The Genetics, Genomics, and Breeding of Cereals and Grain Legumes: Traits and Technologies for Future Food Security



{kind=link}
{kind=link}
{kind=link}
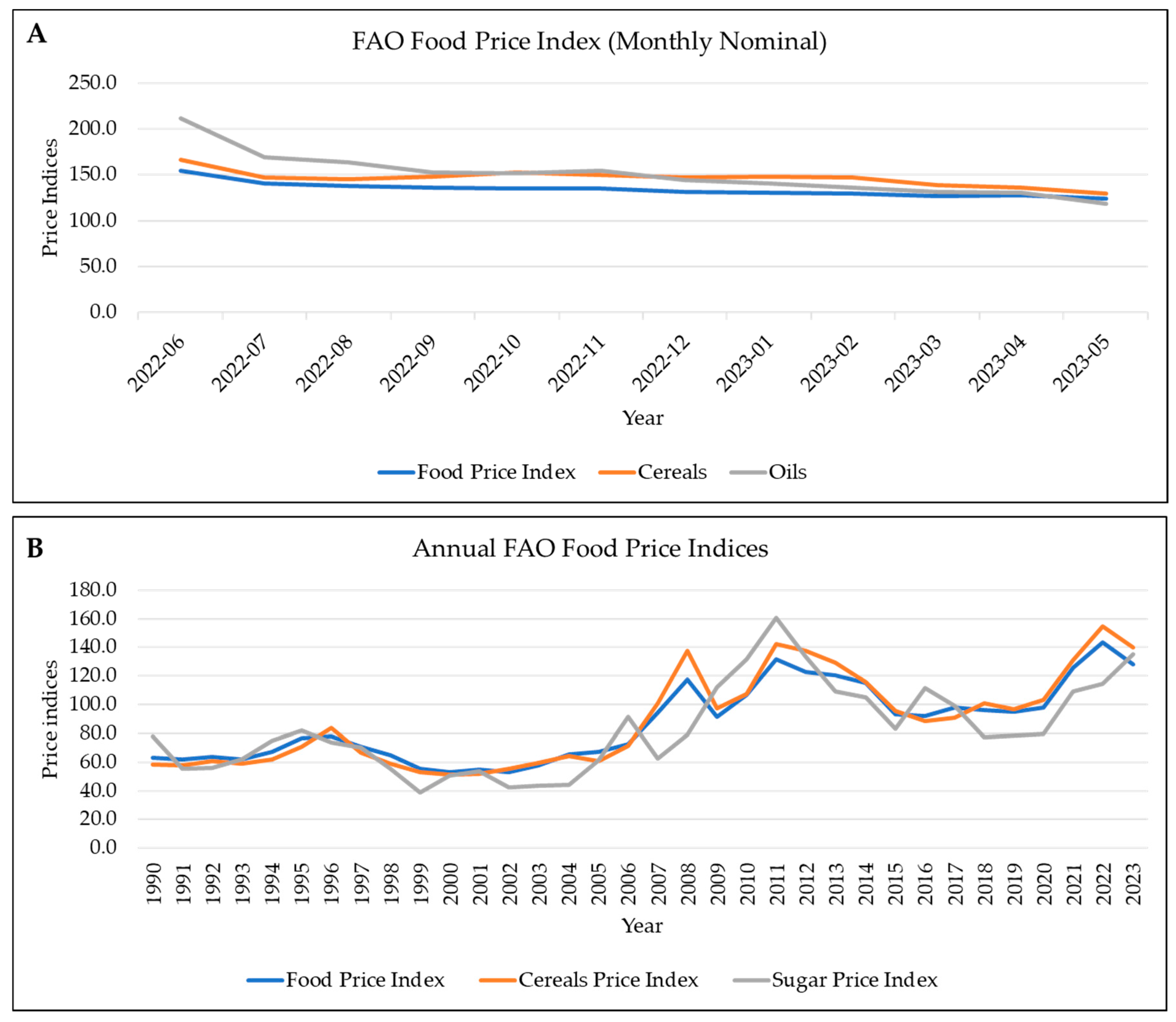
1. Global Population and Food Security
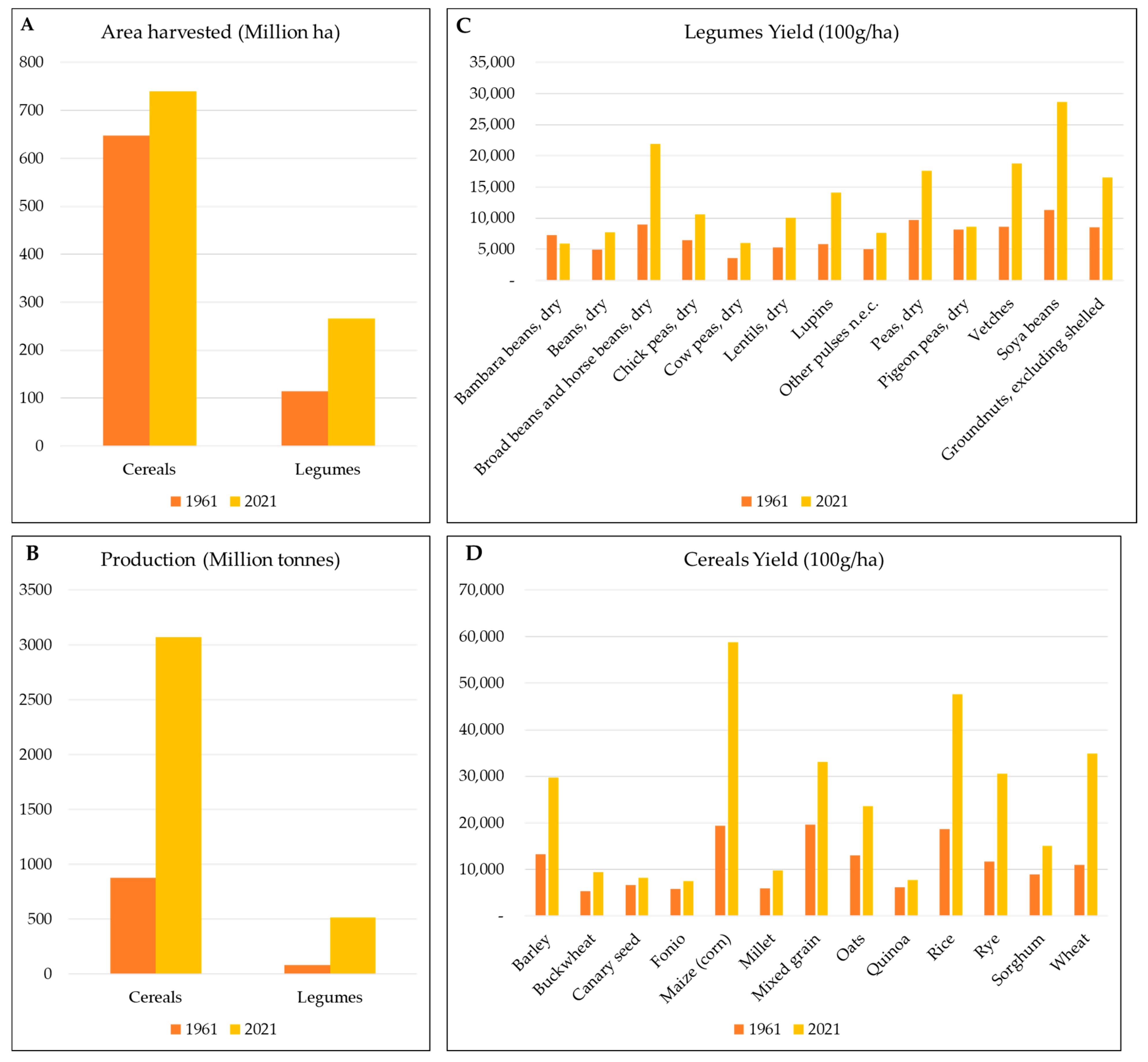
2. The Current Cereals and Legumes Situation
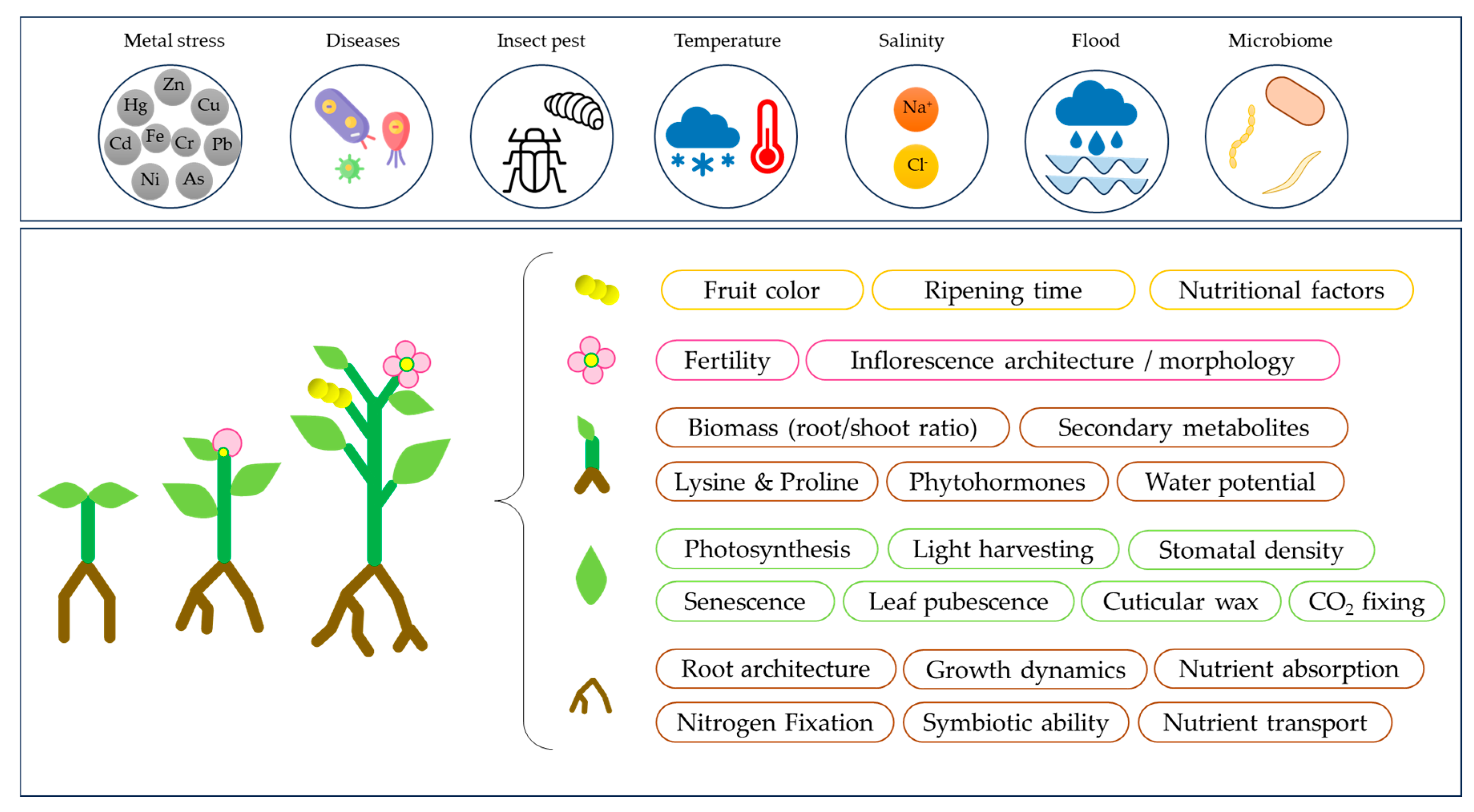
3. The Way Forward in the Genetics, Genomics, and Breeding of Cereals and Grain Legumes
4. Special Issue Overview
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. How to Feed the World in 2050: High-Level Expert Forum; FAO: Rome, Italy, 2009. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef] [PubMed]
- FAO. Crop Prospects and Food Situation—Quarterly Global Report No. 1; Food and Agriculture Organization: Rome, Italy, 2023; p. 46. [Google Scholar]
- Food and Agriculture Organization. The State of Food Security and Nutrition in the World—Repurposing Food and Agricultural Policies to Make Healthy Diets More Affordable; Food and Agriculture Organization of the United Nations: Rome, Italy; International Fund for Agricultural Development: Rome, Italy; United Nations Children’s Fund: New York, NY, USA; United Nations World Food Programme: Rome, Italy; World Health Organization: Rome, Italy, 2022; p. 260. [Google Scholar]
