



Pollen Visualization of Turkish Flora of Selected Plant Species under Light, Scanning, and Transmission Microscopy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Plant İdentification
2.2. Light Microscopy (LM)
2.3. Scanning Electron Microscopy (SEM)
2.4. Transmission Electron Microscopy (TEM)
2.5. Cluster Analysis and PCA (Principal Component Analysis)
3. Result
3.1. Pollen Descriptions
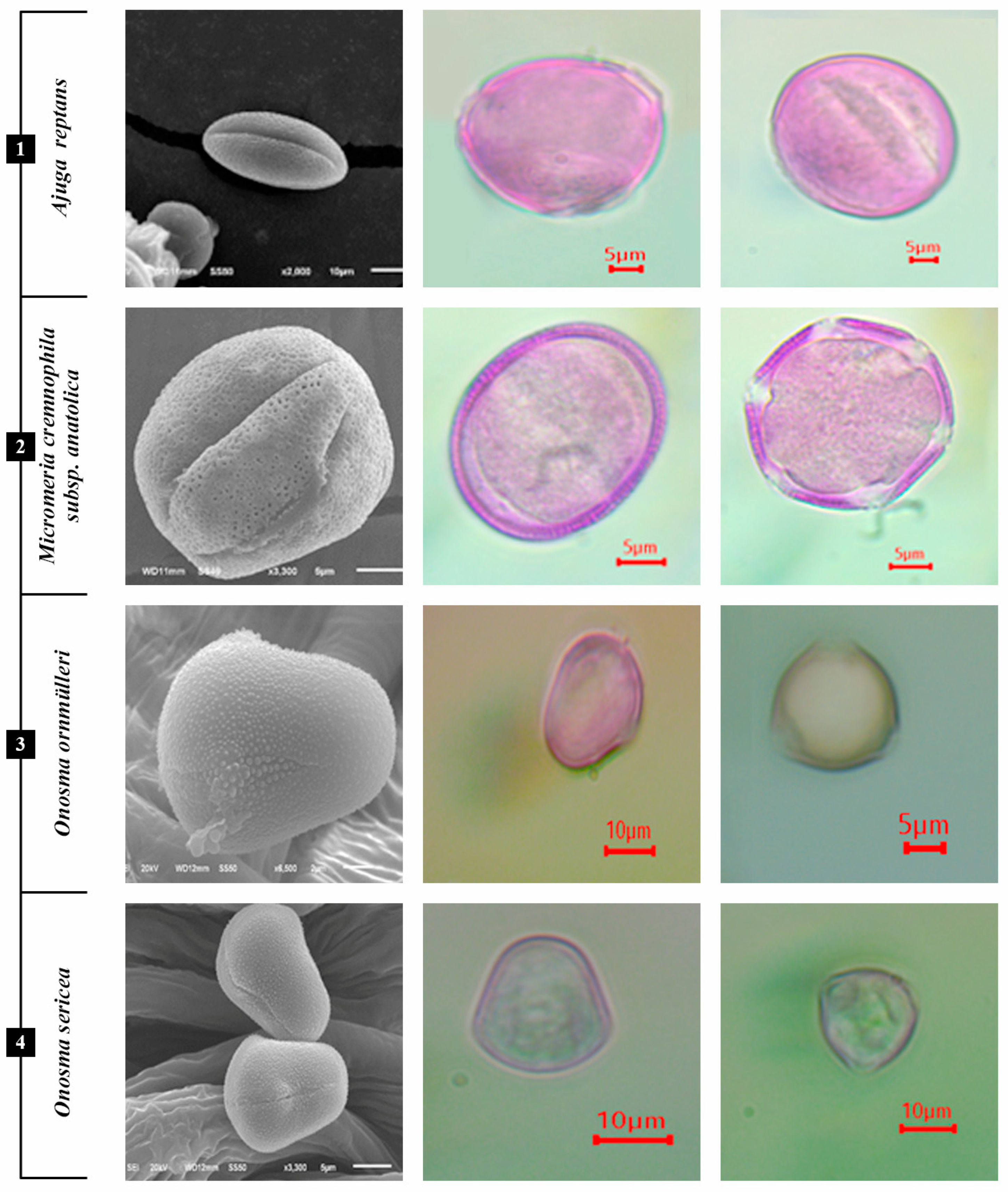
3.1.1. A. reptans
3.1.2. M. cremnophila subsp. anatolica
3.1.3. O. bornmuelleri and O. sericeum
3.1.4. R. caespitosa
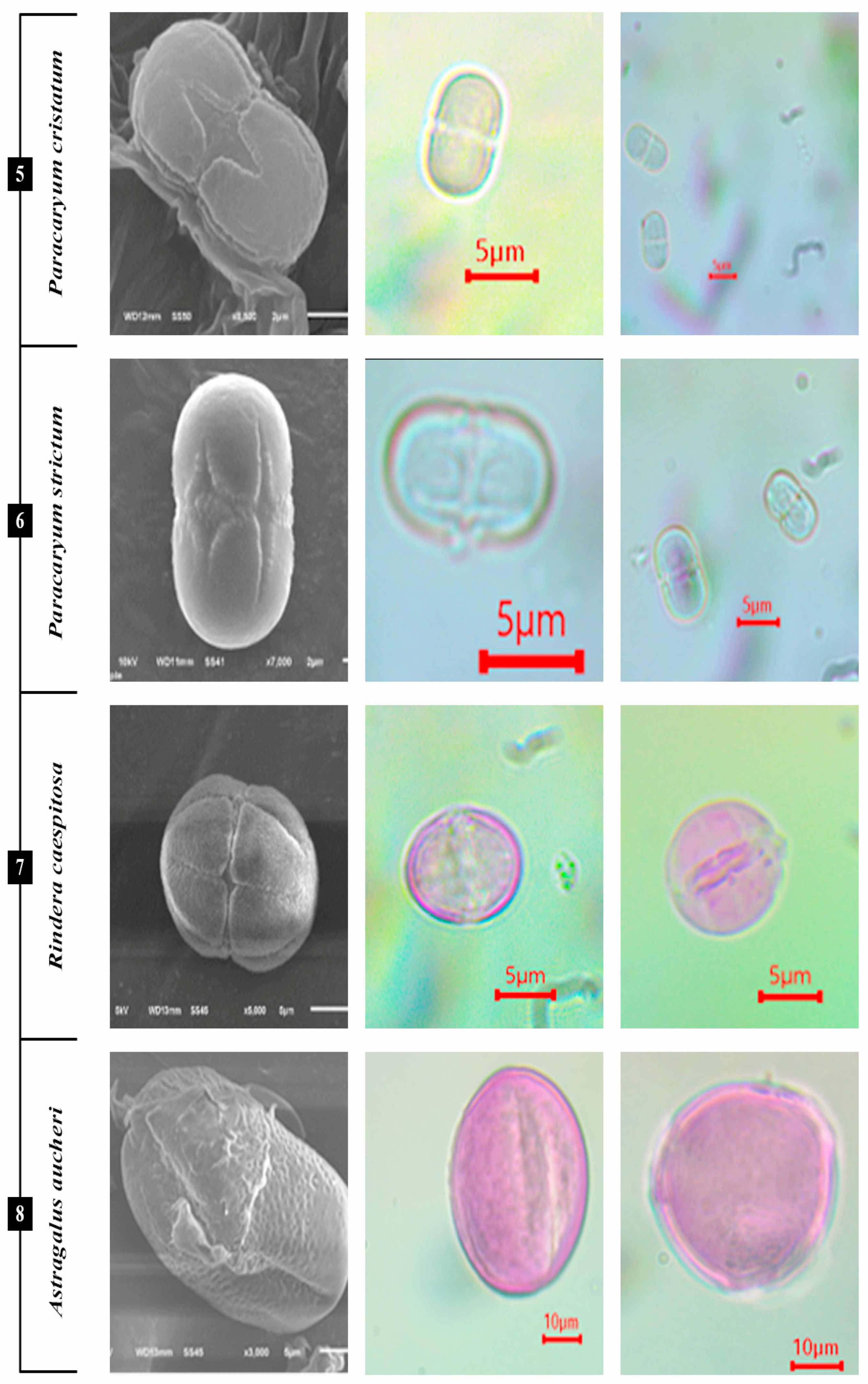
3.1.5. P. cristatum subsp. cristatum, P. strictum
3.1.6. A. aucheri, A. aureus, A. dasycarpum
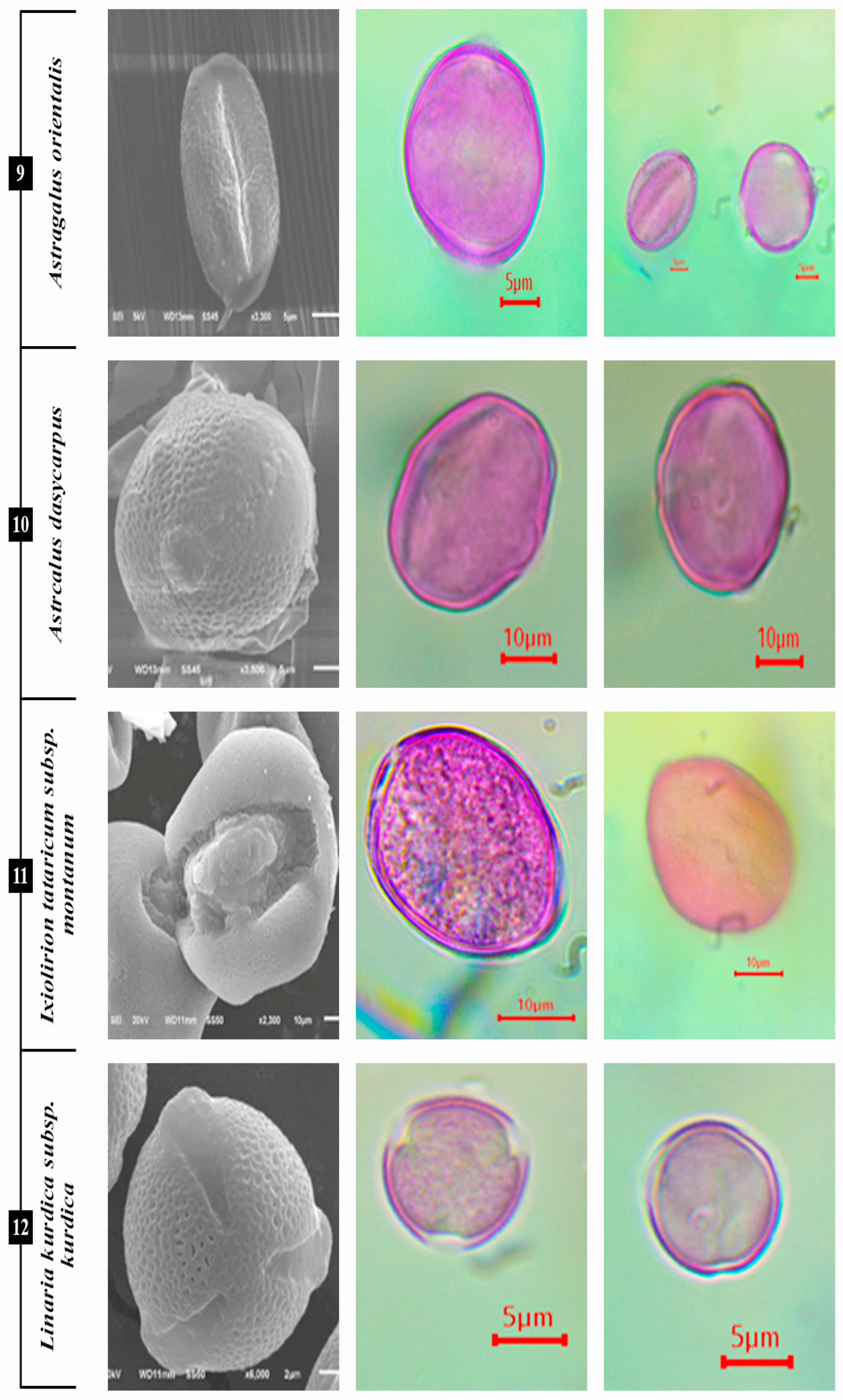
3.1.7. I. tataricum subsp. montanum
3.1.8. L. kurdica subsp. kurdica
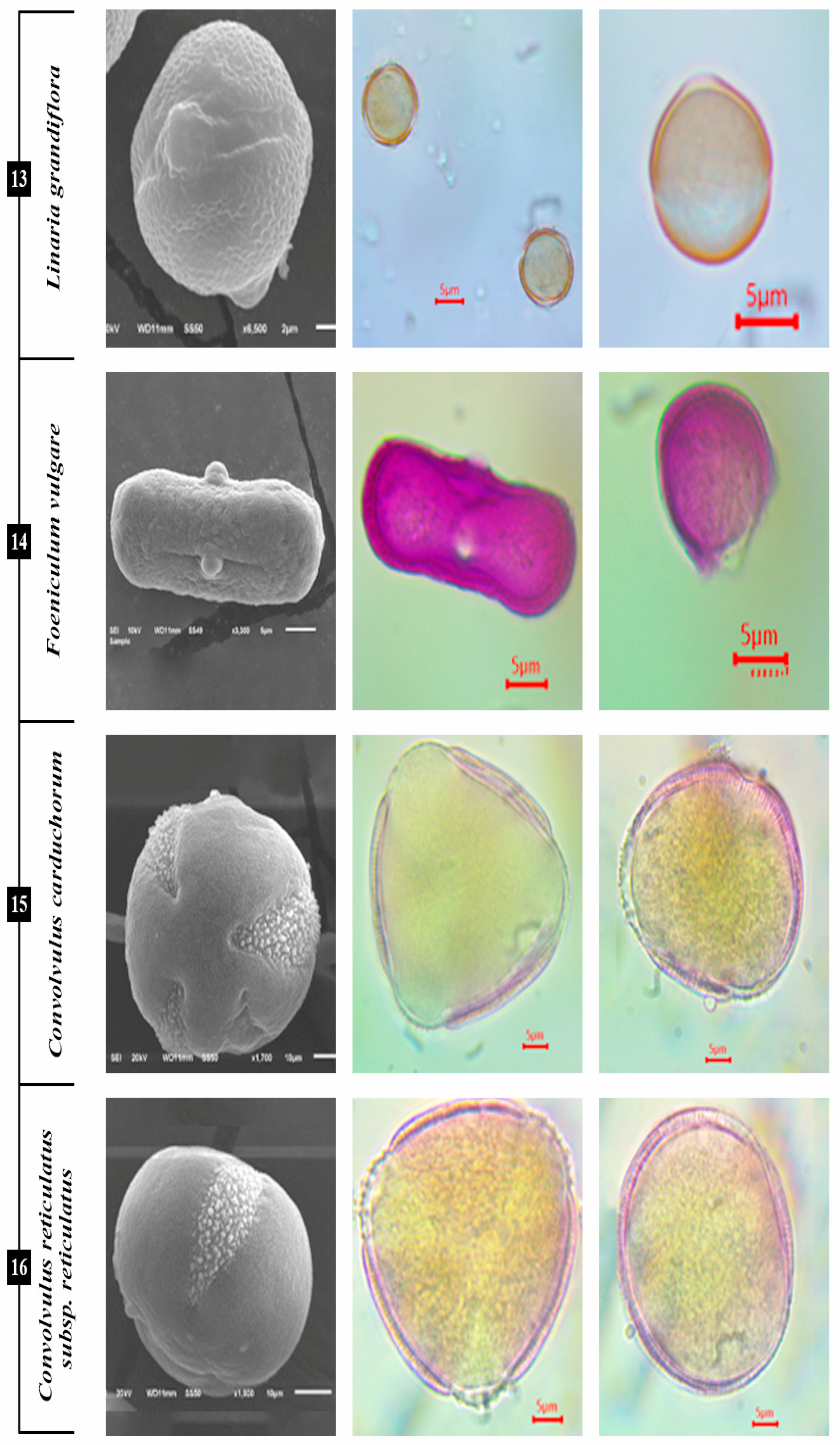
3.1.9. L. grandiflora
3.1.10. F. vulgare
3.1.11. C. carduchorum and C. reticulatus subsp. reticulatus
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Munsif, S.; Khan, M.A.; Ahmad, M.; Zafar, M.; Shah, G.M.; Yasmin, G. Comparative pollen studies of the genera Lantana, Verbena and Vitex of family Verbenaceae from Pakistan. Int. J. Agric. Biol. 2007, 42, 545–549. [Google Scholar]
- Hyde, H.A.; Williams, D.A. The right word. Pollen Anal. Circ. 1944, 8, 6. [Google Scholar]
- Lopes, A.C.V.; Souza, C.N.D.; Saba, M.D.; Gasparino, E.C. Pollen morphology of Malvaceae from Cerrado forest fragments: Details of aperture and ornamentation in the pollen types definition. Palynology 2022, 46, 1–15. [Google Scholar] [CrossRef]
- Soares, E.L.; Landi, L.A.D.C.; Gasparino, E.C. Additions to the knowledge of the pollen morphology of some Fabaceae from Cerrado forest patches of Brazil. Palynology 2021, 45, 269–281. [Google Scholar] [CrossRef]
