

Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Field Management and Cultural Practices
2.3. Data Collection
2.4. Statistical Analysis
3. Results
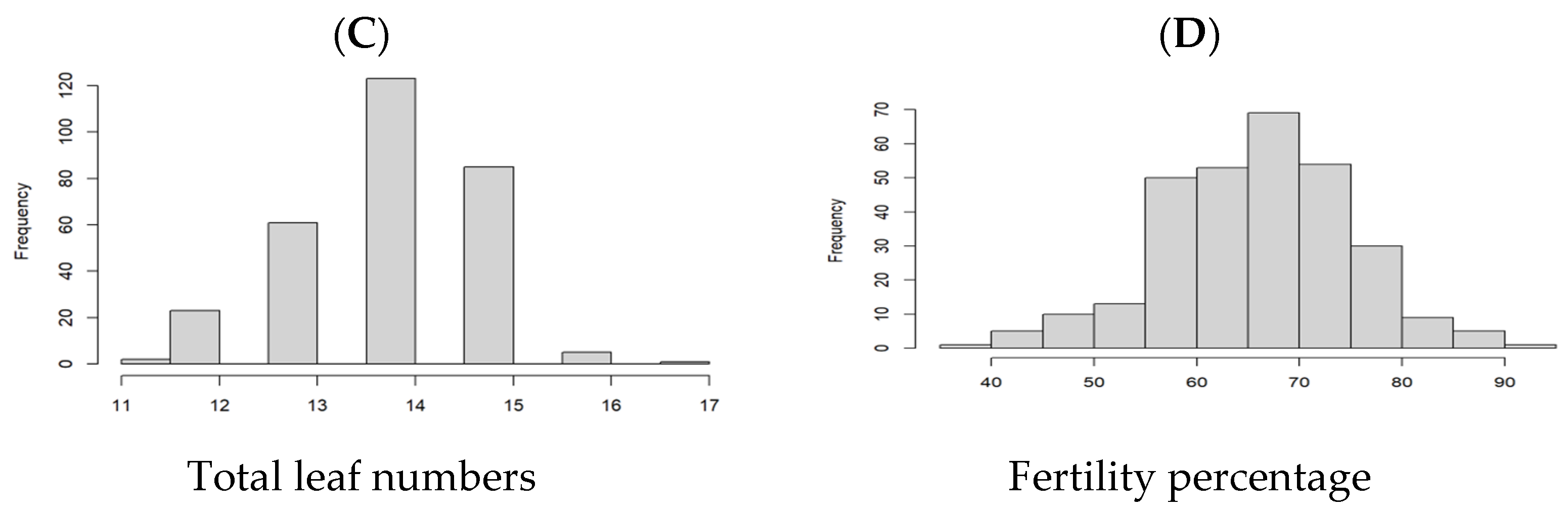
3.1. Phenology, Leaf Numbers, and Fertility Percentage
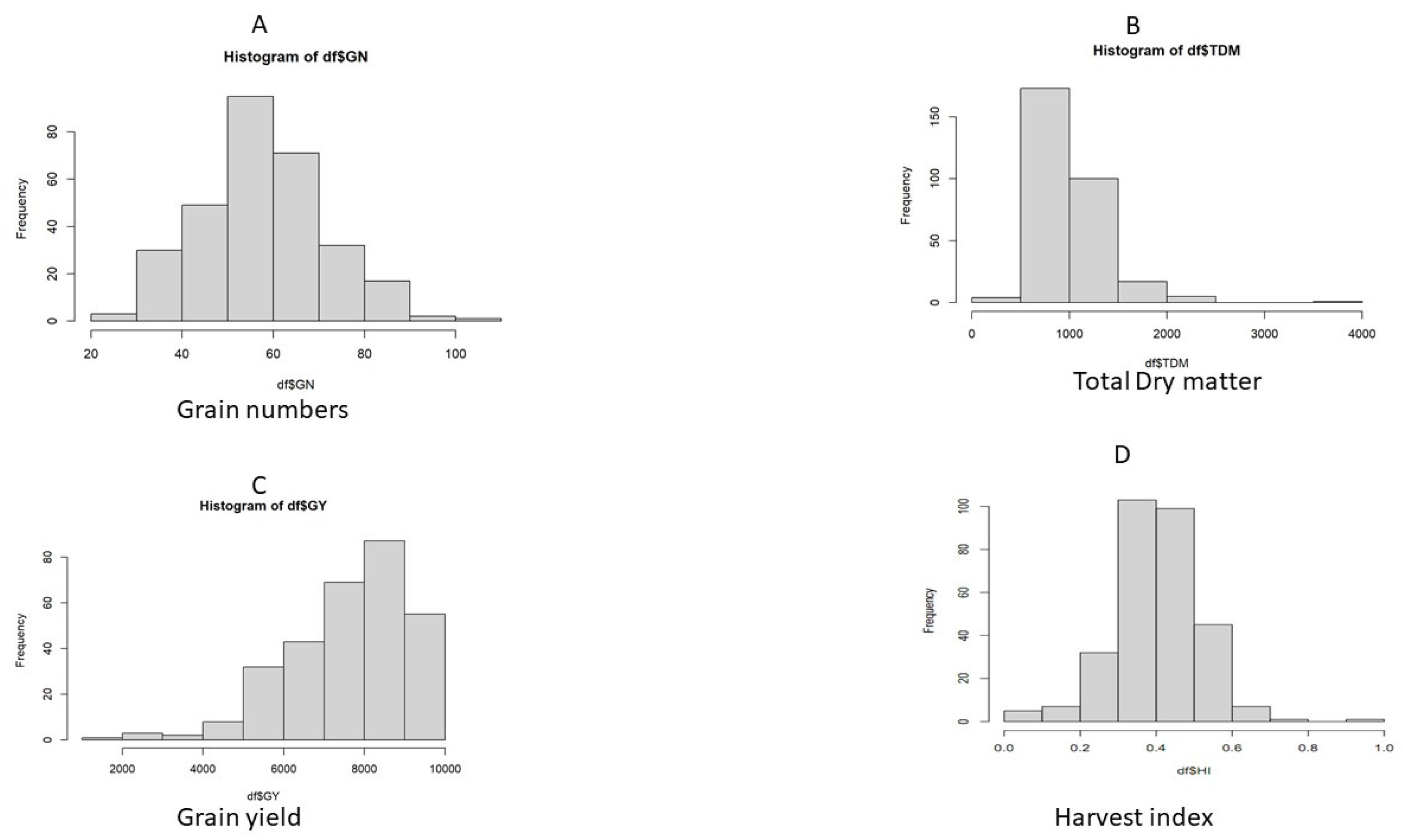
3.2. Total Dry Matter, Yield Components, and Harvest Index