- Smýkal, P.; von Wettberg, E.J.; McPhee, K. Legume Genetics and Biology: From Mendel’s Pea to Legume Genomics. Int. J. Mol. Sci. 2020, 21, 3336. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Stuart, A.M.; Laborte, A.G.; Rattalino Edreira, J.I.; Dobermann, A.; Kien, L.V.N.; Thúy, L.T.; Paothong, K.; Traesang, P.; Tint, K.M. Southeast Asia must narrow down the yield gap to continue to be a major rice bowl. Nat. Food 2022, 3, 217–226. [Google Scholar] [CrossRef]
- Espe, M.B.; Cassman, K.G.; Yang, H.; Guilpart, N.; Grassini, P.; Van Wart, J.; Anders, M.; Beighley, D.; Harrell, D.; Linscombe, S. Yield gap analysis of US rice production systems shows opportunities for improvement. Field Crops Res. 2016, 196, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Senapati, N.; Semenov, M.A.; Halford, N.G.; Hawkesford, M.J.; Asseng, S.; Cooper, M.; Ewert, F.; van Ittersum, M.K.; Martre, P.; Olesen, J.E. Global wheat production could benefit from closing the genetic yield gap. Nat. Food 2022, 3, 532–541. [Google Scholar] [CrossRef]
- Fischer, R.; Byerlee, D.R.; Edmeades, G.O. Can Technology Deliver on the Yield Challenge to 2050? Food and Agriculture Organization of the United Nations, Economic and Social Development Department: Rome, Italy, 2009. [Google Scholar]
- Rong, L.-b.; Gong, K.-y.; Duan, F.-y.; Li, S.-k.; Ming, Z.; Jianqiang, H.; Zhou, W.-b.; Qiang, Y. Yield gap and resource utilization efficiency of three major food crops in the world—A review. J. Integr. Agric. 2021, 20, 349–362. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Zhao, M.; Wang, K.; Li, S.; Gao, Z.; Shi, X.; Chu, Q. Prospects for soybean production increase by closing yield gaps in the Northeast Farming Region, China. Field Crops Res. 2023, 293, 108843. [Google Scholar] [CrossRef]
