Chitosan Reduces Damages of Strawberry Seedlings under High-Temperature and High-Light Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Design
2.2. Determination Indicators and Methods
2.2.1. Chl Content Determination
2.2.2. Determination of Gas Exchange Parameters and Chl Fluorescence Parameters
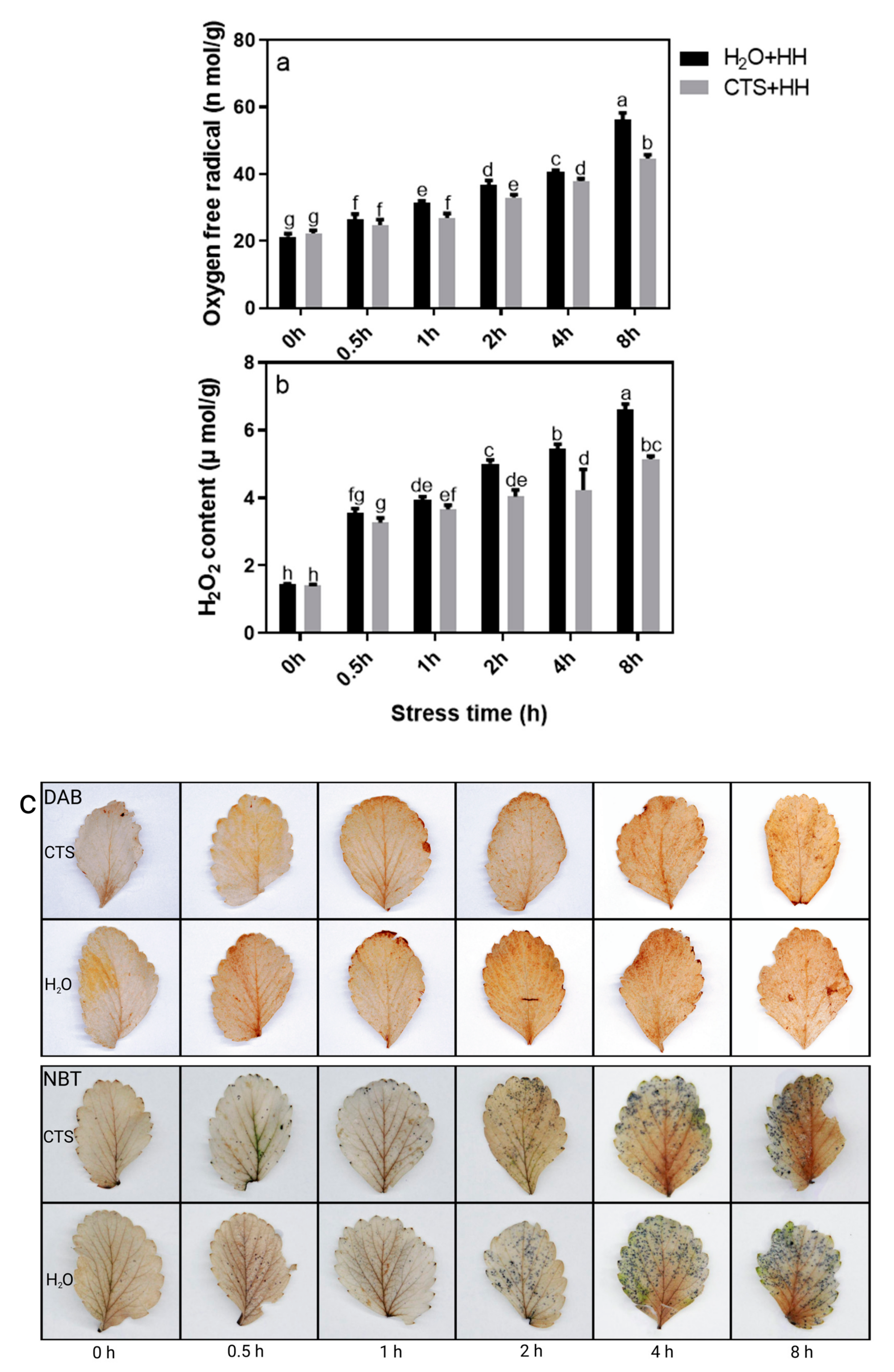
2.2.3. Tissue Staining and Determination of the Contents of H2O2 and O2−
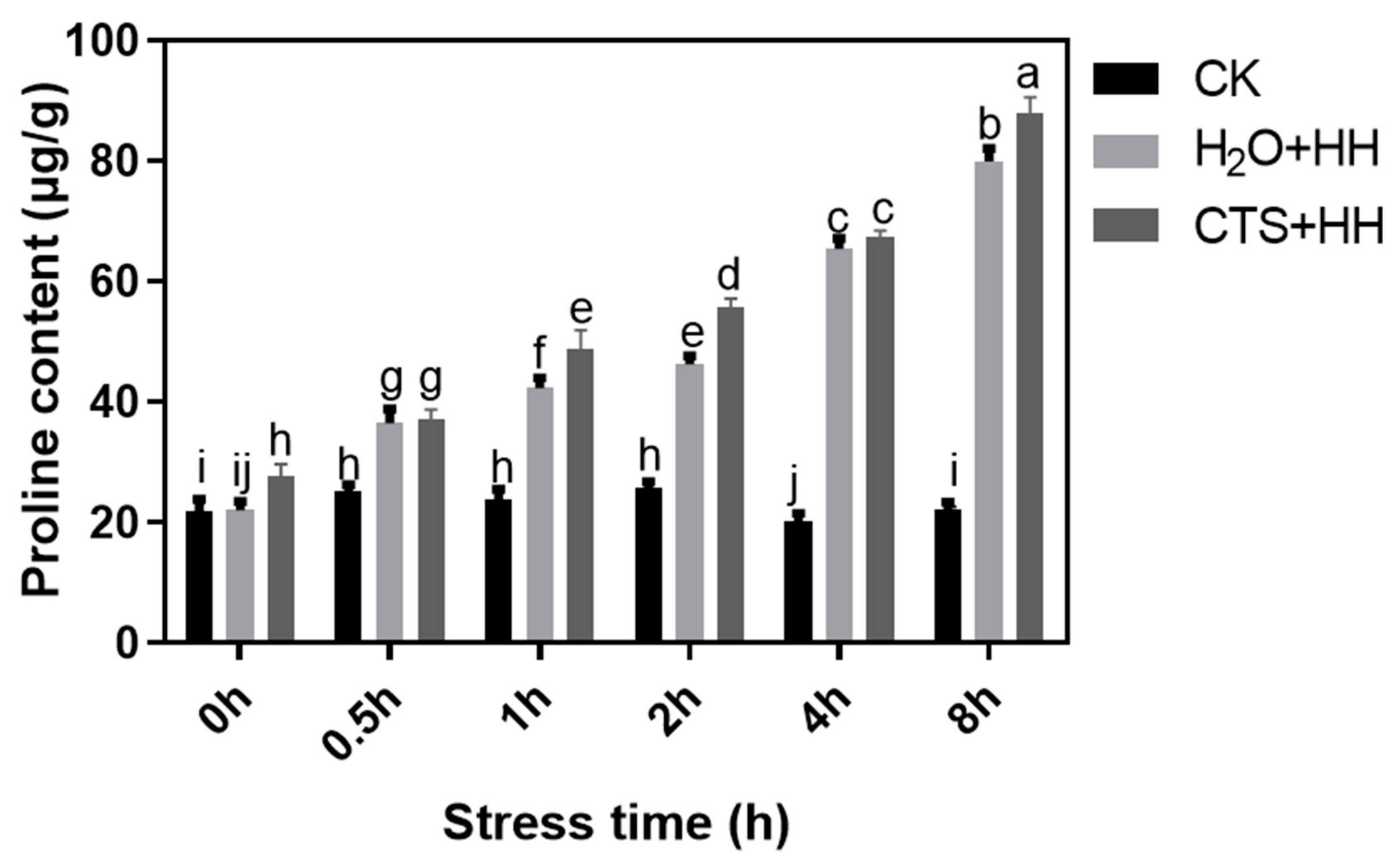
2.2.4. Determination of the Proline Content
2.2.5. Determination of the Relative Conductivity
2.2.6. Determination of the Malondialdehyde (MDA) Content
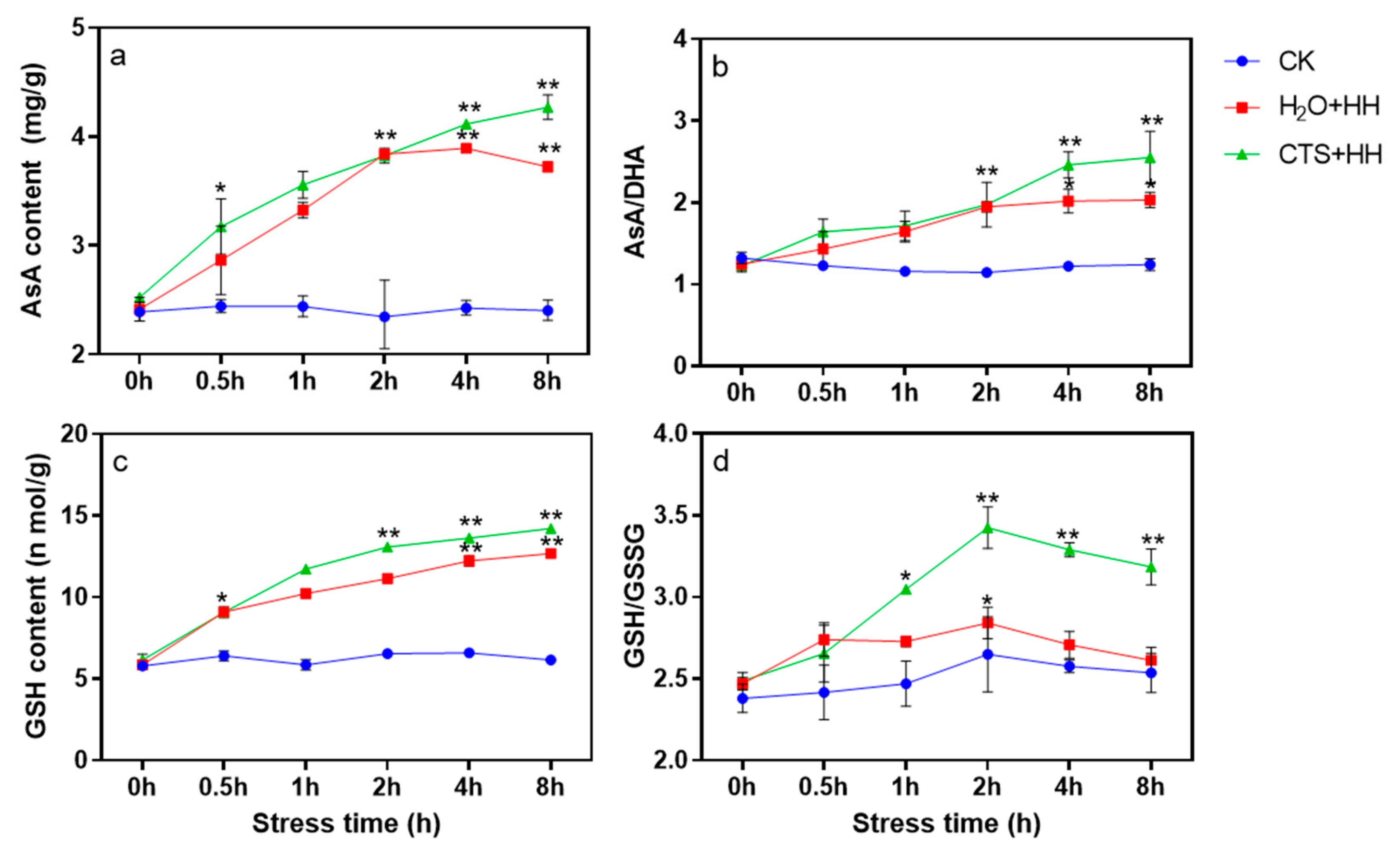
2.2.7. AsA and GSH Contents and Determination of the AsA/DHA GSH/GSSG Ratios
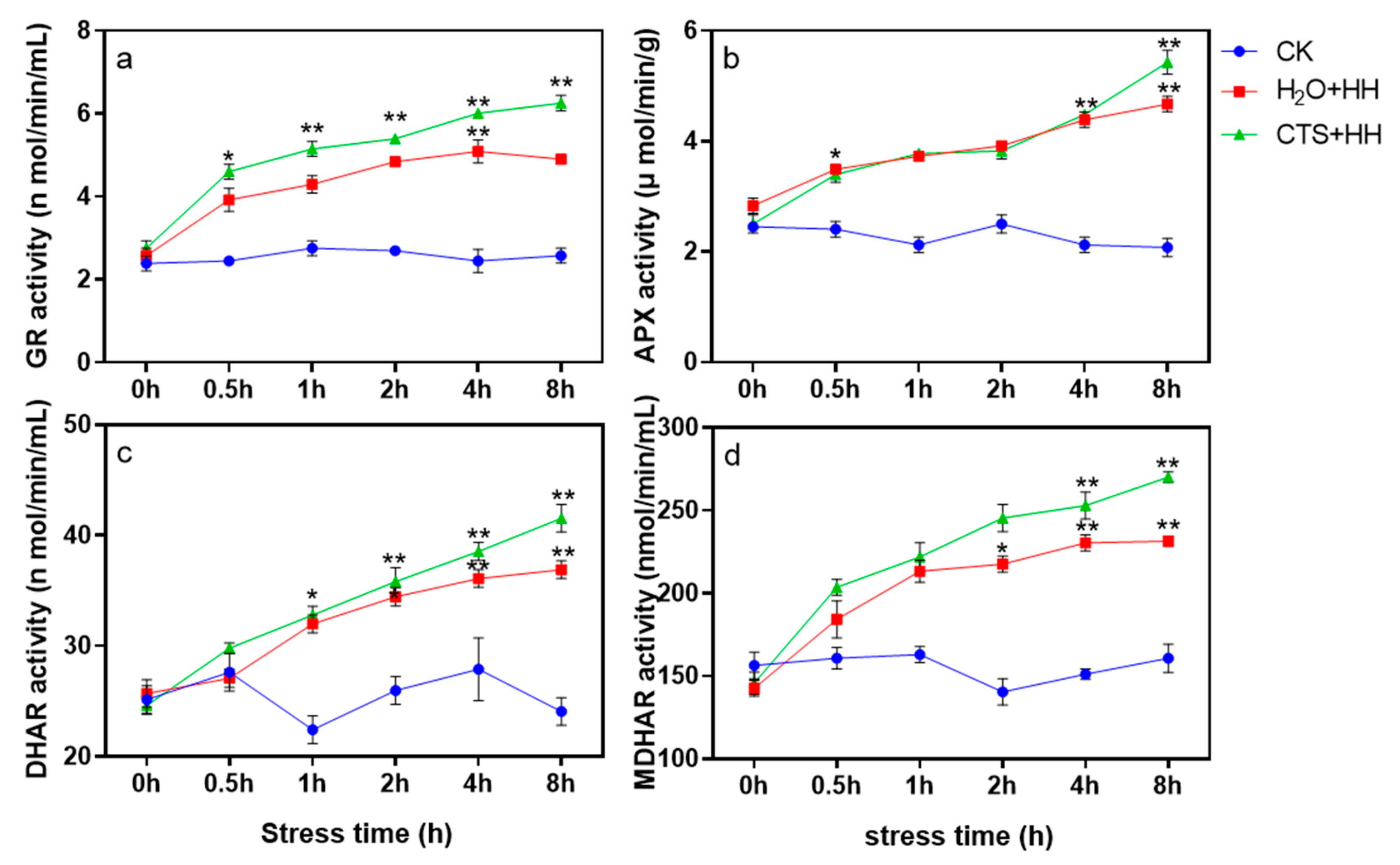
2.2.8. Determination of AsA-GSH Cycle-Related Enzyme Activities
2.3. Statistics and Data Analysis
3. Results
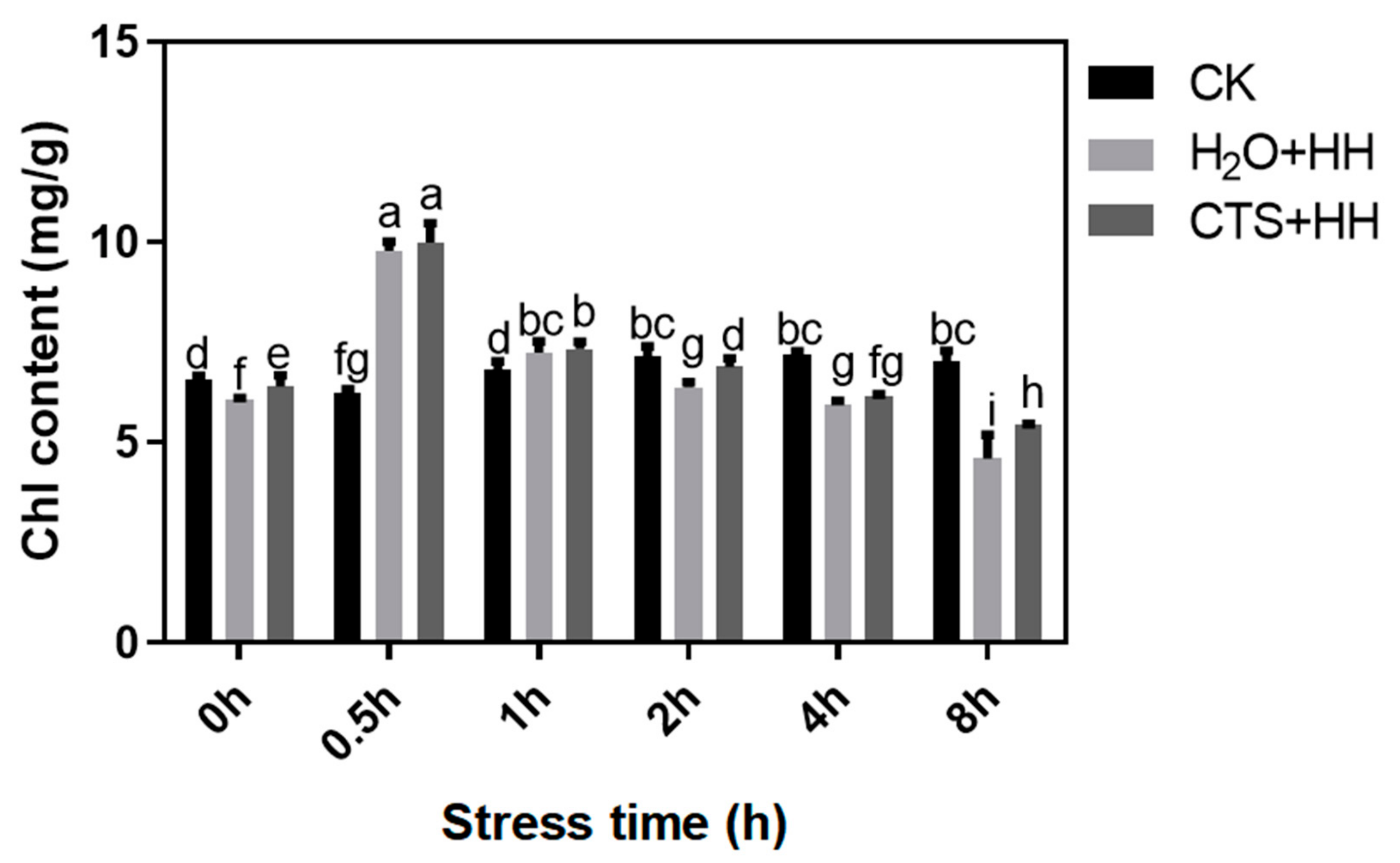
3.1. Effect of CTS on Chl Content
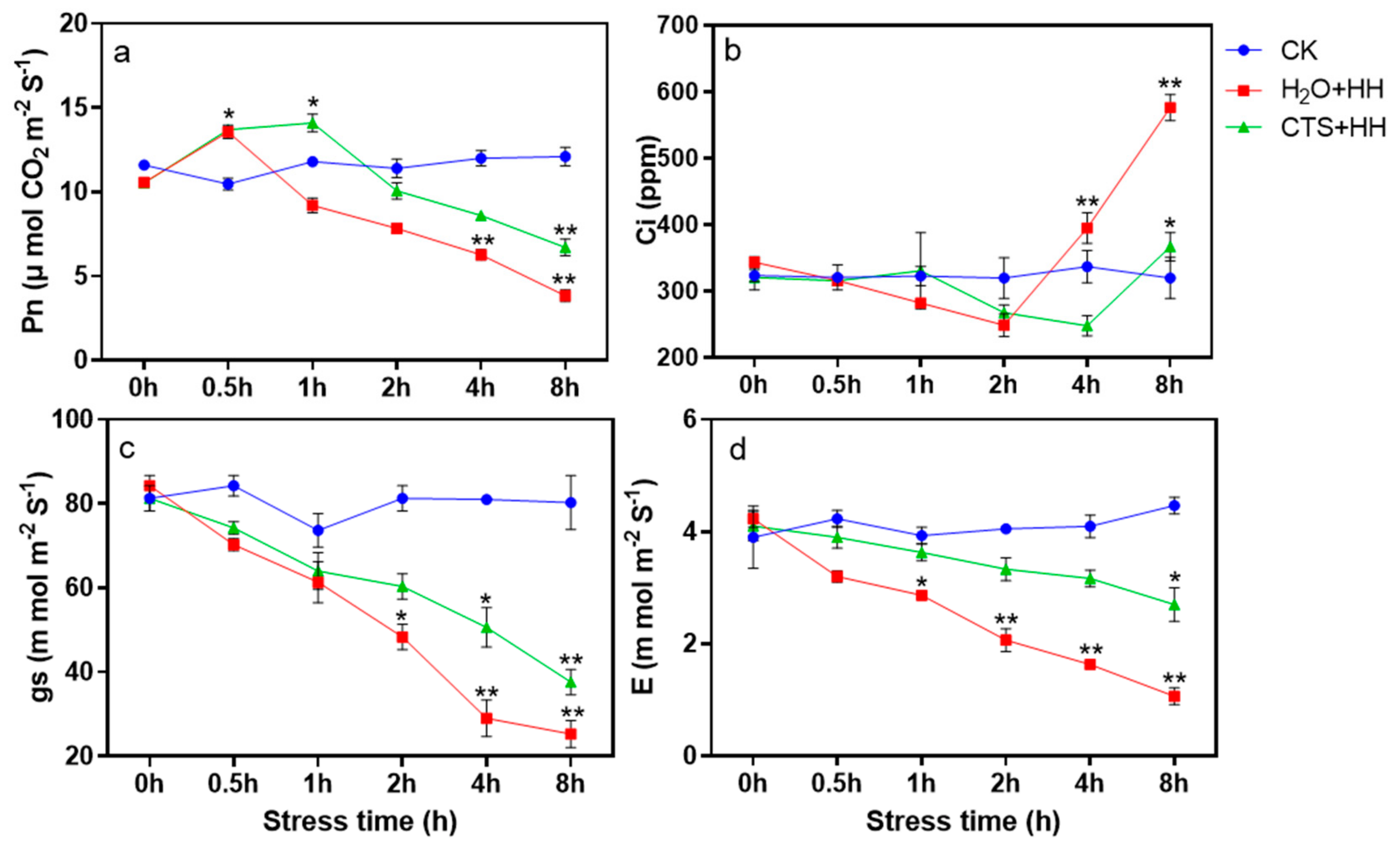
3.2. Effect of CTS Treatment on Gas Exchange Parameters
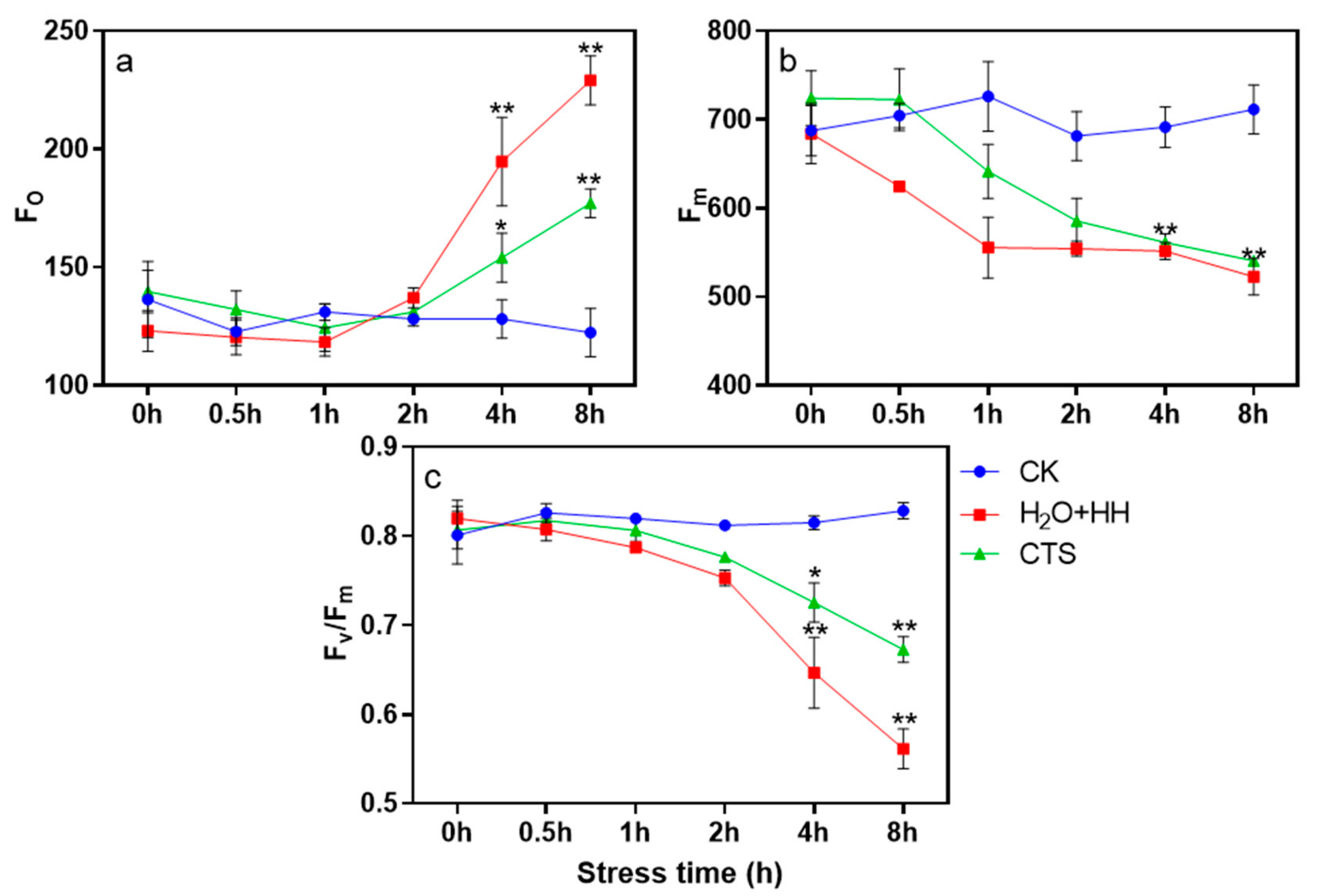
3.3. Effect of CTS on PSII Photoinhibition
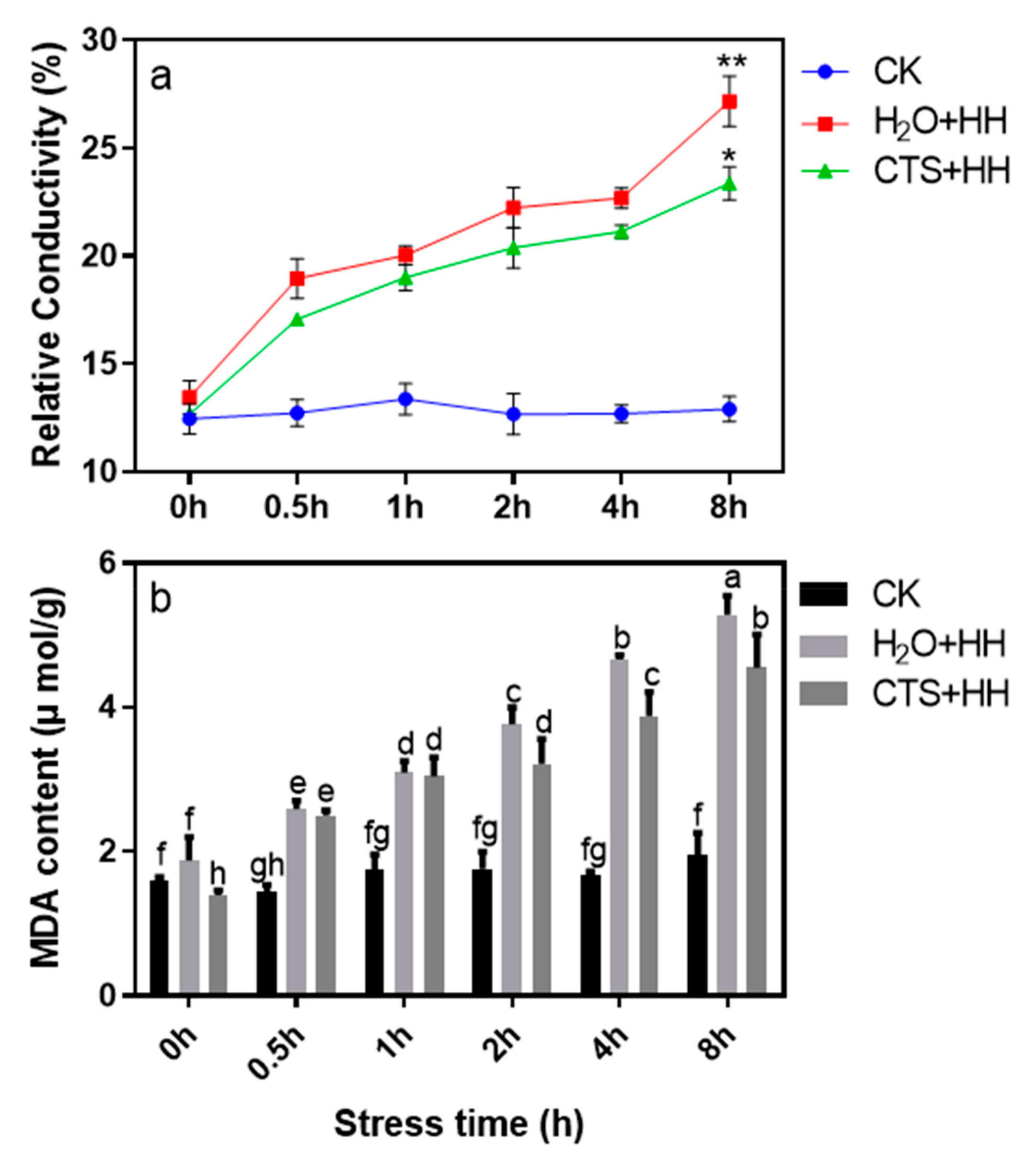
3.4. Effect of CTS on Membrane Peroxidation Level
3.5. Effect of CTS on the H2O2 and O2− Contents
3.6. Effect of CTS on the Proline Content
3.7. Effect of CTS on the AsA and GSH Contents and the AsA/DHA and GSH/GSSG Ratios
3.8. Effect of CTS on the Activities of Key Enzymes of the AsA-GSH Cycle
