2. Materials and Methods
In the experiment, the salt-contaminated soil was collected at the depth of 0–20 cm in An Thuan commune, Thanh Phu district, Ben Tre province. The collected soil was dried up, separated from plant residues, crushed, mixed well and transferred to a black plastic pot whose large bottom × small bottom × height was 23 cm × 17 cm × 18 cm in an amount of 8 kg before being put into application. Then, the soil was supplied with 2 L of tap water to create muds before sowing. The chemical and physical properties of the saline soil are briefly described in
Table 1.
Bacteria source: The ALA-producing
Luteovulum sphaeroides W22 and
L. sphaeroides W47 were isolated and screened from the rice-shrimp saline soil in Thanh Phu, Ben Tre, with accession numbers of MW819907 and OQ819480 [
9].
Fertilizer formula: Types of fertilizers used in the current study included urea (46% N), super phosphate (16% P2O5) and potassium (60% K2O).
Experiment design: Experiment 1 was conducted according to a completely randomized block design with 2 factors. Therein, the first factor (A) was frequencies of applying saline irrigation (none, one, two, three and four times); the other one (B) was the supplementation of ALA-producing purple non-sulfur bacteria (the single strain of L. sphaeroides W22, the single strain of L. sphaeroides W47 and a mixture of L. sphaeroides W22 and W47). In this research, we focused on the differences between single and mixed strains, so the negative control was not designed. However, we mentioned it in the following experiment. Experiment 2 was carried out in the saline soil following the same design as the other experiment with 3 treatments, including (i) no saline irrigation applied, (ii) applying saline irrigation but no bacteria applied and (iii) applying both saline irrigation and the mixture of L. sphaeroides W22 and W47. Experimental pots were placed in the greenhouse in the Agricultural Research and Practice Station, College of Agriculture, Can Tho University. In the greenhouse, the light and dark hours per day were 11 and 13, respectively; the temperature was 36 °C (10.029783 N, 105.767414 E).
Bacterial inoculation: The inoculation of bacteria was referred to from the studies by Khuong et al. [
4,
9,
11]. A single strain of the bacteria was supplied in an amount of 4 mL and a density of 1 × 10
8 CFU mL
−1, while, in the bacterial mixture, the volume of application was 2 mL for each strain. The bacterial supplementation was conducted 4 times at 25, 35, 45 and 60 days after sowing (DAS); i.e., the final bacterial density was approximately 20 × 10
4 CFU g
−1.
Seeds preparation: The rice used in the current study was the OM5451 rice cultivar purchased from the Cuu Long Delta Rice Research Institute, Vietnam. Its seeds were sterilized by ethanol and sodium hypochlorite 1%. Then, they were rinsed by sterile deionized water to ensure that the seeds were free from bacteria. After that, they were incubated for 24 h to germinate. Finally, 6 rice seeds were sowed directly into each pot.
Fertilization: It followed a recommended formula of 90N–60P2O5–30K2O (kg ha−1). For the P fertilizer, it was applied 100% to the ground. For the N fertilizer, it was divided into 3 portions of 30–40–30% and applied each portion at 10, 20 and 45 DAS, respectively. For the K fertilizer, it was separated in 2 halves and fertilized at 10 and 45 DAS with each half.
Irrigation: Tap water was applied continuously at 3–5 cm height during the experiment, except when the saline irrigation was in use.
Saline irrigation: A solution of NaCl at a dose of 4‰was used in experiment 1 and at a dose of 5‰ in experiment 2, and 10 mL of saline irrigation was poured into each pot at 10, 20, 30 and 40 DAS.
Soil analysis: Soil samples at the beginning and at the end of the crop were drilled, collected and brought back to the laboratory. They were left to dry, removed from plant residues and crushed through a 0.5 mm and 2.0 mm sieve. The methods for the soil analysis were summarized as follows: either pH
H2O or pH
KCl was extracted with a ratio of 1:2.5 (soil–water) or 1:2.5 (soil-KCl 1 M), respectively, and measured by a pH meter. The solution from the pH
H2O extraction was reused to measure the electrical conductivity (EC) by an EC meter. To determine the total N content, the samples were digested by a mixture of H
2SO
4saturated: CuSO
4: Se with a ratio 100:10:1, and the Kjeldahl method (Velp UDK129, VELP Scientifica Srl, Italy) was applied to distill samples before they were titrated by H
2SO
4 0.01 N. The blue phenol method was used to determine the amount of the available N at the 640 nm wavelength. The total P content was turned into inorganic forms by a mixture of H
2SO
4saturated—HClO
4; the inorganic solution was revealed in colors by ascorbic acid at the 880 nm wavelength. The available P content was determined by the Bray II method. It was extracted from the soil by HCl 0.1 N and NH
4F 0.03 N with soil–water ratio of 1:7, and the colorimetric method was conducted by ascorbic acid at the 880 nm wavelength. The unavailable P compounds, including ferrous phosphate (Fe-P), aluminum phosphate (Al-P) and calcium phosphate (Ca-P), were extracted by solutions, including NaOH 0.1 M, NH
4F 0.5 M and H
2SO
4 0.25 M, respectively, then detected by ascorbic acid and a spectrometer (UV–Vis 1800 Shimadzu) at the 880 nm wavelength. The cation exchange capacity (CEC) was determined by being extracted from the soil by BaCl
2 0.1 M and titrated by EDTA 0.01 N. Concentrations of K
+, Na
+, Ca
2+ and Mg
2+ from the CEC extracted solution were measured by an atomic absorption spectrometer ICP–OES (Icap 6300 Duo Thermo, Thermo Fisher Scientific Inc., Waltham, MA, USA) at wavelengths of 766.5, 589.0, 422.7 and 285.2 nm, respectively [
18].
