Abstract
Cowpea is an important leguminous crop in arid and semi-arid tropical regions. However, it is sensitive to the temperature and photoperiod. In this study, 41 new lines of cowpea were evaluated in sowing seasons of spring (SpS), summer (SuS), and autumn (AuS) at Wuming, Guangxi province, China. Nine agronomic and yield-related traits were investigated and analyzed. The results showed that the pod width (0.8 cm) was similar among sowing seasons. For all other traits, the values were lowest at SuS, except the number of branches per plant, which was lowest (2.5) at AuS. Strong correlations were detected for the growth period, plant height, pod length, pod width, and 100-seed weight (p < 0.01) across the sowing seasons. A principal component analysis revealed that the first three components could explain 72.47% of the total variations. On the basis of the possible yield, growth period, and growth habit, we identified nine new lines suitable for different sowing seasons in Guangxi. The results of this study suggest these new cowpea lines may be useful for increasing crop production and for breeding new varieties.
1. Introduction
Cowpea [Vigna unguiculata (Linn.) Walp.] (2n = 2x = 22), which belongs to the family Leguminosae [1], is one of the most important pulse crops because its seeds are rich in proteins, minerals, and vitamins, making them a valuable component of food and feed [2,3,4]. Cowpea is native to Africa and was traditionally cultivated in tropical and subtropical regions. Its ability to fix atmospheric nitrogen makes cowpea useful for intercropping with other species and may help to explain its relatively broad distribution [5,6]. According to FAO data (https://www.fao.org/faostat/en/#data/QCL (accessed on 1 September 2022)), the total cowpea planting area has fluctuated slightly and the total cowpea yield has increased somewhat in recent years, reaching approximately 10 million tons (from 15 million hectares) in 2021 [7].
Frequent climate changes result in environmental stresses, especially drought and heat, that seriously affect plant growth [8]. For example, previous studies showed that in the dry season, the yield of six main crops can decrease by 0.6–10% with every 1 °C increase in temperature [9,10]. Cowpea can grow in fertile [11] or saline soil [12], while also tolerating high temperatures and drought conditions [13], making it a popular choice for crop cultivation in saline-alkaline regions as well as for improving soil conditions (e.g., soil adversely affected by natural disasters). Cowpea may help to address the major challenges to food and nutrition security and agricultural sustainability due to global climate changes [14,15]. However, being a typical short-day species, most cowpea cultivars are sensitive to the photoperiod and temperature. Additionally, the introduction of cowpea to new regions or its cultivation in different seasons may lead to a prostrate growth habit and abnormal or incomplete maturation before the next planting season or the onset of winter, resulting in a poor mechanical harvest and low yields.
In China, cowpea is a minor crop that has traditionally been grown in the mountainous regions, usually as a monoculture in terraced fields and gardens or intercropped with other crops and fruit trees, such as peanut [16], cherry [17], and apple trees [18]. In 2021, the total cowpea cultivation area in China was approximately 15,000 hectares and the total yield was about 15,000 tons, which were 10% and 44% increases from the corresponding data in 2016 [7].
The effects of the temperature and photoperiod on cowpea growth remain relatively unknown. Guangxi province, which is located in southern China, has light and temperature conditions that are suitable for cowpea growth from spring to late autumn. Accordingly, there has been a recent rapid increase in the production of this crop in Guangxi. However, the photoperiod and temperature vary greatly among seasons, which influences the adaptability of different varieties [19,20]. Hence, assessing the adaptability of cowpea lines sown on different dates may be useful for optimizing cowpea production and the associated economic benefits for local farmers.
In a previous study, we developed a set of new cowpea lines via hybridizations and evaluated these lines in Guangxi in the summer of 2019 and 2020. Most of the tested lines completed their life cycles and were relatively productive. In the current study, we selected 41 lines and examined their growth in different seasons (spring, summer, and autumn) in Wuming (Guangxi) to analyze their adaptability and utility across seasons. The findings of this study may provide the theoretical basis for improving cowpea germplasm and may also enable local farmers to select ideal varieties for southern China or other global regions with similar environmental conditions.