- Mekonnen, A.; Getnet, M.; Nebiyu, A.; Abebe, A.T. Quantifying Potential Yield and Yield Gaps of Soybean Using CROPGRO-Soybean Model in the Humid Tropics of Southwestern Ethiopia. Int. J. Plant Prod. 2022, 16, 653–667. [Google Scholar] [CrossRef]
- Sentelhas, P.C.; Battisti, R.; Câmara, G.; Farias, J.; Hampf, A.; Nendel, C. The soybean yield gap in Brazil–magnitude, causes and possible solutions for sustainable production. J. Agric. Sci. 2015, 153, 1394–1411. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.F.; Mourtzinis, S.; Edreira, J.I.R.; Conley, S.P.; Gaska, J.; Kandel, H.J.; Lindsey, L.E.; Naeve, S.; Nelson, S.; Singh, M.P. Field validation of a farmer supplied data approach to close soybean yield gaps in the US North Central region. Agric. Syst. 2022, 200, 103434. [Google Scholar] [CrossRef]
- Grassini, P.; Eskridge, K.M.; Cassman, K.G. Distinguishing between yield advances and yield plateaus in historical crop production trends. Nat. Commun. 2013, 4, 2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Chung, G. Genetic improvement of cereals and grain legumes. Genes 2020, 11, 1255. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Bernard, S.M.; Andersen, G.L.; Chen, W. Developing microbe–plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2009, 2, 428–440. [Google Scholar] [CrossRef]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integr. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Yang, S.H.; Chung, G. Wild soybeans: An opportunistic resource for soybean improvement. In Rediscovery of Landraces as a Resource for the Future; IntechOpen: London, UK, 2018. [Google Scholar]
