
Secondary Metabolites, Other Prospective Substances, and Alternative Approaches That Could Promote Resistance against Phytophthora infestans
 and
and 
Abstract
:1. Introduction
2. Phytophthora infestans
3. Resistance to P. infestans
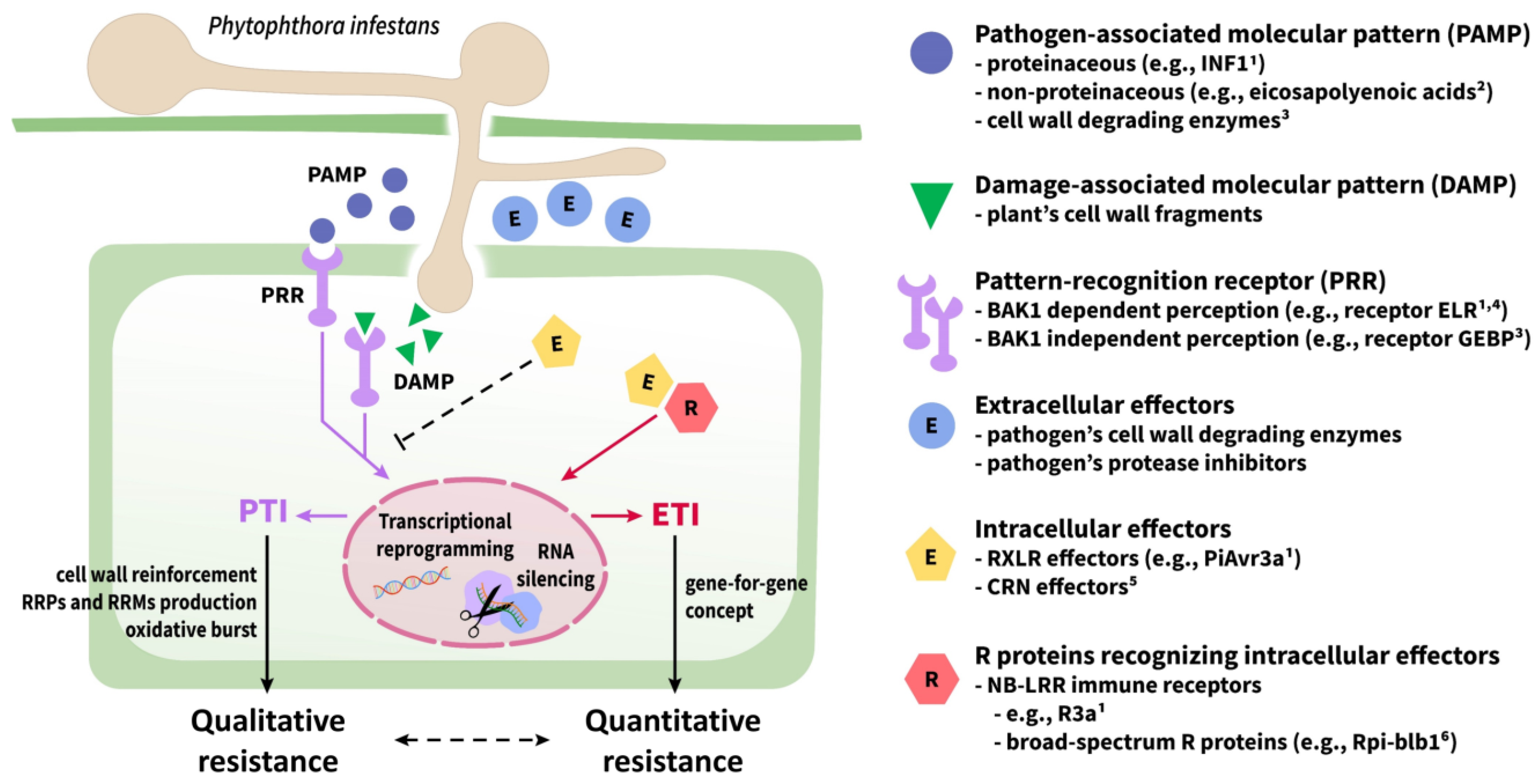
3.1. Plant Innate Immunity

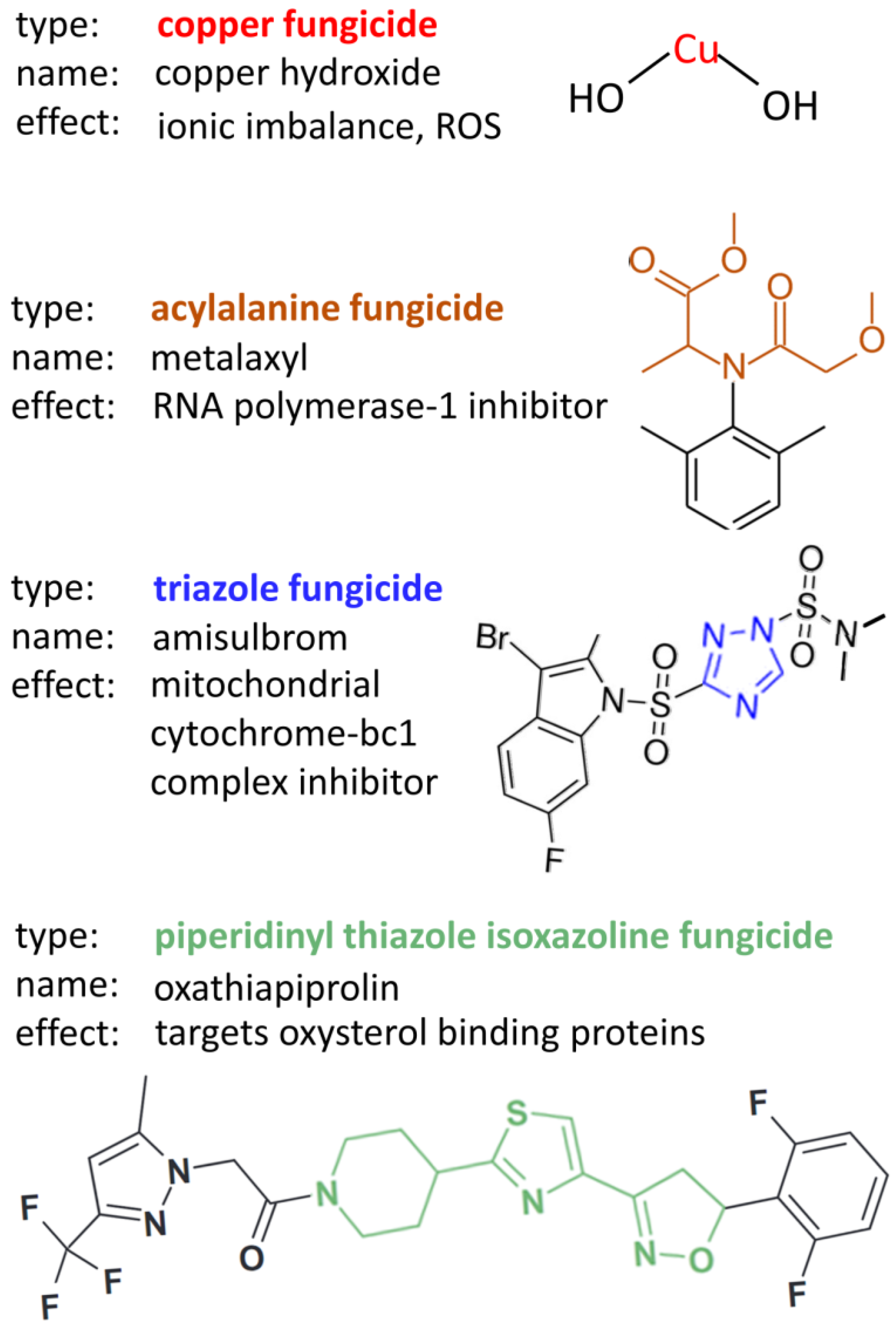
3.2. Chemical Treatment
3.3. Biological Control
3.4. Resistant Cultivars
3.4.1. Natural Resistance
3.4.2. Traditional Breeding
3.4.3. New Breeding Technologies for Potato Improvement
3.4.4. Somatic Hybridization
4. Marker-Assisted Selection
4.1. Genome-Based Analyses
4.2. Metabolome-Based Analyses
4.2.1. Alkaloids
4.2.2. Phenylpropanoids
4.2.3. Terpenoids
4.2.4. Polyamines
4.2.5. Lipidome
4.2.6. Volatiles and Other Compounds of Interest
4.3. Proteome and Peptide-Based Analyses
4.3.1. Candidate Proteins for MAS
4.3.2. Antimicrobial Peptides—Prospective Targets for Enhanced Resistance
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louderback, L.A.; Pavlik, B.M. Starch granule evidence for the earliest potato use in North America. Proc. Natl. Acad. Sci. USA 2017, 114, 7606–7610. [Google Scholar] [CrossRef] [PubMed]
- Hardigan, M.A.; Laimbeer, F.P.E.; Newton, L.; Crisovan, E.; Hamilton, J.P.; Vaillancourt, B.; Wiegert-Rininger, K.; Wood, J.C.; Douches, D.S.; Farré, E.M.; et al. Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. USA 2017, 114, E9999–E10008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Haan, S.; Rodriguez, F. Potato Origin and Production. In Advances in Potato Chemistry and Technology; Jaspreet, S., Lovedeep, K., Eds.; Elsevier: Cambridge, MA, USA, 2016; pp. 1–32. ISBN 978-0-12-800002-1. [Google Scholar]
- Rahaman, M.M.; Shehab, M.K. Water consumption, land use and production patterns of rice, wheat and potato in South Asia during 1988–2012. Sustain. Water Resour. Manag. 2019, 5, 1677–1694. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, P.; Tang, D.; Shang, Y.; Li, C.; Huang, S.; Zhang, C. Inheritance of steroidal glycoalkaloids in potato tuber flesh. J. Integr. Agric. 2019, 18, 2255–2263. [Google Scholar] [CrossRef]
- Yuen, J. Pathogens which threaten food security: Phytophthora infestans, the potato late blight pathogen. Food Secur. 2021, 13, 247–253. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Salazar, L.F. Emerging and Re-emerging Potato Diseases in the Andes. Potato Res. 2006, 49, 43–47. [Google Scholar] [CrossRef]
- Saville, A.C.; Ristaino, J.B. Global historic pandemics caused by the FAM-1 genotype of Phytophthora infestans on six continents. Sci. Rep. 2021, 11, 12335. [Google Scholar] [CrossRef]
- Yoshida, K.; Schuenemann, V.J.; Cano, L.M.; Pais, M.; Mishra, B.; Sharma, R.; Lanz, C.; Martin, F.N.; Kamoun, S.; Krause, J.; et al. The rise and fall of the Phytophthora infestans lineage that triggered the Irish potato famine. eLife 2013, 2, e00731. [Google Scholar] [CrossRef]
- Goss, E.M.; Tabima, J.F.; Cooke, D.E.L.; Restrepo, S.; Fry, W.E.; Forbes, G.A.; Fieland, V.J.; Cardenas, M.; Grünwald, N.J. The Irish potato famine pathogen Phytophthora infestans originated in central Mexico rather than the Andes. Proc. Natl. Acad. Sci. USA 2014, 111, 8791–8796. [Google Scholar] [CrossRef]
- Martin, M.D.; Vieira, F.G.; Ho, S.Y.W.; Wales, N.; Schubert, M.; Seguin-Orlando, A.; Ristaino, J.B.; Gilbert, M.T.P. Genomic Characterization of a South American Phytophthora hybrid mandates reassessment of the geographic origins of Phytophthora infestans. Mol. Biol. Evol. 2016, 33, 478–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haverkort, A.J.; Struik, P.C.; Visser, R.G.F.; Jacobsen, E. Applied Biotechnology to Combat Late Blight in Potato Caused by Phytophthora infestans. Potato Res. 2009, 52, 249–264. [Google Scholar] [CrossRef]
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; de la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging oomycete threats to plants and animals. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abad, Z.G.; Burgess, T.I.; Redford, A.J.; Bienapfl, J.C.; Srivastava, S.; Mathew, R.; Jennings, K. IDphy: An International Online Resource for Molecular and Morphological Identification of Phytophthora. Plant Dis. 2023, 107, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Schoina, C.; Govers, F. The oomycete Phytophthora infestans, the irish potato famine pathogen. In Principles of Plant-Microbe Interactions—Microbes for Sustainable Agriculture; Springer Cham: Heidelberg, Germany, 2015; pp. 371–378. [Google Scholar]
- Chepsergon, J.; Motaung, T.E.; Bellieny-Rabelo, D.; Moleleki, L.N. Organize, Don’t Agonize: Strategic Success of Phytophthora Species. Microorganisms 2020, 8, 917. [Google Scholar] [CrossRef]
- Bronkhorst, J.; Kasteel, M.; van Veen, S.; Clough, J.M.; Kots, K.; Buijs, J.; van der Gucht, J.; Ketelaar, T.; Govers, F.; Sprakel, J. A slicing mechanism facilitates host entry by plant-pathogenic Phytophthora. Nat. Microbiol. 2021, 6, 1000–1006. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Cooke, D.E.L.; Acuña, I.; Muñoz, M. CHAPTER 6: The threat of late blight to global food security. In Emerging Plant Diseases and Global Food Security; Ristaino, J.B., Records, A., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2020; pp. 101–132. [Google Scholar]
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, J.W. Trichoderma Volatile Organic Compounds as a Biofumigation Tool against Late Blight Pathogen Phytophthora infestans in Postharvest Potato Tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How does Phytophthora infestans evade control efforts? Modern insight into the late blight disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five reasons to consider Phytophthora infestans a reemerging pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [Green Version]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U.E.U. Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Fry, W.E. Phytophthora infestans: New tools (and old ones) lead to new understanding and precision management. Annu. Rev. Phytopathol. 2016, 54, 529–547. [Google Scholar] [CrossRef]
- Ni, M.; Feretzaki, M.; Sun, S.; Wang, X.; Heitman, J. Sex in fungi. Annu. Rev. Genet. 2011, 45, 405. [Google Scholar] [CrossRef]
- Tzelepis, G.; Hodén, K.P.; Fogelqvist, J.; Åsman, A.K.M.; Vetukuri, R.R.; Dixelius, C. Dominance of mating type A1 and indication of epigenetic effects during early stages of mating in Phytophthora infestans. Front. Microbiol. 2020, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Fry, W.E. Phytophthora infestans: The itinerant invader; “late blight”: The persistent disease. Phytoparasitica 2020, 48, 87–94. [Google Scholar] [CrossRef]
- Li, Y.; Shen, H.; Zhou, Q.; Qian, K.; Van Der Lee, T.; Huang, S. Changing ploidy as a strategy: The Irish potato famine pathogen shifts ploidy in relation to its sexuality. Mol. Plant-Microbe Interact. 2017, 30, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Naveed, Z.A.; Wei, X.; Chen, J.; Mubeen, H.; Ali, G.S. The PTI to ETI continuum in Phytophthora-plant interactions. Front. Plant Sci. 2020, 11, 2030. [Google Scholar] [CrossRef] [PubMed]
