
Apple–Soybean Mixed Stand Increased Fine Root Distribution and Soil Water Content with Reduced Soil Nitrate Nitrogen

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
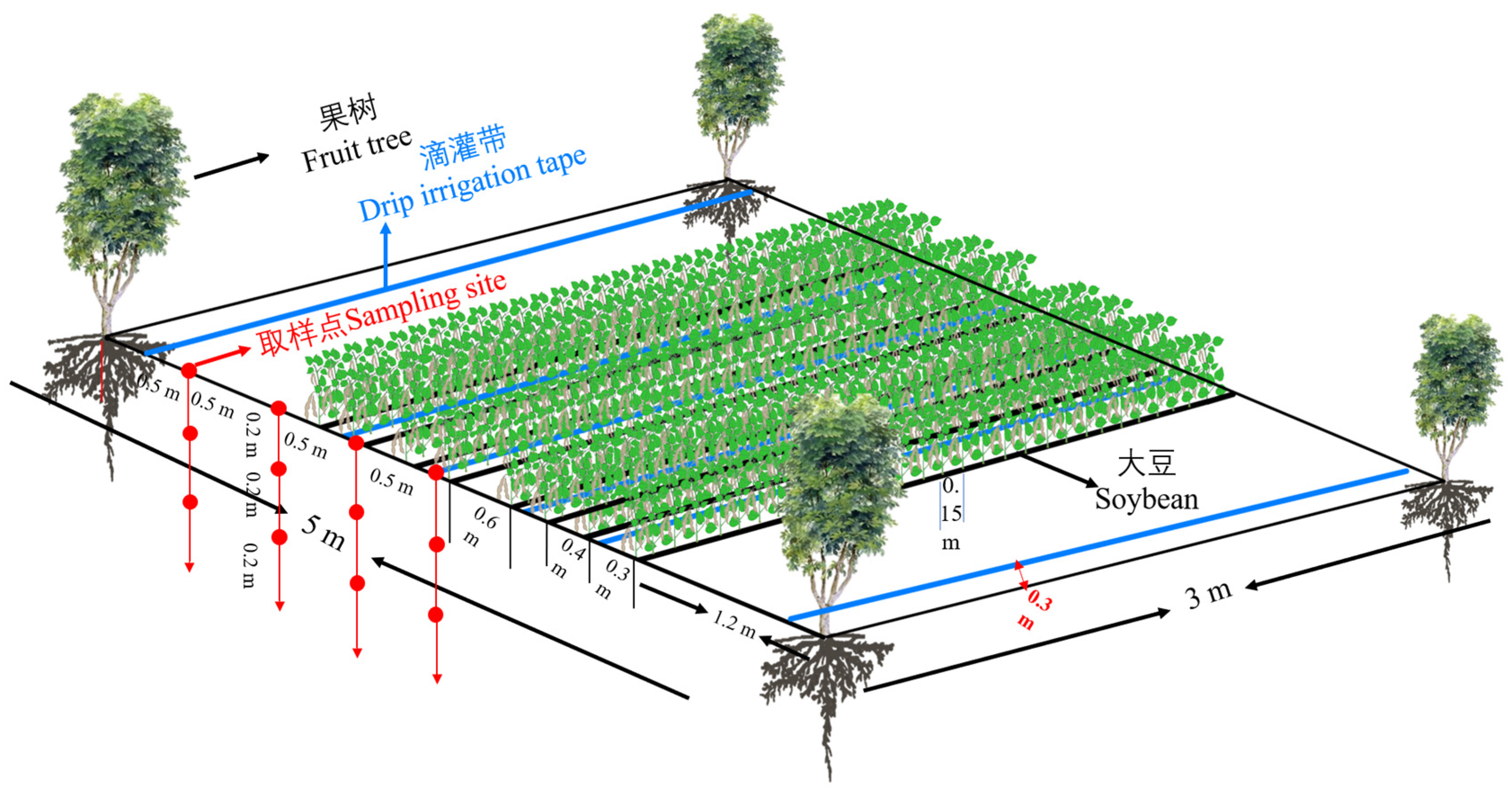
2.2. Study Site and Experimental Design
2.3. Collection of Root and Soil Samples
2.4. Determination of soybean Agronomic Characters
2.5. Fine root Analysis
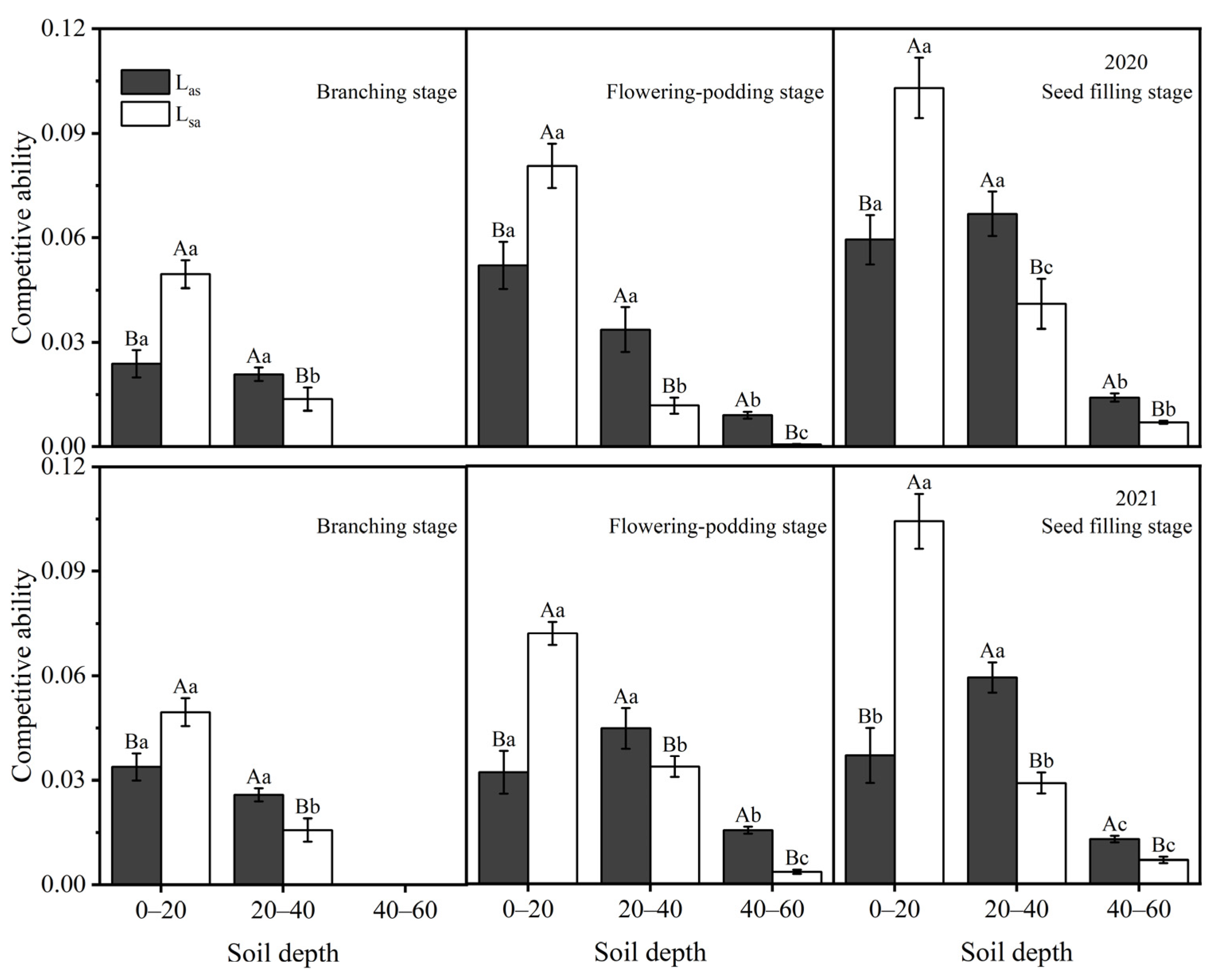
2.6. Competitive Ability
2.7. Soil Analysis
2.8. Yield and Land Equivalent Ratio (LER)
2.9. Statistical Analyses
3. Results
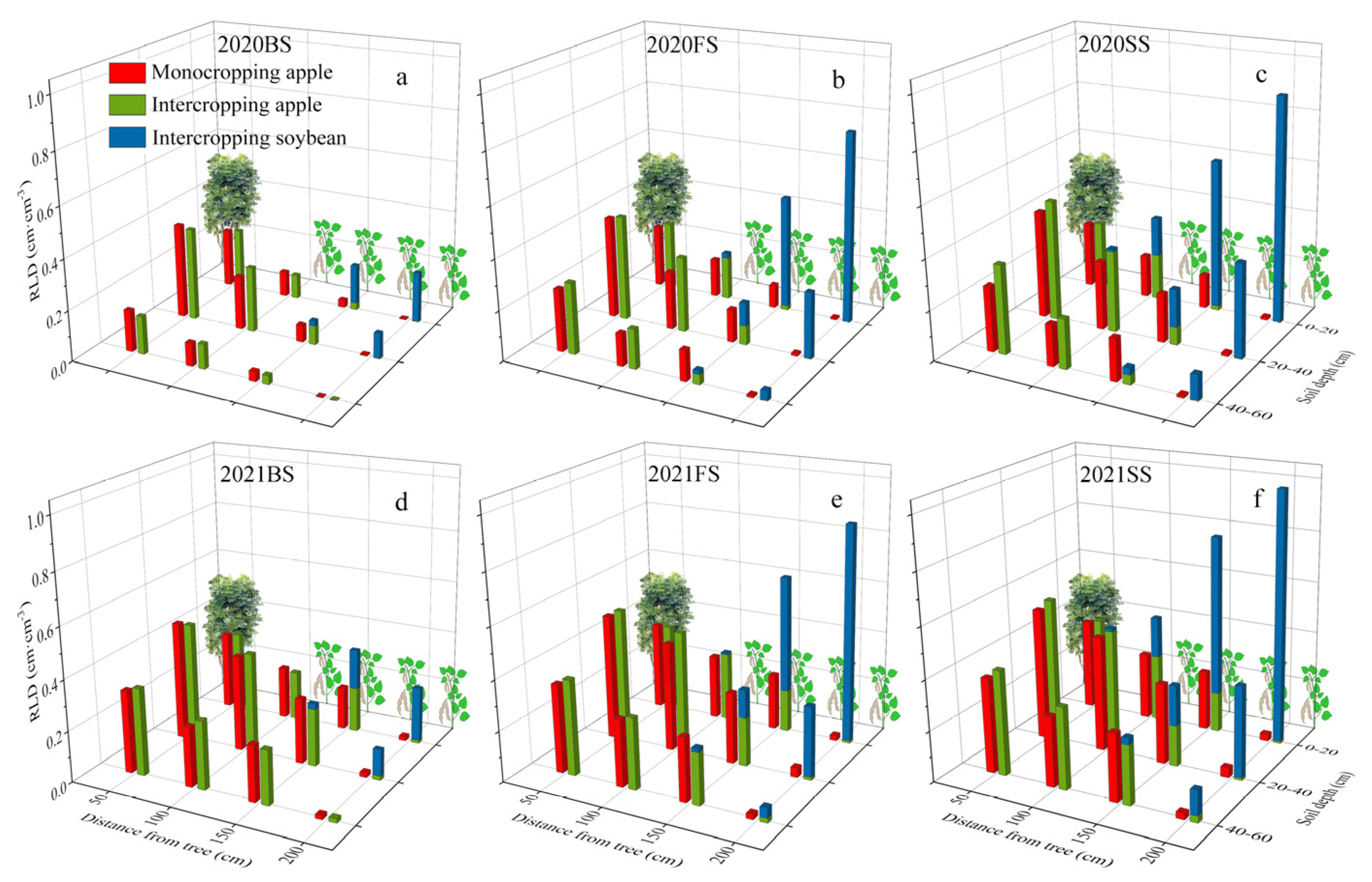
3.1. Fine Root Length Density Distribution in the Intercropping System
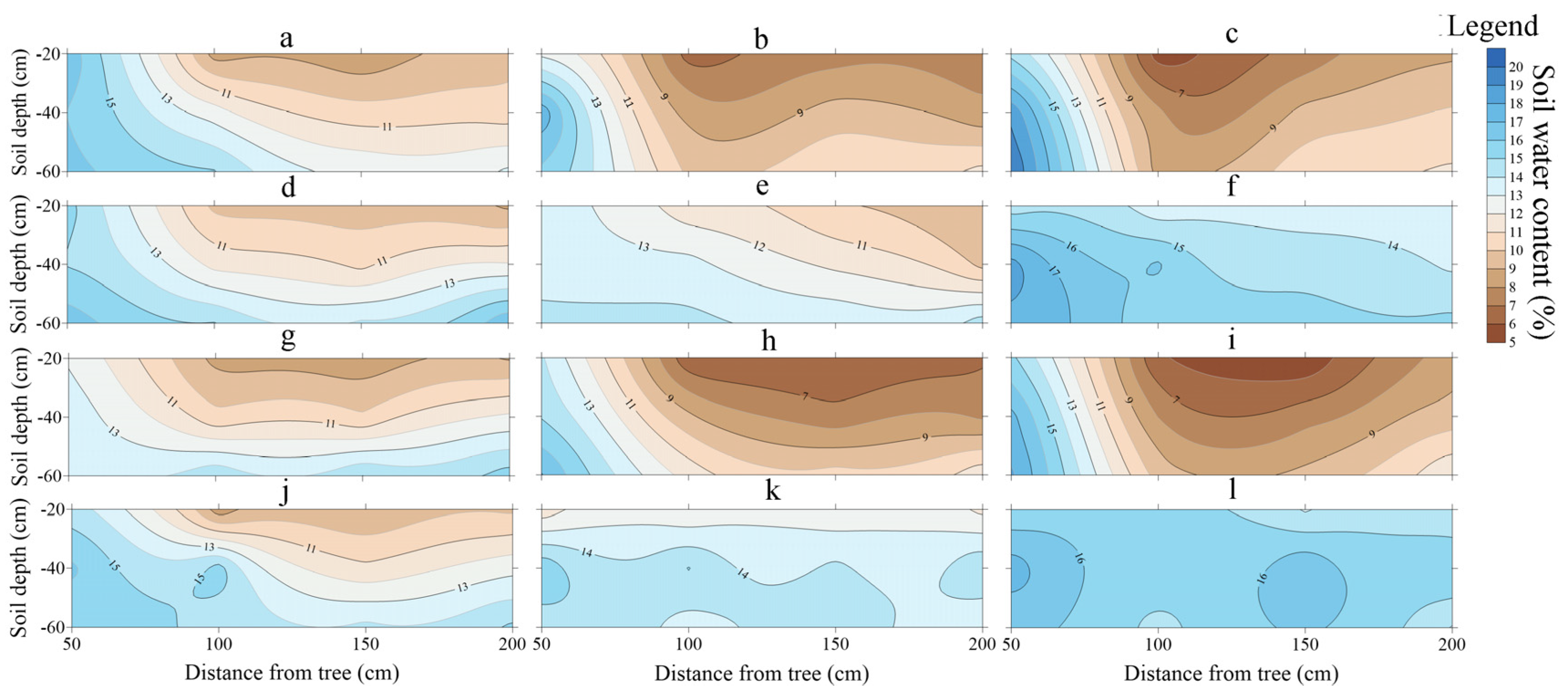
3.2. Soil Water Content Distribution
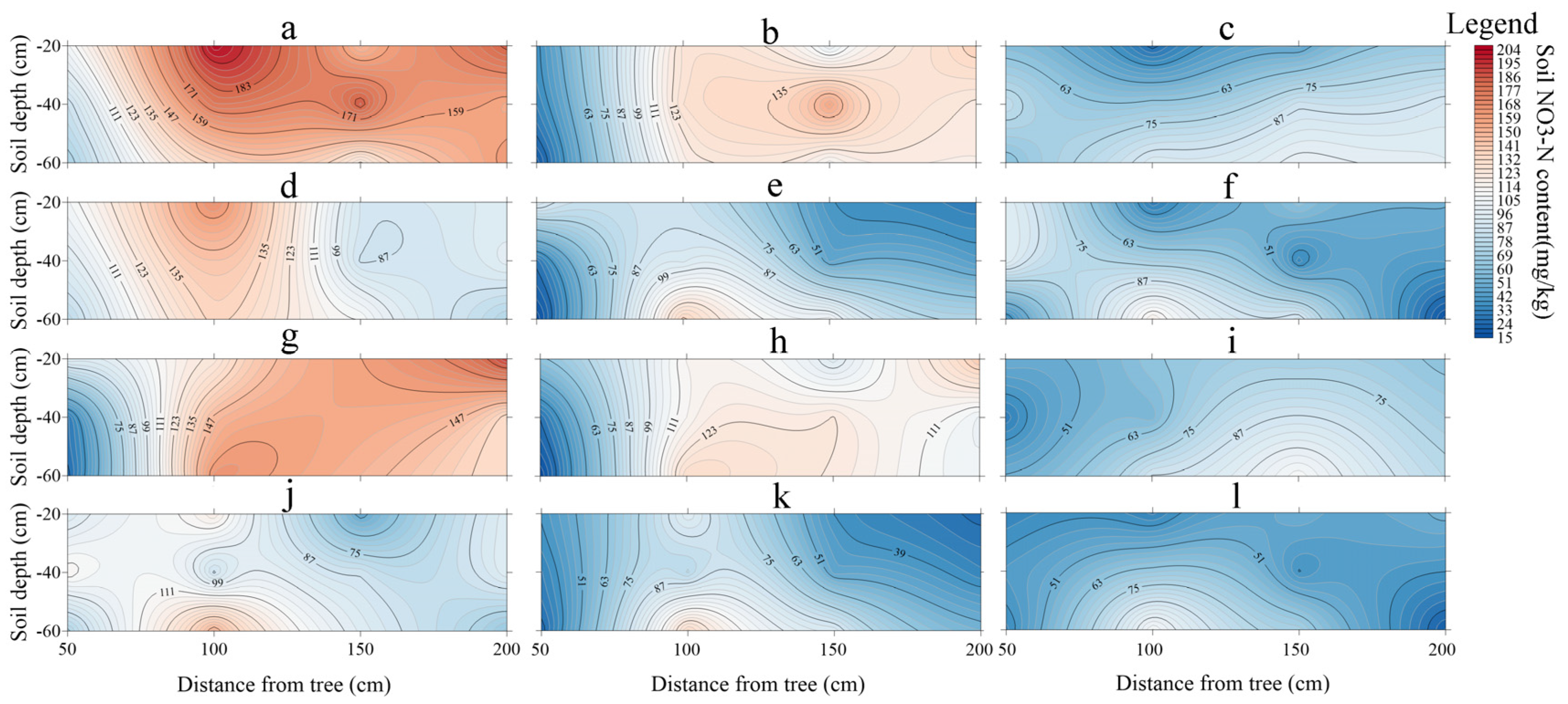
3.3. Soil NO3−−N Content Distribution
3.4. The Relationship between FRLD and Spatiotemporal Distribution of Soil Resources
3.5. Competitiveness Index for Apple–Soybean Intercropping Systems
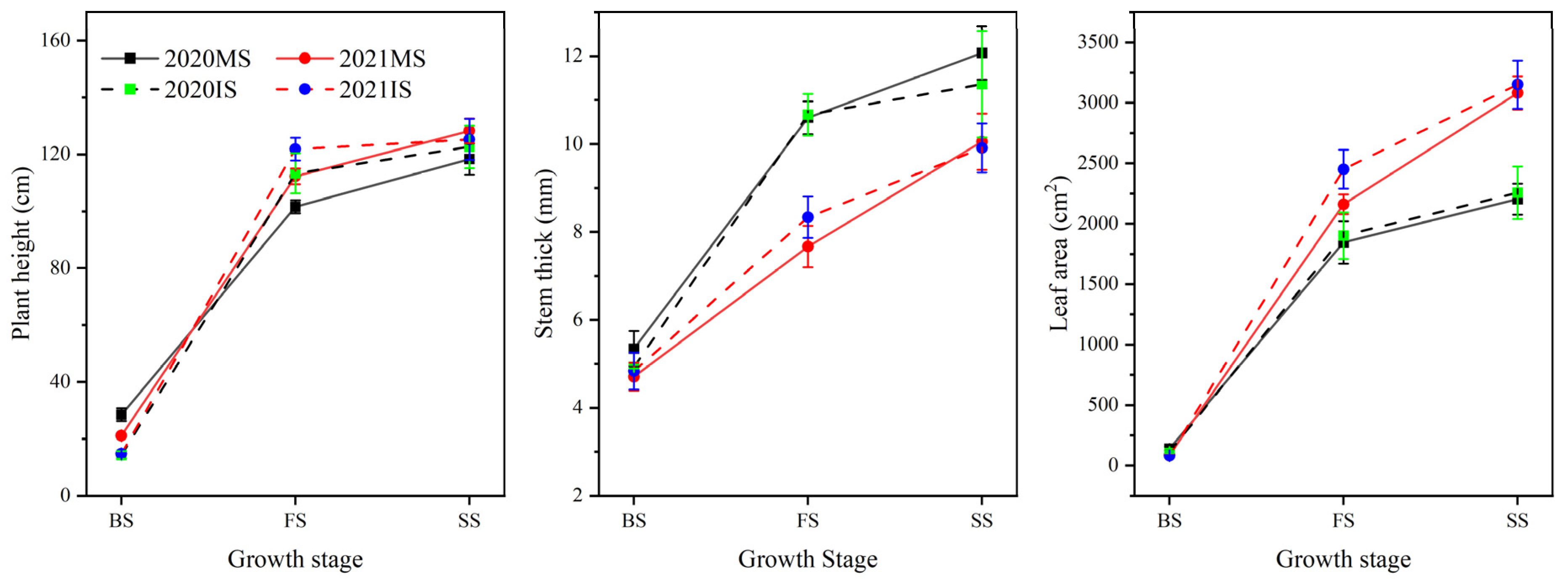
3.6. The Growth of Soybeans
3.7. System Yield and Yield Composition Factors
4. Discussion
4.1. The Distribution of Fine Roots
4.2. Distribution of Soil Water Content
4.3. Distribution of Soil NO3−-N Content
4.4. Interspecific Competition
4.5. Crop Growth and Yield
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Malézieux, E.; Crozat, Y.; Dupraz, C.; Laurans, M.; Makowski, D.; Ozier-Lafontaine, H.; Rapidel, B.; Tourdonnet, S.; Valantin-Morison, M. Mixing plant species in cropping systems: Concepts, tools and models. A review. Agron. Sustain. Dev. 2009, 29, 43–62. [Google Scholar] [CrossRef]
- Pan, R.Z. Plant Physiology, 5th ed.; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Gao, X.; Wu, P.; Zhao, X.; Wang, J.; Shi, Y. Effects of Land Use on Soil Moisture Variations in a Semi-Arid Catchment: Implications for Land and Agricultural Water Management. Land Degrad. Dev. 2014, 25, 163–172. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, P.; Gao, X.; Tian, L.; Li, H. Changes of soil hydraulic properties under early-stage natural vegetation recovering on the Loess Plateau of China. Catena 2014, 113, 386–391. [Google Scholar] [CrossRef]
- Lehmann, L.M.; Smith, J.; Westaway, S.; Pisanelli, A.; Russo, G.; Borek, R.; Sandor, M.; Gliga, A.; Smith, L.; Ghaley, B.B. Productivity and Economic Evaluation of Agroforestry Systems for Sustainable Production of Food and Non-Food Products. Sustainability 2020, 12, 5429. [Google Scholar] [CrossRef]
- Yang, J.F.; Zhang, T.R.; Wang, Y.J.; Zhang, R.Q.; Li, H.K. Intercropped relationship change the developmental pattern of apple and white clover. Bioengineered 2019, 10, 218–228. [Google Scholar] [CrossRef]