2. Materials and Methods
2.1. Plant Materials
A total of 41 cowpea lines were used in this study, including 39 new lines and two landraces (Table 1). These 39 new lines were derived from hybridizations between a local Chinese landrace and introduced cowpea lines in the experimental field of the Chinese Academy of Agricultural Sciences (Beijing) in the summer of 2013. They then underwent north-south shuttle breeding and were purified over several generations until they satisfied certain criteria (i.e., distinctness, uniformity, and stability).

Table 1.
Characteristics of 41 cowpea lines.
2.2. Field Trial Design
A field trial involving three sowing seasons was conducted in 2021 in Wuming (23°26′ N, 108°03′ E), Guangxi, China. The study site mainly contains loose lateritic red soil, with good ventilation and fertility. It was previously used to grow pea, potato, and rice in spring, summer, and autumn, respectively. The spring sowing date was 7 April 2021 (SpS), the summer sowing date was 23 June 2021 (SuS), and the autumn sowing date was 6 August 2021 (AuS). The seeds of each line were sown in three rows that were 5 m long, with 0.5 m between rows and 0.15 m between plants. Each plot comprised 200 seedlings. The field trial was repeated twice. Weeds were removed by hand and conventional field practices were applied to prevent and control diseases and insect pests. The daily average temperature and precipitation during the three growing seasons are presented in Figure 1.
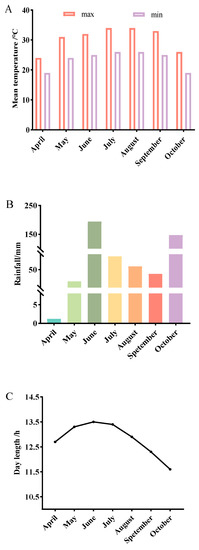
Figure 1.
Wuming climate conditions during the cowpea growing seasons. (A) Mean temperature in Wuming across three sowing seasons. (B) Precipitation in Wuming across three sowing seasons. (C) Day length in Wuming across three sowing seasons. The temperature and rainfall data were obtained from https://www.tianqi24.com/wuming/history202104.html (accessed on 29 September 2022), whereas the day length data were obtained from https://richurimo.bmcx.com/wumingxian__richurimo/?ivk_sa=1023345p (accessed on 29 September 2022).
2.3. Evaluation of Agronomic Traits
Ten individual plants were randomly selected for each line and each replicate to analyze the growth period (GP), plant height (PH), number of branches per plant (NBP), number of pods per plant (NPP), number of seeds per pod (NSP), pod length (PL), pod width (PW), and 100-seed weight (100-SW) as previously described [21]. Briefly, 100-SW was measured using an electronic balance and 100 randomly selected seeds. Additionally, PL was measured using a ruler, whereas PW was measured using a vernier caliper. The number of days from emergence to harvest was recorded as the GP. Plot yield (PY) was measured and the average yield for each line in each sowing season was calculated. The growth habit was determined on the basis of a visual examination of growing plants.
2.4. Statistical Analysis
All phenotypic traits were analyzed using Microsoft Excel 2019. The least significant difference test (p < 0.05), which was performed to assess the significance of the differences between mean values, as well as the analysis of variance (ANOVA), correlation analysis, and principal component analysis (PCA) were conducted using SPSS 26.0. The additive main effects and multiplicative interaction (AMMI) model analysis was performed using QTL IciMapping 4.2 [22].
3. Results
3.1. Climate Conditions in Wuming
Wuming, which is a county in southern China, has a humid subtropical monsoon climate with lots of sunshine and rainfall and an annual average temperature of approximately 21.7 °C. The temperature, rainfall, and photoperiod varied among seasons. From April to October, the day length initially increased, peaking in mid-to-late June, and then decreased (Figure 1C). Changes in the daily maximum and minimum temperatures were not obvious, but the temperatures were generally the highest between June and September (Figure 1A). Rainfall was the greatest in June and October (Figure 1B).
