Abstract
Italian ryegrass (Lolium multiflorum Lam.) is a persistent weed species that poses significant management challenges in key agricultural crops such as wheat, corn, cotton, and soybean. This study investigated the prevalence of resistance to ACCase inhibitor herbicides, specifically diclofop and pinoxaden, among field-collected Italian ryegrass populations. The survey revealed widespread resistance to diclofop and emerging cross-resistance to pinoxaden. To elucidate the physiological mechanism of ACCase herbicide resistance, we investigated mutations in the carboxyl-transferase (CT) domain of the ACCase enzyme, a critical region for herbicide sensitivity. Using dCAPS assays and CT domain sequencing, several known resistance-conferring mutations were detected in diclofop survivors, including I1781L, W2027C, I2041N, D2078G, and C2088R. Additionally, other mutations such as L1701M, E1874A, N1878H, G1946E/Q, V1992D, and E2039D were identified. To understand the functional role of these mutations in herbicide resistance, homology modeling was performed using AutoDock Vina for selected mutation combinations. The computational analysis revealed that all mutations and their combinations resulted in reduced binding affinity with diclofop and pinoxaden compared to the wild-type ACCase CT domain. Computational binding energy predictions indicated that the G1946E mutation and the L1701M + I1781L + E1874A + N1878H combination exhibited the lowest affinities for diclofop and pinoxaden, respectively. This study provides valuable insights into the molecular basis of ACCase inhibitor resistance in Italian ryegrass. However, further research is needed to validate the functional significance of each new substitution and its combinations in conferring herbicide resistance.
1. Introduction
Ryegrass species, such as annual ryegrass (Lolium rigidum Gaud.) and Italian ryegrass (Lolium multiflorum Lam.), is a problematic weed in several agricultural systems. It is particularly troublesome in wheat, barley, oats, cotton, and corn production. Italian ryegrass competes vigorously with crops and reduces yields significantly if it is not effectively controlled. Its rapid growth and ability to establish dense populations make it a significant concern for small grain growers, especially in the southern and southeastern United States [1,2]. Ryegrass species produce large quantities of seeds (up to 45,000 per plant) [3] that can spread rapidly. It is a cool-season grass, which is widespread in temperate regions and capable of evolving quickly in response to environmental pressures, including herbicide selection. Mutations or genetic recombination events can give rise to new resistant biotypes, which can spread within and between populations, further aggravating resistance problems.
Group 1 herbicides, or ACCase inhibitors, provide effective options for controlling Italian ryegrass. Diclofop-methyl was the first commercial ACCase herbicide introduced in the late 1970s [4,5]. ACCase inhibitor herbicides are broadly categorized into three groups based on their chemical structures. The two common classes are aryloxyphenoxypropionates (also known as FOPs) and cyclohexanediones (also known as DIMs). A third class is phenylpyrazolines or phenylpyrrole herbicides (DENs).
In plant metabolism, ACCase is an important enzyme catalyzing the conversion of acetyl-coA to malonyl-coA, the first step of the fatty acid biosynthesis pathway in the chloroplast. It is critical for cellular, physiological, and defensive roles. Therefore, ACCase is an excellent target enzyme for herbicides used in agriculture. There are two isoforms of ACCase enzymes in plants: plastidic and cytoplasmic isoforms [6]. There is a substantial difference in the structure between these two isoforms, and ACCase inhibitors can interact only with plastidic ACCase in grass species by binding to the catalytic site and blocking its activity [7,8,9]. As a result, the synthesis of fatty acids, which are essential components of plant cell membranes and lipid storage molecules, is disrupted. Prolonged inhibition of ACCase results in fatty acid depletion that leads to alterations in the composition and structure of the cell membranes, causing leakage of cellular contents and, ultimately, cell death [5,10,11]. The FOPs, DIMs, and DENs inhibit ACCase enzymes by binding to a specific site in the CT domain [5,11,12,13,14].
Resistance to ACCase inhibitors is widespread in grass weed species. Approximately 51 weed species have been reported to evolve resistance to ACCase inhibitors in many countries across various continents, including North America, Europe, Australia, and Asia [15]. Many single mutations in the plastidic ACCase gene, resulting in amino acid substitutions endowing resistance to ACCase herbicides, have been identified from resistant weed populations and identified as the mechanism of resistance to ACCase-inhibiting herbicides [16]. Extensive research has been conducted on blackgrass (Alopecurus myosuroides Huds.), focusing on understanding the mechanisms of resistance, including target-site mutations in the ACCase gene [17,18]. Therefore, ACCase amino acid substitutions found across grass species are referenced with the corresponding position of plastidic ACCase (Genbank accession AJ310767) in blackgrass to indicate that the same substitution has been observed in blackgrass and is known to confer resistance. The most studied and validated resistance-conferring amino acid substitutions are I1781V/T, W1999L/C/S, W2027S/L/C, I2041V/N, D2078E/G, C2088R, and G2096S/A of the ACCase CT in blackgrass, ryegrass, and barnyard grass (Echinochloa crus-galli (L.) P. Beauv.) [10,18,19,20]. Along with TSMs, non-target-site resistance mechanisms like enhanced herbicide metabolism are also reported as resistance mechanisms [21,22]. Studying ACCase-resistant weeds helps researchers understand how resistance evolved at the molecular level, which results in reduced sensitivity of the ACCase enzyme to herbicide inhibition. This study was conducted with the following aims: (1) Evaluate the response to ACCase inhibitors using field-collected ryegrass populations, (2) Analyze the target site in survivors of diclofop using a derived Cleaved Amplified Polymorphic Sequence (dCAPS) assay and sequencing CT domain (from survivors of selected populations), (3) Carry out the computational modeling of single and multiple mutations found in this study.