- Forey, O.; Temani, F.; Wery, J.; Jourdan, C.; Metay, A. Effect of combined deficit irrigation and grass competition at plantation on peach tree root distribution. Eur. J. Agron. 2017, 91, 16–24. [Google Scholar] [CrossRef]
- Fletcher, E.H.; Thetford, M.; Sharma, J.; Jose, S. Effect of root competition and shade on survival and growth of nine woody plant taxa within a pecan [Carya illinoinensis (Wangenh.) C. Koch] alley cropping system. Agrofor. Syst. 2012, 86, 49–60. [Google Scholar] [CrossRef]
- Abbasi, S.A.; Nazari, M.; Fallah, S.; Iranipour, R.; Mousavi, A. The competitive effect of almond trees on light and nutrients absorption, crop growth rate, and the yield in almond–cereal agroforestry systems in semi-arid regions. Agrofor. Syst. 2020, 94, 1111–1122. [Google Scholar] [CrossRef]
- Bouttier, L.; Paquette, A.; Messier, C.; Rivest, D.; Olivier, A.; Cogliastro, A. Vertical root separation and light interception in a temperate tree-based intercropping system of Eastern Canada. Agrofor. Syst. 2014, 88, 693–706. [Google Scholar] [CrossRef]
- Paul, C.; Weber, M.; Knoke, T. Agroforestry versus farm mosaic systems-Comparing land-use efficiency, economic returns and risks under climate change effects. Sci. Total Environ. 2017, 587, 22–35. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, W.; Chen, J.; Bruijnzeel, L.A.; Mao, Z.; Yang, X.; Cardinael, R.; Meng, F.R.; Sidle, R.C.; Seitz, S.; et al. Reductions in water, soil and nutrient losses and pesticide pollution in agroforestry practices: A review of evidence and processes. Plant Soil 2019, 453, 45–86. [Google Scholar] [CrossRef]
- Abbas, G.; Ali, A.; Khan, M.; Mahmood, H.Z.; Wahab, S.A.; Amir-ud-Din, R. The Transition from Arid Farming Systems to Agroforestry Systems in Pakistan: A Comparison of Monetary Returns. Small–Scale For. 2021, 20, 325–350. [Google Scholar] [CrossRef]
- Pratiwi, A.; Suzuki, A. Reducing Agricultural Income Vulnerabilities through Agroforestry Training: Evidence from a Randomised Field Experiment in Indonesia. Bull. Indones. Econ. Stud. 2018, 55, 83–116. [Google Scholar] [CrossRef]
- Gao, L.; Xu, H.; Bi, H.; Xi, W.; Bao, B.; Wang, X.; Bi, C.; Chang, Y. Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. PLoS ONE 2018, 8, e70739. [Google Scholar] [CrossRef]
- Kubo, S.; Fuku, Y.; Shimamoto, T.; Kuwayama, A.; Ohya, M.; Amano, H.; Maruo, T.; Goto, T.; Komiya, T.; Kadota, K. Vascular Injury Caused by Retrieval of Ruptured and Detached Balloon Valvuloplasty Catheter During Transcatheter Aortic Valve Replacement. JACC Cardiovasc. Interv. 2017, 10, 1593–1595. [Google Scholar] [CrossRef]
