1. Introduction
To meet the demands of the expanding population, wheat cultivation requires the rigorous use of agricultural inputs, mainly nitrogen (N) fertilizers, to ensure satisfactory yields and superior grain quality. Although N fertilizer is indispensable for global food security, its depletion through various processes can lead to catastrophic environmental consequences such as soil acidification, nutrient runoff, eutrophication, reduced biological diversity, and greenhouse gas emissions [
1]. The prevailing notion is that farm input, particularly inorganic N sources, has been introduced to assist farmers in enhancing crop productivity. However, the scarcity of knowledge and technological proficiency among farmers has resulted in an unwarranted application of these inputs, leading to unfavorable consequences [
2,
3]. An undeniable truth is that the use of high amounts of N fertilizers during crop growing periods is heavily dependent on copious amounts of nonrenewable energy sources [
4]. According to estimates from UN-FAO (United Nations Food and Agriculture Organization), approximately 17% of global nitrogen fertilizer consumption is used in wheat production processes [
5]. Utilization of such a massive amount unquestionably results in possible losses of a noteworthy quantity of applied N, coupled with extensive energy consumption during production processes and application in the field. Applying N fertilizer to meet plant demands in the absence or insufficiency thereof is a challenging task to accomplish. Undoubtedly, both of these parameters have the potential to inflict a negative effect on the quality and quantity of crop yields. Considering the persistent challenge of achieving efficient use of N in agriculture, the identification of genotypes that exhibit a high, medium, or low N response remains a promising approach. This strategy has the potential to improve the NUE of crops, which is currently below 50% [
6]. However, more research is needed on the identification of genotypes that can thrive at specific N rates. A recent investigation has revealed that the latest cultivars have advanced yield output coupled with improved efficiency in nutrient utilization [
7], indicating that it is expected that the efficiency of elevated nutrient use is augmented indirectly through the preference for high yield.
One crucial requirement for the identification, selection, and development of N-efficient germplasm is the availability of genetic diversity. Previous studies on wheat cultivars with N inputs reported that the genetic variation for NUE and its component traits was available [
8,
9,
10,
11]. Previous investigations have also shown that the use of N inputs in different seasons and years can result in diverse responses in yields and NUE among different wheat cultivars [
12]. Thus, it is crucial to investigate genotypes with different N rates in particular soil and environmental conditions that enable farmers to enhance crop yields, curtail production expenses, and ensure sustainable agricultural practices. Looking at this point of view, the innovation and nurturing of N-efficient wheat cultivars can curtail the N utilized as input without compromising the grain output. The NUE of wheat has been estimated to be <60% [
13,
14,
15], suggesting that nitrogen is not optimally used for grain production. Several strategies have been employed to increase the nitrogen use efficiency in cereal crops, viz., the use of slow-release N fertilizers, nitrification inhibitors, modified urea materials, including neem-coated urea, precise nitrogen management, leaf color charts, method, rate and time of N application, balanced fertilization [
16], etc. In addition, the identification of nitrogen-efficient cultivars is yet another promising strategy to increase the NUE.
Depending on the cultivars grown, the N input of N
80 (80 kg ha
−1) to N
120 (120 kg ha
−1) can result in a range of 28.8 to 40 kg of wheat grains with an NUE of 1 kg of grain per kg of N applied [
17]. The accumulation of phytomass and the concentration of chlorophyll in leaves are intrinsically linked to the efficiency of N utilization, which enables them to serve as indirect selection markers for cultivars that exhibit a high degree of efficiency in nitrogen utilization. Thus, the judicious application of an optimal dosage of N fertilizer, tailored to the specific cultivar, can be perceived as the main strategy for maximizing economic yields, improving N uptake, and improving the effectiveness of N utilization in wheat. Despite advances in breeding techniques, there are instances where superior cultivars are developed without considering their ability to sustain growth and productivity on soils with low fertility. These cultivars were specifically chosen for their ability to yield more under high-input fertilizer regimes. Therefore, to satisfy the growing global food demands of populations, it is crucial to constantly assess N-efficient varieties and choose breeds based on their genetic outputs. The objectives of this study were as follows: (i) to evaluate the growth, yield components, and N use efficiencies of wheat cultivars under control versus half and recommended N supplies; (ii) to investigate the differences between wheat cultivars in terms of economic yield and harvest index; and (iii) to screen the wheat cultivars based on grain yield efficiency index.