- Anar, M.; Ahmad, M.; Zafar, S.; Elnaggar, A.Y.; Zafar, M.; Sultana, S.; Tariq, A.; Anjum, F.; Hussein, E.E.; Kiliç, Ö.; et al. Palyno-morphological diversity of Asteraceous and Poaceous allergenic plant using microscopic techniques in lesser Himalaya-Pakistan. Microsc. Res. Tech. 2022, 85, 2061–2075. [Google Scholar] [CrossRef]
- Ahmad, S.; Ahmad, M.; Fawzy Ramadan, M.; Sultana, S.; Papini, A.; Ullah, F.; Saqib, S.; Ayaz, A.; Ahmed Bazai, M.; Zaman, W.; et al. Palynological Study of Fossil Plants from Miocene Murree Formation of Pakistan: Clues to Investigate Palaeoclimate and Palaeoenvironment. Agronomy 2023, 13, 269. [Google Scholar] [CrossRef]
- Usma, A.; Ahmad, M.; Zafar, M.; Sultana, S.; Ullah, F.; Saqib, S.; Ayaz, A.; Zaman, W. Palynological Study of Weed Flora from Potohar Plateau. Agronomy 2022, 12, 2500. [Google Scholar] [CrossRef]
- Karaismailoğlu, M.C. Taxonomical, morphological, palynological, anatomical and ecological investigations on monotypic genus Pachyphragma from Turkey. Pak. J. Bot. 2019, 51, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Doğan, C.; Çakır, N. Pollen Morphology of the Genus Arenaria L. (Subgenus Arenaria) (Caryophyllaceae) in Turkey. Pak. J. Bot. 2019, 51, 6. [Google Scholar]
- Khan, S.A.; Tahir, S.S.; Mangi, J.U. Colpate, colporate and porate pollen isolated from the shale of bara formation, Lakhra, Sindh, Pakistan. Pak. J. Bot. 2022, 52, 2095–2099. [Google Scholar] [CrossRef]
- Raza, J.; Ahmad, M.; Zafar, M.; Athar, M.; Sultana, S.; Majeed, S.; Yaseen, G.; Imran, M.; Nazish, M.; Hussain, A. Comparative foliar anatomical and pollen morphological studies of Acanthaceae using light microscope and scanning electron microscope for effective microteaching in community. Microsc. Res. Tech. 2020, 83, 1103–1117. [Google Scholar] [CrossRef] [PubMed]
- Asgari Nematian, M.; Ranjbar, M. Comparative pollen morphology and its systematic implications in Resedaceae in Iran. Nord. J. Bot. 2022, 2, 03394. [Google Scholar] [CrossRef]