- Sun, Y.; Bi, H.; Xu, H.; Duan, H.; Peng, R.; Wang, J. Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy 2018, 8, 280–295. [Google Scholar] [CrossRef]
- Mulia, R.; Dupraz, C. Unusual Fine Root Distributions of Two Deciduous Tree Species in Southern France: What Consequences for Modelling of Tree Root Dynamics? Plant Soil 2006, 281, 71–85. [Google Scholar] [CrossRef]
- Noordwijk, M.V.; Cadisch, G.; Ong, C.K. Below-Ground Interactions in Tropical Agroecosystems Concepts and Models with Multiple Plant Components; CABI Pub: Wallingford, UK; Cambridge, MA, USA, 2004. [Google Scholar]
- Xia, H.Y.; Zhao, J.H.; Sun, J.H.; Bao, X.G.; Christie, P.; Zhang, F.S.; Li, L. Dynamics of root length and distribution and shoot biomass of maize as affected by intercropping with different companion crops and phosphorus application rates. Field Crops Res. 2013, 150, 52–62. [Google Scholar] [CrossRef]
- Baptista, I.; Ritsema, C.; Querido, A.; Ferreira, A.D.; Geissen, V. Improving rainwater-use in Cabo Verde drylands by reducing runoff and erosion. Geoderma 2015, 1, 237–297. [Google Scholar] [CrossRef]
- Yao, S.; Merwin, I.A.; Bird, G.W.; Abawi, G.S.; Thies, J.E. Orchard floor management practices that maintain vegetative or biomass groundcover stimulate soil microbial activity and alter soil microbial community composition. Plant Soil 2005, 271, 377–389. [Google Scholar] [CrossRef]
- Ling, Q.; Gao, X.; Zhao, X.; Huang, J.; Li, H.; Li, L.; Sun, W.; Wu, P. Soil water effects of agroforestry in rainfed jujube (Ziziphus jujube Mill.) orchards on loess hillslopes in Northwest China. Agr. Ecosyst. Environagr. Ecosyst. Environ. 2017, 247, 343–351. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, B.; Hill, R. Water use assessment in alley cropping systems within subtropical China. Agrofor. Syst. 2012, 84, 243–259. [Google Scholar] [CrossRef]
- Pavlidis, G.; Tsihrintzis, V.A. Environmental Benefits and Control of Pollution to Surface Water and Groundwater by Agroforestry Systems: A review. Water Resour Manag 2017, 32, 1–29. [Google Scholar] [CrossRef]
- Rodriguez, C.; Carlsson, G.; Englund, J.E.; Flöhr, A.; Pelzer, E.; Jeuffroy, M.H.; Makowski, D.; Jensen, E.S. Grain legume-cereal intercropping enhances the use of soil-derived and biologically fixed nitrogen in temperate agroecosystems. A meta-analysis. Eur. J. Agron. 2020, 118, 126077. [Google Scholar] [CrossRef]
- Zhang, D.; Du, G.; Sun, Z.; Bai, W.; Wang, Q.; Feng, L.; Zheng, J.; Zhang, Z.; Liu, Y.; Yang, S.; et al. Agroforestry enables high efficiency of light capture, photosynthesis and dry matter production in a semi-arid climate. Eur. J. Agron. 2018, 94, 1–11. [Google Scholar] [CrossRef]