3.2. Analysis of Phenotypic Traits across Sowing Seasons
The analysis of the 41 cowpea lines revealed that the ranges in GP, PH, NBP, NPP, PL, and PY were wider for SpS than for the other two sowing seasons. In contrast, the ranges for NSP and 100-SW were widest for SuS. For most traits, the ranges were smallest for AuS (Table 2).
Table 2.
Mean values, ranges, and coefficients of variation among sowing seasons.
The mean GP and PH values for the 41 cowpea lines were lower for SuS (GP: 51 days and PH: 57.8 cm) than for AuS (GP: 59.7 days and PH: 62.4 cm) and SpS (GP: 69.2 days and PH: 103.7 cm). Similar trends were observed for the mean NPP, NSP, PL, and PY values. The mean NBP value was highest for SuS (3.4), followed by SpS (3.1) and AuS (2.5). The mean 100-SW value was highest for AuS (14.0 g), followed by SpS (13.0 g) and SuS (12.5 g). There were no obvious differences in PW among the sowing seasons (Table 2).
The coefficient of variation (CV) for GP, NSP, PL, PW, and 100-SW was low across the sowing seasons (CV < 20%), which was in contrast to the relatively high CV for NBP and PY (CV > 20%). Moreover, the CV for NBP was slightly higher for SpS (24.7%) than for SuS (21.4%) and AuS (23.5%), whereas the CV for PY was higher for SuS (27.0%) than for SpS (25.7%) and AuS (23.0%). The CV for PH was lowest for SuS (13.7%), while the CV for NPP was lowest for AuS (14.6%) (Table 2).
The ANOVA results (Table 3) indicated there was no significant block effect on the cowpea traits, with the exception of PH (p < 0.001), PW (p < 0.05), and 100-SW (p < 0.05). However, the genotype (G) and sowing date (D) significantly affected all traits (p < 0.001). The G × D interaction also significantly affected all traits (p < 0.001 and p < 0.01).
Table 3.
Factors influencing the variations in cowpea traits.
The correlation analysis showed GP, PH, PL, PW, and 100-SW were highly correlated across SpS, SuS, and AuS (p < 0.01). In addition, NBP and PW were also highly correlated, but only for SpS and AuS (p < 0.01). For NNP, NSP, and PY, there were no obvious correlations across the sowing seasons (Table 4).
Table 4.
Correlation analysis of various traits in different sowing seasons.
3.3. Variations among Lines across Sowing Seasons
The analysis of the data for each line (Table S1) revealed GP was most consistent for line BJ–28, followed by BJ–29 and ZJ–1, whereas GP varied considerably for BJ–20, BJ–21, and BJ–-18. The CV for PH across three sowing seasons was less than 20% for BJ–3, BJ–38, BJ–25, BJ–-1, BJ–22, BJ–4, BJ–35, BJ–19, and BJ–13. In contrast, PH differed substantially across sowing seasons for BJ–33, BJ–15, BJ–10, BJ–27, and BJ–28. Among the examined lines, the NBP, NSP, PL, PW, and 100-SW values were stable among the sowing seasons for BJ–13, BJ–19, BJ–25, BJ–28, BJ–29, and BJ–30. Lines BJ–3 and BJ–4 had stable traits (except for NBP). Similarly, with the exception of PH, BJ–23 also had stable traits. All examined traits were stable for BJ–22. In addition, the lines with a highly variable PH also had unstable NBP and PY values (e.g., BJ–27, BJ–28, BJ–29, BJ–33, and GX18–21). Overall, the growth performance of BJ–3, BJ–4, BJ–22, and BJ–23 was relatively unchanged across the three sowing seasons.
