Abstract
A substrate-based production system is a simple and low-cost method for arbuscular mycorrhizal (AM) fungal inoculum production. However, it is time-consuming and typically yields low numbers of AM fungal spores due to several factors affecting plant growth efficiency. Our study investigated the use of light-emitting diode (LED) technology to expedite AM fungal spore production in planta. We performed experiments with Rhizophagus irregularis inoculated in maize (Zea mays L.), contrasting LED light with greenhouse (GH) conditions. Our results exhibited a significant improvement in AM fungal colonization and spore production, as well as a reduction in the production period from 120 to 90 days under the LED light condition. This was achieved using a red-and-blue light ratio of 60:40 with a total light intensity of 300 µmol m−2 s−1. The LED light treatments improved maize growth by increasing nitrogen (N) and phosphorus (P) concentrations in shoots and roots, respectively. Our gene expression analyses revealed that in AMF-inoculated plants, genes related to photosynthesis were significantly upregulated under LED light compared to the GH condition. Moreover, LED increased the expression of marker genes linked to the AM fungi-related cell cycle, indicating enhanced AM fungal growth during symbiosis. These findings advance our comprehension of LED applications in agriculture, offering promising prospects for acceleration of AM fungal spore production.
1. Introduction
Arbuscular mycorrhizal (AM) fungi are a type of soil fungi that form mutually beneficial relationships with most plant roots [1,2]. These fungi play a critical role in plant growth and ecosystem functioning, especially under phosphorus-limited conditions [3,4,5]. Consequently, AM fungi are among the most effective biofertilizers worldwide. In recent years, two main techniques have been used for AM fungal inoculum production: in vitro, which involves controlled lab cultivation, and in vivo, which relies on live host plants, each with distinct benefits and challenges [6]. Commercial AM fungal inoculants are typically produced using in vitro methods to ensure product purity. However, these high-quality AM fungal inoculants are quite costly and often inaccessible to farmers in developing countries. Consequently, a substrate-based production system might provide a more promising and affordable solution for large-scale AM fungal inoculum production in these countries. This type of inoculum production system relies on the host plant, where AM fungi are grown in a substrate like vermiculite or sand and spores are collected from the substrate to be used as an inoculum [7,8]. To date, substrate-based production systems utilizing organic waste have been implemented in AMF production technology. These systems include associated regulatory mechanisms to ensure both the quality and cost-effectiveness of the end product [9]. In addition, the use of saline soil is a cost-effective alternative for producing AMF inoculum with suitable plant hosts [10]. The efficiency of AM fungal spore production within a substrate-based system can depend on plant compatibility and overall growth efficiency, which can influence AM fungal colonization and propagation [11,12,13]. The production of AM fungal biofertilizers in this system necessitates the presence of actively growing plant roots to complete the AM fungal life cycle. However, this method can be time-consuming and often results in a low yield of AM fungal spore production, typically taking up to 120 days using traditional methods [14,15]. The promotion of host plant growth has been identified as a key factor in enhancing AM fungal spore production and potentially reducing time consumption in such a production system.
Light is one of the most important factors affecting plant growth and development [16]. In recent years, the use of light-emitting diodes (LEDs) to enhance the growth and development of various plant species has piqued interest. LEDs are highly effective in promoting plant growth and development [17,18,19]. Red and blue LED lights, in particular, have been extensively studied due to their capability to stimulate various physiological processes in plants. Red light, essential for photosynthesis, is particularly effective in promoting plant growth and flowering, whereas blue light regulates plant growth and development, including stem elongation, leaf expansion, and chloroplast formation [20,21,22]. The use of light to stimulate biological processes is a well-established practice in many fields, including agriculture and horticulture [23,24]. LED lighting systems possess numerous advantages over traditional lighting methods, including increased energy efficiency, reduced heat production, and the ability to customize the spectral output. This versatility allows the lighting to be tailored to meet specific plant needs, such as promoting photosynthesis, influencing flowering, and enhancing nutrient uptake [25,26].
LEDs have emerged as a transformative tool for enhancing the growth and development of AM fungi, particularly in terms of increasing spore production. Light supplied jointly by red and blue LEDs, with wavelengths of 630 nm and 450 nm, respectively, has proven highly effective in stimulating spore production in Gigaspora margarita within bahia grass [27]. While the mechanism by which LED lights stimulate AM fungal spore production is not thoroughly explained, the observation suggests that LEDs might be exploited to stimulate spore production in other host plants. C4 plants such as maize (Zea mays L.) and sorghum (Sorghum bicolor (L.) Moench) are frequently used for the production of AM fungal inoculum under a substrate-based system due to their high photosynthetic efficiency and resilience to environmental stress [28]. Furthermore, maize is readily available in the local market. In this study, maize was employed as the host plant for AM fungal inoculum production.
Therefore, this study aimed to evaluate the potential advantages of employing LED lights to enhance the spore production of Rhizophagus irregularis in maize under a substrate-based system. Additionally, it proposed a mechanism for AM fungal spore production enhancement, investigating plant growth, nutrients, and the expression of certain plant and fungal genes involved in metabolism and plant–AM fungus symbiosis in response to LED lights. Thus, this study explored the synergistic impact of red and blue LED lights at a specific ratio, revealing their ability to not only promote maize growth but also enhance AM fungal colonization. This innovative LED-based approach also reduces the time required for AM fungal inoculum production.