- Phiri, A.T.; Njira, K. Grain Legume-Based Cropping Systems’ Effects on Soil Organic Carbon and Nutrient Dynamics. Agric. Res. 2022, 12, 45–52. [Google Scholar] [CrossRef]
- Chamkhi, I.; Cheto, S.; Geistlinger, J.; Zeroual, Y.; Kouisni, L.; Bargaz, A.; Ghoulam, C. Legume-based intercropping systems promote beneficial rhizobacterial community and crop yield under stressing conditions. Ind. Crops Prod. 2022, 183, 114958. [Google Scholar] [CrossRef]
- Muoni, T.; Jonsson, M.; Duncan, A.J.; Watson, C.A.; Bergkvist, G.; Barnes, A.P.; Oborn, I. Effects of management practices on legume productivity in smallholder farming systems in sub-Saharan Africa. Food Energy Secur. 2022, 11, e366. [Google Scholar] [CrossRef]
- Chen, C.; Liu, W.; Wu, J.; Jiang, X.; Zhu, X. Can intercropping with the cash crop help improve the soil physico-chemical properties of rubber plantations? Geoderma 2019, 335, 149–160. [Google Scholar] [CrossRef]
- Li, L.P.; Hai, Y.; Anwar, M.; Tang, Z.Y.; Fang, J.Y. Community Structure and Conservation of Wild Fruit Forests in the Ili Valley, Xinjiang. Arid Zone Res. 2011, 28, 7–13. (In Chinese) [Google Scholar]
- Smit, A.L. Root Methods : A Handbook; Springer: Berlin, Germany; New York, NY, USA, 2000. [Google Scholar]
- Wu, F.; Liao, B.W.; Chen, Y.J.; Jiang, Z.M.; Guo, Y.P.; Li, M. Niches of nine mangrove species in a Sonneratia apetala-colonized area of Dongzhai Harbor, Hainan Island, China. Ecol. Evol. 2020, 10, 11838–11846. [Google Scholar] [CrossRef]
- Lu, R.K. Soil Agrochemical Analysis Methods; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Tofinga, M.P.; Paolini, R.; Snaydon, R.W. A study of root and shoot interactions between cereals and peas in mixtures. J. Agr. Sci. 1993, 120, 13–24. [Google Scholar] [CrossRef]
- Seran, T.H.; Brintha, I. Review on maize based intercropping. J. Agron. Crops Sci. 2010, 9, 135–145. [Google Scholar]
- Yang, C.; Chai, Q.; Huang, G. Root distribution and yield responses of wheat/maize intercropping to alternate irrigation in the arid areas of Northwest China. Plant Soil Environ. 2010, 56, 253–262. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.L.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Li, X.; Šimůnek, J.; Shi, H.; Yan, J.; Peng, Z.; Gong, X. Spatial distribution of soil water, soil temperature, and plant roots in a drip-irrigated intercropping field with plastic mulch. Eur. J. Agron. 2017, 83, 47–56. [Google Scholar] [CrossRef]
- Forey, O.; Metay, A.; Bertrand, N.; Wery, J. Two years old peach trees intercropped with a grass mixture can grow through a lithic discontinuity to access water the grass cannot. Agrofor. Syst. 2021, 95, 151–163. [Google Scholar] [CrossRef]