3.4. Principal Component Analysis of Traits
According to the PCA, the first three principal components contributed 72.47% of the total variability in the examined traits (Table 5). The first component (PC1) explained 38.06% of the variation and was positively correlated with GP (0.90), PH (0.80), PW (0.80), and PY (0.59). The second component (PC2) explained 21.93% of the variation and was positively correlated with NPP (0.65), NSP (0.59), PL (0.57), and PY (0.68). The third component (PC3) explained 12.48% of the variation and was correlated with NPP (−0.50), NSP (0.41), and PL (0.65).
Table 5.
Principal component analysis of cowpea traits.
3.5. AMMI Model Analysis
The AMMI biplot of the average yield (Figure 2) suggested the selected lines were high-yielding cowpea varieties. The y-axis of the AMMI biplot presents the effect of the interaction between varieties and sowing seasons. A low absolute value for PCA1 reflects a small interaction effect (i.e., a relatively stable variety across sowing seasons). The average yield was highest for G41 (GX18–21), followed by G19 (BJ–19) and G27 (BJ–27). Moreover, the effect of the interaction between the variety and the sowing season was the greatest for G27 (BJ–27). Among the three sowing seasons, the average yield was highest and lowest for SpS (E1) and SuS (E2), respectively. In the AMMI biplot, most of the lines were concentrated at E3 (i.e., AuS), suggesting they were appropriate for autumn sowing.
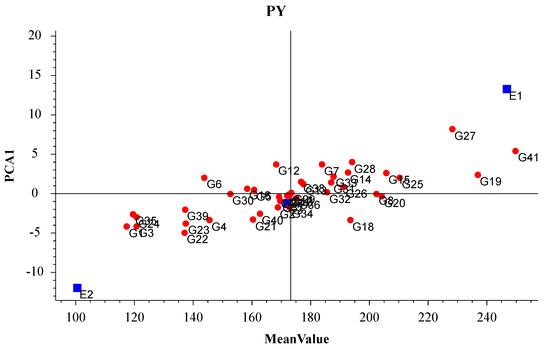
Figure 2.
AMMI biplot (PCA1: average yield). E1, E2, and E3 represent spring, summer, and autumn, respectively. G1–38 represents BJ1–38; G39 represents ZJ–1; and G40 and G41 represent GX18–11 and GX18–21, respectively.
3.6. Selection of Cowpea Lines Suitable for Local Production
According to the yields for the three sowing seasons, 27 lines with high yield potentials were selected for SpS (>250 kg), SuS (>120 kg), and AuS (>200 kg). Of these lines, BJ–8 and BJ–19 had high yield potentials across the sowing seasons. Lines BJ–20, BJ–25, BJ–26, BJ–32, and GX18–21 were suitable for both spring and summer sowing. Lines BJ–13, BJ–14, BJ–15, BJ–28, and BJ–33 were suitable for spring and autumn sowing. Only BJ-18 was suitable for summer and autumn sowing.
Some of the lines with high yield potentials exhibited a prostrate growth habit, which is not conducive for mechanical harvesting. These lines included BJ–11, BJ–13, BJ–15, BJ–18, BJ–19, BJ–-38, and GX18–21. Additionally, the high GP values for BJ–6, BJ–7, BJ–13, BJ–17, and BJ–20 might affect the growth of the intercropped species. Specifically, BJ–20 had a GP that exceeded 76 days in autumn, making it relatively susceptible to powdery mildew infections, which typically occur during the late cowpea growth stage.
Finally, we selected nine elite lines useful for cowpea production in three sowing seasons (Table 6).
Table 6.
Cowpea lines suitable for each sowing season.
4. Discussion
Frequent climate changes might lead to crop yield losses. Cowpea can withstand various environmental stresses, such as drought and high temperatures, and it can grow in low-quality soils. Hence, it is considered to be an important crop for dealing with natural disasters [5], especially in the mountainous regions of China, where the soil quality is low and droughts are common. There have recently been considerable advances in cowpea breeding in China [23]. Additionally, a set of elite lines has been developed and evaluated in different eco-regions [19]. The climate in Guangxi province is appropriate for cowpea cultivation from spring to autumn. Cowpea is often intercropped with young fruit trees or sugarcane in Guangxi. However, the seasonal variations in the photoperiod and rainfall in this part of China affect the growth, development, and productivity of cowpea, which is sensitive to light and temperature. Therefore, the adaptability of new cowpea lines should be investigated to determine which seasons are ideal for their cultivation; similar evaluations have been conducted for many other crops [24,25,26].