- Carlquist, S. Tribal interrelationships and phylogeny of the Asteraceae. Aliso A J. Syst. Florist. Bot. 1976, 8, 465–492. [Google Scholar] [CrossRef] [Green Version]
- Erdtman, G. Pollen Morphology and Plant Taxonomy: Angiosperms; Brill Archive: Leiden, The Netherlands, 1986; Volume 1. [Google Scholar]
- Gentry, A.H.; Tomb, A.S. Taxonomic implications of Bignoniaceae palynology. Ann. Mo. Bot. Gard. 1979, 66, 756–777. [Google Scholar] [CrossRef]
- Ulukuş, D.; Tugay, O. Türkiye için endemik bir bitki olan Cousinia foliosa Boiss. & balansa (asteraceae) üzerine anatomik ve palinolojik çalışmalar. Selçuk Üniversitesi Fen Fakültesi Fen Derg. 2020, 46, 118–125. [Google Scholar]
- Bahadur, S.; Taj, S.; Long, W.; Ahmad, M. Pollen morphology and its implication in the taxonomy of some selected tribes of the Asteraceae of Hainan Island South China. Bot. Rev. 2022, 88, 271–298. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Long, W.; Hanif, U. Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance. Agronomy 2022, 12, 1122. [Google Scholar] [CrossRef]
- Umber, F.; Zafar, M.; Ullah, R.; Bari, A.; Khan, M.Y.; Ahmad, M.; Sultana, S. Implication of light and scanning electron microscopy for pollen morphology of selected taxa of family Asteraceae and Brassicaceae. Microsc. Res. Tech. 2022, 85, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.; Zafar, M.; Ahmad, M.; Zafar, S.; Ghufran, A.; Ayoub, M.; Sultana, S.; Yaseen, G.; Raza, J. Morpho-palynological and anatomical studies in desert cacti (Opuntia dillenii and Opuntia monacantha) using light and scanning electron microscopy. Microsc. Res. Tech. 2022, 85, 2801–2812. [Google Scholar] [CrossRef]