3.3. Genotypic Variability
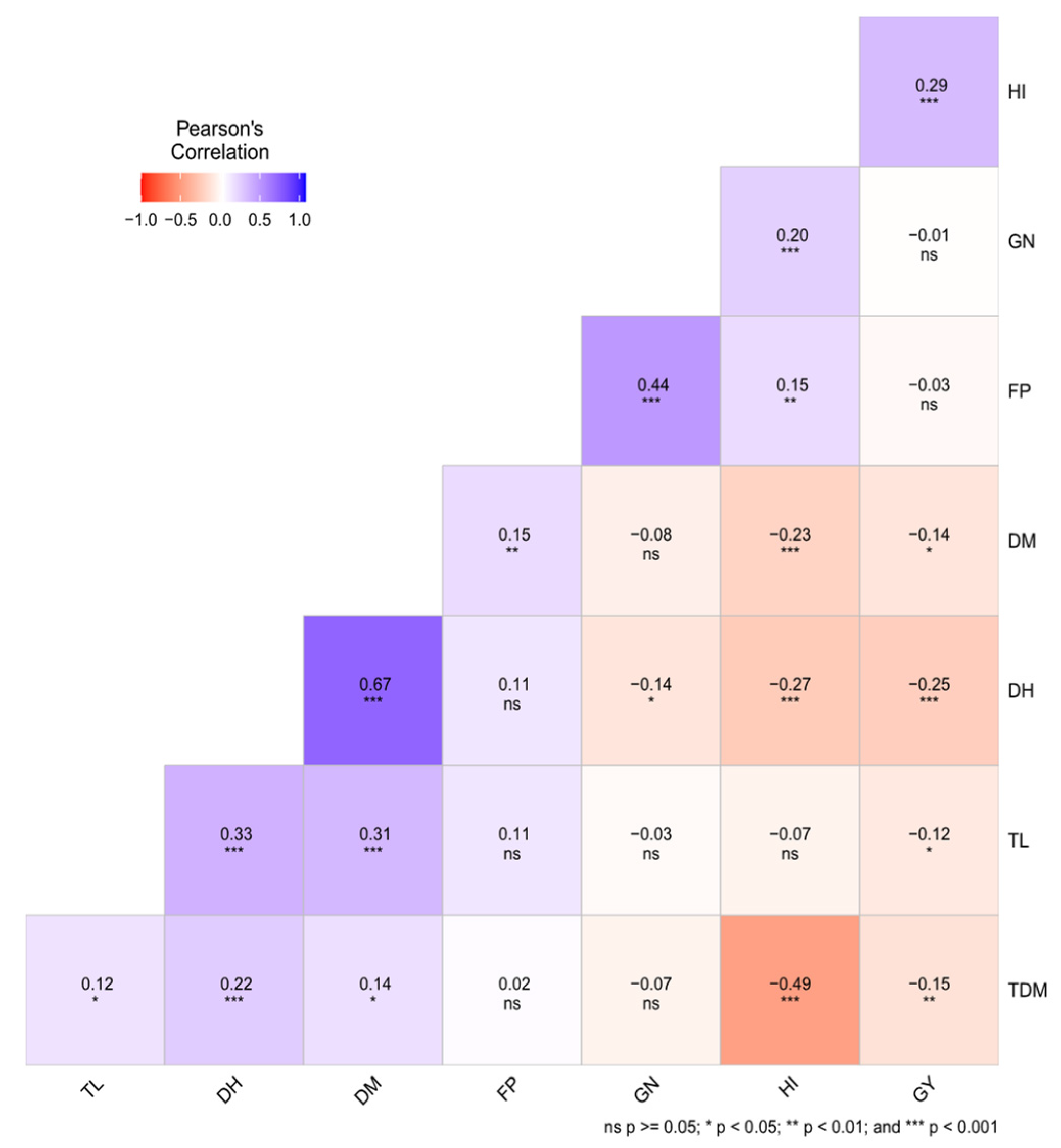
3.4. Correlation Study
3.5. Path Analysis
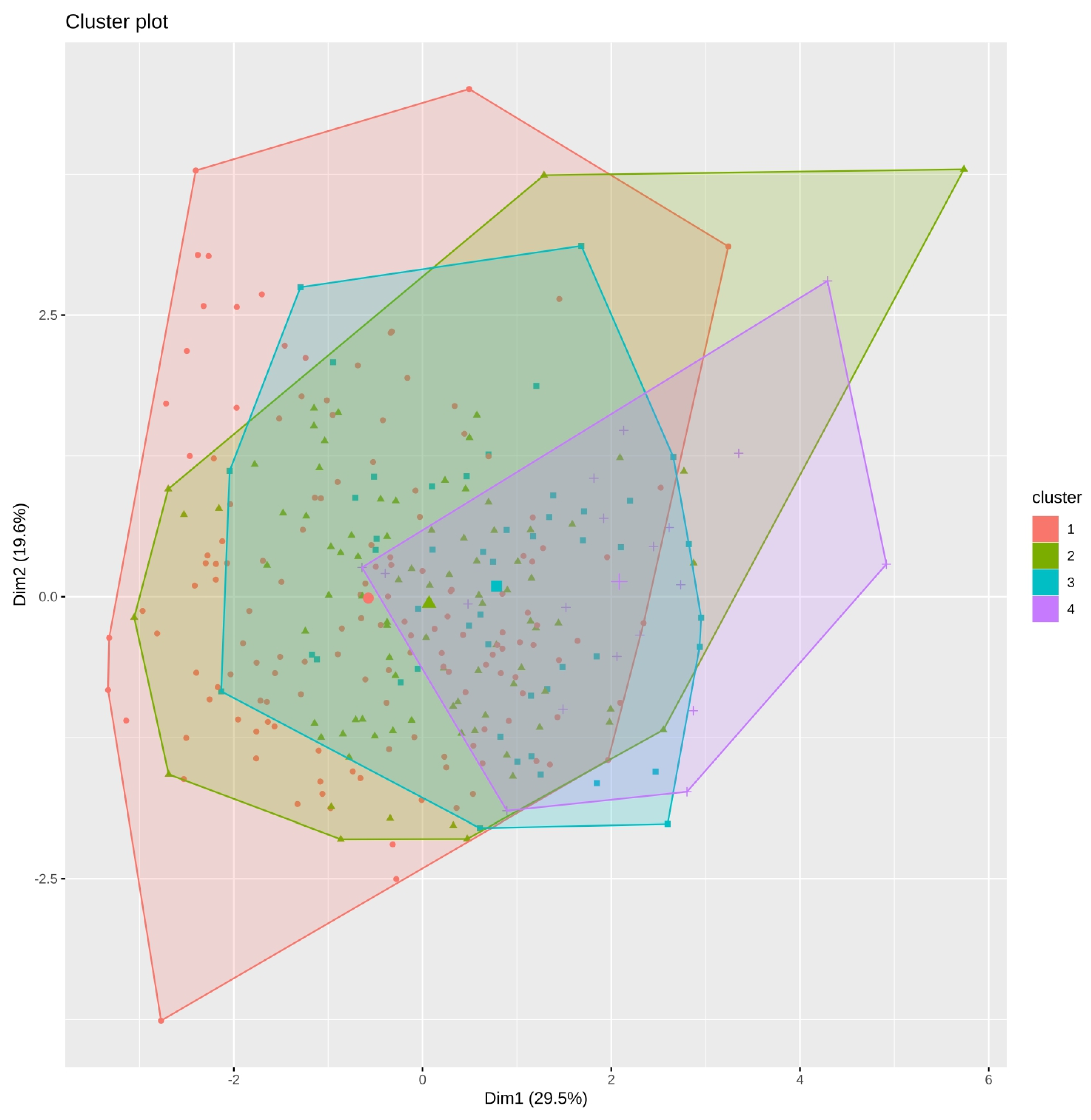
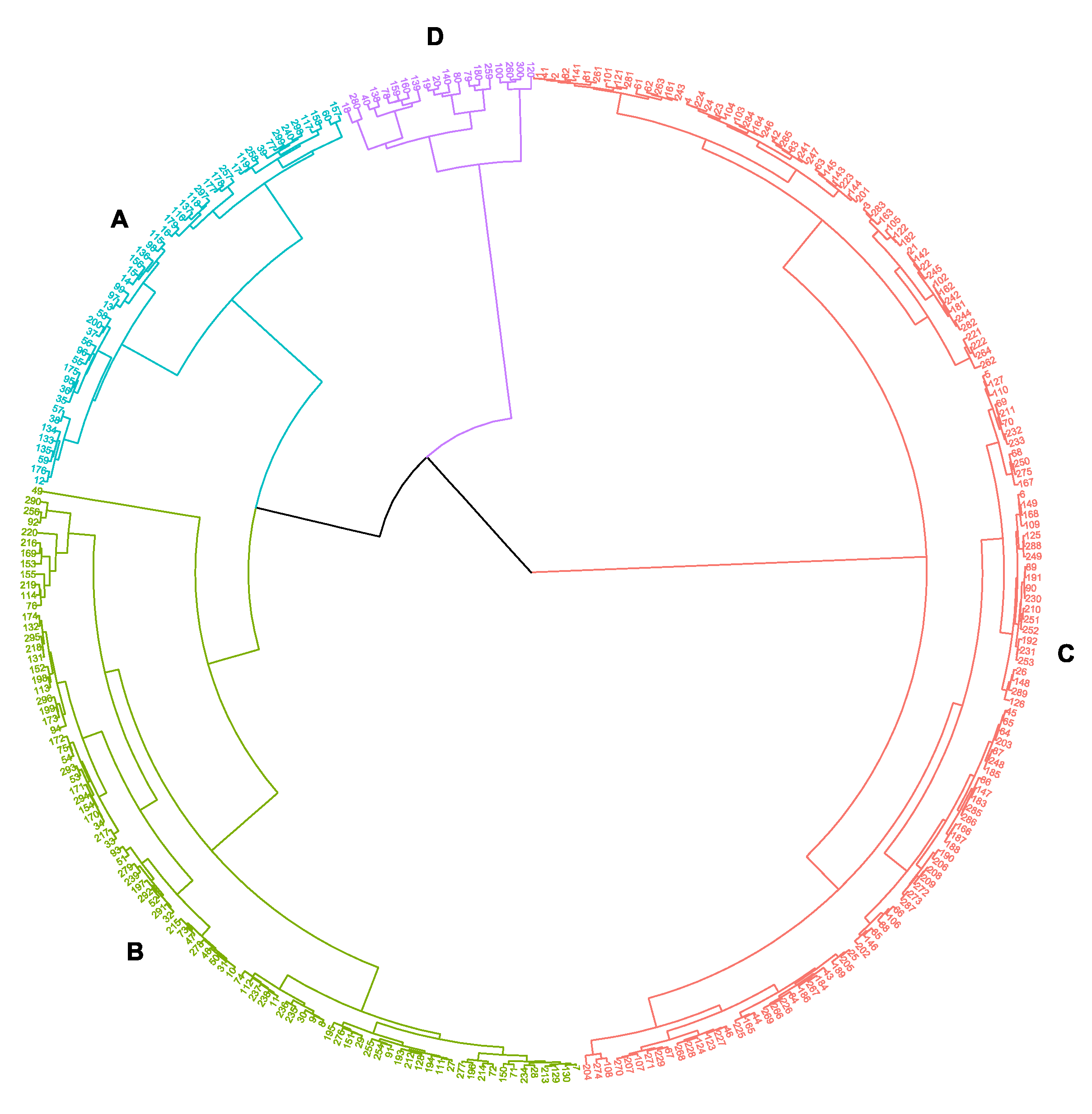
3.6. Principal Component Analysis and Classification of Accessions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmadi, N.; Bouman, B. Riz et rizicultures, enjeux économiques, écologiques et scientifiques. Cah. Agric. 2013, 2222, 333–335. [Google Scholar] [CrossRef]
- Mendez Del Villar, P.M.; Bauer, J.M. Le riz en Afrique de l’Ouest: Dynamiques, politiques et perspectives. Cah. Agric. 2013, 2222, 336–344. [Google Scholar]
- Maclean, J.; Hardy, B.; Hettel, G. Rice Almanac: Source Book for One of the Most Important Economic Activities on Earth, 3rd ed.; IRRI: Manila, Philippines, 2013; pp. 216–217. ISBN 0-85199-636-1. [Google Scholar]
- FAOSTAT Statistical Database; FAO (Food and Agriculture Organization of the United Nations): Rome, Italy, 2020; Available online: http://www.fao.org/faostat/en/#home (accessed on 21 June 2020).