Biochemical properties of rice: The proline content in plants was analyzed at 45 DAS. Leaf and stem samples were collected to determine their moisture by drying stem and leaves of immature rice at 70 °C for 72 h. Proline was examined by the Ninhydrin method of Bates et al. [
19], which proceeded as follows: 0.5 g of the fresh rice samples were weighed and put into a 13 mm × 100 mm tube, and then 10 mL of sulfosalicylic acid 3% was added to completely digest the samples, which were then put on a reciprocal shaker in 30 min and centrifuged at 3000 rpm for 15 min. After the debris was removed, the clear extract was collected. Then, 2.0 mL of the sample extract was reacted with 2.0 mL of Ninhydrin and 2 mL of glacial acetic acid in a tube, well mixed and covered. The samples were put into an incubator for 1 h at 100 °C. They were cooled in ice water, reacted with 4 mL of toluene, shaken for 15–20 s and measured on the atomic absorption spectrophotometer at the 520 nm wavelength.
N, P, K and sodium (Na) content in plants: Concentrations of N, P, K and Na in plants were measured according to the method of Walinga et al. [
20]. Collected stems, leaves and seeds were dried up. Subsequently, they were milled well and used to analyze the concentrations of N, P, K and Na in stems, leaves and seeds. The samples were digested by a mixture of saturated H
2SO
4 and salicylic acid. The amount of N in the samples was determined by the Kjeldahl distilling method. The P proportion was measured at the 880 nm wavelength by the spectrometer. Concentrations of K and Na were detected by the atomic absorption spectrometer. Biomass in stem, leaves and seeds was checked by drying the stovers at 70 °C for 72 h and weighing them afterward.
Growth, yield components and grain yield: All growth and agronomic parameters were measured following the description of IRRI [
21]. The growth parameters included the plant height and panicle length, which were checked on 8 plants or panicles per pot when it was at 90 DAS. The plant height was measured from the ground to the peak of a plant. The panicle length was measured from the neck to the end of a panicle. Yield components were measured as follows: the panicle number per pot: the number of panicles of a pot was counted; the seeds number per panicle: the total of seeds was counted on 8 panicles of a pot; the filled seeds percentage: it was equal to number of filled seeds divided by the total number of seeds; the 1000-seed weight: the weight of 1000 seeds in each treatment was determined. The actual grain yield: the seed weight and moisture of each pot was measured at harvesting and converted into the weight at 14% humidity.
Statistical Analysis: The numeric data were subjected to a two-way analysis of variance (ANOVA) by the SPSS software, version 13.0. The Duncan’s post hoc test was utilized for comparing the differences between means of treatments at 5% significance. A relationship between the bacterial density of the PNSB, Na exchange content, Na content in seeds, yield and proline content produced in rice was evaluated by the correlation analysis. Correlations between the bacterial density and the Na exchange content, between the bacterial density and the Na content in seeds, between the Na exchange content and the proline content, between the Na content in seeds and the proline content, between the bacterial density and proline content and between the bacterial density and the yield were expressed by the Pearson correlation coefficient (R
2). The r value ranging between −1 and +1 indicated linearity between 2 variances, and, when it was equal to 0, there was no correlation between variances. Values, including +1, +0.8, +0.5 and +0.1, showed proportionally high, moderate and low correlations, whereas those including −0.8, −0.5 and −0.1 showed inversely high, moderate and low correlations, respectively [
22].
4. Discussion
Supplying the mixture of
L. sphaeroides W22 and W47 was highly promising in enhancing a reduction in stress caused by salinity because the accumulation of the proline content within a plant appeared to be lower (
Table 2). This was also stated by a lower proline content in the treatments supplied with the mixture of the W22 and W47 (3.02 µmol g
−1 DW) compared with that in the control treatments without bacteria (4.39 µmol g
−1 DW) (
Table 6). When the frequency of applying the saline irrigation increased to three and four times, the proline content in the treatments supplied with the mixture of W22 and W47 was lower than that in the treatments supplied with each individual strain (
Figure S1, Supplementary Data). Lutts et al. [
23] claimed that, in rice, the accumulation of proline is a sign of being damaged rather than having the ability to tolerate salinity; that is, saline-tolerant plants accumulate less proline than saline-susceptible ones. According to Siddique et al. [
24], proline is a beneficial amino acid, gathered in cultivars living in different stressful environments, including drought, heavy metals contamination and salinization.