2. Materials and Methods
2.1. Plant Growth Condition
Maize (Zea mays L.) cultivar Suwan-5 seeds underwent surface sterilization using a 95% ethanol immersion for 30 s, followed by a 12 min treatment with a 3% (v/v) sodium hypochlorite (NaOCl) (Loba Chemie Pvt. Ltd., Maharashtra, India) solution. Sterilized seeds were then rinsed thoroughly six times with sterilized water and allowed to soak overnight in sterilized water in an incubator shaker operating at 50 rpm. After this soaking period, the seeds were once again rinsed with sterile water, deposited on sterilized paper towels, and stored in darkness for 2 days. Each pot was then prepared with 0.5 kg of sterilized river sand with particle sizes from 0.2 to 5 mm and used for planting 1 plant/pot. The experiment was conducted at Suranaree University of Technology, Nakhon Ratchasima, Thailand (14°52021.400, E:102°1020.200) during February–April, 2023. For greenhouse (GH) condition, plants were cultivated under natural light conditions, with an average temperature of 34.5 ± 0.16 °C and light intensity of 900–1100 µmol m−2 s−1 during daytime. For LED light condition, plants were cultivated with a 16/8 h light/dark cycle under standard ambient temperatures for the LED setup, with an average temperature of 30.8 ± 0.21 °C and a light intensity of 300 µmol m−2 s−1. The plants were watered with 200 mL of a half-strength Hoagland’s solution containing a low concentration of KH2PO4 (100 µM) every two days [29]. The half-strength Hoagland’s solution composition used in this study is described in Table S1. Each experiment included five replications.
For each experiment with AM fungal inoculation, a total of 250 spores of Rhizophagus irregularis DAOM197198 (Mycorise, Premier Tech, Rivi-ère-du-Loup, QC, Canada) were suspended in 5 mL of distilled water and inoculated at the center of each pot around the root zone using a 5 mL pipette. The inoculation site was then covered with sand.
2.2. Optimizing the Conditions of Light-Emitting Diodes (LEDs) for Promoting Maize Growth and Fungal Abundance
(i) To optimize the conditions of light-emitting diodes (LEDs) for promoting maize growth, transplanted maize without AM fungal inoculation was exposed to varying LED light conditions. The intensity of red and blue lights was measured using a LIGHTSCOUT quantum light meter (Spectrum Technologies, Inc., Aurora, IL, USA). The ratio of red light to blue light (R:B) was adjusted across four conditions: 80:20, 60:40, 40:60, and 20:80. All treatments were carried out under a consistent light intensity of 300 µmol m−2 s−1. Measurements of plant height, chlorophyll content (using a SPAD502 Plus spectrophotometer) (Konica Minolta Sensing Singapore Pte. Ltd., Crescent, Singapore), and biomass were obtained 45 days after inoculation (DAI). Shoots and roots were separated and dried at 70 °C for 48 h before their respective dry weights were recorded [30].
(ii) Following these assessments, the optimal light ratio was used to examine the effects of various LED light intensities at 200, 300, and 400 µmol m−2 s−1 on enhancing AM fungal spore production. To determine the LED light intensity that affected fungal abundance in maize root, AM fungus was inoculated into the plants as described above.
2.3. Assessment of AM Fungal Colonization
The harvested roots underwent a meticulous preparation process to eliminate pigments. Initially, these roots were submerged in a 10% (w/v) potassium hydroxide (KOH) solution and boiled at 90 °C for 10 min, which was followed by multiple rinses with sterile water. Afterward, the roots were acidified using a 2% (v/v) HCl solution. After acidification, the cleared roots were stained using a 0.05% trypan blue solution in lactic acid at 90 °C for 10 min. After the staining process, the roots were thoroughly rinsed to eliminate any excess stain and were then mounted on microscope slides in a lactoglycerol solution [31]. The extent of AM fungal colonization within the roots was viewed using a light microscope at 10× magnification. The percent of root length colonized by AM fungi was quantified using a standard scoring method [32]. Each parameter was calculated using the following equations:
- 1.
- Frequency of mycorrhiza in the root system (F%)
F% = (number (nb) of fragments myco/total nb) * 100
- 2.
- Intensity of mycorrhizal colonization in the root system (M%)
M% = (95n5 + 70n4 + 30n3 + 5n2 + n1)/(nb total)
- 3.
- Intensity of mycorrhizal colonization in mycorrhizal root fragments (m%)
m% = M*(nb total)/(nb myco),
- 4.
- Arbuscule abundance in mycorrhizal root fragments (a%)
a% = (100mA3 + 50mA2 + 10mA1)/100
mA3 = ((95n5A3 + 70n4A3 + 30n3A3 + 5n2A3 + n1A3)/nb myco) * 100/m
- 5.
- Arbuscule Abundance in the Entire Root System (A%)
A% = a*(M/100)
2.4. Isolation of R. irregularis Spores
After transplanting, maize plants were maintained with regular watering until 75 DAIs, at which point watering was discontinued. The plants were provided with LED lighting up to 90 DAIs. AMF (arbuscular mycorrhizal fungi) spores were collected from each soil sample using the methods of wet sieving and decanting [33,34], adhering to the isolation procedure described by Kiddee et al. [35]. The AMF spores were subsequently counted using a stereomicroscope.