- Yun, L.; Bi, H.; Gao, L.; Zhu, Q.; Ma, W.; Cui, Z.; Wilcox, B.P. Soil Moisture and Soil Nutrient Content in Walnut-Crop Intercropping Systems in the Loess Plateau of China. Arid Land Res. Manag. 2012, 26, 285–296. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qiu, K.; López-Vicente, M.; Shen, W.; Wu, G.L. Soil-water deficit in deep soil layers results from the planted forest in a semi-arid sandy land: Implications for sustainable agroforestry water management. Agr. Water Manag. 2021, 254, 106985. [Google Scholar] [CrossRef]
- Bosi, C.; Pezzopane, J.R.M.; Sentelhas, P.C. Soil water availability in a full sun pasture and in a silvopastoral system with eucalyptus. Agrofor. Syst. 2019, 94, 429–440. [Google Scholar] [CrossRef]
- Thierfelder, C.; Chivenge, P.; Mupangwa, W.; Rosenstock, T.S.; Lamanna, C.; Eyre, J.X. How climate-smart is conservation agriculture (CA)?—Its potential to deliver on adaptation, mitigation and productivity on smallholder farms in southern Africa. Food Secur. 2017, 9, 537–560. [Google Scholar] [CrossRef]
- Rachman, A.; Anderson, S.H.; Gantzer, C.J.; Alberts, E.E. Soil hydraulic properties influenced by stiff-stemmed grass hedge systems. Soil Sci. Soc. Am. J. 2004, 68, 1386–1393. [Google Scholar] [CrossRef]
- Walsh, B.D.; Salmins, S.; Buszard, D.J.; Mackenzie, A.F. Impact of soil management systems on organic dwarf apple orchards and soil aggregate stability bulk density, temperature and water content. Can. J. Soil Sci. 1996, 76, 203–209. [Google Scholar] [CrossRef]
- Hanson, B.; May, D. Effect of subsurface drip irrigation on processing tomato yield, water table depth, soil salinity, and profitability. Agric. Water Manag. 2004, 68, 1–17. [Google Scholar] [CrossRef]
- El-Nesr, M.N.; Alazba, A.A.; Šimůnek, J. HYDRUS simulations of the effects of dual-drip subsurface irrigation and a physical barrier on water movement and solute transport in soils. Irrig. Sci. 2014, 32, 111–125. [Google Scholar] [CrossRef]
- Campi, P.; Palumbo, A.D.; Mastrorilli, M. Effects of tree windbreak on microclimate and wheat productivity in a Mediterranean environment. Eur. J. Agron. 2009, 30, 220–227. [Google Scholar] [CrossRef]
- Lin, B.B. The role of agroforestry in reducing water loss through soil evaporation and crop transpiration in coffee agroecosystems. Agr. Forest Meteorol. 2010, 150, 510–518. [Google Scholar] [CrossRef]
- Teodoro, A.V.; Sousa-Souto, L.; Klein, A.M.; Tscharntke, T. Seasonal contrasts in the response of coffee ants to agroforestry shade-tree management. Environ. Entomol. 2010, 39, 1744–1750. [Google Scholar] [CrossRef]