- Amaro, T.M.M.M.; Thilliez, G.J.A.; Motion, G.B.; Huitema, E. A perspective on CRN proteins in the genomics age: Evolution, classification, delivery and function revisited. Front. Plant Sci. 2017, 8, 99. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Verzaux, E.; Chaparro-Garcia, A.; Bijsterbosch, G.; Keizer, L.C.P.; Zhou, J.; Liebrand, T.W.H.; Xie, C.; Govers, F.; Robatzek, S.; et al. Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants 2015, 1, 15034. [Google Scholar] [CrossRef]
- Zhang, H.; Li, F.; Li, Z.; Cheng, J.; Chen, X.; Wang, Q.; Joosten, M.H.A.J.; Shan, W.; Du, Y. Potato StMPK7 is a downstream component of StMKK1 and promotes resistance to the oomycete pathogen Phytophthora infestans. Mol. Plant Pathol. 2021, 22, 644–657. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.A.A.; Raffaele, S.; Vossen, J.H.; Champouret, N.; Oliva, R.; Segretin, M.E.; Rietman, H.; Cano, L.M.; Lokossou, A.; Kessel, G.; et al. Understanding and exploiting late blight resistance in the age of effectors. Annu. Rev. Phytopathol. 2011, 49, 507–531. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Ma, W. How to win a tug-of-war: The adaptive evolution of Phytophthora effectors. Curr. Opin. Plant Biol. 2021, 62, 102027. [Google Scholar] [CrossRef] [PubMed]
- Ah-Fong, A.M.V.; Shrivastava, J.; Judelson, H.S. Lifestyle, gene gain and loss, and transcriptional remodeling cause divergence in the transcriptomes of Phytophthora infestans and Pythium ultimum during potato tuber colonization. BMC Genom. 2017, 18, 764. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Gu, B.; Huang, G.; Tian, Y.; Quan, J.; Lindqvist-Kreuze, H.; Shan, W. Conserved RXLR effector genes of Phytophthora infestans expressed at the early stage of potato infection are suppressive to host defense. Front. Plant Sci. 2017, 8, 2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Kamoun, S.; Zody, M.C.; Jiang, R.H.Y.; Handsaker, R.E.; Cano, L.M.; Grabherr, M.; Kodira, C.D.; Raffaele, S.; Torto-Alalibo, T.; et al. Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 2009, 461, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Chen, H.; Zhang, X.; Gao, C.; Huang, J.; Lü, L.; Shen, D.; Wang, L.; Huang, C.; Ye, W.; et al. Genome analysis of two newly emerged potato late blight isolates sheds light on pathogen adaptation and provides tools for disease management. Phytopathology 2021, 111, 96–107. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.L.; Moury, B.; Caffier, V.; Montarry, J.; Kerlan, M.C.; Fournet, S.; Durel, C.E.; Delourme, R. Quantitative resistance to plant pathogens in pyramiding strategies for durable crop protection. Front. Plant Sci. 2017, 8, 1838. [Google Scholar] [CrossRef] [Green Version]
- Kahlon, P.S.; Verin, M.; Hückelhoven, R.; Stam, R. Quantitative resistance differences between and within natural populations of Solanum chilense against the oomycete pathogen Phytophthora infestans. Ecol. Evol. 2021, 11, 7768–7778. [Google Scholar] [CrossRef] [PubMed]
- Saubeau, G.; Perrin, F.; Marnet, N.; Andrivon, D.; Val, F. Hormone signalling pathways are differentially involved in quantitative resistance of potato to Phytophthora infestans. Plant Pathol. 2016, 65, 342–352. [Google Scholar] [CrossRef]
- Mundt, C.C. Durable resistance: A key to sustainable management of pathogens and pests. Infect. Genet. Evol. 2014, 27, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Rojas, D.K.J.; Sedano, J.C.S.; Ballvora, A.; Léon, J.; Vásquez, T.M. Novel organ-specific genetic factors for quantitative resistance to late blight in potato. PLoS ONE 2019, 14, e0213818. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, M.F.; Angarita, M.; Delgado, M.C.; García, C.; Jiménez-Gomez, J.; Gebhardt, C.; Mosquera, T. Identification of novel associations of candidate genes with resistance to late blight in Solanum tuberosum group phureja. Front. Plant Sci. 2017, 8, 1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosquera, T.; Alvarez, M.F.; Jiménez-Gómez, J.M.; Muktar, M.S.; Paulo, M.J.; Steinemann, S.; Li, J.; Draffehn, A.; Hofmann, A.; Lübeck, J.; et al. Targeted and untargeted approaches unravel novel candidate genes and diagnostic SNPs for quantitative resistance of the potato (Solanum tuberosum L.) to Phytophthora infestans causing the late blight disease. PLoS ONE 2016, 11, e0156254. [Google Scholar] [CrossRef] [Green Version]
- Santa, J.D.; Berdugo-Cely, J.; Cely-Pardo, L.; Soto-Suárez, M.; Mosquera, T.; Galeano, C.H.M. QTL analysis reveals quantitative resistant loci for Phytophthora infestans and Tecia solanivora in tetraploid potato (Solanum tuberosum L.). PLoS ONE 2018, 13, e0199716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogendra, K.N.; Kushalappa, A.C.; Sarmiento, F.; Rodriguez, E.; Mosquera, T.; Yogendra, K.N.; Kushalappa, A.C.; Sarmiento, F.; Rodriguez, E.; Mosquera, T. Metabolomics deciphers quantitative resistance mechanisms in diploid potato clones against late blight. Funct. Plant Biol. 2014, 42, 284–298. [Google Scholar] [CrossRef]
- Meade, F.; Hutten, R.; Wagener, S.; Prigge, V.; Dalton, E.; Kirk, H.G.; Griffin, D.; Milbourne, D. Detection of novel QTLs for late blight resistance derived from the wild potato species Solanum microdontum and Solanum pampasense. Genes 2020, 11, 732. [Google Scholar] [CrossRef]
- Lokossou, A.A.; Rietman, H.; Wang, M.; Krenek, P.; Van Der Schoot, H.; Henken, B.; Hoekstra, R.; Vleeshouwers, V.G.A.A.; Van Der Vossen, E.A.G.; Visser, R.G.F.; et al. Diversity, distribution, and evolution of Solanum bulbocastanum late blight resistance genes. Mol. Plant. Microbe Interact. 2010, 23, 1206–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedyukhina, E.G.; Kamzolova, S.V.; Vainshtein, M.B. Arachidonic acid as an elicitor of the plant defense response to phytopathogens. Chem. Biol. Technol. Agric. 2014, 1, 18. [Google Scholar] [CrossRef] [Green Version]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete interactions with plants: Infection strategies and resistance principles. Microbiol. Mol. Biol. Rev. 2015, 79, 263–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Si, W.; Gao, R.; Zhang, X.; Yang, S. Evolutionary analysis of RB/Rpi-blb1 locus in the Solanaceae family. Mol. Genet. Genom. 2015, 290, 2173–2186. [Google Scholar] [CrossRef]
- Kolbe, W. Importance of potato blight control exemplified by Höfchen long-term trial (1943–1982), and historical development. Pflanzenschutz Nachr. Bayer 1982, 35, 247–290. [Google Scholar]
- La Torre, A.; Righi, L.; Iovino, V.; Battaglia, V. Control of late blight in organic farming with low copper dosages or natural products as alternatives to copper. Eur. J. Plant Pathol. 2019, 155, 769–778. [Google Scholar] [CrossRef]
- Liu, Y.; Langemeier, M.R.; Small, I.M.; Joseph, L.; Fry, W.E. Risk Management Strategies using precision agriculture technology to manage potato late blight. Agron. J. 2017, 109, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Y. Root treatment with oxathiapiprolin, benthiavalicarb or their mixture provides prolonged systemic protection against oomycete foliar pathogens. PLoS ONE 2020, 15, e0227556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, E.R.; Migliano, L.E.; Chen, Y.; Gevens, A.J. Mefenoxam Sensitivity in US-8 and US-23 Phytophthora infestans from Wisconsin. Plant Health Prog. 2021, 22, 272–280. [Google Scholar] [CrossRef]