2.5. Gene References and Primer Design
We conducted a thorough investigation into the genes associated with plant photosynthesis, using the maize reference genome Zm-B73 (GCF_902167145.1) obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_902167145.1/) (accessed on 20 February 2023). We selected genes relevant to photosynthesis and plant–AM fungus symbiosis for analysis. We designed primers to determine the expression of these genes in maize and R. irregularis, both of which have reportedly participated in plant–AM fungus symbiosis [36]. Using Clone Manager 9 Professional Edition, we carried out the primer design process. The primers used for the amplification of photosynthetic genes, the AM-induced genes in maize, and the symbiosis marker genes for R. irregularis are outlined in Tables S2 and Table S3, and S4, respectively.
2.6. RNA Extraction and qRT-PCR
RNA extraction was conducted using the RNA prep Pure Plant Plus Kit (TIANGEN Biotech, Beijing, China). The RNA samples were treated with DNase I solution (Thermo Fisher Scientific, Waltham, MA, USA) to eliminate any DNA contamination. Following the manufacturer’s guidelines, cDNA synthesis was carried out using the High-Capacity cDNA Reverse Transcription Kit (iScript, Bio-Rad, Hercules, CA, USA). The CFX Opus 96 Real-Time PCR System (Bio-Rad, USA) with Luna Universal qPCR Master Mix (NEB, Biolabs Inc., Durham, NC, USA) was used for quantitative real-time PCR (qRT-PCR) analysis. Z. mays L. 18S rRNA gene was employed as an internal control for the normalization of gene expression in chloroplast-encoded photosynthetic genes, while beta-actin (ZmACT1) [37] was used for AM-induced gene expression in maize. For transcript normalization in R. irregularis, the translation elongation factor 1 α (RiTEF) served as the reference gene. To validate gene expression levels, three biological replicates were performed, each using approximately 10 ng of cDNA template per reaction. Quantitative real-time PCR (qRT-PCR) was conducted using a SYBR Green detection system to ensure accuracy in detecting transcript levels. The relative expression levels of the target genes were calculated using the 2−ΔΔCT method, with each sample normalized to its respective internal control [38].
2.7. Statistical Analyses
Statistical analyses were conducted using R software (version 4.2.1). Variations in plant physiological parameters (height and chlorophyll content), dry biomass, and AM fungal abundance under different LED light ratios were assessed using Tukey’s HSD test for multiple comparisons. One-way ANOVA and Tukey’s post hoc comparisons were employed to compare AM fungal spore numbers across all groups under LED light and GH conditions, within both non-AMF and AMF inoculations. Student’s t-test was used to compare the two groups under different LED light and greenhouse (GH) conditions. Data were visualized using GraphPad Prism (version 9.1.1).
3. Results
3.1. Optimum Blue-and-Red LED Light Ratio Expedited Maize Seedling Growth and Increased Biomass
In this study, we explored the optimal ratios of red light to blue (R:B) light for promoting maize growth during the seedling stage at 7 and 14 days (Figure 1A,B, Table 1). The results suggest that using R:B ratios of 60:40 and 80:20 can enhance maize height in comparison to ratios of 20:80 and 40:60. Contrarily, a higher blue-light-to-red-light ratio enhanced the chlorophyll content at 7 days (Table 1). Furthermore, a higher red-light-to-blue-light ratio (80:20) significantly impacted maize plant height compared to other treatments, but induced a lower chlorophyll content at 14 days. Additionally, the red-to-blue light ratios of 60:40 and 80:20 significantly elevated shoot dry weight compared to ratios of 20:80 and 40:60. Furthermore, treatments with a higher proportion of red light also demonstrated amplified root dry weight compared to those with higher blue light ratios. Consequently, the R:B ratio of 60:40 was selected for further light intensity optimization to promote fungal abundance.
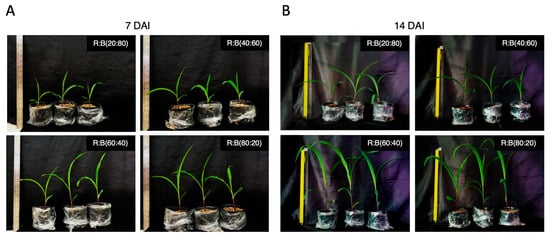
Figure 1.
Evaluating the impact of red–blue (R:B) light ratios on the growth of maize (Zea mays L.) cultivar Suawan-5 at (A) 7 and (B) 14 days after exposure (DAE), with a total light intensity of 300 µmol m−2 s−1.

Table 1.
The influence of various red–blue (R:B) light ratios on the physiological responses of Suwan 5 maize after 7 and 14 days of exposure (DAE). Four distinct treatments were implemented, with different combinations of red-and-blue light ratios at an approximate total intensity of 300 µmol m−2 s−1.