4. Discussion
Our scientific investigation aimed to assess the performance of wheat cultivars under different levels of nitrogen (N) inputs in the North-West region of India. The objective was to enhance both the economic yield and quality of the crops while also minimizing nitrogen losses to the environment. Nitrogen management strategies included the timing of application of N at the appropriate stages (three splits, namely the basal, CRI, and tillering stages) [
26]. The comprehensive findings indicated that among all cultivars, ‘HD 3249’ and ‘HD 3117’ showed superior performance with N
150-split treatments, resulting in higher GY and its components. Across N inputs, N
150 resulted in the highest seed/spike, dry matter accumulation, and grain yield (GY) (
Table 2,
Table 4 and
Table 6). However, plant traits responses to heterogeneous N inputs can vary depending on soil properties, crop genotypic constitution, agronomic management methodologies, and methodology circumstances throughout the entire course of the growth season [
27]. Yano et al. [
28] reported nonsignificant variations in GY regarding N sources and application timings. However, notable disparities in GY were observed when different N inputs were used. Our result aligns with the prevailing results that show a positive correlation between GY and N inputs [
14]. A notable improvement in GY was observed up to the N input of N
150 in certain cultivars. The N
0 application did not show a significant increase in GY; however, a substantial increase in GY was observed when comparing the application rates of N
75 and N
150. Numerous studies have proven that the optimal N rate for wheat cultivation in North-West India can vary between 50 and 185 kg of N ha
–1 to attain the desired GY [
29]. Cultivar GY outcomes were dissimilar, likely due to the unique genetic potential of each cultivar, soil characteristics, and weather conditions prevalent during the conductance of field trials [
30,
31]. Some previous studies have shown that selecting phenotypes adapted to specific environmental conditions and considering early generation selection of partitioning traits, such as harvest index (HI) and GY, can lead to high HI and GY in wheat [
32].
Fuertes-Mendizabal et al. [
33] suggest that the amalgamation of an intensified N rate with delayed N application could induce the expansion of the crop canopy, increase photosynthetic efficiency, increase carbohydrate accumulation during the vegetative stages, and ultimately result in a boost in GY. Our study produced compelling results, suggesting that cultivars ‘HD 3249’ and ‘HD 3117’ exhibited notable improvements in yield when N
150 was distributed between the split treatments of 75–37.5–37.5 kg N ha
−1. These observations signify the remarkable potential of wheat for yield production. In particular, the application of N
150 resulted in a higher number of spikes per square meter, SY, biological yield (BY), harvest index (HI), dry matter accumulation (DMA), and tiller production in most genotypes (
Table 2,
Table 4 and
Table 6). However, the response varied in each wheat cultivar, as the capacity for N uptake is generally low during the initial growth season and rapidly increases during the vegetative growth phase. Based on studies conducted by Zhang et al. [
34], the N input application at three times and two different times resulted in a 15% and 7% increase in GY compared to a single-dose application. Similarly, Singh et al. [
35] reported that the increase in GY was feasible by applying 50% to 75% of N inputs at the sowing stage in loam soil, with urea spread/drilled in rows. Despite variations in N inputs and application timing, the utilization of N
150 leads to a superior yield compared to the N rates of N
75 and N
0. The rationale behind this can be that NCU/NOCU is a slow-release type and undergoes gradual conversion to ammonia in soil, making it plantable over an extended period compared to N inputs N
75 and N
0. Singh et al. [
36] reported that the use of NCU/NOCU in wheat shows an approximate 5–6% increase in GY as compared to urea at the same levels. However, it is essential to acknowledge that the response of GY to N inputs may be influenced by several factors, such as climatic conditions, soil properties, and the specific cultivar of the crop.