- Mboup, M.K.; Sweigard, J.W.; Carroll, A.; Jaworska, G.; Genet, J. Genetic mechanism, baseline sensitivity and risk of resistance to oxathiapiprolin in oomycetes. Pest Manag. Sci. 2022, 78, 905–913. [Google Scholar] [CrossRef]
- Liljeroth, E.; Lankinen, Å.; Wiik, L.; Burra, D.D.; Alexandersson, E.; Andreasson, E. Potassium phosphite combined with reduced doses of fungicides provides efficient protection against potato late blight in large-scale field trials. Crop. Prot. 2016, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Mulugeta, T.; Abreha, K.; Tekie, H.; Mulatu, B.; Yesuf, M.; Andreasson, E.; Liljeroth, E.; Alexandersson, E. Phosphite protects against potato and tomato late blight in tropical climates and has varying toxicity depending on the Phytophthora infestans isolate. Crop. Prot. 2019, 121, 139–146. [Google Scholar] [CrossRef]
- Liljeroth, E.; Lankinen, Å.; Andreasson, E.; Alexandersson, E. Phosphite integrated in late blight treatment strategies in starch potato does not cause residues in the starch product. Plant Dis. 2020, 104, 3026–3032. [Google Scholar] [CrossRef]
- Sharma, S.; Sundaresha, S.; Tiwari, R.K.; Sagar, V.; Lal, M. Effect of phosphorous acid on late blight disease mitigation and minimization of fungicide doses under field conditions. J. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Robinson, J.R.; Isikhuemhen, O.S.; Anike, F.N. Fungal–metal interactions: A review of toxicity and homeostasis. J. Fungi 2021, 7, 225. [Google Scholar] [CrossRef]
- Pasteris, R.J.; Hanagan, M.A.; Bisaha, J.J.; Finkelstein, B.L.; Hoffman, L.E.; Gregory, V.; Andreassi, J.L.; Sweigard, J.A.; Klyashchitsky, B.A.; Henry, Y.T.; et al. Discovery of oxathiapiprolin, a new oomycete fungicide that targets an oxysterol binding protein. Bioorg. Med. Chem. 2016, 24, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.J.; Hayes, A.L. Mode of action of the systemic fungicides furalaxyl, metalaxyl and ofurace. Pestic. Sci. 1982, 13, 330–339. [Google Scholar] [CrossRef]
- Dreinert, A.; Wolf, A.; Mentzel, T.; Meunier, B.; Fehr, M. The cytochrome bc complex inhibitor Ametoctradin has an unusual binding mode. Biochim. Biophys. Acta-Bioenerg. 2018, 1859, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Lal, M.; Sharma, S.; Yadav, S.; Kumar, S. Management of late blight of potato. In Potato—From Incas to All Over the World; InTech: London, UK, 2018. [Google Scholar]
- Miles, L.A.; Lopera, C.A.; González, S.; de García, M.C.C.; Franco, A.E.; Restrepo, S. Exploring the biocontrol potential of fungal endophytes from an Andean Colombian Paramo ecosystem. BioControl 2012, 57, 697–710. [Google Scholar] [CrossRef]
- El-Hasan, A.; Ngatia, G.; Link, T.I.; Voegele, R.T. Isolation, identification, and biocontrol potential of root fungal endophytes associated with solanaceous plants against potato late blight (Phytophthora infestans). Plants 2022, 11, 1605. [Google Scholar] [CrossRef]
- de Vries, S.; von Dahlen, J.K.; Schnake, A.; Ginschel, S.; Schulz, B.; Rose, L.E. Broad-spectrum inhibition of Phytophthora infestans by fungal endophytes. FEMS Microbiol. Ecol. 2018, 94, fiy037. [Google Scholar] [CrossRef] [Green Version]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P.A. Screening for novel biocontrol agents applicable in plant disease management—A review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Lalaymia, I.; Naveau, F.; Arguelles Arias, A.; Ongena, M.; Picaud, T.; Declerck, S.; Calonne-Salmon, M. Screening and efficacy evaluation of antagonistic fungi against Phytophthora infestans and combination with arbuscular mycorrhizal fungi for biocontrol of late blight in potato. Front. Agron. 2022, 4, 948309. [Google Scholar] [CrossRef]
- Yusuf, Y.; Izzet, K.; E Ayhan, G.K.; Ibrahim, D.; Nezhun, G.R.; Halit, C.A.; Mark, W. In vitro antifungal activities of 26 plant extracts on mycelial growth of Phytophthora infestans (Mont.) de Bary. Afr. J. Biotechnol. 2011, 10, 2625–2629. [Google Scholar] [CrossRef] [Green Version]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Arafa, R.A.; Kamel, S.M.; Taher, D.I.; Solberg, S.Ø.; Rakha, M.T. Leaf extracts from resistant wild tomato can be used to control late blight (Phytophthora infestans) in the cultivated tomato. Plants 2022, 11, 1824. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, L.M.; Trexler, R.V.; Malik, R.J.; Hockett, K.L.; Bell, T.H. The inherent conflicts in developing soil microbial inoculants. Trends Biotechnol. 2019, 37, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, X.; Hou, Y.; Xia, X.; Zhu, Z.; Huang, A.; Feng, S.; Li, P.; Shi, L.; Dong, P. Isolation and screening of antagonistic endophytes against Phytophthora infestans and preliminary exploration on anti-oomycete mechanism of Bacillus velezensis 6-5. Plants 2023, 12, 909. [Google Scholar] [CrossRef] [PubMed]
- Troussieux, S.; Gilgen, A.; Souche, J.-L. A New biocontrol tool to fight potato late blight based on Willaertia magna C2c Maky lysate. Plants 2022, 11, 2756. [Google Scholar] [CrossRef]
- De Vrieze, M.; Germanier, F.; Vuille, N.; Weisskopf, L. Combining different potato-associated Pseudomonas strains for improved biocontrol of Phytophthora infestans. Front. Microbiol. 2018, 9, 2573. [Google Scholar] [CrossRef]
- Stephan, D.; Schmitt, A.; Carvalho, S.M.; Seddon, B.; Koch, E. Evaluation of biocontrol preparations and plant extracts for the control of Phytophthora infestans on potato leaves. Eur. J. Plant Pathol. 2005, 112, 235–246. [Google Scholar] [CrossRef]
- Wu, Z.H.; Ma, Q.; Sun, Z.N.; Cui, H.C.; Liu, H.R. Biocontrol mechanism of Myxococcus fulvus B25-I-3 against Phytophthora infestans and its control efficiency on potato late blight. Folia Microbiol. 2021, 66, 555–567. [Google Scholar] [CrossRef]
- Kadiri, M.; Sevugapperumal, N.; Nallusamy, S.; Ragunathan, J.; Ganesan, M.V.; Alfarraj, S.; Ansari, M.J.; Sayyed, R.Z.; Lim, H.R.; Show, P.L. Pan-genome analysis and molecular docking unveil the biocontrol potential of Bacillus velezensis VB7 against Phytophthora infestans. Microbiol. Res. 2023, 268, 127277. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Kida, M.; Arita, M.; Takahashi, M. Induction of systemic resistance by Fusarium oxysporum MT0062 in solanaceous crops. Japanese J. Phytopathol. 1992, 58, 16–22. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- Bethke, P.C.; Halterman, D.A.; Jansky, S.H. Potato germplasm enhancement enters the genomics era. Agronomy 2019, 9, 575. [Google Scholar] [CrossRef] [Green Version]
- Machida-Hirano, R. Diversity of potato genetic resources. Breed. Sci. 2015, 65, 26–40. [Google Scholar] [CrossRef] [Green Version]
- Slater, A.T.; Cogan, N.O.I.; Hayes, B.J.; Schultz, L.; Dale, M.F.B.; Bryan, G.J.; Forster, J.W. Improving breeding efficiency in potato using molecular and quantitative genetics. Theor. Appl. Genet. 2014, 127, 2279–2292. [Google Scholar] [CrossRef]
- Rogozina, E.V.; Gurina, A.A.; Chalaya, N.A.; Zoteyeva, N.M.; Kuznetsova, M.A.; Beketova, M.P.; Muratova, O.A.; Sokolova, E.A.; Drobyazina, P.E.; Khavkin, E.E. Diversity of Late Blight Resistance Genes in the VIR Potato Collection. Plants 2023, 12, 273. [Google Scholar] [CrossRef] [PubMed]