- Mendez Del Villar, P.; Bauer, J.M.; Maiga, A.; Ibrahim, L. Crise Rizicole, Évolution des Marchés et Sécurité Alimentaire en AFRIQUE de l’Ouest; Appraisal Report; CIRAD: Montpellier, France, 2011. [Google Scholar]
- Seck, P.A.; Togola, A.; Touré, A.; Diagne, A. Propositions pour une optimisation des performances de la riziculture en Afrique de l’Ouest. Cah. Agric. 2013, 2222, 361–368. [Google Scholar]
- Panda, D.; Mohanty, B.; Behera, P.K.; Barik, J.; Mishra, S.S. Harnessing leaf photosynthetic traits and antioxidant defence for multiple stress tolerance in three premium indigenous rice landraces of Jeypore tract of Odisha, India. Funct. Plant Biol. 2020, 47, 99–111. [Google Scholar] [CrossRef]
- Singh, B.; Mishra, S.; Bisht, D.S.; Joshi, R. Growing Rice with Less Water: Improving Productivity by Decreasing Water Demand. In Rice Improvement; Ali, J., Wani, S.H., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Naz, M.; Hussain, S.; Javed, T.; Aslam, S.; Raza, A.; Ali, H.M.; Siddiqui, M.H.; Salem, M.Z.M.; et al. Exogenous salicylic acid-induced drought stress tolerance in wheat (Triticum aestivum L.) grown under hydroponic culture. PLoS ONE 2021, 16, e0260556. [Google Scholar] [CrossRef]
- Abdelhamid, H.; Sherbiny, E.; El-hashash, E.F.; El-enin, M.M.A.; Nofal, R.S.; El-mageed, T.A.A.; Bleih, E.M.; El-Saadony, M.T.; El-Tarabily, K.A.; Shaaban, A. Exogenously Applied Salicylic Acid Boosts Morpho-Physiological Traits, Yield, and Water Productivity of Lowland Rice under Normal and Deficit Irrigation. Agronomy 2022, 12, 1860. [Google Scholar]
- Hussain, T.; Hussain, N.; Tahir, M.; Raina, A.; Ikram, S.; Maqbool, S.; Fraz Ali, M.; Duangpan, S. Impacts of Drought Stress on Water Use Efficiency and Grain Productivity of Rice and Utilization of Genotypic Variability to Combat Climate Change. Agronomy 2022, 12, 2518. [Google Scholar] [CrossRef]
- Singhal, R.K.; Fahad, S.; Kumar, P.; Choyal, P.; Javed, T.; Jinger, D.; Singh, P.; Saha, D.; Md, P.; Bose, B.; et al. Beneficial elements: New Players in improving nutrient use efficiency and abiotic stress tolerance. Plant Growth Regul. 2023, 100, 237–265. [Google Scholar] [CrossRef]
- Agarwal, P.; Parida, S.K.; Raghuvanshi, S.; Kapoor, S.; Khurana, P.; Khurana, J.P.; Tyagi, A.K. Rice Improvement through Genome-Based Functional Analysis and Molecular Breeding in India. Rice 2016, 9, 1. [Google Scholar] [CrossRef]
- Janni, M.; Gullì, M.; Maestri, E.; Marmiroli, M.; Valliyodan, B.; Nguyen, H.T.; Marmiroli, N. Molecular and genetic bases of heat stress responses in crop plants and breeding for increased resilience and productivity. J. Exp. Bot. 2020, 71, 3780–3802. [Google Scholar] [CrossRef]
- Song, Y.; Wang, C.; Linderholm, H.W.; Fu, Y.; Cai, W.; Xu, J.; Zhuang, L.; Wu, M.; Shi, Y.; Wang, G.; et al. The negative impact of increasing temperatures on rice yields in southern China. Sci. Total Environ. 2022, 820, 153262. [Google Scholar] [CrossRef] [PubMed]
- Lawas, L.M.F.; Li, X.; Erban, A.; Kopka, J.; Jagadish, S.V.K.; Zuther, E.; Hincha, D.K. Metabolic responses of rice cultivars with different tolerance to combined drought and heat stress under field conditions. GigaScience 2019, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Bocco, R.; Lorieux, M.; Futakuchi, K.; Manneh, B.; Baimey, H.; Ndjiondjop, M.N. Agro-morphological characterization of a population of introgression lines derived from crosses between IR 64 (O. sativa indica) and TOG 5681 (O. glaberrima) for drought tolerance. Plant Sci. 2012, 183, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.S.; Behera, P.K.; Kumar, V.; Lenka, S.K.; Panda, D. Physiological characterization and allelic diversity of selected drought tolerant traditional rice (Oryza sativa L.) landraces of Koraput, India. Physiol. Mol. Biol. Plants 2018, 24, 1035–1046. [Google Scholar] [CrossRef]
- Muehe, E.M.; Wang, T.; Kerl, C.F.; Planer-Friedrich, B.; Fendorf, S. Rice production threatened by coupled stresses of climate and soil arsenic. Nat. Commun. 2019, 10, 4985. [Google Scholar] [CrossRef]
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; et al. Rational design of high-yield and superior-quality rice. Nat. Plants 2017, 3, 17031. [Google Scholar] [CrossRef]
- Shrestha, A.; Cosenza, F.; van Inghelandt, D.; Wu, P.-Y.; Li, J.; Casale, F.A.; Weisweiler, M.; Stich, B. The double round-robin population unravels the genetic architecture of grain size in barley. J. Exp. Bot. 2022, 73, 7344–7361. [Google Scholar] [CrossRef]