2. Materials and Methods
2.1. Plant Material
This research involved Italian ryegrass populations collected from fields in Arkansas, Alabama, Louisiana, Georgia, North Carolina, and Mississippi. A susceptible (S) population was included in every experiment.
2.2. Response to Foliar-Applied ACCase Inhibitors
This study examined 21 field populations, comprising 20 from Arkansas and one from Alabama (Table S1), with experiments conducted between October 2015 and 2016. Seeds were sown in 50-well cellular trays filled with commercial potting mixture (Sunshine Mix®, Sun Gro Horticulture Inc., Bellevue, WA 98008, USA). Seedlings were thinned to one plant per cell (total of 50 seedlings) one week after emergence. The experiment was conducted in a completely randomized design with two replications. Cellular trays were kept in the greenhouse with day temperatures ranging from 22 to 35 °C and night temperatures ranging from 21 to 25 °C. At the three- and four-leaf stages, Italian ryegrass seedlings were treated with the field-recommended dose of diclofop (1× = 1120 g ai ha−1 Hoelon herbicide, Bayer CropScience, Research Triangle Park, NC 27709, USA) and pinoxaden (1× = 60 g ai ha−1 Axial XL, Syngenta Crop Protection, Inc., Greensboro, NC 27419, USA). Diclofop and pinoxaden were applied with 1 and 0.7% non-ionic surfactant (Induce®, Helena Chemical Co., Collierville, TN 38017, USA), respectively. Herbicide treatments were applied in a spray cabinet using a motorized boom equipped with flat fan nozzles (TeeJet spray nozzles, Spraying Systems Co., Wheaton, IL 60189, USA) delivering 187 L ha at 241 kPa. Plants were returned to the greenhouse immediately after herbicide treatment. Visible injury was assessed at 4 weeks after treatment (WAT) relative to the non-treated control using a 0–100% scale, where 0 corresponds to the absence of symptoms and 100% corresponds to plant death. Populations were categorized based on the range of visible injury: 0 to 30% as highly resistant (HR), 31 to 80% as moderately resistant (MR), and 81 to 100% as susceptible (S). The frequency of survival was calculated as the number of plants surviving 4 WAT divided by the total number of treated plants.
Data were analyzed using hierarchical clustering in JMP Pro v. 15 (SAS Institute, Cary, NC, USA) for injury of survivors. This experiment was carried out in two independent runs.
2.3. Genotyping by dCAPS Assay
For this experiment, 22 Italian ryegrass populations (Table S1) were treated with 2× (2240 g ai ha−1) diclofop using 15 seedlings established as described in Section 2.2 The greenhouse study was conducted between March and June 2017. Before herbicide application, leaf tissue from each plant was collected in an Eppendorf tube and kept at −80 °C until processed for DNA extraction. All survivors from each population were tagged as R, and five of those (R) samples were used for dCAPS assay. Leaf samples (100 mg) were ground to a fine powder using liquid nitrogen. Genomic DNA was extracted using the CTAB-based method. The CTAB extraction buffer (1.4 M NaCl, 100 mM Tris-HCl pH 8.0, 20 mM EDTA pH 8.0, 2% CTAB) was added to the powdered leaf tissue. After incubation at 65 °C for 1 h, the samples were purified with phenol/chloroform/isoamyl alcohol (25:24:1), and the DNA was precipitated with isopropanol. The DNA pellet was washed with 1 mL of 70% ethanol and air-dried. Finally, the DNA pellet was resuspended in 50 µL sterile water. All universal primers used were from [23]. To target codon 1781, selected reverse dCAPS primer was used in PCRs with the universal ACCase-targeting primer ACcp1 (5-CAACTCTGGTGCTIGGATIGGCA). PCR mixes were prepared as described by [24]. The same procedure was used to target codons 2027, 2041, 2078, 2088, and 2096. Forward or reverse dCAPS primers were used in PCRs with the universal ACCase-targeting primers ACcp2R or ACcp4. Cycling programs consisted of 95 °C 5 min followed by 37 cycles of 95 °C 5 s, Tm 10 s, and 72 °C 30 s. Digestions were performed at 37 °C for 3 h using 1 μL of the PCR mixes added with 5 U enzyme, 1.5 μL 10× compatible buffer, and 7 μL water. PCR products were visualized by electrophoresis on 3% agarose gels run in 0.5× TBE buffer.
2.4. Accumulation of Multiple Target-Site Mutations
Seeds of five populations—15AR-WHI-4, 13AL-2, 11NC-4, 12AR-WR-E, and 10GA-1 (Table S1)—were sown in 11 cm pots filled with commercial soil. Seedlings were thinned to one plant per pot two weeks after emergence. Plants were watered daily and fertilized with MiracleGro complete fertilizer (MiracleGro, The Scott’s Co., Marysville, OH 43041, USA) every week. Pots were kept in the greenhouse with day temperatures ranging from 22 to 35 °C and night temperatures ranging from 21 to 25 °C. Tillers of 20 plants from each population were divided into five pots to produce clones of seedlings. Four sets of clones were sprayed with 1× dose of either diclofop, pinoxaden, mesosulfuron, or pyroxsulam. One set of clones per accession was used as non-treated control. In this paper, we present data on diclofop- and pinoxaden-treated clones. Plants were rated for visible injury, as described in Section 2.2. The greenhouse work for this experiment was conducted between June and September 2017.