3.2. LED Light Intensity Affected Fungal Abundance in Maize Root
In this study, we used the chosen R:B ratio to examine the effect of LED light at various total intensities of 200, 300, and 400 µmol m−2 s−1, termed RB200, RB300, and RB400, respectively, on fungal abundance. In particular, we focused on the colonization and spore production of R. irregularis when compared to a GH condition. We observed that at 45 DAIs, the application of LED light treatments at varying intensities significantly increased AM fungal infection levels in maize roots compared to the GH condition (Supplementary Table S5). Additionally, AM fungal colonization exhibited an increase at 45 DAIs (Figure 2). Notably, our observations pointed out an increased vesicle density on maize roots with the application of LED light, especially at a total light intensity of 300 µmol m−2 s−1 (RB300) (Figure 2A). While no significant difference in average vesicle size was observed, RB300 had the highest average vesicle size of 78 µm, compared to 71, 58, and 66 µm under GH, RB200, and RB400, respectively (Figure 2B). Interestingly, LED light use not only promoted AM fungal infection but also significantly increased AM fungal spore counts in the sand, reaching up to 228 spores per plant at an intensity of 300 µmol m−2 s−1 (Figure 2C). Based on these findings, the specific LED light intensity of 300 µmol m−2 s−1 was determined as the optimal condition for promoting the development of AM fungal spores in maize roots.
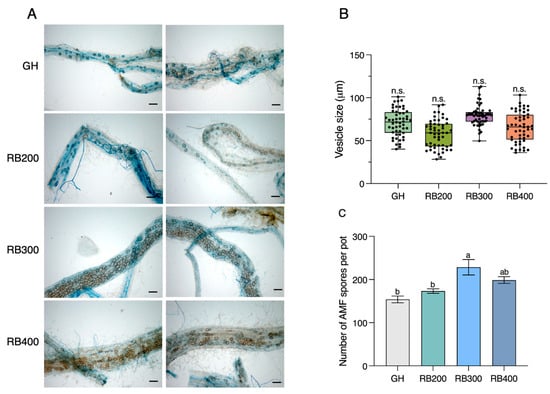
Figure 2.
Impact of LED light on AM fungal colonization, vesicle size, and spore number of R. irregularis in maize roots at 45 DAIs. (A) The AM fungal abundance in roots of maize grown under greenhouse (GH) condition and red–blue (R:B) light ratio of 60:40 at different light intensities of 200, 300, and 400 µmol m−2 s−1 conditions, with scale bars of 100 μm. (B) The average vesicle size, with the vesicles (n = 52) from 10 root fragments randomly measured for each treatment, and (C) the number of AM fungal spores per pot. Values are represented as means ± s.e.m (n = 3). Bars with different letters indicate significant differences at a p-value < 0.05 based on Tukey’s HSD test. n.s., not significant.
3.3. The Optimized LED Condition Enhanced Nutrient Accumulation, AM Fungal Colonization, and Spore Production in Maize Better than the GH Condition
Once the LED condition had been optimized, the experiment was conducted again for comparison with the GH condition. We collected data on plant physiological parameters to investigate the correlation between plant growth and AM fungal abundance over a shorter period than usual. This was aimed at accelerating AM fungal spore production under specific LED light intensities after 45 and 90 days, instead of 120 days. Our findings revealed that AM fungal inoculation under LED light (LED + AMF) at 45 days provided the optimal plant height and chlorophyll content (Figure 3). Without AM fungal inoculation, a significant difference was observed in plant height, but not in chlorophyll content, under the LED light condition when compared with the GH condition (Figure 3B,C). However, both plant height and chlorophyll content were significantly higher under LED light compared to the GH condition following inoculation with AM fungus (Figure 3B,C). Furthermore, the LED + AMF treatment demonstrated a significant increase in shoot and root dry weights when compared with other treatments (Figure 4A). Conversely, a lower dry biomass was observed in the non-AMF treatment under the GH condition, while the non-AMF treatment under LED light did not differ from the GH + AMF treatment. In addition, we analyzed phosphorus (P) and nitrogen (N) concentrations, which are essential nutrients transferred during AM symbiosis with maize roots. At 45 days, root P nutrition in the LED + AMF treatment showed a significant increase compared to the GH + AMF treatment (Figure 4B). However, there were no significant differences observed in the N concentration in both the shoot and root of the maize plants.
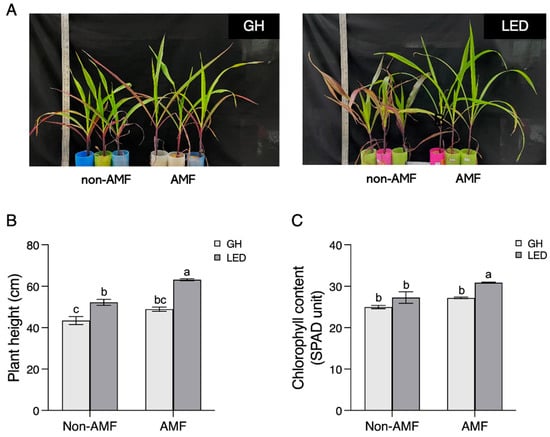
Figure 3.
Plant physiology of Suwan-5 maize, showing growth at 45 days after inoculation (DAIs) under greenhouse (GH) and LED light conditions. (A) Plants were grown without R. irregularis (non-AMF) and with R. irregularis (AMF) inoculation. (B) Plant height and (C) chlorophyll content of maize were measured. Significant differences among all groups, including LED light and GH conditions within both non-AMF and AMF inoculations, were analyzed using one-way ANOVA. Standard error bars (s.e.m.) were calculated with n = 3. Groups that share the same letter show no significant difference, as determined by Tukey’s post hoc test at p < 0.05.