- Perez, W.; Alarcon, L.; Rojas, T.; Correa, Y.; Juarez, H.; Andrade-Piedra, J.L.; Anglin, N.L.; Ellis, D. Screening South American potato landraces and potato wild relatives for novel sources of late blight resistance. Plant Dis. 2022, 106, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Khiutti, A.; Spooner, D.M.; Jansky, S.H.; Halterman, D.A. Testing taxonomic predictivity of foliar and tuber resistance to Phytophthora infestans in wild relatives of potato. Phytopathology 2015, 105, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Karki, H.S.; Jansky, S.H.; Halterman, D.A. Screening of wild potatoes identifies new sources of late blight resistance. Plant Dis. 2021, 105, 368–376. [Google Scholar] [CrossRef]
- Van Weymers, P.S.M.; Baker, K.; Chen, X.; Harrower, B.; Cooke, D.E.L.; Gilroy, E.M.; Birch, P.R.J.; Thilliez, G.J.A.; Lees, A.K.; Lynott, J.S.; et al. Utilizing “omic” technologies to identify and prioritize novel sources of resistance to the oomycete pathogen Phytophthora infestans in potato germplasm collections. Front. Plant Sci. 2016, 7, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann-Pfabe, S.; Hammann, T.; Kruse, J.; Dehmer, K.J. Screening of wild potato genetic resources for combined resistance to late blight on tubers and pale potato cyst nematodes. Euphytica 2019, 215, 48. [Google Scholar] [CrossRef]
- Bethke, P.C.; Halterman, D.A.; Jansky, S. Are We Getting Better at Using Wild Potato Species in Light of New Tools? Crop Sci. 2017, 57, 1241–1258. [Google Scholar] [CrossRef] [Green Version]
- Paluchowska, P.; Śliwka, J.; Yin, Z. Late blight resistance genes in potato breeding. Planta 2022, 255, 127. [Google Scholar] [CrossRef] [PubMed]
- Blossei, J.; Gäbelein, R.; Hammann, T.; Uptmoor, R. Late blight resistance in wild potato species—Resources for future potato (Solanum tuberosum) breeding. Plant Breed. 2022, 141, 314–331. [Google Scholar] [CrossRef]
- Abuley, I.K.; Hansen, J.G. Characterization of the Level and Type of Resistance of Potato Varieties to Late Blight (Phytophthora infestans). Phytopathology 2022, 112, 1917–1927. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato Breeding at the Scottish Plant Breeding Station and the Scottish Crop Research Institute: 1920–2008. Potato Res. 2009, 52, 141–172. [Google Scholar] [CrossRef]
- Jansky, S.H.; De Jong, W.S.; Douches, D.S.; Haynes, K.G.; Holm, D.G. Cultivar Improvement with Exotic Germplasm: An Example from Potato. In The Wild Solanums Genomes. Compendium of Plant Genomes; Carputo, D., Aversano, R., Ercolano, M.R., Eds.; Springer: Cham, Switzerland, 2021; pp. 215–230. ISBN 978-3-030-30342-6. [Google Scholar]
- Akino, S.; Takemoto, D.; Hosaka, K. Phytophthora infestans: A review of past and current studies on potato late blight. J. Gen. Plant Pathol. 2014, 80, 24–37. [Google Scholar] [CrossRef]
- Rietman, H.; Bijsterbosch, G.; Cano, L.M.; Lee, H.R.; Vossen, J.H.; Jacobsen, E.; Visser, R.G.F.; Kamoun, S.; Vleeshouwers, V.G.A.A. Qualitative and Quantitative Late Blight Resistance in the Potato Cultivar Sarpo Mira Is Determined by the Perception of Five Distinct RXLR Effectors. Mol. Plant-Microbe Interact. 2012, 25, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Blatnik, E.; Horvat, M.; Berne, S.; Humar, M.; Dolničar, P.; Meglič, V. Late Blight Resistance Conferred by Rpi-Smira2/R8 in Potato Genotypes In Vitro Depends on the Genetic Background. Plants 2022, 11, 1319. [Google Scholar] [CrossRef]
- Ali, A.; Moushib, L.I.; Lenman, M.; Levander, F.; Olsson, K.; Carlson-Nilson, U.; Zoteyeva, N.; Liljeroth, E.; Andreasson, E. Paranoid potato. Plant Signal. Behav. 2012, 7, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.; Eusufzai, T.K.; Ansarey, F.H.; Hasan, M.M.; Nahiyan, A.S.M. A breeding approach to enhance late blight resistance in potato. J. Hortic. Sci. Biotechnol. 2022, 97, 719–729. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Review and Analysis of Limitations in Ways to Improve Conventional Potato Breeding. Potato Res. 2017, 60, 171–193. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Vossen, J.H.; Visser, R.G.F. Durable late blight resistance in potato through dynamic varieties obtained by cisgenesis: Scientific and societal advances in the DuRPh project. Potato Res. 2016, 59, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Keijzer, P.; van Bueren, E.T.L.; Engelen, C.J.M.; Hutten, R.C.B. Breeding Late Blight Resistant Potatoes for Organic Farming—A Collaborative Model of Participatory Plant Breeding: The Bioimpuls Project. Potato Res. 2022, 65, 349–377. [Google Scholar] [CrossRef]
- Zhu, S.; Li, Y.; Vossen, J.H.; Visser, R.G.F.; Jacobsen, E. Functional stacking of three resistance genes against Phytophthora infestans in potato. Transgenic Res. 2012, 21, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Witek, K.; Lin, X.; Karki, H.S.; Jupe, F.; Witek, A.I.; Steuernagel, B.; Stam, R.; van Oosterhout, C.; Fairhead, S.; Heal, R.; et al. A complex resistance locus in Solanum americanum recognizes a conserved Phytophthora effector. Nat. Plants 2021, 7, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Taoutaou, A.; Berindean, I.V.; Chemmam, M.K.; Beninal, L.; Rida, S.; Khelifi, L.; Bouznad, Z.; Racz, I.; Ona, A.; Muntean, L. Defeated stacked resistance genes induce a delay in disease manifestation in the pathosystem Solanum tuberosum—Phytophthora infestans. Agronomy 2023, 13, 1255. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Thieme, R.; König, J.; Nachtigall, M.; Hammann, T.; Denes, T.-E.E.; Kruppa, K.; Molnár-Láng, M. Introgression of Two Broad-Spectrum Late Blight Resistance Genes, Rpi-Blb1 and Rpi-Blb3, From Solanum bulbocastanum Dun Plus Race-Specific R Genes into Potato Pre-breeding Lines. Front. Plant Sci. 2020, 11, 699. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Plich, J.; Janiszewska, M.; Smyda-Dajmund, P.; Sobkowiak, S.; Śliwka, J. Marker-assisted pyramiding of potato late blight resistance genes Rpi-rzc1 and Rpi-phu1 on di- and tetraploid levels. Mol. Breed. 2020, 40, 89. [Google Scholar] [CrossRef]
- Haesaert, G.; Vossen, J.H.; Custers, R.; De Loose, M.; Haverkort, A.; Heremans, B.; Hutten, R.; Kessel, G.; Landschoot, S.; Droogenbroeck, B.; et al. Transformation of the potato variety Desiree with single or multiple resistance genes increases resistance to late blight under field conditions. Crop Prot. 2015, 77, 163–175. [Google Scholar] [CrossRef]