- Schwab, N.; Schickhoff, U.; Fischer, E. Transition to agroforestry significantly improves soil quality: A case study in the central mid-hills of Nepal. Agr. Ecosyst. Environ. 2015, 205, 57–69. [Google Scholar] [CrossRef]
- Fredes, I.; Moreno, S.; Díaz, F.P.; Gutiérrez, R.A. Nitrate signaling and the control of Arabidopsis growth and development. Curr. Opin. Plant Biol. 2019, 47, 112–118. [Google Scholar] [CrossRef]
- Miller, A.J.; Cramer, M.D. Root Nitrogen Acquisition and Assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Zhu, L.X.; Zhang, W.J. Effects of Controlled-Release Urea Combined with Conventional Urea on Nitrogen Uptake, Root Yield, and Quality of Platycodon Grandiflorum. J. Plant Nutr. 2017, 40, 662–672. [Google Scholar] [CrossRef]
- Fan, P.P.; Jiang, Y.X. Nitrogen dynamics differed among the first six root branch orders of Fraxinus mandshurica and Larix gmelinii during short-term decomposition. J. Plant Res. 2010, 123, 433–438. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; Neergaard, A.; Jensen, L.S.; Luxhøi, J.; Jensen, C.R. Alternate partial root-zone irrigation induced dry/wet cycles of soils stimulate N mineralization and improve N nutrition in tomatoes. Plant Soil 2010, 337, 167–177. [Google Scholar] [CrossRef]
- Ramachandran, N.P.K.; Mohan, K.B.; Nair, V.D. Agroforestry as a strategy for carbon sequestration. J. Plant Nutr. Soil 2009, 172, 10–23. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Rao, M. Modeling of waterflow and nitrate transport under surface drip fertigation. Trans. Asae 2005, 48, 627–637. [Google Scholar]
- Kahlon, M.S.; Josan, A.S.; Khera, K.L. A comparative study on soil and plant parameters under furrow and drip irrigated tomatoes with poor quality water. Res. Crops 2008, 9, 62–72. [Google Scholar]
- Li, J.S.; Zhang, J.J.; Rao, M.J. Simulation of water and nitrate transport under drip irrigation using HYDRUS-2D-model verification. In Proceedings of the 7th International Conference on Environment and Water, Beijing, China, 11–14 October 2004; pp. 221–229. [Google Scholar]
- Wang, Z.; Li, J.; Li, Y. Effects of drip irrigation system uniformity and nitrogen applied on deep percolation and nitrate leaching during growing seasons of spring maize in semi-humid region. Irrig. Sci. 2014, 32, 221–236. [Google Scholar] [CrossRef]
- Yang, B.; Meng, X.; Singh, A.K.; Wang, P.; Song, L.; Zakari, S.; Liu, W. Intercrops improve surface water availability in rubber-based agroforestry systems. Agric. Ecosyst. Environ. 2020, 298, 106937. [Google Scholar] [CrossRef]