Yang and Baker [
37] suggested that the variability in GY between cultivars across different environments can be due to the expression of diverse gene sets in those different environments or to discrepancies in the reactions of the same gene set to varying environmental conditions. Furthermore, the variations of yield and yield traits may be due to the genetic composition of the cultivars influencing the grain filling and transfer of biochemical assimilates from source to sink. ElMetwally et al. [
38], EI-Habbal et al. [
39], and Hassan et al. [
40] reported on varietal disparities in yield attributes of wheat cultivars. Elevating N levels to N
150 resulted in an improvement in yield and yield-contributing characteristics in wheat and was due to the influential function of N as a vital element of chlorophyll on the accumulation of dry matter. N inputs influence carbohydrates biosynthesis by modifying the leaf area, which is responsible for capturing solar energy and assimilating CO
2. Furthermore, they enhance the efficiency of the photosynthesis process. Previous studies by Fu et al. [
41] reported that GY and its components significantly increased with an optimal increase in N inputs.
The application of N inputs has significantly increased the agro-morphological performance measures of cultivars, such as tillers and chlorophyll content. Each cultivar exhibits distinct characteristics based on its unique genetic potential and traits. Certain efficient genotypes demonstrate reduced N losses and improved the growth and development processes. N inputs play a regulatory role in several plant hormones, such as auxin and cytokinin, as well as gene expression, thereby influencing tillering emergence and growth [
42,
43]. The accumulation of dry matter in wheat cultivars varies, which could be attributed to the genotypic diversity influencing the distribution of photoassimilates in the leaves, resulting in different plant growth patterns and assimilate allocation to reproductive organs [
44,
45]. Factors such as plant height, spike size, and hormones are likely responsible for the variations observed in tillering capacity among wheat cultivars [
46,
47]. The ‘HD 3249’ and ‘HD 3117’ genotypes exhibited the highest yield attributes, possibly due to a larger surface area of wheat roots, improved growth and development, and higher photosynthetic efficiency of the leaf area index during flowering and physiological maturity stages.
Nitrogen assumes a pivotal and irreplaceable role in the proliferation and maturation of plants, acting as a vital macronutrient that orchestrates their intricate physiological processes. The results of our investigation showed that the plant height of wheat cultivars was shortest under N input N
0 compared to N
75 and N
150 (
Table 1). Similar kinds of results were reported by Tanka et al. [
48] and Wolf et al. [
49]. However, the response to nitrogen input differs in each cultivar due to their unique genetic potential and characteristics [
50,
51]. Chlorophyll has a significant role as a photosynthetic pigment and an effective indicator of leaf functions amid the detrimental impact of various ecological stressors [
52]. In addition, it serves as a critical parameter for observing nitrogen uptake in wheat [
48]. Our results show that, compared to the untreated plants, there was an increase in total chlorophyll content as the N input increased (
Table 3). The elevated levels of N and chlorophyll in the leaves exhibited a positive correlation with the net photosynthetic rate, resulting in enhanced carbohydrate production. Consequently, this metabolic shift led to increased biomass accumulation in both the above-ground and below-ground plant components [
53,
54].
The N × genotypes interaction (NCI) shows a nonsignificant relationship with both grain protein content (GPC) and 1000-grain weight (G.W.). Although statistically non-significant, the measurements of GPC and G.W. displayed notable variation across the different wheat cultivars used in our experiments (
Table 5). The detailed results reveal that N
150 and N
75 increased the G.W. and GPC of wheat cultivars as compared to N
0. The variation in GPC and G.W. between genotype ‘HD 3249’ and ‘HD 3298’ can be attributed to the different genotypic characteristics of each cultivar. In fact, splitting and N inputs’ application timing have more significant effects on GPC [
14] and other quality traits of wheat [
51] than simply increasing N inputs. Fuertes-Mendizabal et al. [
33] reported that splitting of N rates enhanced quality parameters, while GY remained unaffected. During our N inputs experiment, we observed that increasing the N inputs results in a favorable effect on GPC and G.W. Wheat cultivar traits show divergent responses to N inputs, and this is due to variations in N availability, N release from slow-release fertilizers, genotype-specific genetic traits, and the influence of soil moisture and temperature levels [
55,
56]. Some previous studies reported that nitrate reductase (NR) activity was two times higher at N input N
105 compared to N
0 [
57]. The NR activity in the cultivar at N
150 might be a reason for higher GY and GPC. Balotf et al. [
58] and Lambeck et al. [
59] reported that NR activities in wheat increase as external nitrate is applied.