4. Discussion
4.1. Effect of CTS on the Photosynthesis of Strawberry Seedlings under High-Temperature and High-Light Stress
4.2. Effects of CTS on PSII Photoinhibition of Strawberry Seedlings under High-Temperature and High-Light Stress
4.3. Effects of CTS on Reactive Oxygen Species Metabolism in Strawberry Seedlings under High-Temperature and High-Light Stress
4.4. Effects of CTS on Membrane Peroxidation of Strawberry Seedlings under High-Temperature and High-Light Stress
4.5. Effects of CTS on the AsA-GSH Cycle in Strawberry Seedlings under High-Temperature High-Light Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tan, C.; Dai, H.; Lei, J. World strawberry production and trade status and development trend. World Agric. 2003, 5, 10–12, 40. [Google Scholar] [CrossRef]
- Gong, X.X. Research on Key Technologies of Strawberry Intensive Seedling Cultivation in High Temperature Season; Anhui Institute of Science and Technology: Huainan, China, 2016. [Google Scholar]
- Chen, W.Y. Key techniques for raising strawberry stolon seedlings. Northwest Hortic. 2021, 3, 23–24. [Google Scholar]
- Xu, K.; Guo, Y.; Zhang, S.; Zhou, H.; Zheng, Y. Response and mechanism of strawberry leaf photosynthesis to strong light. J. Appl. Ecol. 2005, 16, 73–78. [Google Scholar]
- Wang, M.; Zhang, C.C.; Du, Y.P. Effect of spraying kaolin on the photosynthetic characteristics of Forg grape leaves under high temperature and strong light stress. Chin. Fruit 2022, 1, 57–61, 67. [Google Scholar] [CrossRef]