In this study, there were substantial morphological variations in the cowpea lines across the three sowing seasons. Moreover, there were significant differences in the main phenological, morphological, and yield-related traits, such as GP, PH, NPP, NSP, and PY, among the sowing seasons, implying these traits were sensitive to changes in the photoperiod and temperature.
Flowering, which is an important event during the life cycle of most crops, is controlled by environmental conditions (e.g., temperature and day length) [27]. The meteorological data for Wuming indicated the temperature is generally lower in spring and autumn than in summer. Low temperatures can delay flowering, thereby prolonging the vegetative phase. Accordingly, delayed sowing can shorten the crop growth period [28,29]. In the present study, cowpea plants flowered in mid-late-May (SpS), late July (SuS), and early September (AuS). Because the temperature was lower in May than in July and September, the flowering time was shorter for SpS than for SuS and AuS. Similarly, the GP values for the 41 examined cowpea lines were higher in spring than in summer and autumn, but the CV for GP was low among the sowing seasons. Previous studies confirmed that the vegetative phase is prolonged when crops are sown early [29,30]. In the current study, the delayed flowering and reproductive growth of cowpea explains why PH was highest for SpS. In addition, the differences in rainfall, which also influences vegetative growth, may help to explain why the GP value was lower for SuS than for AuS and the PH value was higher for SuS than for AuS.
Early sowing enables crops to fully utilize soil moisture and nutrients during relatively long growing seasons, which may lead to increased crop productivity [29,31,32]. In our results, the PY value was higher for SpS than for SuS and AuS, which is consistent with the results of previous studies [33,34]. However, the PY value was lower for SuS than for AuS, likely because of the higher temperatures in summer. Several studies determined that high temperatures could accelerate the phenological period of crops, with detrimental effects on yield [35,36,37]. Additionally, the rainfall in Wuming occurs mostly from June to October. Thus, the high temperatures and humidity levels during this period adversely affect cowpea pollen viability, causing flowers and pods to fall, which ultimately leads to yield losses [38]. Although the cowpea yield was highest for SpS, if cowpea is sown too early, several critical environmental factors (e.g., abrupt temperature decreases) may negatively modulate germination and seedling growth.
The AMMI model may be combined with ANOVA and PCA [39] to elucidate the effect of the interaction between the genotype and environment on specific traits [40]. Because of the intuitiveness of the associated analysis, the AMMI model has been widely used to in rice [41], wheat [42], maize [43], and cotton [44] yields. According to the AMMI biplot presented herein, lines GX18–21, BJ–19, and BJ–27 may produce high yields in Guangxi. Furthermore, sowing in spring may optimize cowpea production in this part of China. However, the high PH values in spring are unfavorable for mechanical harvesting. The AMMI biplot also indicated that most of the analyzed cowpea lines are suitable for growth in autumn. Because of their sensitivity to the photoperiod and temperature, the suitability of several cowpea lines (e.g., BJ–20, BJ–36, BJ–37, and GX18–21) for specific environmental conditions must be assessed before they are introduced to new regions. The wide adaptability of some of the lines (e.g., BJ–22), which was reflected by the relatively stable traits across sowing seasons, should be exploited by cowpea breeding programs.
Finally, we selected nine lines that satisfied the following criteria: high yield, short growth period, and erect growth habit. These criteria were selected because yield is the key index for evaluating the production potential of crops, but an erect growth habit is critical for the mechanical harvesting systems used in modern agriculture. However, we analyzed the lines for only 1 year. Thus, additional tests will need to be conducted before these elite lines are cultivated over a wide region. Nevertheless, we believe these new cowpea lines will be useful for increasing cowpea production in Guangxi and in other regions with similar environmental conditions.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy13020551/s1, Table S1: The analysis of the data for each line.