Plant N accumulation and N grain production efficiency (NUEg) at maturity are influenced by a variety of factors such as N inputs, cultivars, prevailing weather conditions, and yearly variations [
9,
60]. Our 2 years of study revealed that the total uptake of N (TUN) was increased by N inputs and ranged between 55.5 and 169.6 kg ha
−1 (
Table 8). According to a study by Singh et al. [
61], under irrigation conditions, the TUN and NUEg were between 49 and 160 kg ha
–1 and 35 and 79 kg kg
–1, respectively. In a 3-year pioneering study by Xue et al. [
62] in the south-eastern Chinese area of Taihu Lake, it was revealed that the TUN by wheat was 132 and 149 kg ha
–1, respectively, when N inputs of 180 and 240 kg ha
–1 were applied. Similarly, Jia et al. [
63] reported that increasing N inputs from 150 to 270 kg ha
–1 led to an increase in TUN in wheat cultivars, from 198 to 238 kg ha
–1. According to Wang et al. [
64], in wheat, N absorption originated from N fertilizer was increased two-fold when N input increased from 96 to 240 kg ha
–1.
Zhu et al. [
65] observed that at N input N
100, the NUEg ranged from 28.8 to 58.4 kg kg
–1. Wu et al. [
66] reported that superior cultivars show comparatively elevated TUN at the same N inputs. This suggests that a greater TUN could potentially lead to a higher NUE. We have noticed that ‘HD 3249’ and ‘HD 3117’ had a comparatively higher TUN than the other cultivars, leading to a higher GY (
Table 8). The data presented for the NHI and NUEg parameters displayed a diverse range of 67.4 to 77.3% and 35.3 to 44.3 kg kg
−1, respectively, across various N inputs (
Table 8). High N uptake in genotypes ‘HD 3249’ and ‘HD 3117’ was due to more developed root systems in these cultivars, which helped in exploring large soil volumes and making a better use of the resources [
67].
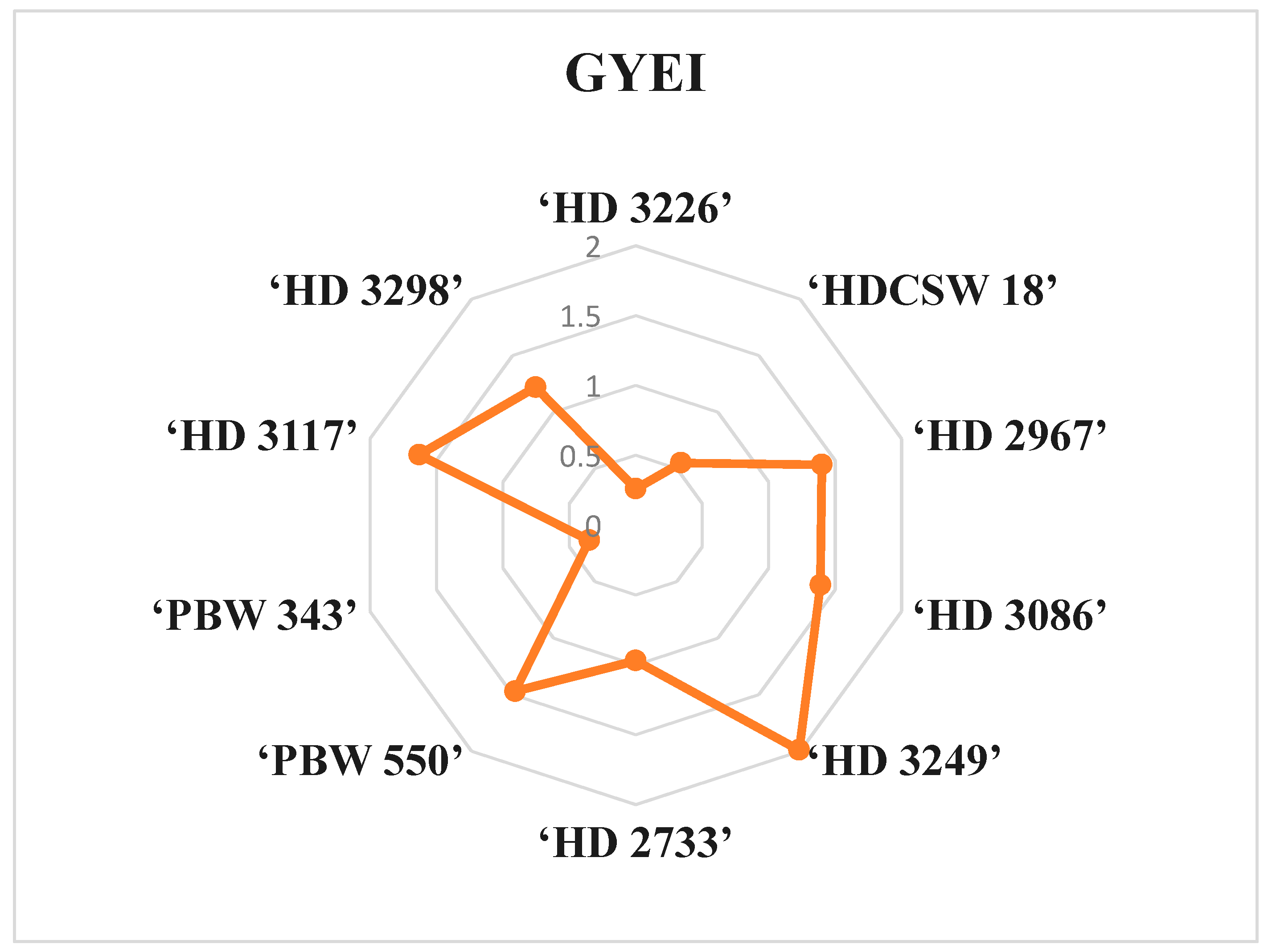
The grain yield efficiency index (GYEI) is a highly reliable indicator of GY and an effective method of evaluating genotypes in terms of their potential grain production [
68]. Among all wheat cultivars, only six cultivars (‘HD 3298’, ‘HD 3117’, ‘PBW 550’, ‘HD 3249’, ‘HD 3086’ and ‘HD 2967’) show a GYEI value of ≥1, indicating favorable responses to various N inputs (