Before herbicide application, leaf tissue from each mother plant was collected in Eppendorf tube and kept at −80 °C for DNA extraction. Five and three plants from each R and S population were selected, respectively, for sequencing the CT domain of the ACCase gene. DNA was extracted as described in Section 2.3. The nearly full-length CT domain was amplified using Acclr9F (5′-ATGGTAGCCTGGATCTTGGACATG-3′) and Acclr6R (5′-GGAAGTGTCATGCAATTCAGCAA-3′) primer pair [17], purified, and sent for sequencing. The PCR was conducted in a 40 μL volume that consisted of 1 μL (60 ng) of DNA, 0.5 μM of each primer, and 20 μL of EmeraldAmp MAX PCR Master Mix (Takara, Kyoto, Japan). The PCR was run with the following profile: 95 °C for 45 s and 32 cycles of 95 °C for 1 min, 56 °C for 1 min, and 72 °C for 2 min, followed by a final extension step for 10 min at 72 °C. The PCR product was purified from agarose gel using Pure-Link Quick Gel Extraction Kit (Invitrogen, Carlsbad, CA, USA), and the purified fragment was divided into three aliquots and sent to Clemson University Genomics Institute—CUGI—for sequencing using forward (Acclr9F), reverse (Acclr6R), and an internal primer (LMAcIPF (5′-TGTTGGAACCATTTCTCTGG-3′)). The resulting overlapping fragments from each R and S plant were assembled into one sequence using Sequencher 5.4.6 (Gene Codes Corporation, Ann Arbor, MI, USA) to obtain the complete coding region of the CT domain of ACCase gene. The nucleotide sequences were translated into open reading frames using the online ExPASy translation tool. The chromatograms for each mutation were examined manually to confirm the presence of the double and a single peak for the interpretation of heterozygous and homozygous mutations, respectively.
2.5. In Silico Prediction of Herbicide Binding
Ligand and receptor preparation: The molecular structures of diclofop and pinoxaden were retrieved from the PubChem database and analyzed using Discovery Studio Visibleizer. Subsequently, the PDB of ligand structure was created using I-Tasser [25,26,27]. The finalized conformations devoid of imaginary frequencies were chosen as initial structures for the docking analysis. We used PYMOL to generate mutations on wild-type ACCase protein. The mutations created in this analysis were the ones that were found in the resistant ryegrass plants in this study.
Molecular docking: The docking simulations were conducted using Autodock Vina. Semi-flexible docking was employed, allowing ligands to explore various conformations while keeping the protein rigid [28]. The docking site was defined within the binding region of the co-crystallized ligand. Analysis of results was performed using Discovery Studio Visibleizer. Hydrogen bonding criteria were set at <2.50 Å between donor and acceptor atoms, with a minimum donor-hydrogen-acceptor angle of 120°, utilizing default docking parameters.
Autodock Vina: The ACCase protein structure was prepared computationally by adding polar hydrogen atoms and assigning Kollman charges. The docking box was centered at the CT domain in the crystal structure. Vina forcefield was used, and the box dimensions were set to 25 Å in each dimension [28]. The exhaustiveness of the global search was set to 8, with a maximum of 9 binding modes considered. Scoring function weights and terms were maintained at default values.
3. Results
3.1. Response to Foliar-Applied ACCase Inhibitors
Among the populations studied (n = 21), 95% (n = 20) and 33% (n = 7) were resistant to the label rate of diclofop and pinoxaden, respectively. At least two populations (15AR-WHI-4 and 15AR-WHI-2) were highly cross-resistant (<20% injury). The rest varied in their response to both herbicides while overall exhibiting high resistance to diclofop and low resistance to pinoxaden (Figure 1). Ten populations were HR and MR, and only one population was susceptible to diclofop. Two, five, and fourteen populations were HR, MR, and S to pinoxaden. The mean survival frequency among all populations was 69% with diclofop and 13% with pinoxaden (Table S2).
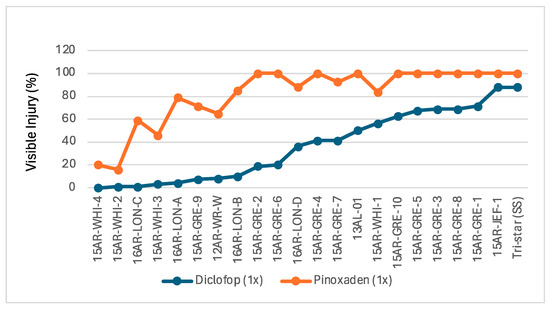
Figure 1.
Visible injury (%) among Italian ryegrass populations in response to ACCase inhibitors. Populations are arranged primarily in increasing levels of susceptibility to diclofop (blue line). Response to pinoxaden is represented by the orange line. Clones of all populations were treated with field recommended rate (1×) of diclofop (1120 g ai ha−1) and pinoxaden (60 g ai ha−1). Each data point is the average of four replications.
3.2. Genotyping of ACCase Mutations by dCAPS Assay
To investigate the molecular mechanism of resistance to diclofop, we selected 22 diclofop-resistant ryegrass populations from a previous [29] and the current study. We analyzed the presence of seven known TSMs—L1781M, W1999C, W2027C, I2041N, D2078G, C2088R, and G2096S—in five survivors (total of 115) from each resistant population (Figure 2). The diclofop response of each population in terms of visible injury was plotted from the current and previous [29] studies, as some populations used in dCAPS analysis were already characterized previously in our laboratory. The results of the dCAPS assay showed that 56 out of 115 plants did not carry any of the seven TS mutations tested. All samples from three populations—10NC-1, 15AR-GRE-1, and 11AR-03—did not carry any of the seven known resistance-conferring mutations tested by the dCAPS assay. The rest of the 20 populations were found to contain at least one resistance-conferring TS mutation. Among all the plants, the frequency of the I2041N mutation was found to be high (48%) along with W2027C (4%), L1781M (2%), and C2088R (1%). None of these populations showed the presence of W1999C, D2078G, or G2096S. The majority of the populations carried a single mutation; however, two populations (15AR-WHI-2 and 11AR-05) harbored two mutations in at least one of the five plants tested. Two populations, 15AR-GRE-1 and 11AR-03, did not show any known mutations and were also in the category of plants having high (70–75%) visible injury.