- Mba, C. Induced mutations unleash the potentials of plant genetic resources for food and agriculture. Agronomy 2013, 3, 200–231. [Google Scholar] [CrossRef] [Green Version]
- Roy, N.S.; Basnet, P.; Ramekar, R.V.; Um, T.; Yu, J.-K.; Park, K.-C.; Choi, I.-Y. Alternative Splicing (AS) Dynamics in Dwarf Soybean Derived from Cross of Glycine max and Glycine soja. Agronomy 2022, 12, 1685. [Google Scholar] [CrossRef]
- Bastia, R.; Pandit, E.; Sanghamitra, P.; Barik, S.R.; Nayak, D.K.; Sahoo, A.; Moharana, A.; Meher, J.; Dash, P.K.; Raj, R. Association Mapping for Quantitative Trait Loci Controlling Superoxide Dismutase, Flavonoids, Anthocyanins, Carotenoids, γ-Oryzanol and Antioxidant Activity in Rice. Agronomy 2022, 12, 3036. [Google Scholar] [CrossRef]
- Viana, J.M.S.; Dias, K.O.d.G.; Silva, J.P.A.d. Comparative Analysis of Pedigree-Based BLUP and Phenotypic Mass Selection for Developing Elite Inbred Lines, Based on Field and Simulated Data. Agronomy 2022, 12, 2560. [Google Scholar] [CrossRef]
- Afzal, M.A.; Azeem, F.; Afzal, S.; Afzal, N.; Rizwan, M.; Seo, H.; Shah, A.A.; Nawaz, M.A. Comparative Omics-Based Identification and Expression Analysis of a Two-Component System in Vigna radiata in Drought Stress. Agronomy 2023, 13, 989. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Zong, X.; Teng, C.; Hou, W.; Li, P.; Du, D. Genetic Diversity of Global Faba Bean Germplasm Resources Based on the 130K TNGS Genotyping Platform. Agronomy 2023, 13, 811. [Google Scholar] [CrossRef]