- Dong, X.X.; Jin, H.H.; Wang, H.B. Research progress on the mechanism of plant photosystem adaptation to high light. J. Plant Physiol. 2016, 52, 1725–1732. [Google Scholar]
- Powles, S.B. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 1984, 35, 15–44. [Google Scholar] [CrossRef]
- Gan, Y.G.; Lian, J. Study on the effect of extracting chlorophyll from leaves of Aurantia chinensis with various extracts. Trop. Agric. Sci. 2021, 41, 86–91. [Google Scholar]
- Qin, S.H.; Li, L.L.; Chen, N.N. Effects of exogenous Ca2+ on morphological and photosynthetic characteristics and chlorophyll fluorescent parameters of squash seedlings under high temperature and strong light stress. J. Appl. Ecol. 2010, 21, 2830–2835. [Google Scholar]
- El Knidri, H.; Belaabed, R.; Addaou, A.; Laajeb, A.; Lahsini, A. Extraction, chemical modification and characterization of chitin and chitosan. Int. J. Biol. Macromol. 2018, 120, 1181–1189. [Google Scholar] [CrossRef]
- Jiang, X.S.; Mo, H.T.; Su, H.J.; Zhang, X.Y. Application of chitin and chitosan in agriculture. Chin. Agric. Sci. Bull. 2013, 29, 170–174. [Google Scholar]
- Zhu, Y.L.; Gu, D.L.; Wang, W.Z.; Du, X.F.; Yang, W.F.; Sun, A.X. Effect of chitosan on cold resistance of rice seedlings. Jiangsu Agric. Sci. 2017, 45, 66–68. [Google Scholar]