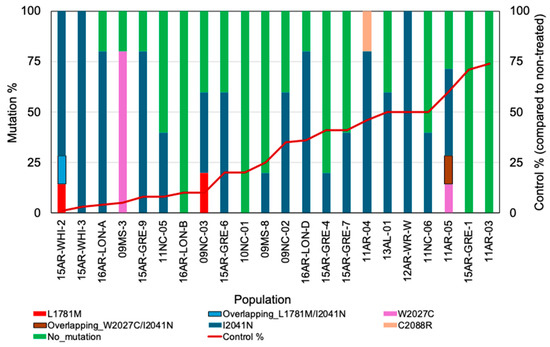
Figure 2.
Frequency of known mutations among the diclofop survivors of L. multiflorum populations. Mutation frequency in a population was calculated as % from the number of each mutation found within the five plants per population. The populations are listed in the order of increasing visible injury (shown on the right y-axis) upon diclofop treatment, which is depicted as a line graph (red line).
3.3. Accumulation of Multiple TSMs
The dCAPS analysis showed that many plants from diclofop-resistant populations did not carry any of the known TSM. We selected five resistant populations showing diverse visible injuries in response to diclofop and pinoxaden from the preliminary screening. The tillers of a single plant were cloned, and two sets of clones were sprayed with diclofop and pinoxaden. Visible injuries of 20 clones per herbicide per population were recorded (Table S3). Five plants per population showing a strong R phenotype to diclofop were selected. Three plants from the S population were also included. The entire CT domain (approximately 1600 bp) of the plastid form of the ACCase gene of the mother plants was Sanger-sequenced. We detected 11 eleven non-synonymous SNPs within these plants, of which four are known resistance-conferring TS mutations: I1781L, I2041N, D2078G, and C2088R. Other substitutions that were found in survivors were L1701M, E1874A, N1878H, G1946E/Q, V1992D, and E2039D. All three S plants did not carry any non-synonymous SNPs in the CT domain of the ACCase gene (Table 1).

Table 1.
Resistance-conferring mutation profile (substitutions are color-shaded) of the ACCase CT domain from five selected Italian ryegrass populations. New SNPS detected in this study are highlighted in bold.
We found two plants, each from 10GA-1 and 13LA-2, that were double mutants of I1781L and D2078G in heterozygous form. The G1946E was the most prevalent (80%), followed by the known I2041N (44%) mutation detected among 25 plants. The 15AR-WHI-4, 12AR-WR-E, 11NC-4, and 13AL-2 showed characteristic mutation profiles, with most of the plants displaying similar composition within the population. All five plants from the 15AR-WHI-4 population showed the presence of two mutations, G1946E and C2088R (that is, G1946E + C2088R; hereafter, the presence of multiple mutations will be presented as a combination), in the homozygous and heterozygous states, respectively. All five plants from the 12AR-WR-E population showed the presence of G1946E/Q + I2041N, with one plant showing the presence of additional L1701M + E1874A + N1878H substitutions. The four plants from the 11NC-4 population displayed a combination of three mutations, V1992D + E2039D + I2041N, and a total of three plants showed the presence of an additional mutation, G1946E. All five plants from 13LA-2 showed the presence of L1701M + I1781L + E1874A + N1878H + G1946E, with one plant displaying an additional combination of V1992D + E2039D + D2078G. Regarding the population, 10GA-1, there was no common profile of mutation found among all plants; however, four plants showed the presence of at least one missense mutation within the CT domain of the ACCase gene.
3.4. Computational Modeling
In our deeper investigation of resistance mechanisms in diclofop- and pinoxaden-resistant Italian ryegrass populations, we conducted computational molecular docking studies to assess the impact of various mutations on the binding affinity of these herbicides to the CT domain of ACCase (Table 2). The wild-type ACCase exhibited strong binding affinities of −6.2 kcal/mol for both herbicides. The mutations I1781M, I2041N, C2088R, and G1946E resulted in decreased binding affinities with binding energies ranging from −4.7 to −5.4 kcal/mol and −5.1 to −5.7 Kcal/mol for diclofop and pinoxaden, respectively. The binding affinity of double mutants G1946E + C2088R and G1946E + I2041N was −4.9 and −4.7 Kcal/mol with diclofop and 5.2 Kcal/mol with pinoxaden, respectively. Additionally, mutations at other positions, including a combination of four mutations (L1701M + I1781L + E1874A + N1878H), resulted in the lowest binding energy (−4.8 Kcal/mol) with pinoxaden.
Table 2.
Binding affinity of single ACCase mutations and combinations of ACCase mutations modeled in CT domain and herbicides (diclofop and pinoxaden) as predicted by AutoDockVina.
4. Discussion
The repetitive use of ACCase inhibitors or other types of herbicides imposes selective pressure, favoring the survival and reproduction of resistant plants. Screening experiments revealed that Italian ryegrass populations in Arkansas are highly resistant to the field-recommended dose of diclofop, which has been the primary herbicide for ryegrass control in wheat production for decades, making it the main selector for Group 1 herbicide resistance. Certainly, more than 95% of Italian ryegrass populations in Arkansas have developed resistance to diclofop. This high level of resistance is a clear indication of the extensive use of this herbicide in wheat production over many years. Pinoxaden, the latest Group 1 herbicide introduced for wheat production, shows a lower frequency of resistance compared to diclofop (Figure 1). This indicates that the resistance to pinoxaden is at an early evolutionary stage. However, it is noteworthy that a substantial proportion (roughly 1/3) of diclofop-resistant populations exhibit cross-resistant to pinoxaden. Previous resistance profiling studies [29] have confirmed this cross-resistance pattern, indicating that several diclofop-resistant populations cannot be effectively controlled by pinoxaden. The observed cross-resistance between diclofop and pinoxaden in Italian ryegrass populations is likely conferred by specific ACCase mutations that reduce binding affinity to both herbicides. This outcome can be explained by the partial overlap in the binding sites of diclofop and pinoxaden on the ACCase enzyme [30]. Similarly, other research groups have reported increasing resistance to pinoxaden among Italian ryegrass populations in wheat fields [31,32,33].