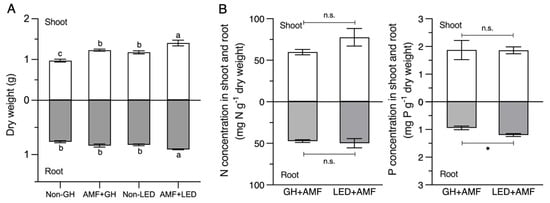
Figure 4.
The data of (A) dry biomass and (B) plant tissue nitrogen (N) and phosphorus (P) concentrations at 45 days after inoculation (DAIs). Average concentrations of nitrogen (N) and phosphorus (P) (mg g−1 dry weight) in the shoot and root of Suwan-5 maize are presented with standard error bars (± s.e.m; n = 3). The analysis of shoot and root biomass was conducted separately, and different letters indicate significant differences based on Tukey’s HSD test. Student’s t-test was performed to analyze nitrogen (N) and phosphorus (P) concentrations (n.s., not significant; *, p < 0.05).
The fungal abundance, in terms of colonization, demonstrated a significant increase in the AM fungal colonization frequency (F%), colonization intensity (M%), and arbuscule abundance (A%) in the root system under LED light treatment, compared to that under GH condition at 45 DAIs. Contrastingly, the colonization intensity (m%) and arbuscule abundance (a%) in colonized roots did not reveal any substantial differences (Figure 5A). Interestingly, the quantification of AM fungal spores in the sand at 45 DAIs increased by up to 1.5 times under the LED light treatment compared to the GH condition (Figure 5B). The number of AM fungal spores peaked at 1500 under LED light at 90 DAIs, whereas fewer than 1000 spores were produced under the GH condition (Figure 5C). This result suggests that LED light conditions could potentially stimulate AM fungal development and spore production in the host plant, potentially accelerating the process of spore production in maize to 90 days compared to the traditional 120 days.
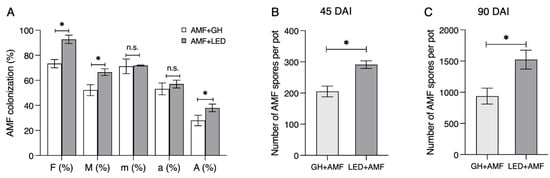
Figure 5.
The abundance of R. irregularis at 45 days after inoculation (DAIs). Maize was inoculated with R. irregularis (AMF) under the greenhouse (GH) and LED light conditions. (A) AMF colonization in maize roots. F(%), the frequency of mycorrhiza in the root system; M(%), the intensity of mycorrhizal colonization in the root system; m(%), the intensity of mycorrhizal colonization in the root fragments; a(%), the arbuscule abundance in the mycorrhizal parts of the root fragments; and A(%), the arbuscule abundance in the root system. (B) The number of R. irregularis spores produced in the soil at 45 DAIs and (C) 90 DAIs. Values are represented as means ± s.e.m. (n = 3). Student’s t-test (n.s., not significant; * p < 0.05) was conducted in (A–C).
3.4. Gene Expression in Maize and R. irregularis during Symbiosis in Response to Growth Treatment under LED Light and GH Conditions
We examined alterations in gene expression, focusing on photosynthesis-related plant genes and specific marker genes involved in the symbiotic response between maize and R. irregularis. We collected data from the root systems at 45 DAIs. The transcript amounts of genes linked to plant photosynthesis comprise intricate parts of the photosynthetic electron transport (PET) process. These include the RuBisCO (large subunit) rbcL, as well as psaA and psbA, which are essential components of photosystems I and II in the electron transport chain, respectively. Remarkably, petA and petD serve as elements of the cytochrome b6f complex subunit IV. In addition, atpB and atpE are fundamental parts of the ATPase subunits β and ε, located within the stromal region, overseeing ATP production. All these genes demonstrated considerable upregulation, showing a roughly 1.5- to 40-fold increase, under the LED light condition compared to the GH condition (Figure 6).
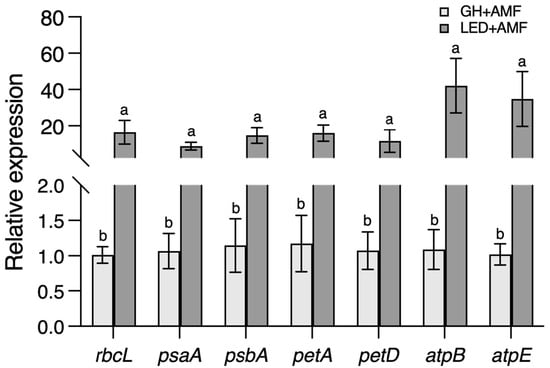
Figure 6.
The expression of maize photosynthesis-related genes at 45 days after inoculation (DAIs). Chloroplast-encoded photosynthetic genes: rbcL: RuBisCO (large subunit); psaA and psbA: electron transport (photosystems I and II); petA and petD: electron transport (cytochrome ƒ and subunit IV of cytochrome b6f complex); and atpB and atpE: ATP synthesis. There are three replications, and the letter above the bar indicates significant difference at p-value < 0.05 using Student’s t-test.