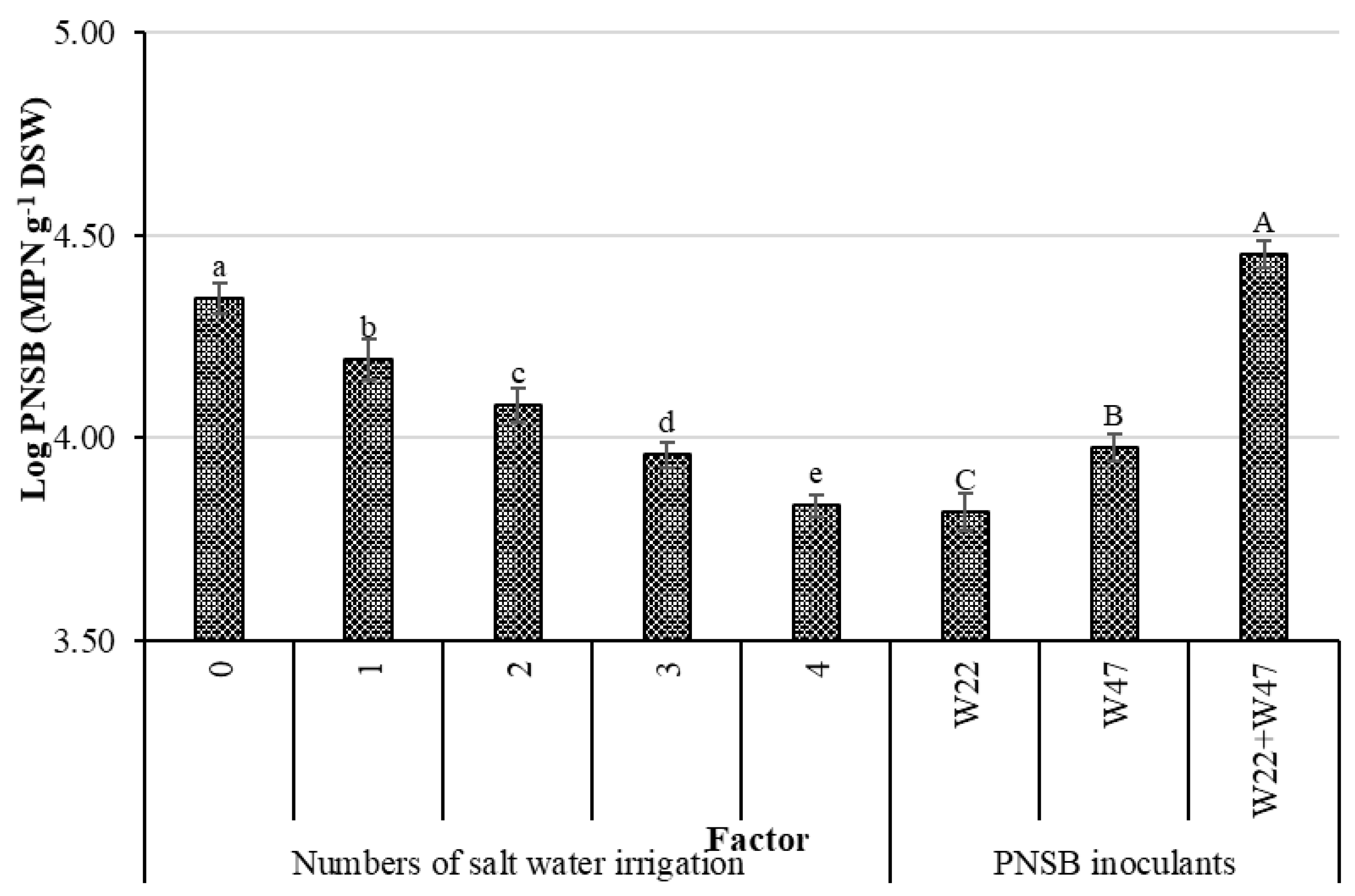
Table 2 indicates that, the more often applying the saline irrigation was, the higher the accumulated proline became. This can be interpreted that the proline production was meant to tolerate saline stress, which agreed with the study by Bhusan et al. [
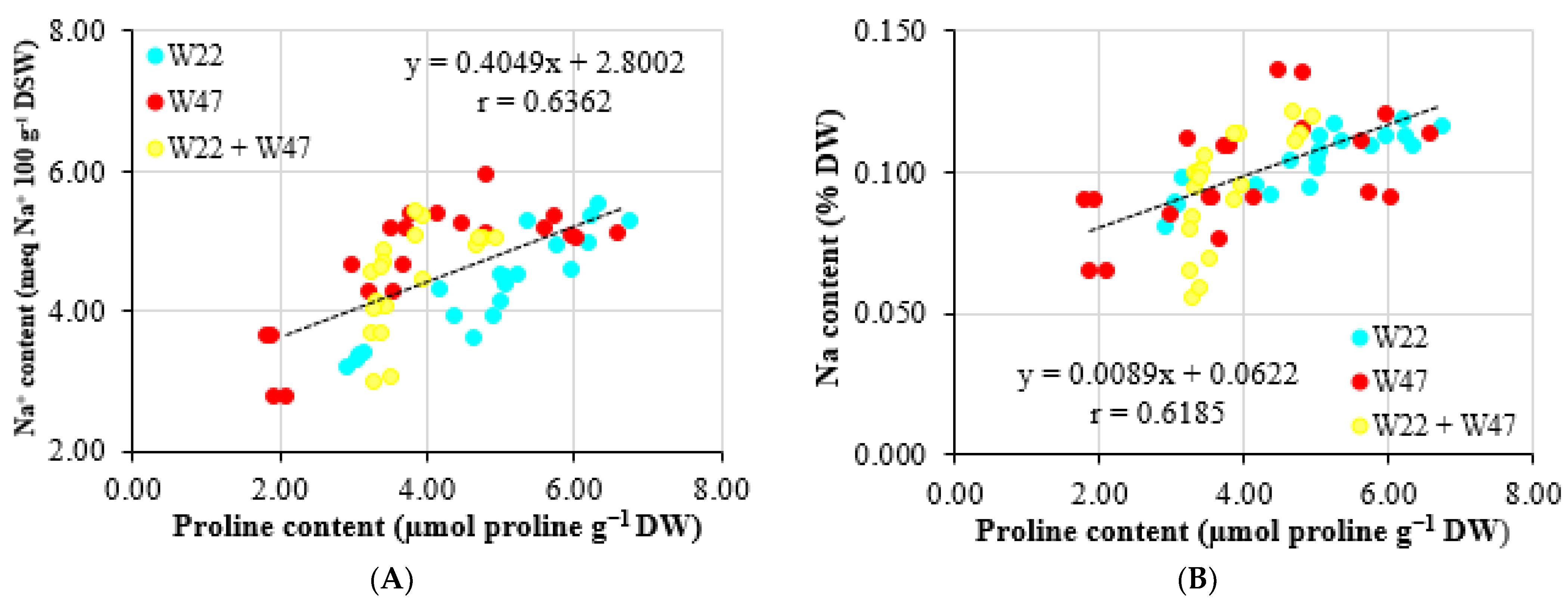
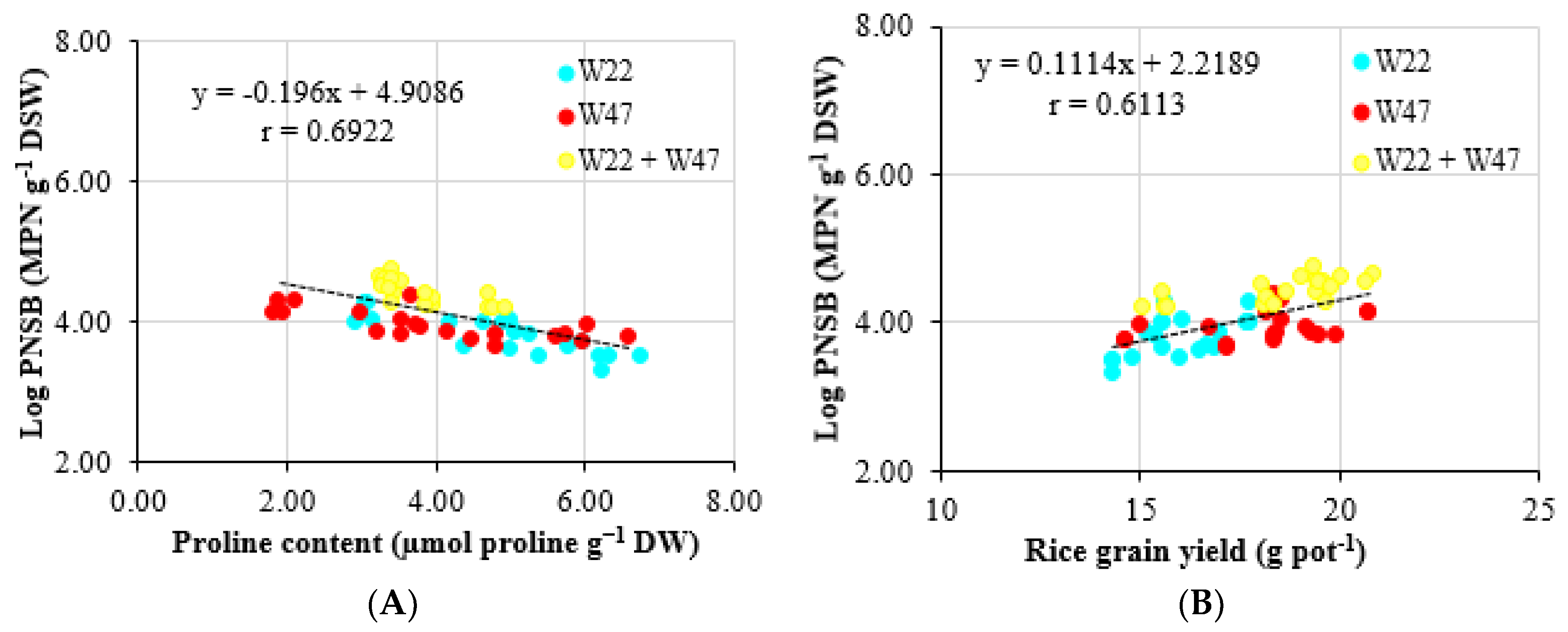
25], where the proline concentration increased in NaCl 25 mM treated conditions in comparison with the no-saline-treated ones. From the above, the bacteria in the current study were able to lessen the stress on rice plants suffered from the saline condition. In other words, the bacteria were suitable to aid plants that were grown in highly saline conditions. Therefore, a close correlation between the soil bacterial density and the proline content was observed, with a correlation coefficient of r = 0.6922 (
Figure 5A). Moreover, ALA seemed to be the key to help plants fight against saline stress. ALA has been observed for a capacity in lower salt concentration in soil and induces salt resistance in crops [
10,
14]. The significance was that supplying the mixture of ALA-producing
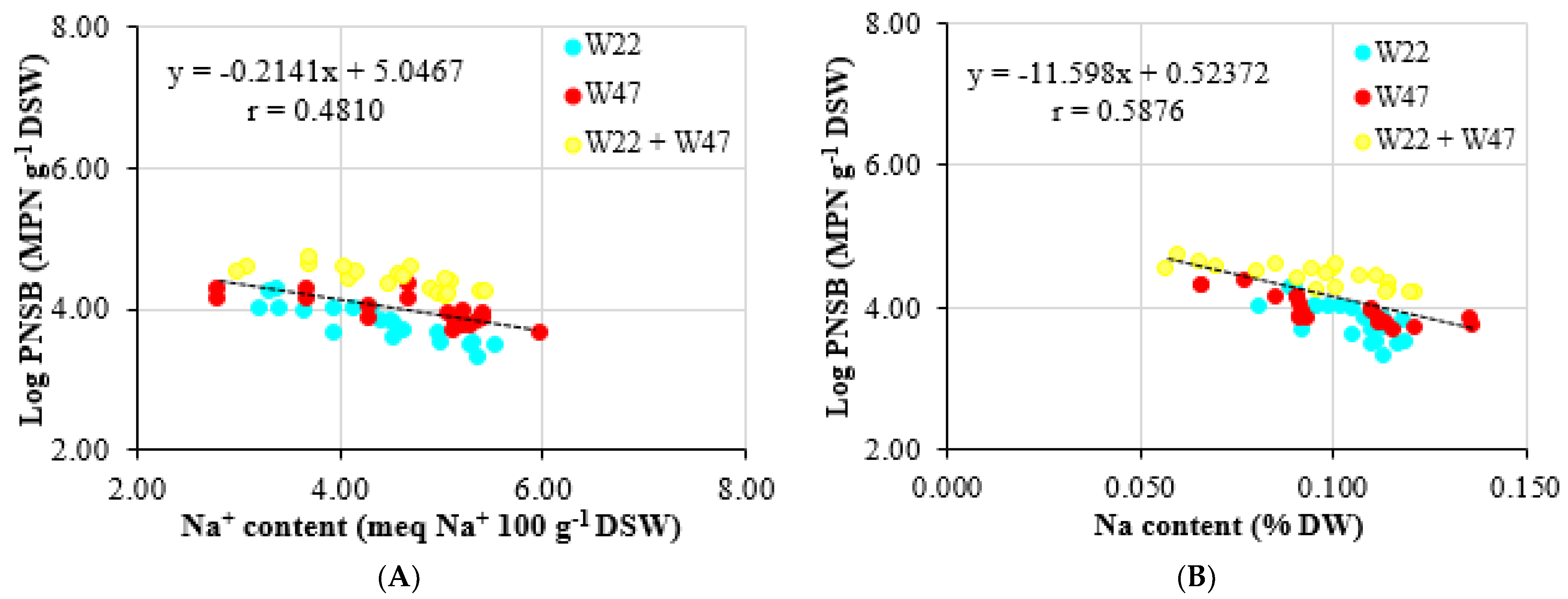
L. sphaeroides W22 and W47 highly negatively correlated to the Na