- Jo, K.R.; Zhu, S.; Bai, Y.; Hutten, R.C.B.; Kessel, G.J.T.; Vleeshouwers, V.G.A.A.; Jacobsen, E.; Visser, R.G.F.; Vossen, J.H. Problematic Crops: 1. Potatoes. In Plant Pathogen Resistance Biotechnology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 171–191. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.R.; Jo, K.R.; Mortazavian, S.M.M.; Huigen, D.J.; Evenhuis, B.; Kessel, G.; Visser, R.G.F.; Jacobsen, E.; Vossen, J.H. Broad spectrum late blight resistance in potato differential set plants MaR8 and MaR9 is conferred by multiple stacked R genes. Theor. Appl. Genet. 2012, 124, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Ghislain, M.; Byarugaba, A.A.; Magembe, E.; Njoroge, A.; Rivera, C.; Román, M.L.; Tovar, J.C.; Gamboa, S.; Forbes, G.A.; Kreuze, J.F.; et al. Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnol. J. 2019, 17, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Zaidi, S.S.-A.; Shakir, S.; Mansoor, S. Applications of New Breeding Technologies for Potato Improvement. Front. Plant Sci. 2018, 9, 925. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Schipper, D.; Jacobsen, E.; Visser, R.G.F.; Govers, F.; Bouwmeester, K.; Bai, Y. Silencing susceptibility genes in potato hinders primary infection with Phytophthora infestans at different stages. Hortic. Res. 2022, 9, uhab058. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Wolters, A.M.A.; Vossen, J.H.; Rouwet, M.E.; Loonen, A.E.H.M.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Silencing of six susceptibility genes results in potato late blight resistance. Transgenic Res. 2016, 25, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Petersen, B.L.; Andreasson, E. Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci. Rep. 2021, 11, 4487. [Google Scholar] [CrossRef] [PubMed]
- Orczyk, W.; Przetakiewicz, J.; Nadolska-Orczyk, A. Somatic hybrids of Solanum tuberosum—Application to genetics and breeding. Plant Cell Tissue Organ Cult. 2003, 74, 1–13. [Google Scholar] [CrossRef]
- Sedlák, P.; Sedláková, V.; Vašek, J.; Zeka, D.; Čílová, D.; Melounová, M.; Orsák, M.; Domkářová, J.; Doležal, P.; Vejl, P. Phenotypic, molecular and biochemical evaluation of somatic hybrids between Solanum tuberosum and S. bulbocastanum. Sci. Rep. 2022, 12, 4484. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Rawat, S.; Luthra, S.K.; Zinta, R.; Sahu, S.; Varshney, S.; Kumar, V.; Dalamu, D.; Mandadi, N.; Kumar, M.; et al. Genome sequence analysis provides insights on genomic variation and late blight resistance genes in potato somatic hybrid (parents and progeny). Mol. Biol. Rep. 2021, 48, 623–635. [Google Scholar] [CrossRef]
- Cruceriu, D.; Erdely-Molnar, I.; Diaconeasa, Z.; Margineanu, A.M.; Aurori, A.; Rakosy-Tican, E. Comparative characterization of somatic hybrids of Solanum bulbocastanum + S. tuberosum Cv. ‘rasant’ with their parents in relation to biochemical responses to wound stress and trichome composition. Stud. Univ. Babes-Bolyai Chem. 2020, 65, 133–148. [Google Scholar] [CrossRef]
- Cruceriu, D.; Molnar, I.; Diaconeasa, Z.; Aurori, A.; Socaciu, C.; Rakosy-Tican, E. In Vitro Culture as a Stressful Factor Triggers Changes in Polyphenols, Flavonoids and Antioxidant Activity in Somatic Hybrids between Solanum tuberosum and S. bulbocastanum and their Respective Parents. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Rakosy-Tican, E.; Thieme, R.; Nachtigall, M.; Molnar, I.; Denes, T.E. The recipient potato cultivar influences the genetic makeup of the somatic hybrids between five potato cultivars and one cloned accession of sexually incompatible species Solanum bulbocastanum Dun. Plant Cell Tissue Organ Cult. 2015, 122, 395–407. [Google Scholar] [CrossRef]
- Kushalappa, A.C.; Gunnaiah, R. Metabolo-proteomics to discover plant biotic stress resistance genes. Trends Plant Sci. 2013, 18, 522–531. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, P.; Jena, A.K.; Deuri, R.; Singh, S.P.; Sarmah, S. Review on Molecular Epidemiology in Relation to Devastating Late Blight Pathogen, P. infestans, de Bary. Int. J. Curr. Microbiol. App. Sci 2018, 7, 4651–4685. [Google Scholar] [CrossRef]
- Naeem, M.; Demirel, U.; Yousaf, M.F.; Caliskan, S.; Caliskan, M.E.; Wehling, P. Overview on domestication, breeding, genetic gain and improvement of tuber quality traits of potato using fast forwarding technique (GWAS): A review. Plant Breed. 2021, 140, 519–542. [Google Scholar] [CrossRef]
- Prodhomme, C.; Vos, P.G.; Paulo, M.J.; Tammes, J.E.; Visser, R.G.F.; Vossen, J.H.; van Eck, H.J. Distribution of P1(D1) wart disease resistance in potato germplasm and GWAS identification of haplotype-specific SNP markers. Theor. Appl. Genet. 2020, 133, 1859–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.K.; Siddappa, S.; Singh, B.P.; Kaushik, S.K.; Chakrabarti, S.K.; Bhardwaj, V.; Chandel, P. Molecular markers for late blight resistance breeding of potato: An update. Plant Breed. 2013, 132, 237–245. [Google Scholar] [CrossRef]
- Jupe, F.; Witek, K.; Verweij, W.; Śliwka, J.; Pritchard, L.; Etherington, G.J.; Maclean, D.; Cock, P.J.; Leggett, R.M.; Bryan, G.J.; et al. Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 2013, 76, 530–544. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement? Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef]
- Sakurai, N. Recent applications of metabolomics in plant breeding. Breed. Sci. 2022, 72, 21065. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Gupta, P.; Priscilla, K.; Sharankumar; Hangargi, B.; Veershetty, A.; Ramrao, D.P.; Suresh, S.; Narasanna, R.; Naik, G.R.; et al. Metabolomics Intervention Towards Better Understanding of Plant Traits. Cells 2021, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Price, E.J.; Drapal, M.; Perez-Fons, L.; Amah, D.; Bhattacharjee, R.; Heider, B.; Rouard, M.; Swennen, R.; Becerra Lopez-Lavalle, L.A.; Fraser, P.D. Metabolite database for root, tuber, and banana crops to facilitate modern breeding in understudied crops. Plant J. 2020, 101, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Nie, T.; Wang, D.; Chen, Q. Anthocyanin regulatory networks in Solanum tuberosum L. leaves elucidated via integrated metabolomics, transcriptomics, and StAN1 overexpression. BMC Plant Biol. 2022, 22, 228. [Google Scholar] [CrossRef]
- Dufková, H.; Berka, M.; Greplová, M.; Shejbalová, Š.; Hampejsová, R.; Luklová, M.; Domkářová, J.; Novák, J.; Kopačka, V.; Brzobohatý, B.; et al. The Omics Hunt for Novel Molecular Markers of Resistance to Phytophthora infestans. Plants 2022, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Hamooh, B.T.; Sattar, F.A.; Wellman, G.; Mousa, M.A.A. Metabolomic and Biochemical Analysis of Two Potato (Solanum tuberosum L.) Cultivars Exposed to In Vitro Osmotic and Salt Stresses. Plants 2021, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Holm, D.G.; Broeckling, C.D.; Prenni, J.E.; Heuberger, A.L. Metabolomics and Ionomics of Potato Tuber Reveals an Influence of Cultivar and Market Class on Human Nutrients and Bioactive Compounds. Front. Nutr. 2018, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef]
- Heretsch, P.; Giannis, A. The Veratrum and Solanum Alkaloids. Alkaloids Chem. Biol. 2015, 74, 201–232. [Google Scholar] [CrossRef]
- Chowański, S.; Adamski, Z.; Marciniak, P.; Rosiński, G.; Büyükgüzel, E.; Büyükgüzel, K.; Falabella, P.; Scrano, L.; Ventrella, E.; Lelario, F.; et al. A Review of Bioinsecticidal Activity of Solanaceae Alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Sarquís, J.I.; Coria, N.A.; Aguilar, I.; Rivera, A. Glycoalkaloid content in Solanum species and hybrids from a breeding program for resistance to late blight (Phytophthora infestans). Am. J. Potato Res. 2000, 77, 295–302. [Google Scholar] [CrossRef]