- Zafar, A.; Zafar, M.; Ahmad, M.; Khan, A.M.; Mahmood, T.; Kilic, O.; Fatima, A.; Habib, D.; Sultana, S.; Majeed, S.; et al. Microscopic (LM and SEM) visualization of pollen ultrastructure among honeybee flora from lower Margalla Hills and allied areas. Microsc. Res. Tech. 2022, 85, 3325–3338. [Google Scholar] [CrossRef]
- Demirpolat, A.; Dogan, G.; Bagci, E. Pollen Morphology and Fruit Micromorphology of the Genus Scandix (Apiaceae). Feb-Fresenius Environ. Bull. 2021, 30, 12413. [Google Scholar]
- Erdtman, G. Pollen morphology and plant taxonomy 4. Labiatae, Verbenaceae and Avicenniaceae. Sven. Bot. Tidskr. 1945, 39, 279–285. [Google Scholar]
- Wodehouse, R.P. Pollen grains. Their Structure, Identification and Significance in Science and Medicine; McGraw-Hill Book Company: New York, NY, USA, 1935. [Google Scholar]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis; Hafner Press: New York, NY, USA, 1975. [Google Scholar]
- Ertman, G. Pollen Morphology and Plant Taxonomy Angiosperms; Ronald Press: New York, NY, USA, 1952. [Google Scholar]
- Kılıç, N.; Yılmaz Dağdeviren, R.; Caner, H.; Akkemik, Ü. Türkiye’de Kullanılmakta Olan Palinoloji ve Polen Terimleri Üzerine Bir Değerlendirme ve Öneriler. Avrasya Terim Derg. 2020, 8, 98–108. [Google Scholar]
- Kose, Y.B.; Erkara, I.; Alan, S. Pollen Morphology of Some Turkish Ajuga L. (Lamiaceae) and its Taxonomic Value. Bangladesh J. Bot. 2011, 40, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Turkmen, Z.; Coşkunçelebi, K.; Makbul, S.; Beyazoğlu, O.; Doğan, C. Pollen Morphology of Onosma L. (Boraginaceae) Taxa Distributed In Anatolia Kuzey-Doğu. Fen Bilim. Enstitüsü Derg. 2011, 4, 71–82. [Google Scholar]
- Bigazzi, M.; Nardi, E.; Selvi, F. Palynological Contribution To The Systematics Of Rindera And The Allied Genera Paracaryum and Solenanthus (Boraginaceae-Cynoglosseae). Willdenowia 2006, 36, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Ekici, M.; Ekim, T. Revision of the Section Hololeuce Bunge of the Genus Astragalus L. (Leguminosae) in Turkey, Turk. J. Bot. 2004, 28, 307–347. [Google Scholar]