Understanding the underlying molecular mechanism of resistance to ACCase inhibitors in Italian ryegrass populations allow us to anticipate the potential for resistance development and design proactive management strategies to prevent it. Research data available so far indicates that the resistance to ACCase inhibitors primarily results from molecular adaptation within the CT domain of the ACCase gene, which is the target site for these herbicides [34]. Molecular adaptation is typically assumed to evolve by sequential fixation of beneficial mutations. Herbicide targets are generally key enzymes in plant biochemical pathways. Due to natural genetic diversity, rare mutations in these enzymes can occur, sometimes resulting in variants that remain functional while being less affected by herbicides. Some enzymes, such as ACCase and ALS (acetolactate synthase), have higher mutation rates than others, such as the glyphosate target EPSPS (5-enolpyruvylshikimate 3-phosphate synthase), which is highly conserved. As a result, resistance to ACCase- and ALS-inhibiting herbicides often develops through several TS mutations, while resistance to glyphosate usually occurs through other mechanisms or a few TS mutations.
In the dCAPS study, we found that the severity of injury was not associated with a certain type of mutation. For example, I2041N was found in plants that were highly resistant (0–20% injury) as well as in moderately resistant plants (40–60% injury) (Figure 2). The I2041N mutation has been associated with resistance to FOP herbicide [10], but it contributes high to moderate resistance across different populations. The zygosity of I2041N may be responsible for differential response, as reported by [35,36]. Alternatively, it also indicates that other mechanisms contribute to resistance in such cases. In our dataset, the contribution to resistance by the NTSR mechanism was also evident by the fact that 50% of R plants did not carry known TS mutation (Figure 2). Resistant plants in two populations, 15AR-GRE-1 and 11AR-03, showed the absence of any known TS mutation with high injury (Figure 2), which further suggests the presence of an NTSR mechanism. Out of seven, three resistance-conferring codon positions, 1781, 1999, and 2041, are directly involved in herbicide-binding sites in the CT domain. The remaining four codon positions—1927, 2078, 2088, and 2096—are in the close vicinity of the herbicide-binding site. Also, the herbicide-binding site overlaps with the substrate (acetyl-CoA)-binding site to some extent in the CT domain [30]. Therefore, mutation at some of the amino acid positions, such as 1999, 2078, and 2088, are reported to be responsible for fitness cost, most likely resulting from a decrease in overall ACCase activity that is required for fatty acid synthesis. This could be the reason for the absence or low frequency of W1999C, D2078G, and C2088R mutations in the resistant plants tested in this study.
Although the dCAPS analysis indicated the presence of NTSR factors, we conducted Sanger sequencing of the ACCase CT domain in selected populations to verify the presence/absence of other mutation/s not covered by dCAPS analysis. The CT domain is functionally critical, and mutations concentrated in this region are likely adaptive and associated with herbicide resistance. Given the diploid nature of the Italian ryegrass genome, heterozygous polymorphic positions would exhibit two nucleotides representing two alleles. We carefully analyzed the chromatographs to accurately identify homozygous and heterozygous polymorphic peaks, which is crucial for determining the zygosity of each mutation. Examination of the ACCase CT domain genotype in all survivors (Table 1) revealed the accumulation of multiple mutations in single ACCase alleles, potentially resulting in substantial structural changes to the herbicide-binding site. The zygosity of these mutations influences the resulting ACCase enzyme variants: two heterozygous mutations may produce either a double-mutant enzyme or two single-mutant forms, while a combination of homozygous and heterozygous mutations would yield both double-mutant and single-mutant enzymes. Our analysis identified several distinctive mutational profiles (highlighted as background color in Table 2) across the populations, suggesting that resistance-conferring mutations become more prevalent over time. We detected substitutions at different positions within the CT domain that are well characterized for their role in resistance to ACCase inhibitors such as I1781L, I2041N, D2078G, and C2088R. Other new substitutions that we found in this study, such as L1701M, E1874A, N1878H, G1946E/Q, V1992D, and E2039D, have not been characterized before. All the new mutations except V1992D and E2039D are reported [17,19], but their linkage to resistance to ACCase inhibitors is not validated. The presence of a single mutation in the CT domain as the basis of resistance to ACCase inhibitors is well known [9], but multiple mutations in a single resistant plant have also been reported [18,37,38]. The G1946E mutation was found to be present at the highest frequency among all five geographically distant resistant populations from Arkansas, Louisiana, North Carolina, and Georgia. To further investigate if G1946E is associated with resistance or concurrently enriched among resistant populations, de novo prediction was carried out using docking studies. Docking studies can help elucidate the molecular mechanisms by simulating the interaction between herbicides and target proteins. It can predict how specific mutations or structural alterations in the target protein affect herbicide binding and efficacy. In this study, we conducted a computational analysis to assess the impact of known and newly detected substitutions and their combination on the binding affinity of the diclofop and pinoxaden with the ACCase CT domain. Results revealed the distinct binding affinities between the wild-type and mutant variants (Table 2) of the CT domain. Lesser binding energy (more negative score) indicates stronger binding of diclofop and pinoxaden to the ACCase catalytic site; conversely, more binding energy (less negative score) indicates a weakened interaction. The wild-type structure showed the most negative score of −6.2 (Table 2) or the strongest binding with diclofop and pinoxaden, as anticipated. The structural change in the CT domain binding site by changing G at 1946th position to E resulted in the lowest negative score and predicted the weakest interaction with both herbicides, which suggests that the herbicide-binding site is affected by G1946E substitution. Also, the addition of G1946E with other known resistance-conferring mutations, such as I2041N and C2088R, weakened the interaction between the CT domain and the herbicide molecules.