- He, H.; Li, D.D.; Pan, F.F.; Wu, Z.; Yang, S.Y. Effects of chitosan priming on rice seed germination and seedling growth under salt stress. J. Jiangxi Agric. Univ. 2022, 44, 1066–1074. [Google Scholar]
- Chen, Y.; Wang, T.; Liao, S.; Huang, Y.; Wu, B.; Kang, Y. Effects of exogenous chitosan on the growth of cucumber seedlings under high temperature stress. Jiangsu Agric. Sci. 2019, 47, 142–146. [Google Scholar]
- Endang, K.; Ramadhani, A.N.; Putri, N.G.A.; Damara, V.C.J. Chlorophyll extraction methods review and chlorophyll stability of katuk leaves (Sauropus androgynous). J. Phys. Conf. Ser. 2021, 1858, 012015. [Google Scholar]
- Tao, L.U.; Lu, J.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Protective roles of D1 protein turnover and the xanthophyll cycle in tomato under sub-high temperature and high light stress. Front. Agric. Sci. Eng. 2021, 8, 262–279. [Google Scholar]
- Lin, Z.F.; Li, S.S.; Lin, G.Z.; Guo, J.Y. Relationship between accumulation of H2O2 in senescent leaves and chloroplasts and membrane lipid peroxidation. J. Plant Physiol. 1988, 1, 16–22. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Zhang, R.; Xu, C.; Bao, Z.; Xiao, R.; Chen, X.; Xiao, W.; Li, D.; Fu, X.; Yang, C.; Li, L. Auxin alters sodium ion accumulation and nutrient accumulation by playing protective role in salinity challenged strawberry. Plant Physiol. Biochem. 2021, 164, 1–9. [Google Scholar] [CrossRef]
- Sun, L. The Mitigation Effect of Exogenous Selenium on Strawberry Seedlings under Low Temperature Stress and Its Effect on AsA-GSH Cycle; Zhejiang University: Zhejiang, China, 2016. [Google Scholar]