- Liu, K.; Harrison, M.T.; Yan, H.; Liu, D.L.; Meinke, H.; Hoogenboom, G.; Bin Wang, B.; Bin Peng, B.; Guan, K.; Jaegermeyr, J.; et al. Silver lining to a climate crisis in multiple prospects for alleviating crop waterlogging under future climates. Nat. Commun. 2023, 14, 765. [Google Scholar] [CrossRef]
- Anantha, M.S.; Patel, D.; Quintana, M.; Swain, P.; Dwivedi, J.L.; Torres, R.O.; Verulkar, S.B.; Variar, M.; Mandal, N.P.; Kumar, A.; et al. Trait combinations that improve rice yield under drought: Sahbhagi Dhan and new drought-tolerant varieties in South Asia. Crop Sci. 2016, 56, 408–421. [Google Scholar] [CrossRef]
- Singh, P.; Singh, S.K.; Korada, M.; Khaire, A.; Singh, D.K.; Habde, S.V.; Majhi, P.K.; Naik, R. Exploring variability and genetic diversity among rice genotypes in Eastern Uttar Pradesh. Electron. J. Plant Breed. 2022, 12, 1367–1374. [Google Scholar] [CrossRef]
- Chuchert, S.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Genetic diversity, genetic variability, and path analysis for yield and its components in indigenous upland rice (Oryza sativa L. var. glutinosa). Songklanakarin J. Sci. Technol. 2018, 40, 609–616. [Google Scholar] [CrossRef]
- Khomphet, T.; Intana, W.; Promwee, A.; Islam, S.S. Genetic Variability, Correlation, and Path Analysis of Thai Commercial Melon Varieties. Int. J. Agron. 2022, 7877239. [Google Scholar] [CrossRef]
- Ichsan, C.N.; Basyah, B.; Zakaria, S.; Efendi, E. Differences of water status and relationship with roots growth and yield of rice under water stress. Syst. Rev. Pharm. 2020, 11, 611–618. [Google Scholar]
- González Guzmán, M.; Cellini, F.; Fotopoulos, V.; Balestrini, R.; Arbona, V. New approaches to improve crop tolerance to biotic and abiotic stresses. Special Issue Article. Physiol. Plant. 2022, 147, e13547. [Google Scholar] [CrossRef]
- Diagne, A.; Mindingoyi, S.; Wopereis, M.; Akintayo, I. The NERICA Success Story: Development, Achievements and Lessons Learned; The Africa Rice Center (Africa Rice): Cotonou, Benin, 2010. [Google Scholar]
- Schober, P.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Lleras, C. Path Analysis. In Encyclopedia of Social Measurement; Kempf-Leonard, K., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2005; pp. 25–30. [Google Scholar] [CrossRef]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Combining ability and heritability for yield and its related traits in Thai. Agric. Nat. Resour. 2020, 54, 229–236. [Google Scholar]
- Raina, A.; Laskar, R.A.; Wani, M.; Jan, B.L.; Ali, S.; Khan, S. Gamma Rays and Sodium Azide Induced Genetic Variability in High-Yielding and Biofortified Mutant Lines in Cowpea [Vigna unguiculata (L.) Walp.]. Front. Plant Sci. 2022, 13, 911049. [Google Scholar] [CrossRef]
- Rashid, M.; Yousaf, Z.; Din, A.; Munawar, M.; Aftab, A.; Riaz, N.; Younas, A.; Alaraidh, I.A.; Okla, M.K.; AbdElgawad, H. Assessment of Mineral Nutrient Efficiency in Genetically Diverse Spinach Accessions by Biochemical and Functional Marker Strategies. Front. Plant Sci. 2022, 13, 889604. [Google Scholar] [CrossRef]
- Arpali, D.; Yagmur, M. The Determination of Selection Criteria Using Path Analysis in Two Rowed Barley (Hordeum vulgare L. Conv. Distichon). Turk. J. Agric. Nat. Sci. 2015, 2, 248–255. [Google Scholar]
- Shamuyarira, K.W.; Shimelis, H.A.; Mathew, I.; Tsilo, T.J. Correlation and path coefficient analyses of yield and yield components in drought-tolerant bread wheat populations. S. Afr. J. Plant Soil 2019, 36, 367–374. [Google Scholar] [CrossRef]
- Islam, S.; Anothai, J.; Nualsri, C.; Soonsuwon, W. Correlation and path analysis of phenological traits of Thai upland rice genotypes. Songklanakarin J. Plant Sci. 2020, 7, 133–142. [Google Scholar]
- Burton, G.W.; DeVane, E.H. Estimating Heritability in Tall Fescue (Festuca arundinacea) from Replicated Clonal Material1. Agron. J. 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Estimates of Genetic and Environmental Variability in Soybeans1. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Duangpan, S.; Buapet, P.; Sujitto, S.; Eksomtramage, T. Early assessment of drought tolerance in oil palm D × P progenies using growth and physiological characters in seedling stage. Plant Genet. Resour. 2018, 16, 544–554. [Google Scholar] [CrossRef]
- Haefele, S.M.; Wopereis, M.C.S.; Wiechmann, H. Long-term fertility experiments for irrigated rice in the West African Sahel: Agronomic results. Field Crop. Res. 2002, 78, 119–131. [Google Scholar] [CrossRef]
- IRRI. Standard Evaluation System (SES) for Rice; International Rice Research Institute: Los Banos, Philippines, 2002; 65p. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 1 December 2021).