+ content in the soil (r = 0.6362) (
Figure 4A) and in seeds (r = 0.6185) (
Figure 4B).
Table 2 presents that the plant height and panicle length decreased as the frequency of applying the saline irrigation increased. In detail, the plant height and panicle length decreased in the treatments applied with the saline irrigation four times by 7.63 and 6.03% compared with those in the treatments without the saline irrigation. Previous studies demonstrated that saline concentrations in te water influenced the agronomic parameters more severely in the vegetative and reproductive stages than in the filling and ripening stages [
26]. This result was consistent with the study by Gerona et al. [
3], where the panicle length decreased from 27.9 to 17.3 cm in CSR28 rice in a condition of 10 dS m
−1 salinity. Prodjinoto et al. [
27] also stated that treatment with NaCl resulted in a reduction in plant height from 62.7 to 33.3 cm compared with that in the control treatment without NaCl. However, the percentage of differences in the current study was not as high as the two previous ones; this may be due to differences in rice cultivars (salt-sensitive ones versus the normal one in the study) or the salt concentrations. Salt-contaminated soil or water changes morphologies and modifies metabolism in rice by restricting their development [
28,
29,
30]. However, supplying the ALA-producing bacteria contributed to a reduction in damages on rice growth caused by NaCl because ALA adjusts crucial physiological processes in plants cultivated in saline conditions [
31]. In the current study, the plant height significantly increased in the treatments supplied with the mixture of W22 and W47 compared with those supplied with a single strain of either W22 or W47 (
Table 2). Kantachote et al. [
32] proved that plants in the treatments supplied with a biofertilizer containing bacteria of
R. palustris TK103 were taller than those in the control treatment. In addition, the treatments supplied with bacteria of
R. palustris TK103, PP803 or TN114 ameliorated the panicle length. However, supplying a single strain of either W22 or W47 resulted in an equivalent panicle length, fluctuating between 19.1 and 19.4 cm (
Table 2). Noticeably, the panicles in the treatments supplied with the mixture of W22 and W47 (17.8 cm) were longer than those in the treatments without bacterial supplementation (16.2 cm) (
Table 6). This revealed the role of W22 and W47 in the improvement in rice growth.