- Faik, A.; Popova, A.; Velitchkova, M. Effects of long-term action of high temperature and high light on the activity and energy interaction of both photosystems in tomato plants. Photosynthetica 2016, 54, 611–619. [Google Scholar] [CrossRef]
- Zhang, S.B.; Deng, Q.; Hao, Y. Effects of high temperature and high light on photosystem II activity in leaves of two Bauhinia species. Photosynthetica 2019, 57, 1094–1099. [Google Scholar] [CrossRef]
- Gang, S.; Liu, Y.F.; Lu, T.; Qi, M.F.; Guan, X.X.; Liu, Y.; Li, T.L. The effects of abscisic acid on photosynthesis in tomato under sub-high temperature and high light stress. J. Comput. Theor. Nanosci. 2016, 13, 7189–7198. [Google Scholar] [CrossRef]
- Chen, F.; Yang, S.L.; Zhang, L.; Tong, Y.X.; Zhang, Z.H.; Yu, F.F.; Jia, L.Y. Effects of exogenous methyl jasmonate on the AsA-GSH cycle of maize seedlings under salt stress. Biol. Bull. 2021, 56, 44–48. [Google Scholar]
- Yue, Y.; Luo, X. Effects of high temperature stress on cotton chlorophyll. Agric. Technol. 2022, 42, 34–37. [Google Scholar] [CrossRef]
- Wei, X.L.; Feng, C.Q.; Huang, Y.X.; Xu, T.L.; Qiu, F.X.; Wu, Q.; Zheng, Y.J.; Li, W.Q.; He, H.Q. Magnesium alleviates high temperature and strong light on photosynthetic physiological damage in tobacco plants. Fujian Agric. J. 2022, 37, 33–41. [Google Scholar] [CrossRef]