- Yogendra, K.N.; Pushpa, D.; Mosa, K.A.; Kushalappa, A.C.; Murphy, A.; Mosquera, T. Quantitative resistance in potato leaves to late blight associated with induced hydroxycinnamic acid amides. Funct. Integr. Genom. 2014, 14, 285–298. [Google Scholar] [CrossRef]
- Pushpa, D.; Yogendra, K.N.; Gunnaiah, R.; Kushalappa, A.C.; Murphy, A. Identification of Late Blight Resistance-Related Metabolites and Genes in Potato through Nontargeted Metabolomics. Plant Mol. Biol. Rep. 2014, 32, 584–595. [Google Scholar] [CrossRef]
- Andrivon, D.; Corbière, R.; Lucas, J.-M.; Pasco, C.; Gravoueille, J.-M.; Pellé, R.; Dantec, J.-P.; Ellissèche, D. Resistance to late blight and soft rot in six potato progenies and glycoalkaloid contents in the tubers. Am. J. Potato Res. 2003, 80, 125–134. [Google Scholar] [CrossRef]
- Deahl, K.L.; Young, R.J.; Sinden, S.L. A Study of the relationship of late blight resistance to glycoalkaloid content in fifteen potato clones. Am. Potato J. 1973, 50, 248–253. [Google Scholar] [CrossRef]
- Dahlin, P.; Müller, M.C.; Ekengren, S.; McKee, L.S.; Bulone, V. The Impact of Steroidal Glycoalkaloids on the Physiology of Phytophthora infestans, the Causative Agent of Potato Late Blight. Mol. Plant-Microbe Interact. 2017, 30, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Potato Glycoalkaloids and Metabolites: Roles in the Plant and in the Diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef] [PubMed]
- Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.; Nebbia, C.S.; Nielsen, E.; et al. Risk assessment of glycoalkaloids in feed and food, in particular in potatoes and potato-derived products. EFSA J. 2020, 18, e06222. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Xiong, Z.; Zou, W.; Shi, Z.; Li, S.; Zhang, X.; Liu, S.; Liu, Y.; Luo, X.; Ren, J.; et al. Anti-oomycete ability of scopolamine against Phytophthora infestans, a terrible pathogen of potato late blight. J. Sci. Food Agric. 2023. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Halim, V.A.; Eschen-Lippold, L.; Altmann, S.; Birschwilks, M.; Scheel, D.; Rosahl, S. Salicylic Acid Is Important for Basal Defense of Solanum tuberosum Against Phytophthora infestans. Mol. Plant-Microbe Interact. 2007, 20, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
- van Aubel, G.; Serderidis, S.; Ivens, J.; Clinckemaillie, A.; Legrève, A.; Hause, B.; Van Cutsem, P. Oligosaccharides successfully thwart hijacking of the salicylic acid pathway by Phytophthora infestans in potato leaves. Plant Pathol. 2018, 67, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Cesarino, I. Structural features and regulation of lignin deposited upon biotic and abiotic stresses. Curr. Opin. Biotechnol. 2019, 56, 209–214. [Google Scholar] [CrossRef]
- Abuley, I.K.; Pedersen, H.A.; Lekfeldt, J.D.S.; Fomsgaard, I.S.; Ravnskov, S. Metabolite profiling of Solanum tuberosum reveals a differential response to Phytophthora infestans dependent on host resistance and pathogen isolate. Plant Pathol. 2023, 72, 924–932. [Google Scholar] [CrossRef]
- Kröner, A.; Marnet, N.; Andrivon, D.; Val, F. Nicotiflorin, rutin and chlorogenic acid: Phenylpropanoids involved differently in quantitative resistance of potato tubers to biotrophic and necrotrophic pathogens. Plant Physiol. Biochem. 2012, 57, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Kuć, J.A. Metabolites accumulating in potato tubers following infection and stress. Teratology 1973, 8, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Yogendra, K.N.; Kumar, A.; Sarkar, K.; Li, Y.; Pushpa, D.; Mosa, K.A.; Duggavathi, R.; Kushalappa, A.C. Transcription factor StWRKY1 regulates phenylpropanoid metabolites conferring late blight resistance in potato. J. Exp. Bot. 2015, 66, 7377–7389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, S.; Ikeda, S.; Tsuda, S.; Someya, N.; Asano, K.; Kikuchi, J.; Chikayama, E.; Ono, H.; Sekiyama, Y. A survey of metabolic changes in potato leaves by NMR-based metabolic profiling in relation to resistance to late blight disease under field conditions. Magn. Reson. Chem. 2017, 55, 120–127. [Google Scholar] [CrossRef]
- Henriquez, M.A.; Adam, L.R.; Daayf, F. Alteration of secondary metabolites’ profiles in potato leaves in response to weakly and highly aggressive isolates of Phytophthora infestans. Plant Physiol. Biochem. 2012, 57, 8–14. [Google Scholar] [CrossRef]
- Gorzolka, K.; Perino, E.H.B.; Lederer, S.; Smolka, U.; Rosahl, S. Lysophosphatidylcholine 17:1 from the leaf surface of the wild potato species Solanum bulbocastanum inhibits Phytophthora infestans. J. Agric. Food Chem. 2021, 69, 5607–5617. [Google Scholar] [CrossRef]
- Hegde, N.; Joshi, S.; Soni, N.; Kushalappa, A.C. The caffeoyl-CoA O-methyltransferase gene SNP replacement in Russet Burbank potato variety enhances late blight resistance through cell wall reinforcement. Plant Cell Rep. 2020, 40, 237–254. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Yue, Y.; Amanullah, S.; Jahangir, M.M.; Fan, Y. Volatile terpenoids: Multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 2017, 246, 803–816. [Google Scholar] [CrossRef]
- Henriquez, M.A.; Soliman, A.; Li, G.; Hannoufa, A.; Ayele, B.T.; Daayf, F. Molecular cloning, functional characterization and expression of potato (Solanum tuberosum) 1-deoxy-d-xylulose 5-phosphate synthase 1 (StDXS1) in response to Phytophthora infestans. Plant Sci. 2016, 243, 71–83. [Google Scholar] [CrossRef]
- Toljamo, A.; Koistinen, V.; Hanhineva, K.; Kärenlampi, S.; Kokko, H. Terpenoid and lipid profiles vary in different Phytophthora cactorum—Strawberry interactions. Phytochemistry 2021, 189, 112820. [Google Scholar] [CrossRef]
- Mansfeld, B.N.; Colle, M.; Kang, Y.; Jones, A.D.; Grumet, R. Transcriptomic and metabolomic analyses of cucumber fruit peels reveal a developmental increase in terpenoid glycosides associated with age-related resistance to Phytophthora capsici. Hortic. Res. 2017, 4, 17022. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.E.; Dennis, C. Antifungal activity of post-infectional metabolites from potato tubers. Physiol. Plant Pathol. 1976, 9, 155–165. [Google Scholar] [CrossRef]
- Takemoto, D.; Shibata, Y.; Ojika, M.; Mizuno, Y.; Imano, S.; Ohtsu, M.; Sato, I.; Chiba, S.; Kawakita, K.; Rin, S.; et al. Resistance to Phytophthora infestans: Exploring genes required for disease resistance in Solanaceae plants. J. Gen. Plant Pathol. 2018, 84, 312–320. [Google Scholar] [CrossRef]
- Camagna, M.; Ojika, M.; Takemoto, D. Detoxification of the solanaceous phytoalexins rishitin, lubimin, oxylubimin and solavetivone via a cytochrome P450 oxygenase. Plant Signal. Behav. 2020, 15, 1707348. [Google Scholar] [CrossRef]
- Baur, S.; Bellé, N.; Frank, O.; Wurzer, S.; Pieczonka, S.A.; Fromme, T.; Stam, R.; Hausladen, H.; Hofmann, T.; Hückelhoven, R.; et al. Steroidal Saponins─New Sources to Develop Potato (Solanum tuberosum L.) Genotypes Resistant against Certain Phytophthora infestans Strains. J. Agric. Food Chem. 2022, 70, 7447–7459. [Google Scholar] [CrossRef]
