused on atmospheric environmental pollution [
37] or the immediate effects on pollen viability and other
in vitro functional attributes [
36,
43]. Hence, our study provides valuable insights into the understanding of the effects of diverse NP foliar applications on sunflower pollen morphology and yield, filling a notable gap in this research area.
In this context, sunflowers serve as excellent models for both agronomic and environmental research.
4.1. The Effect of ZnO-NPs and Conventional Agrochemical ZnSO4 on Sunflower Pollen Morphology and Their Impact on Yield
Our findings indicate a positive impact of ZnO-NPs foliar application on sunflower yield (
Table 8), yield-related parameters, and seasonal physiology during the 2019 season, as previously described in a study by Ernst et al. [
60].
For the Neostar hybrid, a demonstrable alteration in pollen morphology was observed, specifically in the spine parameter. In 2019, it exhibited a lower value, while in 2020, it showed a higher value compared to the control. Notably, this parameter displayed significant variability, especially in 2020, among all the NP-treated variants.
In the context of sunflower cultivation, the correlation between temperature and annual precipitation, as highlighted by Astiz and Hernández [
77], may have subsequent implications for nutrient uptake, absorption, or the response to NP foliar deposition. Our results clearly show that during pollen maturation in July, there was a marked absence of rainfall, combined with normal temperatures. In contrast, the vegetation season of 2020 was characterized by minimal precipitation and exceptionally cold temperatures (see
Table 2 and
Table 3). In addition, in 2020, statistically significant enhancements were noted in pollen morphology parameters, specifically in the elongation of polar axes and the APG parameter.
The Edison hybrid exhibited no statistically significant differences in pollen grain morphology and its morphological parameters compared to the control variant. In fact, it displayed slightly negative characteristics, though not statistically significant (see
Table 6). This could be attributed to the inherent variability of the Edison hybrid [
62], which shows lower sensitivity to metal-based NP foliar application or other external abiotic-induced stressors, including seasonal–climatic fluctuations.
Nevertheless, in these practical, in-field experiments where ZnO-NPs were applied to sunflowers, we have, for the first time, confirmed that despite the varied responses in pollen morphology, the sunflower yield consistently demonstrated a positive trend over three growing seasons for both sunflower hybrids evaluated. In this context, Zn appears to play a significant role in the development of male reproductive organs and gametes. Zinc deficiency results in delayed flowering, reduced size of the anthers, pollen production, decreased size of pollen grains, and causes morphological changes in pollen, especially in the exine structure of lentils [
40] and black gram [
41]. Additionally, reduced pollen viability has been found in maize [
78], lentils [
40] and black gram [
41], and chickpeas [
79].
Considering the sunflower plant response, including pollen morphology, we can anticipate two effects after the application of ZnO-NPs. First, as we know, when colloidal dispersion is applied, ZnO-NPs undergo photodegradation, leading to Zn assimilation and subsequent redistribution by the leaves/plant to various geochemical species [
80]. This process could potentially enhance other physiological, metabolic, and biochemical pathways in the plant [
8]. In terms of physiology and higher yields, we have previously observed such effects at low, comparable concentrations in the case of
Setaria italica [
57], lentils [
58], and sunflowers [
81]. The interaction with ZnO-NPs could have a direct impact on the morphology, structure, and function of pollen, as outlined by Yoshihara et al. [
82]. This interaction resulted in reduced pollen grain germination and tube elongation in the case of
Lilium longiflorum. Moreover, the application of commercial nanofertilizer ZnO-NPs at a concentration of 7.5 mL·L
−1 caused pollen aberrations and structural changes in the sporoderm, along with degenerative changes in the content of peach pollen grains [
45].
On the other side, the negative impact of Zn deficiency on the morphology and viability of pollen grains can be mitigated by adding ZnSO
4 [
79]. Although Zn supplementation at a dose of 500 mg/pot ZnSO
4 has a positive effect on the size of pea pollen, a high dose of 1000 mg/pot of this element causes abnormalities in sporoderm of pollen grains and deformation of the pollen [
83]. Our result confirmed that after the application of ZnSO
4, sunflower pollen grains clearly reached the highest values for several evaluated size parameters, reflecting the positive influence of Zn and S on pollen development.