- Ma, P.F.; Li, L.; Yang, Y.; Zhao, H.; Fu, X.; Zhang, C. Effects of salicylic acid on D1 protein phosphorylation and PS II performance in wheat leaf chloroplasts under high temperature and high light stress. J. Appl. Ecol. 2008, 19, 2632–2636. [Google Scholar]
- Zhao, H.Z.; Guo, H.J.; Zhao, L.S.; Gu, J.L.; Zhao, S.R.; Li, J.H.; Liu, L.X. Major agronomic traits and photosynthetic properties of spatial environmentally mutagenic wheat chlorophyll deletion mutants. J. Crop 2011, 37, 119–126. [Google Scholar]
- Xu, Y.F.; Fu, J.; Chu, X.; Sun, Y.; Zhou, H.; Hu, T. Nitric oxide mediates abscisic acid induced light-tolerance in leaves of tall fescue under high-light stress. Sci. Hortic. 2013, 162, 1–10. [Google Scholar] [CrossRef]
- Yu, Q.L.; Sun, W.; Han, Y.; Hao, J.; Qin, X.; Liu, C.; Fan, S. Exogenous spermidine improves the sucrose metabolism of lettuce to resist high-temperature stress. Plant Growth Regul. 2022, 96, 497–509. [Google Scholar] [CrossRef]
- Li, T.L.; Lu, T.; Liu, Y.F.; Qi, M.F.; Sun, Z.P. Mechanism of PSI of PS and photoinhibition in higher plants. J. Shenyang Agric. Univ. 2016, 47, 513–519. [Google Scholar]
- Chen, M.M.; Nie, G.P.; Li, X.; Zhou, L.; Yang, L.Y.; Zhang, Y.C. Analysis of chlorophyll fluorescence under natural high temperature. Chin. Agric. Bull. 2022, 38, 86–95. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Wang, C.; Guo, Y.; Zhang, W.W. Effect of exogenous chitosan on antioxidant system of vegetable soybean chloroplast under NaCl stress. J. Plant Nutr. Fertil. 2016, 22, 1356–1365. [Google Scholar]