Author Contributions
Experimental designed, L.W.; accessions, L.W. and S.W.; fieldwork, L.J., G.L., Y.C. and D.G.; data analysis, D.G.; writing—original draft preparation, D.G.; writing—review and editing, L.W.; funding acquisition, L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the China Agriculture Research System of MOF and MARA (CARS-08), the Agricultural Science and Technology Innovation Program (ASTIP) from CAAS, the Central Public-interest Scientific Institution Basal Research Fund (No. Y2022XK08), the Program of Protection of Crop Germplasm Resources in China (grant Nos. 2019NWB036-07 and 19200385-6).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boukar, O.; Abberton, M.; Oyatomi, O.; Togola, A.; Tripathi, L.; Fatokun, C. Introgression breeding in cowpea [Vigna unguiculata (L.) Walp.]. Front. Plant Sci. 2020, 11, 567425. [Google Scholar] [CrossRef] [PubMed]
- Huynh, B.L.; Ehlers, J.D.; Ndeve, A.; Wanamaker, S.; Lucas, M.R.; Close, T.J.; Roberts, P.A. Genetic mapping and legume synteny of aphid resistance in African cowpea (Vigna unguiculata L. Walp.) grown in California. Mol. Breed 2015, 35, 36. [Google Scholar] [CrossRef] [PubMed]
- Sombié, P.A.E.D.; Compaoré, M.; Coulibaly, A.Y.; Ouédraogo, J.T.; Tignégré, J.S.; Kiendrébéogo, M. Antioxidant and phytochemical studies of 31 Cowpeas (Vigna unguiculata (Walp L.)) genotypes from Burkina. Foods 2018, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Ginka, A.A.; Tsvetelina, D.S.; Maria, M.I. Proximate and lipid composition of cowpea (Vigna unguiculata L.) cultivated in Bulgaria. J. Food Compos. Anal. 2014, 33, 146–152. [Google Scholar]
- Carvalho, M.; Lino-Neto, T.; Rosa, E.; Carnide, V. Cowpea: A legume crop for a challenging environment. J. Sci. Food Agric. 2017, 97, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.S.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef]
- FAOSTAT [EB/OL]. Available online: https://www.fao.org/faostat/zh/#data (accessed on 1 September 2022).
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.B.; Huang, J.L.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.H.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Field, C.B. Global scale climate–Crop yield relationships and the impacts of recent warming. Environ. Res. Lett. 2007, 2, 14002. [Google Scholar] [CrossRef]
- Timko, M.P.; Rushton, P.J.; Laudeman, T.W.; Bokowiec, M.T.; Chipumuro, E.; Cheung, F.; Town, C.D.; Chen, X.F. Sequencing and analysis of the gene-rich space of cowpea. BMC Genomics 2008, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Tlahig, S.; Bellani, L.; Karmous, I.; Barbieri, F.; Loumerem, M.; Muccifora, S. Response tosalinity in legume species: An insight on the effects of salt stress during seed germination and seedling growth. Chem. Biodivers. 2021, 18, e2000917. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Castro, I.; Moutinho-Pereira, J.; Correia, C.; Egea-Cortines, M.; Matos, E.; Rosa, E.; Carnide, V.; Lino-Neto, T. Evaluating stress responses in cowpea under drought stress. J. Plant Physiol. 2019, 241, 153001. [Google Scholar] [CrossRef] [PubMed]