- Takahashi, Y. The Role of Polyamines in Plant Disease Resistance. Environ. Control Biol. 2016, 54, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Chacón, A.L.; Camperos-Reyes, J.E.; Diazgranados, R.A.Á.; Romero, H.M. Biochemical and physiological responses of oil palm to bud rot caused by Phytophthora palmivora. Plant Physiol. Biochem. 2013, 70, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Hamzehzarghani, H.; Vikram, A.; Abu-Nada, Y.; Kushalappa, A.C. Tuber metabolic profiling of resistant and susceptible potato varieties challenged with Phytophthora infestans. Eur. J. Plant Pathol. 2015, 145, 277–287. [Google Scholar] [CrossRef]
- Berková, V.; Berka, M.; Griga, M.; Kopecká, R.; Prokopová, M.; Luklová, M.; Horáček, J.; Smýkalová, I.; Čičmanec, P.; Novák, J.; et al. Molecular Mechanisms Underlying Flax (Linum usitatissimum L.) Tolerance to Cadmium: A Case Study of Proteome and Metabolome of Four Different Flax Genotypes. Plants 2022, 11, 2931. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Kuklová, A.; Hoehenwarter, W.; Fragner, L.; Novák, O.; Rotková, G.; Jedelský, P.L.; Žáková, K.; Šmehilová, M.; Strnad, M.; et al. Proteome and metabolome profiling of cytokinin action in Arabidopsis identifying both distinct and similar responses to cytokinin down- and up-regulation. J. Exp. Bot. 2013, 64, 4193–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlin, L.; Baroukh, C.; Genin, S. Polyamines: Double agents in disease and plant immunity. Trends Plant Sci. 2021, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.-H.; Singhal, R.; Kachroo, A.; Kachroo, P. Fatty Acid-and Lipid-Mediated Signaling in Plant Defense Review View project Systemic Acquired Resistance View project. Artic. Annu. Rev. Phytopathol. 2017, 55, 505–536. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Kliebenstein, D.J.; Bostock, R.M.; Dehesh, K. Fatty acids and early detection of pathogens. Curr. Opin. Plant Biol. 2013, 16, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: The molecular point of view. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Saito, K. Roles of lipids as signaling molecules and mitigators during stress response in plants. Plant J. 2014, 79, 584–596. [Google Scholar] [CrossRef]
- Weber, H.; Chételat, A.; Caldelari, D.; Farmer, E.E. Divinyl Ether Fatty Acid Synthesis in Late Blight–Diseased Potato Leaves. Plant Cell 1999, 11, 485–493. [Google Scholar] [CrossRef]
- Fauconnier, M.-L.; Rojas-Beltran, J.; Dupuis, B.; Delaplace, P.; Frettinger, P.; Gosset, V.; du Jardin, P. Changes in oxylipin synthesis after Phytophthora infestans infection of potato leaves do not correlate with resistance. Plant Physiol. Biochem. 2008, 46, 823–831. [Google Scholar] [CrossRef]
- Sumayo, M.S.; Kwon, D.K.; Ghim, S.Y. Linoleic acid-induced expression of defense genes and enzymes in tobacco. J. Plant Physiol. 2014, 171, 1757–1762. [Google Scholar] [CrossRef]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological Control of Phytopathogenic Fungi by Fatty Acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef]
- Shimada, T.L.; Takano, Y.; Hara-Nishimura, I. Oil body-mediated defense against fungi: From tissues to ecology. Plant Signal. Behav. 2015, 10, e989036. [Google Scholar] [CrossRef] [Green Version]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green leaf volatile production by plants: A meta-analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Ameye, M.; Mänd, M.; Audenaert, K.; Landschoot, S.; Haesaert, G. Green Leaf Volatile Confers Management of Late Blight Disease: A Green Vaccination in Potato. J. Fungi 2021, 7, 312. [Google Scholar] [CrossRef]
- Brus-Szkalej, M.; Andersen, C.B.; Vetukuri, R.R.; Grenville-Briggs, L.J. A family of cell wall transglutaminases is essential for appressorium development and pathogenicity in Phytophthora infestans. bioRxiv 2021, preprint. [Google Scholar] [CrossRef]
- Fraser-Pitt, D.J.; Mercer, D.K.; Smith, D.; Kowalczuk, A.; Robertson, J.; Lovie, E.; Perenyi, P.; Cole, M.; Doumith, M.; Hill, R.L.R.; et al. Cysteamine, an Endogenous Aminothiol, and Cystamine, the Disulfide Product of Oxidation, Increase Pseudomonas aeruginosa Sensitivity to Reactive Oxygen and Nitrogen Species and Potentiate Therapeutic Antibiotics against Bacterial Infection. Infect. Immun. 2018, 86, e00947-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronek, P.; Thielert, M.; Voytik, E.; Tanzer, M.C.; Hansen, F.M.; Willems, S.; Karayel, O.; Brunner, A.-D.; Meier, F.; Mann, M. Rapid and In-Depth Coverage of the (Phospho-)Proteome with Deep Libraries and Optimal Window Design for dia-PASEF. Mol. Cell. Proteom. 2022, 21, 100279. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Polle, A. What the transcriptome does not tell—Proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Alexandersson, E.; Sandin, M.; Resjö, S.; Lenman, M.; Hedley, P.; Levander, F.; Andreasson, E. Quantitative proteomics and transcriptomics of potato in response to Phytophthora infestans in compatible and incompatible interactions. BMC Genom. 2014, 15, 497. [Google Scholar] [CrossRef] [Green Version]
- Chawade, A.; Alexandersson, E.; Bengtsson, T.; Andreasson, E.; Levander, F. Targeted Proteomics Approach for Precision Plant Breeding. J. Proteome Res. 2016, 15, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.L.; Andreu, A.B.; Korgan, S.; Lobato, M.C.; Huarte, M.; Walling, L.L.; Daleo, G.R. PLPKI: A novel serine protease inhibitor as a potential biochemical marker involved in horizontal resistance to Phytophthora infestans. Plant Breed. 2014, 133, 275–280. [Google Scholar] [CrossRef]
- Sharma, N.; Gruszewski, H.A.; Park, S.W.; Holm, D.G.; Vivanco, J.M. Purification of an isoform of patatin with antimicrobial activity against Phytophthora infestans. Plant Physiol. Biochem. 2004, 42, 647–655. [Google Scholar] [CrossRef]
- Tian, Z.; He, Q.; Wang, H.; Liu, Y.; Zhang, Y.; Shao, F.; Xie, C. The Potato ERF Transcription Factor StERF3 Negatively Regulates Resistance to Phytophthora infestans and Salt Tolerance in Potato. Plant Cell Physiol. 2015, 56, 992–1005. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Huang, M.; Gao, J.; Wang, Z.; Zhang, D.; Zhang, Y.; Yan, L.; Yu, X.; Li, B.; Shen, Y. Comparative proteomics of three Chinese potato cultivars to improve understanding of potato molecular response to late blight disease. BMC Genom. 2020, 21, 880. [Google Scholar] [CrossRef]
- Resjö, S.; Zahid, M.A.; Burra, D.D.; Lenman, M.; Levander, F.; Andreasson, E. Proteomics of PTI and Two ETI Immune Reactions in Potato Leaves. Int. J. Mol. Sci. 2019, 20, 4726. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Gao, J.; Zhang, Y.; Wang, Z.; Zhang, D.; Chen, Q.; Ye, X.; Xu, Y.; Yang, G.; Yan, L.; et al. Quantitative Proteomics of Potato Leaves Infected with Phytophthora infestans Provides Insights into Coordinated and Altered Protein Expression during Early and Late Disease Stages. Int. J. Mol. Sci. 2019, 20, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burra, D.D.; Lenman, M.; Levander, F.; Resjö, S.; Andreasson, E. Comparative Membrane-Associated Proteomics of Three Different Immune Reactions in Potato. Int. J. Mol. Sci. 2018, 19, 538. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.B.; Pagano, M.R.; Daleo, G.R.; Guevara, M.G. Hydrophobic proteins secreted into the apoplast may contribute to resistance against Phytophthora infestans in potato. Plant Physiol. Biochem. 2012, 60, 59–66. [Google Scholar] [CrossRef]