A remarkable finding is the change in the shape of the pollen of the Neostar variety after the application of ZnO-NPs in 2019, when all the treated variants exhibited a shape modification of the pollen grains compared to the control. While we defined the pollen grains in the control variant as subprolate based on the ratio of the polar and equatorial diameter, in the treated variants including ZnO-NPs, they were prolate spheroidal (
Table 7). However, in the following year, this change in shape occurred only in the variant with ZnSO
4 treatment.
This shape modification can be associated with seasonal weather fluctuations (see
Table 2 and
Table 3) and could potentially impact pollen collection and distribution by sunflower pollinators [
84]. In this context, the use of agronomically and conventionally applied ZnSO
4 appears to be more suitable than treatment with “pure” ZnO-NPs. ZnSO
4 is readily bioavailable and mobile in an ion-soluble form. Additionally, the SO
4 anion may function as a macronutrient [
79], potentially leading to a more substantial agronomic impact (
Table 8).
This was further confirmed at a statistically more significant level in terms of yield (refer to
Table 8) and all relevant parameters of pollen morphology, namely the length of polar axis, equatorial diameter, and area of pollen grains (APGa) (see
Table 5). In addition, since SO
4 is a biologically available form of sulphur, it can participate in the formation of pectins [
85], which are polymers integrated into the intine of pollen grains [
86], naturally supporting the function of pollen grain germination. It can also contribute to the production and regulation of phytohormones, especially cytokinin, which could be reflected in the development of sunflower pollen [
87]. In this scenario, larger pollen grains have an advantage in germination and growth towards fertilizing ovules when compared to smaller pollen grains, as demonstrated in
Hibiscus syriacus with improved fruit quality [
88].
4.2. The Impact of Iron Nanoparticles on Pollen Morphology and Their Associations with Sunflower Yield
In terms of pollen morphology, the application of Fe
3O
4-NPs appears to have a statistically insignificant impact. This is evident from the smaller pollen grains (APGb) and shorter spines compared to the other variants, including the NP-free control. These findings may be linked to plain seasonal physiological responses and relatively indistinct yield (see
Table 8) and yield-related parameters, including oil content, which are comparable to the control without NP application, as documented in the study by Ernst et al. [
60].
In heavy metal-contaminated soils, with Zn and Fe concentrations sometimes up to seven times higher where
Chenopodium botrys L. was grown, Yousefi et al. [
89] observed other development irregularities that affected plant fertility and survival. These irregularities also led to changes in anther morphology and a reduced pollen count in plants from the contaminated site.
In our study, one of the stress-inducing factors associated with the properties of Fe
3O
4-NPs, particularly concerning pollen morphology, is their small size, which does not exceed 5 nm. In this size range, we can expect a nano-domain effect, resulting in a certain level of metastability and an associated increase in the reactivity of these NPs, as pointed out by Lin et al. [
90]. Assuming monodisperse NPs with high crystallinity and a higher concentration (76 mg·L
−1), a hypothesis could be formulated regarding the increased proliferation, translocation, and enormous reactivity of Fe
3O
4-NPs or their residues into pollen grains during the flowering phenophase. However, this hypothesis is challenged by the overall lower Fe concentrations in final-quality sunflower grains, as reported by Ernst et al. [
60].
While soluble Fe is recognized as a micronutrient, the transport, mobility, and effects, as well as the destiny of Fe in the NP nature within plants or their structures, may differ. This variance has been demonstrated in pumpkin plants (
Cucurbita maxima L.) [
91] and is evident in its negative impact on the overall development of tobacco (
Nicotiana tabacum) [
92].
If Fe
3O
4-NPs could directly impact pollen grains, we should take into consideration their higher mechanical and chemical stability, which could modify various pollen properties, such as a reduction in pollen viability or the abortiveness of
Arabidopsis with a subsequent reduction in seed yield [
93].
4.3. The Examined Impact of AuSi NPs on Pollen Morphology and Their Correlation with Sunflower Crop Yield
The evaluation of the AuSi-NP variant after application at the same concentration (0.1 mg·L
−1 Au and 10 mg·L
−1 SiO
2) in two growing seasons and with two hybrids, Neostar and Edison, yielded somewhat ambiguous results. In 2019, for the Neostar hybrid, when considering pollen morphology, statistically significant differences were observed only in the case of an elongated equatorial diameter. There were also indications of potential changes in polar diameter and spine length, but these were statistically insignificant. The other parameters favored the control variant, but with insignificant differences (see
Table 4).