- Qu, D.Y.; Gu, W.R.; Zhang, L.G.; Chen, X.C.; Li, J.; Li, C.F.; Li, S. Effects of chitosan on the activities of antioxidant enzymes and endogenous hormones in maize seedling roots under cadmium stress. J. Northwest Bot. 2017, 37, 719–727. [Google Scholar]
- Deng, T.; Zhang, J.L.; Feng, J.N.; Wang, D. Chitosan stimulates antifreeze response of walnut under low temperature stress. J. Northwest For. Univ. 2016, 31, 60–64. [Google Scholar]
- Wang, G.P.; Xue, X.M.; Lu, C.; Nie, P.X.; Wang, J.Z. The effect and mechanism of betaine on improving plant stress resistance. Jiangxi Agric. J. 2014, 26, 22–26. [Google Scholar] [CrossRef]
- Yan, D. Study on Physiological Indexes and Membrane Damage of Tomato Seedlings under Low Temperature Stress by Exogenous Substances; Northeast Agricultural University: Harbin, China, 2007. [Google Scholar]
- Peng, C.C.; Zhang, G.H. Research progress on the relationship between biofilm and plant resistance. J. Hubei Univ. Nat. 1998, 6, 5–10. [Google Scholar]
- Deng, T.; Zhang, J.L.; Feng, J.N.; Wang, D. Chitosan on walnut freezing response under cold stress. J. Northwest For. Coll. 2016, 31, 60–64. [Google Scholar]
- Jing, Z.Z.; Zhu, H.; Zhu, H.; Tao, Y.; Liu, C.; Liu, J.; Yang, F.; Li, M. Exogenous ABA enhances the antioxidant defense system of maize by regulating the AsA-GSH cycle under drought stress. Sustainability 2022, 14, 3071. [Google Scholar] [CrossRef]