Table 7 and
Figure 2). Our results suggest that GYEI varied among all wheat cultivars. There is an abundance of literature available on how various cultivars and species within the wheat family exhibit various nitrogen utilization mechanisms [
69,
70]. The diversity in wheat cultivars for NUE was due to multiple factors, including root morphology, root exudation patterns, uptake capacity, transport mechanisms, metabolic pathways, and the partitioning of assimilates. In addition, variations in physiological and biochemical traits such as photosynthetic efficiency, transpiration rates, nitrogen assimilation and remobilization, and sink strength also contribute to differences in NUE [
71,
72].
Pearson’s correlation analysis revealed that, except for some of the parameters, all traits of wheat cultivars were highly positively correlated with each other at
p < 0.01 (
Figure 3), [
66,
73,
74]. The HI shows a significant positive correlation with GY in our study [
75]. Our results were consistent with those of Peng et al. [
76], Ayadi et al. [
77], Kubar et al. [
78], Boulelouah et al. [
79], and Wang et al. [
80]. Multiple research studies have shown that it is possible to achieve high GY and NUE by focusing on the distribution of biomass into the grains rather than solely increasing its concentrations [
80]. Thus, understanding N metabolism in a specific genotype, and identifying wheat cultivars with greater NUE in conjunction with optimizing N input strategies may improve NUE and GY, and reduce the negative impacts of excess fertilizer application.
The N-use-efficient wheat cultivars accumulated more N in their grains and biomass, suggesting a higher uptake of N in comparison to the N-use-inefficient cultivars. Thus, it can be easily stipulated that the increased uptake of N by efficient N users reduces the losses of N. At a particular point in time, the quantity of nitrate-nitrogen available can be removed by the plants or lost to the environment. Thus, the efficient N-user wheat cultivars can have efficient N uptake, and very little N may be available for loss in comparison to the N-user-inefficient cultivars.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}