The resistant population, 15AR-WHI-4, was found to be highly resistant to diclofop and cross-resistant pinoxaden (Figure 1 and Table S3). Detection of known C2088R mutation in all the five plants from 15AR-WHI-4 can explain the cross-resistance as the mutation at this codon is reported to confer broad resistance to all commonly used ACCase-inhibiting herbicides [39]. All plants within this population presented a double mutation of C2088R + G1946E in the same allele of ACCase (Table 1), which suggests that the quantitative effect of two substitutions may provide additional benefit to the diclofop and pinoxaden binding site. This additive effect was supported by the docking study (Table 2), which showed a weakened interaction of C2088R + G1946E compared to C2088R with diclofop and pinoxaden. The C2088R mutation did not complement the yeast ACC1 null mutation, suggesting cytosine at position 2088 is essential for total ACCase activity [30]. Therefore, mutation at this position compromises the ACCase activity and is found to be associated with fitness penalty in resistant plants [5,18]. In agreement with these reports, we found the presence of a heterozygous C2088R allele in all R plants from the highly cross-resistant population, 15AR-WHI-4, which allows plants to survive on wild-type ACCase alleles under diclofop and pinoxaden selection. The 12AR-WR-E population exhibited cross-resistance (Figure 1 and Table S3), with the majority of resistant plants carrying the I2041N + G1946E double mutation. While I2041N is known to confer resistance specifically to FOPs, it has not been associated with resistance to pinoxaden [10,36]. The computational study showed that the addition of G1946E to I2041N significantly lowered the diclofop binding affinity but not the pinoxaden compared to I2041N, and therefore, it is difficult to predict if G1946E would be involved in pinoxaden resistance. It is likely that NTS factors may be responsible for cross-resistance in the 12AR-WR-E population. The plants from 11NC-1 also carried the I2041N + G1946E mutation, but this population was characterized as highly diclofop resistant and slightly cross-resistant to pinoxaden [29] and (Table S3), which supports our interpretation that I2041N + G1946E is mainly responsible for diclofop resistance. Interestingly, the simulation of a combination of L1701M + I1781L + E1874A + N1878H that was found in each R plant from 13LA-2 predicted the most reduced pinoxaden binding affinities compared to the wild-type or I1781L mutant protein, which suggests the molecular bases of pinoxaden binding specificity. The amino acid position 1781 within ACCase protein is directly involved in the herbicide-binding site and reported to confer resistance to all three classes of ACCase herbicides [10,30]. The presence of multiple substitutions along with I1781L in R plants from 13LA-2, which is cross-resistant to pinoxaden (Table S3), indicates that the pinoxaden binding site is significantly altered and may be responsible for cross-resistance. Although, it is difficult to interpret from our data that all new substitutions will be responsible for resistance, too, and further investigation is required to validate its role. A single plant from the 10GA-1 population showed the absence of any ACCase mutation in the CT domain, which strongly suggests the presence of other resistant factors that may have segregated within the population and contributed to resistance together with other TSM in the rest of the plants.
The evolution of herbicide resistance through target-site mutations (TSMs) is driven by several evolutionary forces, including herbicide selection pressure, fitness costs, genetic variation, gene flow, and multiple resistance mechanisms [16,40,41,42]. The repeated use of herbicides over extended periods imposes strong selection pressure on weed populations, favoring the emergence of de novo mutations in target sites that enable plants to survive herbicide applications. The new mutations identified in this study are found to co-occur with the known mutation on the same allele, suggesting they may have arisen through sequential accumulation over multiple generations. Accumulation of additional mutations may enhance resistance or mitigate fitness costs associated with the first mutation [42,43]. Although the computational study predicted the G1946E substitution as the stronger mutation that can result in the weakest interaction with the herbicide, we did not find the R plant with only the G1946E substitution. The G1946E was always found with other known resistance-conferring mutations, which suggests that G1946E itself might not confer strong resistance but may offer an additive effect. Overall, the docking studies provided valuable insight into the newly found substitutions with the diclofop and pinoxaden binding that accumulation of multiple substitutions may exhibit synergistic effects, further enhancing herbicide resistance. However, it is essential to experimentally validate the contribution of each mutation or combination of mutations to verify its effect on enzyme function to ascertain its relevance in agricultural environments. Therefore, potential future studies may include in vitro enzymatic assays with herbicides and substrate using recombinant ACCase enzyme with double mutation and single mutation.
5. Conclusions
This study demonstrates widespread resistance to diclofop in Italian ryegrass populations, with emerging resistance to pinoxaden, indicating the evolution of resistance to multiple Group 1 herbicides. Molecular studies conducted with diclofop- and pinoxaden-resistant populations identified four known TSMs (I1781L, I2041N, D2078G, and C2088R) in the CT domain of the plastidic ACCase gene. Computational prediction of the lower binding affinity of ACCase protein modeled with G1946E and L1701M + I1781L + E1874A + N1878H with ACCase inhibitors warrants further studies with larger datasets and an in vivo validation approach.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy14102316/s1. Table S1: Collection details of ryegrass populations used in this study; Table S2: Mortality response of Italian ryegrass populations to foliar-applied diclofop and pinoxaden; Table S3: Visible injury response of Italian ryegrass populations to foliar-applied diclofop and pinoxaden.