- Ogundele, O.M.; Gbashi, S.; Oyeyinka, S.A.; Kayitesi, E.; Adebo, O.A. Optimization of infrared heating conditions for precooked cowpea production using response surface methodology. Molecules 2021, 26, 6137. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, C.; Sun, Z.; Wang, L.L.; Duanmu, D.Q.; Fan, Q.L. Genome editing in cowpea Vigna unguiculata using CRISPR-Cas9. Int. J. Mol. Sci. 2019, 20, 2471. [Google Scholar] [CrossRef]
- Zou, X.J.; Liu, Z.Q.; Niu, S.W. Preliminaryreport on fertilizer saving and benefit increasing experiment of peanut cowpea intercropping system. South China Agric. 2019, 13, 3–4. [Google Scholar]
- Niu, Y.; Bo, X.F.; Qin, Z.X.; Meng, Q.X. Effects of different legume intercropping on sapling growth, soil properties and soil microbial population in cherry orchard. Northern Horticultur. 2019, 4, 38–44. [Google Scholar]
- Wang, H.N.; Shang, C.; Shao, J.Z.; Sun, J.S. Effects of intercropping with cowpea (Vigna unguiculata L. Walp) on physiological parameters and frost hardiness of young apple tree. J. Agric. Univ. Hebei 2013, 36, 26–30. [Google Scholar]
- Gong, D.; Luo, G.L.; Zhang, X.Y.; Zhu, X.; Yin, Z.G.; Wang, S.H.; Sha, A.H.; Wang, L.X. Assessment of adaptability for 34 new cultivars of cowpea under different eco-environments. Crops 2022, 2, 89–95. [Google Scholar]
- Zhang, P.; Wang, S.H.; Pan, X.W.; Gong, D.; Chen, H.L.; Cheng, X.Z.; Wang, L.X. The breeding of new cowpea cultivar Zhongjiang No 10. China Seeds 2022, 5, 89–90. [Google Scholar]
- Wang, P.Z.; Li, X.X. Descriptors and Data Standard for Cowpea [Vigna unguiculata (L.) Walp.]; China Agriculture Press: Beijing, China, 2006. [Google Scholar]
- Wang, J.K.; Li, H.H.; Zhang, L.Y.; Lei, M. Version 4.1 of QTL IciMapping: Integrated software for genetic linkage map construction and QTL mapping in bi-parental populations. In Proceedings of the 7th International Crop Science Congress (7th ICSC), Beijing, China, 14–19 August 2016. [Google Scholar]
- Cheng, X.Z.; Wang, S.M. Edible Beans Cultivars in China; China Agricultural Science and Technology Press: Beijing, China, 2009. [Google Scholar]
- Ke, F.L.; Ma, X.L. Responses of maize hybrids with contrasting maturity to planting date in Northeast China. Sci. Rep. 2021, 11, 15776. [Google Scholar] [CrossRef]
- Lamari, N.; Zhendre, V.; Urrutia, M.; Bernillon, S.; Maucourt, M.; Deborde, C.; Prodhomme, D.; Jacob, D.; Ballias, P.; Rolin, D.; et al. Metabotyping of 30 maize hybrids under early-sowing conditions reveals potential marker-metabolites for breeding. Metabolomics 2018, 14, 132. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Fayyaz-ul-Hassan. Response of spring wheat (Triticum aestivum L.) quality traits and yield to sowing date. PLoS ONE 2015, 10, e126097. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Tuttolomondo, T.; Virga, G.; Rossini, F.; Anastasi, U.; Licata, M.; Gresta, F.; Bella, S.L.; Santonoceto, C. Effects of environment and sowing time on growth and yield of upland cotton (Gossypium hirsutum L.) cultivars in Sicily (Italy). Plants 2020, 9, 1209. [Google Scholar] [CrossRef]
- Richards, M.F.; Preston, A.L.; Napier, T.; Jenkins, L.; Maphosa, L. Sowing date affects the timing and duration of key chickpea (Cicer arietinum L.) crowth phases. Plants 2020, 9, 1257. [Google Scholar] [CrossRef] [PubMed]
- Khan, A. Planting density and sowing date strongly influence growth and lint yield of cotton crops. Field Crops Res. 2017, 209, 129–135. [Google Scholar] [CrossRef]