Applying the saline irrigation during the vegetative stage adversely affects the yield components and grain yield of plants [
33]. To be more specific, rice is restricted in the biomass, plant height, number of panicles, 1000-seed weight and ratio of filled seeds [
34,
35]. The results in
Table 2 indicate that the yield components, including the number of panicles per pot and the percentage of filled seeds, decreased as the frequency of applying the saline irrigation increased. In detail, the number of panicles per pot started to drop with applying the saline irrigation twice, while, with applying the saline irrigation once, the percentage of filled grain decreased compared with no applied saline irrigation. This result was in accordance with the studies by Hakim et al. and Thitisaksakul et al. [
36,
37], where the number of panicles per pot and the ratio of filled seeds were also decreased by saline irrigation at 4 dS m
−1, in comparison with the control treatment. Moreover, the percentage of filled seeds in the CSR28 rice under the 10 dS m
−1 saline condition was lower than that in the control treatment, with 71% compared to 90.2% [
3]. However, 1000-seed weight in the current study had not been significantly affected by the saline irrigation application (
Table 2). Meanwhile, in the study by Bhusan et al. [
25], the saline-susceptible rice (BRRI dhan 29) in treatments applied with 25 mM NaCl had lower seed weight per pot compared with that in the treatment without irrigated salinity. This could indicate that the OM5451 rice cultivar used in the current study was neither venerable nor tolerant to salinity. Moreover, during the supplementation of the bacterial mixture, the number of panicles per pot and the percentage of filled seeds increased in comparison with those in the treatments supplied with a single strain of either W22 or W47. This was in accordance with studies by Khuong et al. [
11,
13,
38], where supplying bacteria of
Rhodopseudomonas spp., also a PNSB, also contributed to improvements in the number of panicles per square meter and the number of filled seeds per panicle in comparison with the control treatment without bacteria. However, the number of seeds per panicle was equivalent to both individual and mixed supplementation (
Table 2), while, in the study by Kantachote et al. [
32], supplying bacteria of
R. palustris PP803 enhanced the number of seeds per panicle in comparison with the control treatment. This could be because the KDML 105 rice variety used in this study has been identified to have salt-tolerance genotype [
39]. The result in
Table 6 also presents that supplying the mixture of W22 and W47 increased the number of panicles per pot, the number of seeds per panicle and the percentage of filled seeds by 23.9, 19.0 and 3.89%, respectively, resulting in a yield improvement.
Applying the saline irrigation 1–4 times at different vegetative and reproductive stages of rice reduced its yield by 5.76–25.9% in comparison with when no saline irrigation was applied. In particular, the lowest grain yield was recorded in the treatment applied with the saline irrigation four times (
Table 2). This result was in accordance with the study by Baxter et al. [
40], where grain yields in different rice varieties all declined at a dose of 2.4 dS m
−1. The bacteria of ALA-excreting
R. palustris TN114 and PP803 have been determined to be capable of supporting plants to overcome saline stress [
14,
41]. The bacteria of
L. sphaeroides have been proven to be capable of providing ALA [
9,
13,
42] to enable rice to tolerate saline stress [
43,
44] for improving rice grain yield. To be more specific, the treatments applied with either the single strain of W47 or the mixture of the W22 and W47 resulted in higher yield than those in the treatments applied with the single strain of W22 (
Table 2). This result was also consistent with the study by Khuong et al. [
38], where supplying a bacterial mixture provided better yield in comparison with the control treatments and the ones applied with a bacterial strain individually. In the current study, both the single strain of W47 or the mixture of ALA-producing W22 and W47 supported rice growth in saline conditions. Kang et al. [
45]; Naeem et al. [
46]; Nunkaew et al. [
14]; Wongkantrakorn et al. [
44] proved that low ALA concentrations (0.01–30 mg L
−1) are still capable of stimulating plant development in order to minimize the damage caused by salinity. Under the condition of cultivating in saline paddy fields, the maximum ALA content synthesized by the
R. palustris TK103, PP803 and P1 strains was in the range of 1.4–1.7 mg ALA L
−1, leading to higher growth and yield [
16]. The result of the current study also certified that the
Luteovulum genus belonging to the PNSB group enhanced rice yield by 21.5% when the saline irrigation was applied four times at 5‰ concentration (
Table 6). This result was also proven via the correlation between bacterial densities and rice grain yield, with the correlation coefficient (r = 0.6113) (
Figure 5B).
The results in
Table 3 illustrate that the salt-contaminated soil had pH
H2O at roughly 5.99–6.16 and pH
KCl at 5.31–5.83, and it was evaluated as acidic according to the categorization of Shannon et al. [
47]. The EC was in the range of 1.85–2.64 mS cm
−1, classified to be from moderate to high based on the evaluation of Shannon et al. [
47]. The proline content started to rise when the EC reached 2.5 dS m
−1 [
40]. High pH and salinity were considered to be a remarkable obstacle for cultivating plants on saline soils, especially rice vulnerable to salinity [
48]. The above result revealed that the pH
H2O was slightly moderate acidity and pH
KCl was intermediate, which altogether allowed sustainable growth. However, the frequency of the saline irrigation and the supplementation of the bacteria had not influenced pH
H2O and pH
KCl yet, while the EC rose when the saline irrigation was applied in comparison with the no saline irrigation. Nonetheless, the treatments supplied with either the mixture of
L. sphaeroides W22 and W47 or the single of W47 contributed to lowering the EC, in comparison with supplying the single strain of W22 (
Table 3), with a correlation coefficient between bacterial densities and Na
+ concentrations (r = 0.4810) (
Figure 3A). In the same line, supplying the bacterial mixture increased the pH by 0.21 and decreased the EC by 0.31 mS cm
−1 in comparison with the no bacteria case (
Table 7).
The frequency of applying the saline water and the supplementation of the bacteria did not affect the concentration of total N and total P in the soil. However, supplying the bacterial mixture improved the concentrations of NH
4+ and available P in comparison with those in supplying the bacteria individually. This was also expressed via a higher amount of NH
4+ and available P by 5.10 and 7.50 mg kg
−1 in the treatments supplied with the mixture of both the W22 and W47 in comparison with the case of supplying with no bacteria (
Table 7). This can be explained regarding that the strains of W22 and W47 were capable of fixing N and solubilizing P [
9]. Thus, the concentration of NH
4+ and available P all rose. Moreover, this can be expressed through a decreased unavailable P content due to its being solubilized into available forms. The PNSB have been determined to be capable of fixing N and solubilizing P [
13,
15,
38,
49].