- Lopez-Diaz, M.L.; Rolo, V.; Moreno, G. Trees’ role in nitrogen leaching after organic, mineral fertilization: A greenhouse experiment. J. Environ. Qual. 2011, 40, 853–859. [Google Scholar] [CrossRef]
- Livesley, S.J.; Gregory, P.J.; Buresh, R.J. Competition in tree row agroforestry systems. 1. Distribution and dynamics of fine root length and biomass. Plant Soil 2000, 227, 149–161. [Google Scholar] [CrossRef]
- Xu, H.S.; Bi, H.X.; Xi, W.M.; Powell, R.L.; Gao, L.B.; Yu, L. Root Distribution Variation of Crops under Walnut-Based Intercropping Systems in the Loess Plateau of China. Pak. J. Agr. Sci. 2014, 51, 773–778. [Google Scholar]
- Sgarbossa, J.; Elli, E.F.; Schwerz, F.; Nardini, C.; Miguel, K.F.; Schmidt, D.; Dal, A.; Caron, B.O. Bean–soybean succession under full sun and in agroforestry systems: Impacts on radiation use efficiency, growth and yield. J. Agron. Crops Sci. 2020, 207, 362–377. [Google Scholar] [CrossRef]
- Wu, Y.; Gong, W.; Yang, W. Shade Inhibits Leaf Size by Controlling Cell Proliferation and Enlargement in Soybean. Sci. Rep. 2017, 7, 9259. [Google Scholar] [CrossRef]
- Caron, B.O.; Sgarbossa, J.; Schwerz, F.; Elli, E.F.; Eloy, E.; Behling, A. Dynamics of solar radiation and soybean yield in agroforestry systems. An. Acad. Bras. Ciências 2018, 90, 3799–3812. [Google Scholar] [CrossRef]
- Ling, Q.; Zhao, X.; Wu, P.; Gao, X.; Sun, W. Effect of the fodder species canola (Brassica napus L.) and daylily (Hemerocallis fulva L.) on soil physical properties and soil water content in a rainfed orchard on the semiarid Loess Plateau, China. Plant Soil 2019, 453, 209–228. [Google Scholar] [CrossRef]
- Liu, C.Q. Challenges and Countermeasures for the supply security of important agricultural products in China. Econ. Asp. 2021, 5, 61–73. (In Chinese) [Google Scholar]
- Chen, Y.F.; Wang, J.Y.; Zhang, F.G. Globalization and the new pattern of food security. J. Nat. Resour. 2021, 36, 1362–1380. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Index | FRLD | SWC | SNC | SD | HD | ST |
|---|---|---|---|---|---|---|---|
| MA | FRLD | 1 | |||||
| SWC | 0.497 ** | 1 | |||||
| SNC | −0.477 ** | −0.280 ** | 1 | ||||
| SD | 0.044 | −0.32 | 0.439 ** | 1 | |||
| HD | −0.871 ** | 0.484 ** | −0.535 ** | 0 | 1 | ||
| ST | 0.177 ** | 0.357 ** | −0.132 | 0 | 0 | 1 | |
| AS | FRLD | 1 | |||||
| SWC | 0.452 ** | 1 | |||||
| SNC | 0.168 * | −0.115 | 1 | ||||
| SD | 0.119 | 0.490 ** | 0.114 | 1 | |||
| HD | −0.854 ** | −0.342 ** | −0.167 * | 0 | 1 | ||
| ST | 0.165 * | 0.283 ** | −0.373 ** | 0 | 0 | 1 |
| Treatment | Model | Nonstandardized Coefficient | Standardized Coefficient | t | Sig. | Multicol Linearity | Model Parameters | |||