- Bártová, V.; Bárta, J.; Jarošová, M. Antifungal and antimicrobial proteins and peptides of potato (Solanum tuberosum L.) tubers and their applications. Appl. Microbiol. Biotechnol. 2019, 103, 5533–5547. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Zhang, Z.; Kang, L.; Li, M.; Zhang, J.; Feng, Y.; Yin, J.; Gong, X.; Zhao, J. Small G Protein StRab5b positively regulates potato resistance to Phytophthora infestans. Front. Plant Sci. 2022, 13, 1065627. [Google Scholar] [CrossRef] [PubMed]
- Colignon, B.; Dieu, M.; Demazy, C.; Delaive, E.; Muhovski, Y.; Raes, M.; Mauro, S. Proteomic Study of SUMOylation During Solanum tuberosum–Phytophthora infestans Interactions. Mol. Plant-Microbe Interact. 2017, 30, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.; He, Q.; Song, J.; Liu, Z.; Yu, J.; Hu, K.; Liu, H.; Mu, Y.; Wu, J.; Tian, Z.; et al. A Phytophthora infestans RXLR effector AVR8 suppresses plant immunity by targeting a desumoylating isopeptidase DeSI2. Plant J. 2023. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.L.; De Souza, C.M.; De Oliveira, K.B.S.; Dias, S.C.; Franco, O.L. The role of antimicrobial peptides in plant immunity. J. Exp. Bot. 2018, 69, 4997–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverstein, K.A.T.; Moskal, W.A.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.K.; Mattoo, A.K. Multitasking antimicrobial peptides in plant development and host defense against biotic/abiotic stress. Plant Sci. 2014, 228, 135–149. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef]
- Oliveira-Lima, M.; Benko-Iseppon, A.; Neto, J.; Rodriguez-Decuadro, S.; Kido, E.; Crovella, S.; Pandolfi, V. Snakin: Structure, Roles and Applications of a Plant Antimicrobial Peptide. Curr. Protein Pept. Sci. 2017, 18, 368–374. [Google Scholar] [CrossRef]
- Almasia, N.I.; Nahirñak, V.; Hopp, H.E.; Vazquez-Rovere, C. Potato Snakin-1: An antimicrobial player of the trade-off between host defense and development. Plant Cell Rep. 2020, 39, 839–849. [Google Scholar] [CrossRef]
- Beliaev, D.V.; Yuorieva, N.O.; Tereshonok, D.V.; Tashlieva, I.I.; Derevyagina, M.K.; Meleshin, A.A.; Rogozhin, E.A.; Kozlov, S.A. High Resistance of Potato to Early Blight Is Achieved by Expression of the Pro-SmAMP1 Gene for Hevein-Like Antimicrobial Peptides from Common Chickweed (Stellaria media). Plants 2021, 10, 1395. [Google Scholar] [CrossRef]
- Colombo, M.; Masiero, S.; Rosa, S.; Caporali, E.; Toffolatti, S.L.; Mizzotti, C.; Tadini, L.; Rossi, F.; Pellegrino, S.; Musetti, R.; et al. NoPv1: A synthetic antimicrobial peptide aptamer targeting the causal agents of grapevine downy mildew and potato late blight. Sci. Rep. 2020, 10, 17574. [Google Scholar] [CrossRef]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef] [PubMed]
- Das, P.R.; Sherif, S.M. Application of Exogenous dsRNAs-induced RNAi in Agriculture: Challenges and Triumphs. Front. Plant Sci. 2020, 11, 946. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.; Ahmad, N.; Khalifa, M.A.S.; Du, Y.; Mandozai, A.; Khattak, A.N.; Piwu, W. Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean. Genes 2023, 14, 369. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Examples of Biocontrol Agents | Organism | Effect | References |
|---|---|---|---|
| Extracts | Xanthium strumarium, Lauris nobilis, Salvia officinalis, Styrax officinalis | Mycelial growth inhibition | [73] |
| Solanum habrochaites | Mycelial growth inhibition, reduced disease progression | [75] | |
| Willaertia magna C2c Maky | Disease reduction | [78] | |
| Trichoderma virens | reduced disease progression | [72] | |
| Bacteria | Pseudomonas strains isolated from the rhizosphere and shoots of potato | Reduced disease progression | [79] |
| Bacillus subtilis | Reduced disease progression | [80] | |
| Myxococcus fulvus | Reduced disease progression | [81] | |
| Bacillus velezensis | Mycelial growth inhibition, improved resistance | [77,82] | |
| Fungi | Fungal endophytes isolated from Solanum spp. | Mycelial growth inhibition | [69] |
| Fusarium oxysporum | Induction of systemic resistance | [83] | |
| Fungal endophytes isolated from Espeletia spp. | Mycelial growth inhibition | [68] | |
| Endophytes Phoma eupatorii | Mycelial growth inhibition, infection prevention | [70] |
| Class | Name | HMDB |
|---|---|---|
| Steroidal saponins | α-Solanine | HMDB0034202 |
| α-Chaconine | HMDB0039353 | |
| Acridone alkaloids | Melicopicine | NA |
| Solanidines | Solanidine | HMDB0003236 |
| Tropane alkaloid | Scopolamine | HMDB0003573 |
| Class | Name | HMDB |
|---|---|---|
| Coumarins | 4-Coumaryl alcohol | HMDB0003654 |
| Scopolin | HMDB0303366 | |
| Scopoletin | HMDB0034344 | |
| Catechols | Paucine | HMDB0029876 |
| Flavonoids | Rutin | HMDB0003249 |
| Hydroxycinnamic acids and derivatives | 1-O-Feruloyl-β-D-glucose | HMDB0302219 |
| 1-O-Sinapoyl-β-D-glucose | HMDB0302379 | |
| Caffeic acid 3-glucoside | HMDB0303040 | |
| Ferulic acid | HMDB0000954 | |
| N-cis-Feruloyltyramine | HMDB0036381 | |
| Subaphylline | HMDB0033463 | |
| Quinic acids and derivatives | 5-O-Feruloylquinic acid | HMDB0240478 |
| p-Coumaroyl quinic acid | HMDB0301709 | |
| Chlorogenic acid | HMDB0003164 | |
| Quinic acid | HMDB0003072 |
| Class | Name | HMDB |
|---|---|---|
| Sesquiterpenoids | Phytuberin | HMDB0035754 |
| Rishitin | HMDB0035593 | |
| Lubimin | NA | |
| Solavetivone | HMDB0035657 | |
| Saponins | Neoindioside D | NA |
| Protoneodioscin | ||
| Barogenin-solatrioside | ||
| Barogenin-chacotrioside |
| Class | Name | HMDB/LM ID |
|---|---|---|
| Glycerophospholipid | Heptadecenoyl-lysophosphatidylcholine (LPC 17:1) | LMGP01050002 |
| Acetate ester | Z-3-Hexenyl acetate | HMDB0040215 |
| Dialkyldisulfide | Cystamine | HMDB0250701 |
| Alkylthiol | Cysteamine | HMDB0002991 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufková, H.; Greplová, M.; Hampejsová, R.; Kuzmenko, M.; Hausvater, E.; Brzobohatý, B.; Černý, M. Secondary Metabolites, Other Prospective Substances, and Alternative Approaches That Could Promote Resistance against Phytophthora infestans. Agronomy 2023, 13, 1822. https://doi.org/10.3390/agronomy13071822
Dufková H, Greplová M, Hampejsová R, Kuzmenko M, Hausvater E, Brzobohatý B, Černý M. Secondary Metabolites, Other Prospective Substances, and Alternative Approaches That Could Promote Resistance against Phytophthora infestans. Agronomy. 2023; 13(7):1822. https://doi.org/10.3390/agronomy13071822
Chicago/Turabian StyleDufková, Hana, Marie Greplová, Romana Hampejsová, Marharyta Kuzmenko, Ervín Hausvater, Břetislav Brzobohatý, and Martin Černý. 2023. "Secondary Metabolites, Other Prospective Substances, and Alternative Approaches That Could Promote Resistance against Phytophthora infestans" Agronomy 13, no. 7: 1822. https://doi.org/10.3390/agronomy13071822
APA StyleDufková, H., Greplová, M., Hampejsová, R., Kuzmenko, M., Hausvater, E., Brzobohatý, B., & Černý, M. (2023). Secondary Metabolites, Other Prospective Substances, and Alternative Approaches That Could Promote Resistance against Phytophthora infestans. Agronomy, 13(7), 1822. https://doi.org/10.3390/agronomy13071822