Although the CEC content was statistically unchanged between the supplementation of either the mixture of
L. sphaeroides W22 and W47 or a single strain of them, the treatments supplied with either the mixture of the W22 and W47 or the single strain of W47 had lower Na
+ concentrations than those in the treatments supplied with the single strain of W47. Further, in the present study, the correlation relationship between bacterial densities and Na
+ concentrations was also determined, with r = 0.4810 (
Figure 3A). Likewise, supplying the bacterial mixture helped to increase K
+ and Mg
2+ concentrations in comparison with the case of using a single strain (
Table 3). Thus, supplying the mixture of both the W22 and W47 helped to reduce Na
+ and enhance K
+ and Mg
2+ in comparison with supplying a single strain of either W22 or W47 (
Table 7). This can be interpreted that the bacteria of W22 and W47 secrete EPS to immobilize Na
+ [
9]. The reason is that EPS contains functional groups, including –OH and -HOOC, which are able to bind with Na
+ ions [
13,
48,
50]. Furthermore, the PNSB group has demonstrated K solubilizing capacity [
51], leading to a higher K
+ concentration in the treatments with bacteria than those in the ones without bacteria (
Table 7).
The result in
Table 4 highlights that increasing the frequency of applying the saline irrigation changed N, P, K and Na concentrations in seeds. In detail, the N, P and Na contents increased in the treatments applied with the saline irrigation four times. However, applying the saline irrigation from two times above led to a reduction in the K content in seeds. Huang et al. [
52] stated that two rice varieties, saline-tolerant rice (Jinyuan85) and saline-susceptible rice (Liaojing763), had decreased N contents in stems and leaves when being treated with NaCl. In addition, the Na content increased along with an increase in the number of times of applying the saline irrigation. Meanwhile, supplying the ALA-producing PNSB increased N and P concentrations in seeds. To be more specific, supplying the mixture of both