The transcript levels of five genes induced by R. irregularis in maize roots were also determined. The expression of the transcription factor gene RAM1, primarily responsible for arbuscule formation, showed a significant change (Figure 7A). Similarly, the phosphate transporter Pht6 and ammonium transporter AMT3, involved in P and N uptake in arbuscular-containing cells within specialized tree-like structures in cortical cells, exhibited no significant differences between the LED light and GH conditions. However, the expression of the glycerol-3-phosphate acyltransferase gene RAM2, which is involved in AM fungal colonization, increased 2-fold under LED light compared to the GH condition. Meanwhile, the expression of the acyl-ACP thioesterase FatM showed no significant difference.
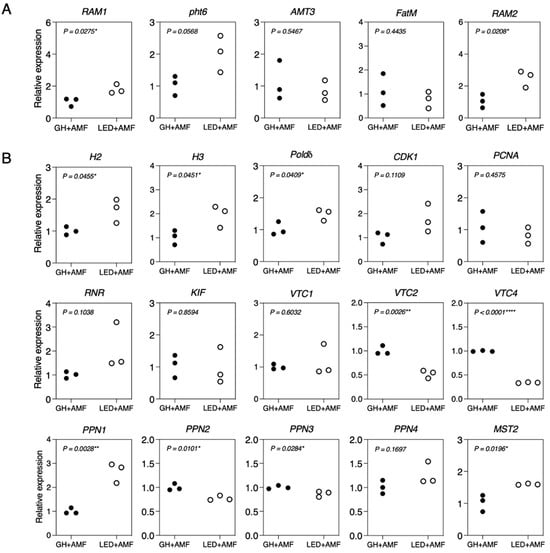
Figure 7.
The gene expression profiles of maize (A) and R. irregularis (B) at 45 days after inoculation (DAIs). Plants were inoculated with R. irregularis (AMF) and grown under the greenhouse (GH) and LED light conditions. P-values were calculated based on Student’s t-test (*, p < 0.05; **, p < 0.01; ****, p < 0.0001). (A) maize genes, including RAM1: GRAS transcription factor; Pht6: phosphate transporter; AMT3: ammonium transporter; FatM: acyl–acyl carrier protein thioesterase; and RAM2: glycerol-3-phosphate acyltransferase. (B) R. irregularis genes, including H2 and H3: histones; Polδ: DNA polymerase delta subunit 4; CDK1: cyclin-dependent kinase; PCNA: proliferating cell nuclear antigen; RNR: ribonucleotide reductase; KIF: kinesin; PPN1-PPN4: endopolyphosphatases; VTC1, VTC2, and VTC4: vacuolar transporter chaperones; and MST2: monosaccharide transporter.
Moreover, we scrutinized the transcript levels of cell cycle-related genes, including H2, H3, Polδ, CDK1, PCNA, RNR, and KIF, and their response to alterations in phosphorus (P) and carbon (C) nutrition during the symbiotic interaction between R. irregularis and maize. The results indicated that out of these seven genes, three (H2, H3, and Polδ) exhibited significant upregulation in gene expression under LED light compared to the GH condition (Figure 7B). We focused on the transcript levels in R. irregularis in response to varying P levels, especially those concerning polyphosphate synthesis and degradation processes. Genes responsible for the synthesis of polyphosphate enzymes, namely VTC1, VTC2, and VTC4 [39], exhibited downregulation under the LED light conditions. On the contrary, the gene tied to endophosphatases, PPN1, exhibited strong upregulation in response to the LED light conditions, whereas PPN2, PPN3, and PPN4 all showed downregulation. We also noted an increase in the expression of the monosaccharide transporter gene MST2, which has a crucial role in sugar uptake in R. irregularis, under LED light compared to the GH condition.
4. Discussion
The production of arbuscular mycorrhizal fungal (AMF) inoculum is essential for various agricultural and ecological applications. Recently, two primary techniques for AMF inoculum production are in vitro and in vivo methods [6]. In vitro systems, while offering precise control over the growth condition and high-density spore production, often involve higher costs due to the need for specialized equipment and media. This can be a significant barrier to their widespread adoption. Recent research has mainly explored various substrate-based methods for AMF inoculum production, using different substrates such as sand, saline soil, and agro-waste [9,10,28]. However, there is less research available on the use of LED light in these systems, which could potentially offer new insights and improvements in AMF production efficiency.
Plants need light for photosynthesis, which is the process by which they transform light energy into chemical energy to stimulate their growth and development. Different wavelengths of light influence plant growth, development, and biomass in varying ways. Red and blue lights play imperative roles in plant growth [40,41,42]. LED light is commonly used to provide light for plants during periods or seasons when natural light intensity is low to prevent chlorosis or impaired leaf development [43,44]. Additionally, LED light has been utilized to stimulate plant growth when provided with optimal light conditions [45,46].
In our study, we specifically optimized the red-and-blue light ratio and light intensity to support maize plant growth and promote AM fungal abundance. The ratios of red and blue lights had a significant impact on maize growth in the early stage, whereas light intensity influenced AM fungal spore production (Figure 1 and Figure 2, Table 1). These findings suggest that a specific LED light ratio and light intensity are required for each plant and for each specific purpose of stimulating AM fungal abundance.