In the case of the Edison hybrid, during the 2021 vegetation season (see
Table 6), the morphological parameters, when compared to the control, were statistically insignificant. Considerable differences from the NP-free variant were observed only in terms of shorter spine length or smaller equatorial diameter, which does not correspond to the results of the Neostar hybrid. Here, spines were the shortest among all variants, including the control. This paradoxical variation in spine length could be seen as an indicator of differential protection against potential environmental effects between the two hybrids.
Like pollen spines, shorter equatorial diameters may also represent a type of plant adaptation mechanism in response to specific environments or preferences for pollinators, aiding the plant in facilitating pollination by certain types of insects or other pollinators [
94]. Surprisingly, it is worth noting that the AuSi nano-composite exhibited effects on pollen morphology like ZnO-NPs in both applied hybrids and growing seasons. However, it demonstrated a significant increasing trend in yield (refer to
Table 8), even in the case of the Edison hybrid, which is known to be less responsive to external factors, and this was at a higher statistical level.
The Au-NPs at a size of approximately 10 nm, with excellent crystallinity, monodispersity, and a dominant spherical shape [
61], along with unique biocompatibility and stability [
95], theoretically suggest that Au NPs could pass through plant membranes and tissues relatively easily [
31], potentially negatively affecting the final quality and content of sunflower achenes. However, this was not chemically and analytically confirmed in our case [
60]. It may be related to the very low concentration we applied the NPs (0.1 mg·L
−1), which was best reflected in terms of physiological parameters, yield, and yield-related parameters [
60]. Mahakham et al. [
96] found in maize that Au-NPs preferentially entered the grains, but translocation into vegetative organs or roots was not recorded.
Hypothetically, Au-NPs may directly affect pollen morphology, potentially causing disruptions in internal and external structures and functionality. Surprisingly, our findings during the 2019 vegetation season demonstrated a higher agro-ecological terrestrial insect population, optimal living conditions, and functionality [
60].
Since Au-NPs were applied together with SiO
2 mesoporous silica
Mallomonas kaline, we also expected a synergistic effect due to the presence of silica as a plant micronutrient [
97] and a kind of plant growth stimulator in NP form [
4]. This could possibly explain the observed twofold increase in Si concentrations in the final seed quality compared to the control [
60]. This form of SiO
2 (mesoporous silica
Mallomonas kaline) is relatively more soluble, more bioavailable, and more mobile than crystallographically structurally more stable and inert nanoforms of SiO
2. This form, in the context of negative effects on selected terrestrial communities, was confirmed by Hussein et al. [
98] with the potential of being used as a nanopesticide.
An alternative hypothesis suggests that the pollen or other generative parts of the plant might incorporate silica due to its functional sorption complex, relative smaller size, porous structure, chemical inertness, or surface modification [
61].
4.4. The Effect of TiO2-NPs Treatment on Sunflower Pollen Morphology and Yield
From our results, it is evident that sunflower yields for both hybrids were significantly higher when applying TiO
2-NPs, not only in comparison with the control but also with all other NP-treated variants (see
Table 8). These findings are quite surprising because, when we consider the relatively negative impact on the pollen size of both the Neostar and Edison hybrids, they showed the lowest values for the polar axis, equatorial diameter, and other related parameters (see
Table 5 and
Table 6). Paradoxically, although smaller pollen grains are generally characterized by lower germination, they may show a higher rate of pollen tube growth. From this point view, even small pollen can produce more vigorous progeny, which can pose a certain selective advantage [
99].
Concurrently, pollen grains in the variant with Ti in both varieties exhibited statistically the highest values for spines compared to other variants. This can be seen as a kind of adaptive mechanism in response to Ti-induced stress, which could, on the other hand, significantly benefit the visitation preferences of individual pollinator types. In effect, some pollinators visit species with large echinate pollen grains less frequently, such as
Knautia arvensis and some
Malvaceae, but also collect similar pollen from
Cucurbita pepo [
100].
In the case of the application of TiO
2-NPs, positive, negative, or no effects are elaborated in connection with plant response in the academic literature [
72]. Here Ti is not presumed to have an essential character but is rather considered as a plant stimulator [
4,
72].