- Lv, X.M.; Yang, Y.; Jin, J.; Bai, R. Effects of CaCl2 on the AsA-GSH cycle of sour jujube seedlings under NaCl stress. Acta Hortic. Sin. 2017, 44, 953–962. [Google Scholar] [CrossRef]
- Lin, S.H.; Zhang, X.J.; Zhang, H.J.; Jia, H.L.; Pan, Y.; Luo, H.X. Research progress on redox defense system of fruits and vegetables. Chin. Fruits Veg. 2021, 41, 33–39. [Google Scholar] [CrossRef]
- Qin, S.Y.; Liu, H.; Nie, Z.; Gao, W.; Li, C.; Lin, Y.; Zhao, P. AsA–GSH cycle and antioxidant enzymes play important roles in Cd tolerance of wheat. Bull. Environ. Contam. Toxicol. 2018, 101, 684–690. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, E.; Zhang, Y.; Li, H.; Wang, X.; Zhang, H.; Xiao, W.; Chen, X.; Li, L. Chitosan Reduces Damages of Strawberry Seedlings under High-Temperature and High-Light Stress. Agronomy 2023, 13, 517. https://doi.org/10.3390/agronomy13020517
Fu E, Zhang Y, Li H, Wang X, Zhang H, Xiao W, Chen X, Li L. Chitosan Reduces Damages of Strawberry Seedlings under High-Temperature and High-Light Stress. Agronomy. 2023; 13(2):517. https://doi.org/10.3390/agronomy13020517
Chicago/Turabian StyleFu, Erting, Yuzheng Zhang, Huiling Li, Xiaozhe Wang, Hanxiao Zhang, Wei Xiao, Xiude Chen, and Ling Li. 2023. "Chitosan Reduces Damages of Strawberry Seedlings under High-Temperature and High-Light Stress" Agronomy 13, no. 2: 517. https://doi.org/10.3390/agronomy13020517