- Grosjean, P.; Ibanez, F. pastecs: Package for Analysis of Space-Time Ecological Series. R Package Version 1.3.21. 2018. Available online: https://CRAN.R-project.org/package=pastecs (accessed on 1 December 2021).
- de Mendiburu, F. agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 1 December 2021).
- Popat, R.; Patel, R.; Parmar, D. variability: Genetic Variability Analysis for Plant Breeding Research. R Package Version 0.1.0. 2020. Available online: https://CRAN.R-project.org/package=variability (accessed on 1 December 2021).
- Olivoto, T.; Lúcio, A.D. metan: An R package for multi-environment trial analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; cluster: Cluster Analysis Basics and Extensions. R Package Version 2.1.4. 2022. Available online: https://CRAN.R-project.org/package=cluster (accessed on 1 December 2021).
- Kassambara, A.; Mundt, F. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 1 December 2021).
- Zeng, L.; Shannon, M.; Grieve, C. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Vendruscolo, T.P.S.; Barelli, M.A.A.; Castrillon, M.A.S.; da Silva, R.S.; de Oliveira, F.T.; Corrêa, C.L.; Zago, B.W.; Tardin, F.D. Correlation and path analysis of biomass sorghum production. Genet. Mol. Res. 2016, 15, 1–106. [Google Scholar] [CrossRef]
- Hussain, N.; Ahmed, M.; Duangpan, S.; Hussain, T.; Taweekun, J. Potential impacts of water stress on rice biomass composition and feedstock availability for bioenergy production. Sustainability 2021, 13, 10449. [Google Scholar] [CrossRef]
- Kalaiselvan, S.; Subramanian, A.; Thirumurugan, T.; Rajanbabu, V. Genetic variability and association studies in F2 population of rice under sodicity. Electron. J. Plant Breed. 2019, 10, 601–613. [Google Scholar] [CrossRef]
- Gunasekaran, A.; Seshadri, G.; Ramasamy, S.; Muthurajan, R.; Karuppasamy, K.S. Identification of Newer Stable Genetic Sources for High Grain Number per Panicle and Understanding the Gene Action for Important Panicle Traits in Rice. Plants 2023, 12, 250. [Google Scholar] [CrossRef]
- Khush, G. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Poehlman, J. Breeding Field Crops; The Avi Publishing Company, Incorporation: Westport, CT, USA, 1979. [Google Scholar]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Path analysis for yield and its components in F1 upland rice hybrids and their parental lines. Songklanakarin J. Sci. Technol. 2019, 41, 1419–1425. [Google Scholar]
- Lu, Y.; Chuan, M.; Wang, H.; Chen, R.; Tao, T.; Zhou, Y.; Xu, Y.; Li, P.; Yao, Y.; Xu, C.; et al. Genetic and molecular factors in determining grain number per panicle of rice. Front. Plant Sci. 2022, 13, 964246. [Google Scholar] [CrossRef]
- Vange, T. Biometrical studies on genetic diversity of some upland rice (Oryza sativa L.) accessions. Nat. Sci. 2009, 7, 21–27. [Google Scholar]
- Sumanth, V.; Suresh, B.G.; Ram, B.J.; Srujana, G. Estimation of genetic variability, heritability and genetic advance for grain yield components in rice (Oryza sativa L.). J. Pharmacogn. Phytochem. 2017, 6, 1437–1439. [Google Scholar]
- Ogunbayo, S.A.; Sié, M.; Ojo, D.K.; Sanni, K.A.; Akinwale, M.G.; Toulou, B.; Shittu, A.; Idehen, E.O.; Popoola, A.R.; Daniel, I.O.; et al. Genetic variation and heritability of yield and related traits in promising rice genotypes (Oryza sativa L.). J. Plant Breed. Crop Sci. 2014, 6, 153–159. [Google Scholar]
- Hussain, T.; Hussain, N.; Ahmed, M.; Nualsri, C.; Duangpan, S. Responses of lowland rice genotypes under terminal water stress and identification of drought tolerance to stabilize rice productivity in southern thailand. Plants 2021, 10, 2565. [Google Scholar] [CrossRef]
- Moukoumbi, Y.D.; El-Namaky, R.; Djaman, K.; Mbodj, D.; Manneh, B. Alternate phenotype–genotype selection for developing superior high-yielding irrigated rice lines. Crop J. 2018, 6, 191–201. [Google Scholar] [CrossRef]