- Pınar, M.N.; Ekici, M.; Aytaç, Z.; Akan, H.; Çeter, T. Pollen morphology of Astragalus L. sect. Onobrychoidei DC. (Fabaceae) in Turkey. Turk. J. Bot. 2009, 33, 291–303. [Google Scholar] [CrossRef]
- Akan, H.; Tatlıdil, S.; Bıçakcı, A. Pollen morphology of Astragalus L. Section Alopecuroidei DC. (Fabaceae) in Turkey. Int. J. Bot. 2005, 1, 50–58. [Google Scholar] [CrossRef]
- Oybak Dönmez, E.; Işık, S. Pollen morphology of Turkish Amaryllidaceae, Ixioliriaceae and Iridaceae. Grana 2008, 47, 15–38. [Google Scholar] [CrossRef]
- Çelenk, S. Afyon Ve Çevresinde Yayılış Gösteren Linaria Miller Türlerinin Polen Morfolojileri. Fırat Üniversitesi Mühendislik Bilim. Derg. 2004, 16, 183–194. [Google Scholar]
- Demirpolat, A.; Kılıç, Ö. A Scanning Electron Microscope Study: Investigation of Some Convolvulus L. Taxa (Convolvulaceae) as Morphological and Palynological. J. Glob. Health Nat. Sci. 2021, 4, 94–101. [Google Scholar]
- Güler, A. Bal Arısı (Apis Mellifera); Ondokuz Mayıs Üniversitesi, Ziraat Fakültesi: Samsun, Turkey, 2006; p. 574. [Google Scholar]
- Baydar, H.; Gürel, F. Antalya doğal florasında bal arısı (Apis mellifera)’ nın polen toplama aktivitesi, polen tercihi ve farklı polen tiplerinin morfolojik ve kalite özellikleri. Turk. J. Agric. For. 1998, 22, 475–482. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Plant Name | Locality | Endemic Situation | Voucher Specimen | |
|---|---|---|---|---|---|
| 1. | Ajuga reptans L. | Bingöl-Tunceli | - | AD-257 | |
| 2. | Micromeria cremnophila Boiss. Et Heldr. subsp. anatolica P. H. Davis | Elazig-Harput castle | Endemic | AD-1011 | |
| 3. | Onosma bornmuelleri Hausskn. | Bingol | Endemic | AD-200 | |
| 4. | Onosma sericeum Wild. | Bingol | - | AD-176 | |