- Yoldas, F.; Esiyok, D. Effects of sowing dates and cultural treatments on growth, quality and yield of processing beans. Pak. J. Biol. Sci. 2007, 10, 2470–2474. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.X.; Guo, J.; Gu, M.F.; Wu, Y.C.; Mei, Y.; Zheng, J.Q. Effects of sowing date and density on yield and some yield characters of red cowpea. Jiangsu Agric. Sci. 2010, 6, 252–253. [Google Scholar]
- Yin, L.j.; Dai, X.L.; He, M.R. Delayed sowing improves nitrogen utilization efficiency in winter wheat without impacting yield. Field Crops Res. 2018, 221, 166–174. [Google Scholar] [CrossRef]
- Gormus, O.; Yucel, C. Different planting date and potassium fertility effects on cotton yield and fiber properties in the çukurova region, Turkey. Field Crops Res. 2022, 78, 141–149. [Google Scholar] [CrossRef]
- Ferrise, R.; Triossi, A.; Stratonovitch, P.; Bindi, M.; Martre, P. Sowing date and nitrogen fertilization effects on dry matter and nitrogen dynamics for durum wheat: An experimental and simulation study. Field Crops Res. 2010, 117, 245–257. [Google Scholar] [CrossRef]
- Liu, K.Z.; Zhang, C.X.; Guan, B.B.; Yang, R.; Liu, K.; Wang, Z.Z.; Li, X.; Xue, K.Y.; Yin, L.J.; Wang, X.Y. The effect of different sowing dates on dry matter and nitrogen dynamics for winter wheat: An experimental simulation study. PeerJ. 2021, 9, e11700. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.L.; Shen, X.; Cai, X.M.; Yan, F.B.; Lu, W.P.; Cheng-Shi, Y. Effects of heat stress during grain filling on the structure and thermal properties of waxy maize starch. Food Chem. 2014, 143, 313–318. [Google Scholar] [CrossRef]
- Zhang, C.M.; Zhao, K.; Tang, S.; Li, Z.S.; Chen, M.; Zhou, Z.G.; Zhou, S.M. Correlation, Principal component and cluster analysis of agronomic traits of six cowpea varieties. Southwest China J. Agric. Sci. 2021, 34, 501–507. [Google Scholar]
- Gilmour, A. Statistical analysis of regional yield trials AMMI analysis of factorial designs. Agric. Ecosyst. Environ. 1994, 48, 307–308. [Google Scholar] [CrossRef]
- Wang, H.X.; Shan, F.H.; Tian, L.P.; Ma, Q.Y.; Zhao, C.P.; Zhang, F.T. Analysis of stability of winter wheat varieties in the regional trails of the northern wheat region of China. Crops 2018, 5, 40–44. [Google Scholar]
- Hasan, M.J.; Kulsum, M.U.; Sarker, U.; Matin, M.Q.I.; Shahin, N.H.; Kabir, M.S.; Ercisli, S.; Marc, R.A. Assessment of GGE, AMMI, Regression, and its deviation model to identify stable rice hybrids in Bangladesh. Plants 2022, 11, 2336. [Google Scholar] [CrossRef]
- Bishwas, K.C.; Poudel, M.R.; Regmi, D. AMMI and GGE biplot analysis of yield of different elite wheat line under terminal heat stress and irrigated environments. Heliyon 2021, 7, e7206. [Google Scholar]
- Shojaei, S.H.; Mostafavi, K.; Omrani, A.; Omrani, S.; Mousavi, S.M.N.; Illés, A.; Bojtor, C.; Nagy, J. Yield stability analysis of maize (Zea mays L.) hybrids using parametric and AMMI methods. Scientifica 2021, 2021, 5576691. [Google Scholar] [CrossRef]
- Farias, F.J.; Carvalho, L.P.; Silva, F.J.L.; Teodoro, P.E. Biplot analysis of phenotypic stability in upland cotton genotypes in Mato Grosso. Genet Mol Res. 2016, 15, gmr8009. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).