Evidently, our results showed a relatively negative impact on the morphological features of sunflower pollen with TiO
2-NPs interaction, while crop yields consistently remained excellent (see
Table 8). In this context, Vear et al. [
101] hypothesize that the energy saved in pollen production can be used to increase the oil content. There are indications of the impact of TiO
2-NPs, evident in the intensified physiology of the same hybrid, Neostar, during the 2018 growing season, along with yield and yield-related parameters, as well as oil content [
81]. These outcomes are comparable to the improvements in yield and oil content seen in
Glycine max, as published by Rezaei et al. [
102].
In our study, we applied the anatase-rutile form of TiO
2-NPs with structural modifications, higher chemical and mechanical stability, and a size corresponding to ~30 nm [
81]. The potential positive impact of our applied concentration range of TiO
2-NPs has been reflected in several studies. Larue et al. [
103] observed increased uptake and translocation of Ti into leaves, as well as root elongation in
Triticum aestivum L., albeit at similar sizes but slightly higher concentrations. Similarly, Servin et al. [
104] noted enhanced root growth in
Cucumis sativus, which also exhibited a strong correlation with their nitrogen content. It is worth noting that in both cases, TiO
2-NPs were applied via the root system rather than through leaf deposition.
Additionally, when considering the analytically determined total content of Ti affecting the final quality of sunflower grains, where TiO
2 is one of the most resistant materials, the NPs have a relatively small size and ability to transfer within the plant. It is surprising that an increased total content was not found in them compared to the NP-free control [
81]. Thus, we can indirectly assert that the transfer of Ti or TiO
2-NPs into the reproductive organs, which affects seed quality, was limited, even though there are certainly indications of modifications in pollen morphology (see
Table 5 and
Table 6).
On the contrary, we postulated that TiO
2-NPs might exert adverse effects due to the pronounced chemical and mechanical stability of NPs associated with the potential negative photocatalytic activity of TiO
2-NPs, generating reactive oxygen species (ROS) when exposed to sunlight, as elucidated by Lei et al. [
19]. Nevertheless, the study conducted by Huang et al. [
105] highlights their beneficial impact in improving pollen germination rates and pollen tube length in litchi male flowers through the application of TiO
2-NPs.
5. Conclusions
In summary, our study found that the application of metal-based NPs as NFs had variable effects on sunflower pollen morphology, but only at non-critical levels, and they did not significantly impact crop yields in field experiments conducted over three vegetative seasons with two hybrids of sunflower. We credit this outcome to the application of eco-friendly, low-concentration NFs, ensuring precise and sustainable agriculture.
(i) There was a relative “negative” trend in the application of TiO2-NPs concerning sunflower pollen morphology; however, this did not manifest in the form of reduced crop yields for both sunflower hybrids, Neostar and Edison. It may be interpreted as an adaptive mode of actions in response to induced Ti or TiO2-NP stress during sunflower development, targeting more to yield production with anatase-rutile minerals at the ~30 nm size.
(ii) At the same Zn concentration corresponding to 10 mg·L−1, the most favorable response to sunflower pollen morphology was observed with the agronomically conventional ionically soluble fertilizer ZnSO4, followed by ZnO-NPs. Furthermore, the use of both NFs led to higher yields compared to the control, where yields were consistently diminished.
(iii) When we consider Au-NPs as a plant-growth stimulator affecting plant morphology corresponding to 10 nm Au-NPs anchored to SiO2 mesoporous silica, there were no clear differences observed in the examined parameters. This might be related to the applied hybrids, with Neostar appearing to be more sensitive than the Edison hybrid, as well as in the control experiments (NP-free). However, the AuSi-NPs variant unexpectedly achieved higher yields, where surprisingly the Edison hybrid showed a more beneficial response to yield at a statistically significant level, even though it is considered to be less sensitive.
(iv) In this study, no statistically significant differences in pollen morphology were observed when magnetite (Fe3O4-NPs), classified as a micro-based nanoform, was utilized at its highest concentration with the smallest particle size. Contrary to our initial hypothesis, which posited higher reactivity due to its relative stability and active surface, this postulate was not confirmed. However, the agronomic yield exhibited a slight improvement when compared to the NP-free control.

 ,
, 



{kind=link}
{kind=link}
{kind=link}