Author Contributions
G.R. and A.C.L. contributed equally to this work; G.R. analyzed the molecular data and wrote and revised the manuscript; A.C.L. and R.A.S.-P. conducted and analyzed greenhouse experiments; A.C.L. conducted molecular experiments; S.A. and C.E.N. conducted computational modeling and analysis; N.R.-B. and G.R. conceptualized this project; J.C.V. analyzed the data; N.R.-B., G.R., S.A. and C.E.N. edited and reviewed the manuscript; N.R.-B. contributed to funding acquisition, collection of Italian ryegrass populations, and project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Hatch Project 2223 (Herbicide-resistant weeds in Arkansas cropping systems).
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Materials; further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors thank Claudia Oliveira for helping with greenhouse assays.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Barnes, M.A.; Peeper, T.F.; Epplin, F.M.; Krenzer, E.G. Effects of Herbicides on Italian Ryegrass (Lolium multiflorum), Forage Production, and Economic Returns from Dual-Purpose Winter Wheat (Triticum aestivum). Weed Technol. 2001, 15, 264–270. [Google Scholar] [CrossRef]
- Salas, R.A.; Dayan, F.E.; Pan, Z.; Watson, S.B.; Dickson, J.W.; Scott, R.C.; Burgos, N.R. EPSPS Gene Amplification in Glyphosate-resistant Italian Ryegrass (Lolium perenne Ssp. Multiflorum) from Arkansas. Pest Manag. Sci. 2012, 68, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Bararpour, M.T.; Norsworthy, J.K.; Burgos, N.R.; Korres, N.E.; Gbur, E.E. Identification and Biological Characteristics of Ryegrass (Lolium Spp.) Accessions in Arkansas. Weed Sci. 2017, 65, 350–360. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S. Recurrent Selection with Reduced Herbicide Rates Results in the Rapid Evolution of Herbicide Resistance in Lolium rigidum. Theor. Appl. Genet. 2005, 110, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Délye, C. Weed Resistance to Acetyl Coenzyme A Carboxylase Inhibitors: An Update. Weed Sci. 2005, 53, 728–746. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Powles, S.B. Mutations of the ALS Gene Endowing Resistance to ALS-inhibiting Herbicides in Lolium rigidum Populations. Pest Manag. Sci. 2008, 64, 1229–1236. [Google Scholar] [CrossRef]
- Nikolskaya, T.; Zagnitko, O.; Tevzadze, G.; Haselkorn, R.; Gornicki, P. Herbicide Sensitivity Determinant of Wheat Plastid Acetyl-CoA Carboxylase Is Located in a 400-Amino Acid Fragment of the Carboxyltransferase Domain. Proc. Natl. Acad. Sci. USA 1999, 96, 14647–14651. [Google Scholar] [CrossRef]
- Raghav, N.; Singh, R.; Chhokar, R.S.; Sharma, D.; Kumar, R. Mutations in the Plastidic ACCase Gene Endowing Resistance to ACCase-Inhibiting Herbicide in Phalaris minor Populations from India. 3 Biotech 2016, 6, 12. [Google Scholar] [CrossRef]
- Takano, H.K.; Ovejero, R.F.L.; Belchior, G.G.; Maymone, G.P.L.; Dayan, F.E. ACCase-Inhibiting Herbicides: Mechanism of Action, Resistance Evolution and Stewardship. Sci. Agric. 2021, 78, e20190102. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to Acetyl-CoA Carboxylase-inhibiting Herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef]
- Herbert, D.; Walker, K.A.; Price, L.J.; Cole, D.J.; Pallett, K.E.; Ridley, S.M.; Harwood, J.L. Acetyl-CoA Carboxylase—A Graminicide Target Site. Pestic. Sci. 1997, 50, 67–71. [Google Scholar] [CrossRef]
- Rendina, A.R.; Craig-Kennard, A.C.; Beaudoin, J.D.; Breen, M.K. Inhibition of Acetyl-Coenzyme A Carboxylase by Two Classes of Grass-Selective Herbicides. J. Agric. Food Chem. 1990, 38, 1282–1287. [Google Scholar] [CrossRef]
- Devine, M.D. Acetyl-CoA Carboxylase Inhibitors. In Herbicide Classes in Development; Böger, P., Wakabayashi, K., Hirai, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 103–113. ISBN 978-3-642-63972-2. [Google Scholar]
- Xia, X.; Tang, W.; He, S.; Kang, J.; Ma, H.; Li, J. Mechanism of Metamifop Inhibition of the Carboxyltransferase Domain of Acetyl-Coenzyme A Carboxylase in Echinochloa crus-galli. Sci. Rep. 2016, 6, 34066. [Google Scholar] [CrossRef] [PubMed]
- Heap, I.M. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 14 May 2024).