- Clever Mukuze, U.M.M.; Badji, A.; Maphosa, M.; Obua, T.; Kweyu, S.V.; Nghituwamhata, S.N.; Rono, E.C.; Kasule, F.; Habwa, P.T. Genome-wide association study for the detection of bruchid re-2 sistance loci in soybean. Agronomy 2023, 13, 31. [Google Scholar]
- Tajibayev, D.; Mukin, K.; Babkenov, A.; Chudinov, V.; Dababat, A.A.; Jiyenbayeva, K.; Kenenbayev, S.; Savin, T.; Shamanin, V.; Tagayev, K.; et al. Exploring the Agronomic Performance and Molecular Characterization of Diverse Spring Durum Wheat Germplasm in Kazakhstan. Agronomy 2023, 13, 1955. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, M.A.; Chung, G.; Golokhvast, K.S. The Genetics, Genomics, and Breeding of Cereals and Grain Legumes: Traits and Technologies for Future Food Security. Agronomy 2023, 13, 2065. https://doi.org/10.3390/agronomy13082065
Nawaz MA, Chung G, Golokhvast KS. The Genetics, Genomics, and Breeding of Cereals and Grain Legumes: Traits and Technologies for Future Food Security. Agronomy. 2023; 13(8):2065. https://doi.org/10.3390/agronomy13082065
Chicago/Turabian StyleNawaz, Muhammad Amjad, Gyuhwa Chung, and Kirill S. Golokhvast. 2023. "The Genetics, Genomics, and Breeding of Cereals and Grain Legumes: Traits and Technologies for Future Food Security" Agronomy 13, no. 8: 2065. https://doi.org/10.3390/agronomy13082065