| 5. | Rindera caespitosa (A. DC.) Bunge | Bingol | Endemic | AD-78 | |
| 6. | Paracaryum Schreber Boiss. subsp. cristatum Schreber Boiss. | Elazig | Endemic | AD-1012 | |
| 7. | Paracaryum strictum (Koch) Boiss. | Elazig | Endemic | AD-1051 | |
| 8. | Astragalus aucheri Boiss. | Elazig | Endemic | AD-1034 | |
| 9. | Astragalus aureus Wild. | Bingol | - | AD-155 | |
| 10. | Astragalus dactylocarpus Boiss. | Bingol | - | AD-230 | |
| 11. | Ixiolirion tataricum Pallas Herbert subsp. montanum Labill. Takht. | Bingöl | - | AD-158 | |
| 12. | Linaria kurdica Boiss. Et HOHEN. subsp. kurdica Boiss. Et Huet | Elazig | - | AD-1098 | |
| 13. | Linaria grandiflora Desf. | Bingol | - | AD-253 | |
| 14. | Foeniculum vulgare Miller | Bingol | - | AD-543 | |
| 15. | Convolvulus carduchorum Davis | Elazig | Endemic | AD-123 | |
| 16. | Convolvulus reticulatus Choisy subsp. reticulatus Choisy | Bingol | - | AD-112 | |
| Plant Name | Shape | Type and Number of Aperture | Exine Ornamentation | Colpi Orientation | ||
|---|---|---|---|---|---|---|
| 1. | Lamiaceae | A. reptans | Oblate-spheroidal | 3-colporate | Tectatae-granulatae | Sunken, angular |
| 2. | M. cremnophila ssp. anatolica | Sub-oblate | 6-colpate | Reticulate | Sunken | |
| 3. | Boraginaceae | O. bornmuelleri | Sub-prolate | 3-colporate | Scabrate-reticulate | Sunken |
| 4. | O. sericea | Sub-prolate | 3-colporate | Scabrate-reticulate | Sunken | |
| 5. | R. caespitosa | Sub-prolate | 3-colporate | Granulate | Pseudocolpus, margo | |
| 6. | P. cristatum ssp. cristatum | Prolate | 6-colporate | Rugulate | Pseudocolpus, margo | |
| 7. | P. strictum | Prolate | 6-colporate | Rugulate | Pseudocolpus, margo | |
| 8. | Fabaceae | A. aucheri | Prolate | 3-colporate | Reticulate | Sunken, angular, margins distinct |
| 9. | A. aureus | Prolate | 3-colporate | Reticulate | Sunken, angular, margins distinct | |