- Xie, W.; Wang, G.; Yuan, M.; Yao, W.; Lyu, K.; Zhao, H.; Yang, M.; Li, P.; Zhang, X.; Yuan, J.; et al. Breeding Signatures of Rice Improvement Revealed by a Genomic Variation Map from a Large Germplasm Collection. Proc. Natl. Acad. Sci. USA 2015, 112, 5411–5419. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenological Stages | Dose | Split Period | |
|---|---|---|---|
| Vegetative phase | 60 kg N ha−1 | or 150–200 kg N.ha−1 N15P15K15 | Tillering corresponding at average 21 days after transplanting (DAT) |
| 60 kg K ha−1 (KCl) | |||
| 60 kg P ha−1 (DAP) | |||
| Reproductive phase | 60 kg N ha−1 | Panicle initiation | |
| 30 kg N ha−1 | Heading stage | ||
| Parameters | Cultivars | Residual | CV (%) |
|---|---|---|---|
| Days to heading | 247.60 *** | 34.50 | 9.20 |
| Days to maturity | 178.20 *** | 33.10 | 6.64 |
| Total leaf numbers | 2.80 *** | 1.00 | 9.49 |
| Grain numbers per panicle | 547.00 *** | 391.00 | 36.60 |
| Fertility percentage | 247.80 *** | 159.50 | 20.70 |
| Grain yield | 66,339,880.0 *** | 2,642,706.00 | 26.10 |
| Total dry matter | 408,954.00 *** | 180,509.00 | 50.30 |
| Harvest index | 0.02 *** | 0.01 | 29.20 |
| Attributes | GCV | PCV | H2 | GA | GAM | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Days to heading | 7.50 | L | 9.14 | L | 67.31 | H | 14.25 | M | 12.68 | M |
| Days to maturity | 5.01 | L | 6.52 | L | 59.11 | M | 11.00 | M | 7.94 | L |
| Total leaf numbers | 5.19 | L | 9.46 | L | 30.07 | M | 0.82 | L | 5.86 | L |
| Grain numbers per panicle | 12.30 | M | 36.12 | H | 11.58 | L | 5.03 | L | 8.62 | L |
| Fertility percentage | 8.21 | L | 20.66 | H | 15.77 | L | 4.46 | L | 6.71 | L |
| Grain yield | 15.09 | M | 26.07 | H | 33.50 | L | 1375.30 | H | 17.99 | M |
| Total dry matter | 27.02 | H | 49.42 | H | 29.89 | L | 311.46 | H | 30.43 | H |
| Harvest index | 16.88 | M | 29.13 | H | 33.58 | M | 0.08 | L | 20.16 | M |
| DH | DM | TL | TDM | FP | HI | GN | |
|---|---|---|---|---|---|---|---|
| DH | −0.1425 | −0.0164 | −0.0104 | −0.0059 | −0.0005 | −0.0848 | 0.0004 |
| DM | −0.0983 | −0.0238 | −0.0087 | −0.0045 | −0.0006 | −0.0745 | 0.00041 |
| TL | −0.0442 | −0.0062 | −0.0334 | −0.0036 | −0.0003 | −0.0514 | −0.0009 |
| TDM | −0.0442 | −0.0057 | −0.0063 | −0.0187 | −0.0004 | −0.1439 | −0.0011 |
| FP | −0.0199 | −0.0036 | −0.0030 | −0.0002 | −0.0039 | 0.0308 | 0.0097 |
| HI | 0.04702 | 0.0069 | 0.0067 | 0.0105 | −0.0005 | 0.2569 | 0.0025 |
| GN | −0.0029 | −0.0005 | 0.0017 | 0.0011 | −0.0020 | 0.0334 | 0.0191 |
| Residual effect = 0.86346888 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moukoumbi, Y.D.; Bayendi Loudit, S.M.; Sikirou, M.; Mboj, D.; Hussain, T.; Bocco, R.; Manneh, B. Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change. Agronomy 2023, 13, 2218. https://doi.org/10.3390/agronomy13092218
Moukoumbi YD, Bayendi Loudit SM, Sikirou M, Mboj D, Hussain T, Bocco R, Manneh B. Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change. Agronomy. 2023; 13(9):2218. https://doi.org/10.3390/agronomy13092218
Chicago/Turabian StyleMoukoumbi, Yonnelle Dea, Sandrine Mariella Bayendi Loudit, Mouritala Sikirou, Daouda Mboj, Tajamul Hussain, Roland Bocco, and Baboucarr Manneh. 2023. "Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change" Agronomy 13, no. 9: 2218. https://doi.org/10.3390/agronomy13092218
APA StyleMoukoumbi, Y. D., Bayendi Loudit, S. M., Sikirou, M., Mboj, D., Hussain, T., Bocco, R., & Manneh, B. (2023). Evaluation of Genotypic Variability and Analysis of Yield and Its Components in Irrigated Rice to Stabilize Yields in the Senegal River Valley Affected by Climate Change. Agronomy, 13(9), 2218. https://doi.org/10.3390/agronomy13092218