L. sphaeroides W22 and W47 had higher N and P contents in seeds in comparison with the treatments supplied with the bacteria individually.
Figure 3B also illustrates the correlation between bacterial densities and accumulated Na contents (r = 0.5876); that is, the bacteria prevented Na from accumulating within the rice. In addition, supplying the mixture of the W22 and W47 enhanced N, P and K concentrations in stems, leaves and in seeds, except for the P content in stems and leaves. It also restricted the Na content in stems, leaves and seeds in comparison with the results in the treatments without bacteria (
Table 8).
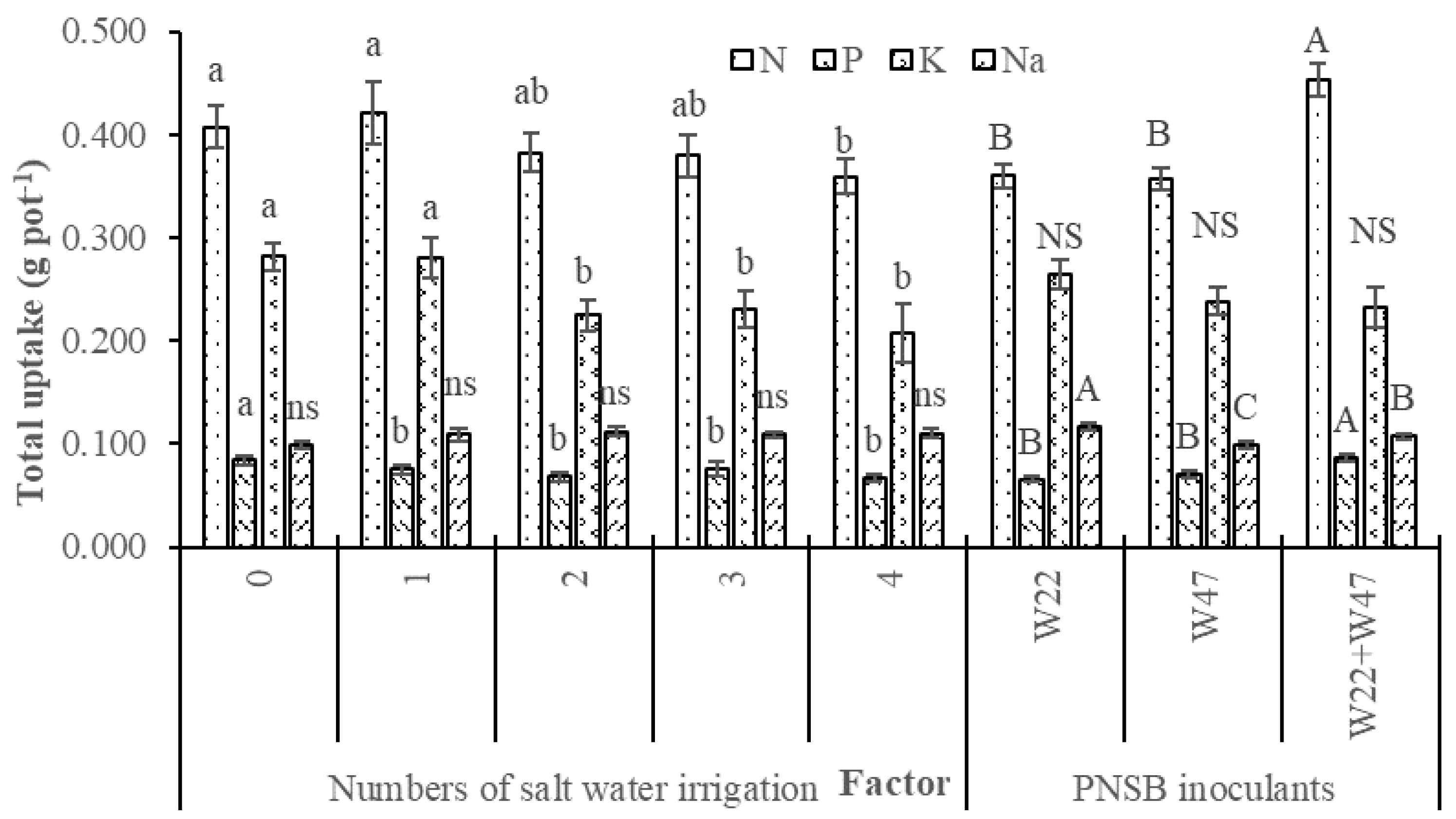
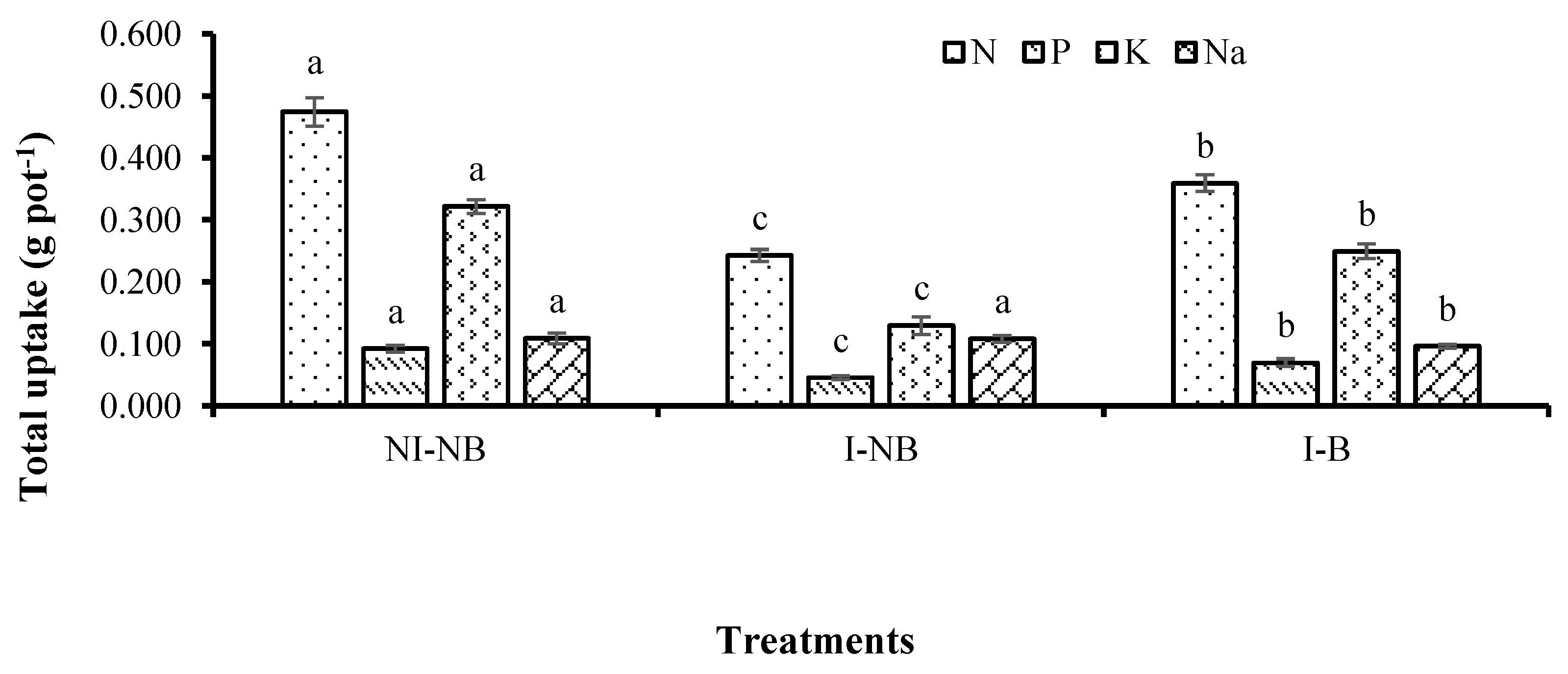
Applying the saline irrigation four times reduced the total N, P and K uptakes and maintained the Na uptake. However, supplying the mixture of
L. sphaeroides W22 and W47 increased the total uptakes of N and P and reduced the total Na uptake in comparison with supplying a single strain of either W22 or W47 (
Figure 2). Moreover, when the mixture of W22 and W47 was applied, the total N, P and K uptakes increased by 32.4, 34.7 and 48.1%, respectively, and the total Na uptake decreased by 11.2% compared with those in the treatments applied with the saline irrigation four times at 4‰ but no bacteria applied (
Figure 2). This demonstrated that the bacteria of W22 and W47 had fixed N and solubilized P and K [
13], resulting in an enhancement in the concentrations of NH
4+, available P and exchangeable K (
Table 7). They also increased in the N, P and K uptakes (
Figure 2), resulting in higher growth and yield components of rice, and ultimately grain yield (
Table 6). This result was consistent with the study by García Morales et al. [
53], where mineral concentrations in the soil or in highly nutritious solution may lead to lower N uptake in rice.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