|---|---|---|---|---|---|---|---|---|---|---|
| B | Standard Error | Toleance | VIF | R2 | Sig. | |||||
| MA | Constant | 0.301 | 0.028 | 10.666 | 2 × 10−21 | 0.7897 | 4.2199 × 10−71 | |||
| SWC | 0.003 | 0.001 | 0.080 | 2.125 | 0.035 | 0.689 | 1.451 | |||
| SNC | 0.0000533 | 0.000 | 0.019 | 0.495 | 0.621 | 0.632 | 1.582 | |||
| HD | −0.097 | 0.005 | −0.837 | −20.503 | 4 × 10−52 | 0.587 | 1.705 | |||
| ST | 0.015 | 0.003 | 0.194 | 5.570 | 8 × 10−8 | 0.806 | 1.241 | |||
| AS | Constant | 0.173 | 0.033 | 5.218 | 4 × 10−7 | 0.7789 | 8.11 × 10−69 | |||
| SWC | 0.008 | 0.002 | 0.152 | 4.220 | 4 × 10−5 | 0.798 | 1.254 | |||
| SNC | 0.0003897 | 0.000 | 0.117 | 3.308 | 0.001 | 0.828 | 1.208 | |||
| HD | −0.086 | 0.004 | −0.782 | −22.345 | 2 × 10−57 | 0.839 | 1.192 | |||
| ST | 0.012 | 0.003 | 0.165 | 4.613 | 7 × 10−6 | 0.802 | 1.247 | |||
| Years | Treatment | Plant Number · ha−1 Trees Number · ha−1 | No. of Pods·Plant−1 No. of Fruits·Plant−1 | No. of Seed·Pod−1 | 100-Grain Weight (g) Single Fruit Weight (g) | Yield (kg · ha−1) |
|---|---|---|---|---|---|---|
| 2020 | Monocropping soybean | 2.4 × 105 | 40.63 ± 7.96 a | 2.3 ± 0.80 a | 29.74 ± 0.76 b | 6636.51 ± 590.56 b |
| Intercropping soybean | 1.1 × 105 | 44.2 ± 12.38 a | 2.96 ± 0.90 b | 26.50 ± 0.68 a | 3759.91 ± 338.53 a | |
| 2021 | Monocropping soybean | 2.4 × 105 | 40.74 ± 7.85 a | 2.47 ± 0.80 a | 30.04 ± 0.88 b | 7243.20 ± 623.99 b |
| Intercropping soybean | 1.1 × 105 | 44.7 ± 10.44 a | 2.88 ± 0.94 a | 26.9 ± 0.87 a | 3815.04 ± 349.07 a | |
| Monocropping apple | 675 | 8.44 ± 1.06 a | 227.62 ± 14.43 a | 1296.75 ± 163.34 a | ||
| Intercropping apple | 675 | 7.94 ± 0.95 a | 217.64 ± 10.95 a | 1148.44 ± 97.82 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Wang, X.; Liu, T.; Wei, W.; Zhang, S.; Zhu, Y.; Tuerti, T.; Li, L.; Zhang, W. Apple–Soybean Mixed Stand Increased Fine Root Distribution and Soil Water Content with Reduced Soil Nitrate Nitrogen. Agronomy 2023, 13, 548. https://doi.org/10.3390/agronomy13020548
Shen L, Wang X, Liu T, Wei W, Zhang S, Zhu Y, Tuerti T, Li L, Zhang W. Apple–Soybean Mixed Stand Increased Fine Root Distribution and Soil Water Content with Reduced Soil Nitrate Nitrogen. Agronomy. 2023; 13(2):548. https://doi.org/10.3390/agronomy13020548
Chicago/Turabian StyleShen, Lei, Xiuyuan Wang, Tingting Liu, Wenwen Wei, Shuai Zhang, Yun Zhu, Tayir Tuerti, Luhua Li, and Wei Zhang. 2023. "Apple–Soybean Mixed Stand Increased Fine Root Distribution and Soil Water Content with Reduced Soil Nitrate Nitrogen" Agronomy 13, no. 2: 548. https://doi.org/10.3390/agronomy13020548