In this case, although blue light regulates plant growth and development, including stem elongation, leaf expansion, and the formation of chloroplasts [20,21,22], higher ratios of blue light to red light promoted chlorophyll content but did not promote maize shoot and root development. Conversely, higher ratios of red light to blue light were highly effective in promoting maize growth, particularly the root biomass, which is where AM fungi colonize and potentially supports AM fungal abundance. As a result, a red-to-blue light ratio of 60:40 was selected to stimulate the growth of maize as a host plant for AM fungal spore production.
Nevertheless, the ratio of red and blue LED lights supplied should vary depending on the plant species [30,47,48]. Therefore, the specific light ratio should be examined for each host plant used for AM fungal propagation. For instance, an optimal blue-and-red light ratio of 25:75 improved leaf photosynthetic capacity, net photosynthetic rate, stomatal conductance, and chlorophyll content in cucumbers [49], while a photon flux ratio of red and blue lights of 7:1 promoted growth in pepper seedlings [50].
Once a specific red-to-blue light ratio was selected, we determined the corresponding LED light intensity to better promote AM fungal spore production compared to GH conditions. Utilizing a red-to-blue LED light ratio of 60:40, with a total intensity of 300 µmol m−2 s−1, yielded the highest number of AM fungal spores, significantly surpassing the numbers observed under the GH conditions. Interestingly, lowering the light intensity to 200 µmol m−2 s−1 or raising it to 400 µmol m−2 s−1 resulted in a decrease in AM fungal colonization and spore numbers within sand (Figure 2). This demonstrates the potential of integrating specific LED light conditions to support plant growth and boost AM fungal inoculum production in maize. The improved plant growth (including height and shoot and root dry weights), higher chlorophyll content, and significantly increased AM fungal colonization and spore production at 45 and 90 DAIs under the LED light conditions (compared to the GH conditions) confirmed the possibility of promoting AM fungal inoculum production in shorter periods (reduced from 120 days to 90 days) using a substrate-based production system under LED lighting (Figure 3, Figure 4A and Figure 5). Previous reports suggest that providing red light followed by blue light encourages AM fungal colonization, while solely utilizing red light stimulates colonization and leads to an increase in spore production in bahia grass, especially in glass bead materials [27]. Similarly, the use of red and blue lights can stimulate the hyphal growth of G. margarita in bahia grass grown in a sand medium. However, applying only blue light was found to suppress AM fungal colonization and decrease spore numbers in bahia grass [27]. These observations indicate that light can have diverse effects on plant growth and subsequent AM fungal development. This is consistent with earlier studies suggesting that LED lighting can stimulate the proliferation of hyphal branches in G. margarita and Glomus spp. (R-10) [51], expand the root area, and thereby encourage AM fungal colonization, leading to the production of numerous AM fungal spores in the soil [27,52,53]. Nevertheless, future study is required to explore the performance of inoculum produced under LED conditions in different plant species to evaluate its effectiveness more comprehensively.
In an attempt to understand how maize reacts to LED lighting, we analyzed the expression levels of several marker genes related to plant photosynthesis. Light acts as a crucial regulator of photosynthetic gene expression, meaning that significant stimulation or reprogramming of gene expressions under LED lights may have an impact on various metabolic processes [53]. Applications of red and blue lights have been noted to increase plant nutrient accumulation, as well as acting as the primary sugar and fatty acid source for AM fungi during symbiosis.
Initially, we looked at genes linked to photosynthesis, which included those responsible for carbon fixation (e.g., rbcL), the electron transport chain (e.g., psaA, psbA, petA, and petD), and ATP synthesis (e.g., atpB and atpE). We studied their reaction to LED and GH conditions, both with and without AM fungal inoculation. The results showed that every tested gene responded more strongly to LED light stimulation compared to their expression under the GH conditions. The expression levels of these genes increased from 1.5 to 40 times under LED lights compared to the GH conditions, where natural sunlight was the principal light source (Figure 6). This considerable response to LED lighting suggests the potential for increased carbon flow, nutrient accumulation, and facilitation of transfers to AM fungi.
We also examined the accumulation of the nutrients P and N, which are integral to nutrient absorption by AM fungi during symbiosis [35,54], in maize’s shoot and root. Nutrient solutions with high concentrations of P and N have been shown to inhibit AMF colonization [55], whereas optimal levels promote colonization and enhance P and N accumulation in host plants. Our findings indicated no noticeable difference in N accumulation in both maize’s shoot and root at 45 DAIs. However, there was a significant increase in P accumulation in the root under LED light treatment, when compared to the GH conditions (Figure 4B). These results indicate that the use of LED lighting with specific light intensity could augment maize growth and development. This enhancement is due to the stimulation of photosynthetic processes. Additionally, the fungal partner enhances plant nutrient absorption, aiding the transfer of phosphorus and nitrogen to their host plants [56]. These outcomes highlight the effectiveness of LED lighting in promoting horticultural and field crop growth and biomass through the encouragement of photosynthesis [23,24,30].