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of Evolved Herbicide Resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Zhang, X.-Q.; Powles, S.B. The Molecular Bases for Resistance to Acetyl Co-Enzyme A Carboxylase (ACCase) Inhibiting Herbicides in Two Target-Based Resistant Biotypes of Annual Ryegrass (Lolium rigidum). Planta 2006, 223, 550–557. [Google Scholar] [CrossRef]
- Yu, Q.; Collavo, A.; Zheng, M.-Q.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of Acetyl-Coenzyme A Carboxylase Mutations in Resistant Lolium Populations: Evaluation Using Clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef]
- Kaundun, S.S. An Aspartate to Glycine Change in the Carboxyl Transferase Domain of Acetyl CoA Carboxylase and Non-target-site Mechanism(s) Confer Resistance to ACCase Inhibitor Herbicides in a Lolium multiflorum Population. Pest Manag. Sci. 2010, 66, 1249–1256. [Google Scholar] [CrossRef]
- Damalas, C.A.; Koutroubas, S.D. Herbicide-resistant Barnyardgrass (Echinochloa crus-galli) in Global Rice Production. Weed Biol. Manag. 2023, 23, 23–33. [Google Scholar] [CrossRef]
- Iwakami, S.; Kamidate, Y.; Yamaguchi, T.; Ishizaka, M.; Endo, M.; Suda, H.; Nagai, K.; Sunohara, Y.; Toki, S.; Uchino, A.; et al. CYP 81A P450s Are Involved in Concomitant Cross-resistance to Acetolactate Synthase and acetyl-CoA Carboxylase Herbicides in Echinochloa phyllopogon. New Phytol. 2019, 221, 2112–2122. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S. Metabolism-Based Herbicide Resistance and Cross-Resistance in Crop Weeds: A Threat to Herbicide Sustainability and Global Crop Production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef]
- Délye, C.; Michel, S. ‘Universal’ Primers for PCR-sequencing of Grass Chloroplastic acetyl-CoA Carboxylase Domains Involved in Resistance to Herbicides. Weed Res. 2005, 45, 323–330. [Google Scholar] [CrossRef]
- Délye, C.; Matéjicek, A.; Gasquez, J. PCR-based Detection of Resistance to acetyl-CoA Carboxylase-inhibiting Herbicides in Black-grass (Alopecurus myosuroides Huds) and Ryegrass (Lolium rigidum Gaud). Pest Manag. Sci. 2002, 58, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A Unified Platform for Automated Protein Structure and Function Prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein Structure and Function Prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. I-TASSER Server for Protein 3D Structure Prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef]
- Xue, Q.; Liu, X.; Russell, P.; Li, J.; Pan, W.; Fu, J.; Zhang, A. Evaluation of the Binding Performance of Flavonoids to Estrogen Receptor Alpha by Autodock, Autodock Vina and Surflex-Dock. Ecotoxicol. Environ. Saf. 2022, 233, 113323. [Google Scholar] [CrossRef]
- Salas, R.A.; Burgos, N.R.; Mauromoustakos, A.; Lassiter, R.B.; Scott, R.C.; Alcober, E.A. Resistance to ACCase and ALS Inhibitors in Lolium perenne Ssp. multiflorum in the United States. J. Crop Weed 2013, 9, 168–183. [Google Scholar]
- Jang, S.; Marjanovic, J.; Gornicki, P. Resistance to Herbicides Caused by Single Amino Acid Mutations in Acetyl-CoA Carboxylase in Resistant Populations of Grassy Weeds. New Phytol. 2013, 197, 1110–1116. [Google Scholar] [CrossRef]
- Singh, V.; Maity, A.; Abugho, S.; Swart, J.; Drake, D.; Bagavathiannan, M. Multiple Herbicide–Resistant Lolium Spp. Is Prevalent in Wheat Production in Texas Blacklands. Weed Technol. 2020, 34, 652–660. [Google Scholar] [CrossRef]
- Rauch, T.A.; Thill, D.C.; Gersdorf, S.A.; Price, W.J. Widespread Occurrence of Herbicide-Resistant Italian Ryegrass (Lolium multiflorum) in Northern Idaho and Eastern Washington. Weed Technol. 2010, 24, 281–288. [Google Scholar] [CrossRef]
- Buddenhagen, C.E.; James, T.K.; Ngow, Z.; Hackell, D.L.; Rolston, M.P.; Chynoweth, R.J.; Gunnarsson, M.; Li, F.; Harrington, K.C.; Ghanizadeh, H. Resistance to Post-Emergent Herbicides Is Becoming Common for Grass Weeds on New Zealand Wheat and Barley Farms. PLoS ONE 2021, 16, e0258685. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.P.C.; Kim, Y.S.; Tong, L. Mechanism for the Inhibition of the Carboxyltransferase Domain of Acetyl-Coenzyme A Carboxylase by Pinoxaden. Proc. Natl. Acad. Sci. USA 2010, 107, 22072–22077. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Peng, Y.; Chen, W.; Yu, Q.; Bai, L.; Pan, L. The ILE-2041-VAL Mutation in the ACCase Gene Confers Resistance to Clodinafop-propargyl in American Sloughgrass (Beckmannia syzigachne Steud). Pest Manag. Sci. 2021, 77, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Ghanizadeh, H.; Harrington, K.C.; Mesarich, C.H. The Target Site Mutation Ile-2041-Asn Is Associated with Resistance to ACCase-Inhibiting Herbicides in Lolium multiflorum. New Zealand J. Agric. Res. 2020, 63, 416–429. [Google Scholar] [CrossRef]
- Vázquez-García, J.G.; Alcántara-de La Cruz, R.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Cruz-Hipólito, H.E.; Torra, J.; Barro, F.; De Prado, R. Accumulation of Target Gene Mutations Confers Multiple Resistance to ALS, ACCase, and EPSPS Inhibitors in Lolium Species in Chile. Front. Plant Sci. 2020, 11, 553948. [Google Scholar] [CrossRef]
- Saini, R.K.; Malone, J.; Preston, C.; Gill, G. Target Enzyme-Based Resistance to Clethodim in Lolium rigidum Populations in Australia. Weed Sci. 2015, 63, 946–953. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Hutchings, S.-J.; Dale, R.P.; McIndoe, E. Broad Resistance to ACCase Inhibiting Herbicides in a Ryegrass Population Is Due Only to a Cysteine to Arginine Mutation in the Target Enzyme. PLoS ONE 2012, 7, e39759. [Google Scholar] [CrossRef]
- Gaines, T.A.; Patterson, E.L.; Neve, P. Molecular Mechanisms of Adaptive Evolution Revealed by Global Selection for Glyphosate Resistance. New Phytol. 2019, 223, 1770–1775. [Google Scholar] [CrossRef]
- Neve, P.; Vila-Aiub, M.; Roux, F. Evolutionary-thinking in Agricultural Weed Management. New Phytol. 2009, 184, 783–793. [Google Scholar] [CrossRef]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the Evolution of Herbicide Resistance in Weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Resistance to AHAS Inhibitor Herbicides: Current Understanding. Pest Manag. Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).