| 10. | A. dactylocarpus | Prolate | 3-colporate | Reticulate | Sunken, angular, margins distinct | |
| 11. | Amaryllidaceae | I. tataricum subsp. montanum | Sub-prolate | 3-colporate | Perforate | Sunken, angular, margins distinct |
| 12. | Scrophulariaceae | L. kurdica subsp. kurdica | Oblate-spheroidal | 3-colporate | Reticulate | Sunken |
| 13. | L. grandiflora | Oblate-spheroidal | 3-colporate | Reticulate | Sunken | |
| 14. | Apiaceae | F. vulgare | Per-prolate | 3-colporate | Scabrate | Sunken |
| 15. | Convolvulaceae | C. carduchorum | Oblate-spheroidal | 3-colporate | Perforate | Sunken |
| 16. | C. reticulatus subsp. reticulatus | Oblate-spheroidal | 3-colporate | Perforate | Sunken |
| Plant Name | Exine Thickness | Polar Diameter | Equatorial Diameter | P/E Ratio | Colpi Length | Colpi Width | Pores Length | Pores Width | |
|---|---|---|---|---|---|---|---|---|---|
| Lamiaceae | A. reptans | 1.1 ± (0.1) | 22.4 ± (2.0) | 22.7 ± (2.1) | 0.98 | 16.7 ± (1.2) | 7.6 ± (1) | - | - |
| M. cremnophila ssp. anatolica | 1.5 ± (1.2) | 40 ± (2.3) | 46.6 ± (1.3) | 0.85 | 23.5 ± (1.3) | 8.3 ± (1.6) | - | - | |
| Boraginaceae | O. bornmuelleri | 1.0 ± (0.5) | 17.3 ± (1.0) | 14.7 ± (1) | 1.17 | 13.9 ± (0.3) | - | 3 ± (0.5) | 3.29 ± (0.5) |
| O. sericea | 1.0 ± (0.5) | 17.8 ± (1.0) | 14.9 ± (0.1) | 1.19 | 13.8 ± (0.1) | - | 2.83 ± (0.3) | 2.29 ± (0.5) | |
| R. caespitosa | 0.5 ± (0.3) | 13 ± (1.0) | 10 ± (0.1) | 1.30 | 7.1 ± (1.0) | 2.5 ± (1.2) | 3.8 ± (0.5) | 5.3 ± (0.3) | |
| P. cristatum ssp. cristatum | 0.24 ±(0.2) | 9.1 ± (0.3) | 5.6 ± (0.1) | 1.62 | 5.3 ± (1.3) | 1.5 ± (1.5) | 2.9 ± (0.3) | 4.3 ± (0.7) | |
| P. strictum | 0.23 ± (0.1) | 10.1 ± (0.5) | 5.2 ± (0.2) | 1.94 | 6.5 ± (1.3) | 1.9 ± (1.1) | 2.8 ± (0.5) | 4.7 ± (0.3) | |
| Fabaceae | A. aucheri | 0.45 ± (0.1) | 35.5 ± (1.9) | 23.6 ± (1.28) | 1.50 | 29.2 ± (1.3) | 4.2 ± (0.3) | 6.3 ± (0.2) | 4.2 ± (0.5) |
| A.aureus | 0.5 ± (0.1) | 35.8 ± (1.2) | 23.5 ± (1.0) | 1.52 | 27.5 ± (0.3) | 3.9 ± (0.9) | 5.9 ± (0.6) | 4.2 ± (0.7) | |
| A. dactylocarpus | 0.5 ± (0.1) | 38.7 ± (2.2) | 25.3 ± (1.5) | 1.52 | 29.8 ± (1.5) | 5.5 ± (1.2) | 7.4 ± (1.2) | 5.7 ± (1.1) | |
| Amaryllidaceae | I. tataricum subsp. montanum | 1.8 ± (0.7) | 51.2 ± (4.3) | 41.6 ± (3.3) | 1.23 | 38.9 ± (1.9) | 15.0 ± (1.8) | - | - |
| Scrophulariaceae | L. kurdica subsp. kurdica | 0.9 ± (0.2) | 10.9 ± (1.2) | 11.2 ± (1.1) | 0.97 | 8.01 ± (2.3) | 2.4 ± (2.3) | 4.4 ± (1.4) | 4.5 ± (1.8) |
| L. grandiflora | 0.82 ± (0.2) | 10.0 ± (1.2) | 10.3 ± (1.3) | 0.97 | 7.85 ± (1.2) | 3.8 ± (0.9) | 4.23 ± (1.2) | 3.98 ± (1.5) | |
| Apiaceae | F. vulgare | 1.8 ± (0.5) | 26.8 ± (1.9) | 10.0 ± (1.8) | 2.68 | 14.9 ± (2.9) | 4.0 ± (2.3) | 3.8 ± (0.9) | 4.1 ± (0.3) |
| Convolvulaceae | C.carduchorum | 2.1 ± (0.4) | 42.3 ± (1.3) | 43 ± (1) | 0.97 | 18.2 ± (1.9) | 13.7 ± (1.1) | - | - |
| C. reticulatus subsp. reticulatus | 1.9 ± (0.3) | 41.9 ± (0.9) | 43.5 ± (1.2) | 0.97 | 17.3 ± (1.2) | 12.4 ± (1.3) | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demirpolat, A.; Sancar, P.Y. Pollen Visualization of Turkish Flora of Selected Plant Species under Light, Scanning, and Transmission Microscopy. Agronomy 2023, 13, 962. https://doi.org/10.3390/agronomy13040962
Demirpolat A, Sancar PY. Pollen Visualization of Turkish Flora of Selected Plant Species under Light, Scanning, and Transmission Microscopy. Agronomy. 2023; 13(4):962. https://doi.org/10.3390/agronomy13040962
Chicago/Turabian StyleDemirpolat, Azize, and Pelin Yılmaz Sancar. 2023. "Pollen Visualization of Turkish Flora of Selected Plant Species under Light, Scanning, and Transmission Microscopy" Agronomy 13, no. 4: 962. https://doi.org/10.3390/agronomy13040962