To better understand how plants and AM fungi respond to LED light during symbiosis, we determined the expression of several plants and AM fungal marker genes involved in symbiosis. We discovered that enhancing AM fungal formation in maize under LED light corresponds with a significant induction of the plant marker gene, RAM1, during the symbiotic process (Figure 7A). Intriguingly, we did not observe significant differences in the expression of genes tied to plant nutrient uptake, such as N and P (specifically AMT3 and Pht6), under LED light conditions. However, we did note an uptick in the expression of genes involved in lipid transfer from the host to the AMF, notably RAM2. We also observed the expression of genes related to cell cycle, phosphate metabolism, and nutrient uptake in AM fungi during symbiosis [34,39,57]. A recent study showed that Bacillus velezensis S141, a plant growth-promoting rhizobacterium (PGPR), can stimulate R. irregularis colonization and spore production in Lotus japonicus by activating genes within this group, while concurrently downregulating genes involved in polyphosphate biosynthesis via vacuolar transporter chaperones (VTCs) and endopolyphosphatases (PPNs) [58]. Except for PPN1, genes associated with phosphate metabolism were generally downregulated. The expression of the monosaccharide transporter MST2 showed an increase, resulting in an uptick in AM fungal propagules. Consequently, our findings suggest that LED light may function as an activator of lipid biosynthesis in the host plant, facilitating transfer of nutrients to AM fungi and promoting their growth and development. This could potentially expedite the AM fungal life cycle by reducing the time needed to produce spores, potentially stimulating spore production over a shorter period.
5. Conclusions
In summary, our study examined the effects of red and blue LED lights on maize growth, biomass, and fungal abundance. We discovered that a red-to-blue light ratio of 60:40, combined with a total light intensity of 300 µmol m−2 s−1, provided the optimal conditions for improving several crucial parameters of maize growth and development. This specific LED light intensity significantly encouraged the colonization and spore production of R. irregularis, compared to the GH conditions. The gene expression analyses in both the host plant and the AM fungus illuminated the molecular mechanisms enabling these improvements. The LED light conditions elevated the expression of critical photosynthesis-related genes, leading to an increased net photosynthetic sugar production. In addition, LED light elevated the expression of genes involved in the symbiotic relationship between maize and R. irregularis, as well as genes related to regulating the fungus’s cell cycle. Our findings show the potential of combining red and blue LED lights as a practical tool for augmenting the efficiency of AM fungal inoculum production under a substrate-based system.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy14102342/s1, Table S1 Preparation for the half-strength Hoagland solution (for mycorrhizal). Table S2 Primers used for the amplification of photosynthetic genes in Zea mays. Table S3 Primers listed of arbuscular mycorrhiza-induced genes in Zea mays. Table S4 Primers of marker genes for R. irregularis. Table S5 The growth of maize and AM colonization at 45 days after inoculation (DAI) across four different conditions: non-inoculation under greenhouse (GH) and LED light (LED), and inoculation with R. irregularis under greenhouse (AMF+GH), and under LED light (AMF+LED) [16,29,35,37,59].
Author Contributions
Conceptualization, N.B., K.S., and N.T.; methodology, J.W., P.P., P.S., and T.G.; software, S.K. and N.L.; validation, K.T.; formal analysis, S.K. and N.L.; investigation, J.W., P.P., P.S., T.G., and K.T.; data curation, S.K. and N.L.; writing—original draft preparation, S.K. and N.L.; writing—review and editing, P.T.; visualization, P.T.; supervision, N.B., K.S., and N.T.; project administration, N.B.; funding acquisition, K.S., N.T., and P.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by The Royal Golden Jubilee (RGJ) Ph.D. Program, The Thailand Research Fund (TRF), Suranaree University of Technology (SUT), The NSRF via the Program Management Unit for Human Resources & Institutional Development and Innovation (PMU-B) under grant number B13F660055, and the Grant-in-Aid for Scientific Research (JSPS 22H02226) from the Japan Society for the Promotion of Science.
Institutional Review Board Statement
The biosecurity concerns of this study were reviewed and approved by Suranaree University of Technology (approval number: SUT-IBC-007/2023) and are in accordance with the levels of risk in pathogens and animal toxins listed in “the Risk Group of Pathogen and Animal Toxin (2017)” issued by the Department of Medical Sciences, Ministry of Public Health, the Pathogen and Animal Toxin Act (2015) and Biosafety Guidelines for Modern Biotechnology, BIOTEC (2016).
Data Availability Statement
The original data analyzed in this study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kivlin, S.N.; Hawkes, C.V.; Treseder, K.K. Global diversity and distribution of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2011, 43, 2294–2303. [Google Scholar] [CrossRef]
- Lee, E.H.; Eo, J.K.; Ka, K.H.; Eom, A.H. Diversity of arbuscular mycorrhizal fungi and their roles in ecosystems. Mycobiology 2013, 41, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, P.; Gianinazzi, S.; Perotto, S.; Turnau, K.; Barea, J.-M. The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fertil. Soils 2003, 37, 1–16. [Google Scholar] [CrossRef]
- Mei, L.; Yang, X.; Cao, H.; Zhang, T.; Guo, J. Arbuscular mycorrhizal fungi alter plant and soil C:N:P stoichiometries under warming and nitrogen input in a semiarid meadow of China. Int. J. Environ. Res. Public Health 2019, 16, 397. [Google Scholar] [CrossRef]
- Prasad, K.; Aggarwal, A.; Yadav, K.; Tanwar, A. Impact of different levels of superphosphate using arbuscular mycorrhizal fungi and Pseudomonas fluorescens on Chrysanthemum indicum L. J. Soil Sci. Plant Nutr. 2012, 12, 451–462. [Google Scholar]
- Sudheer, S.; Johny, L.; Srivastava, S.; Adholeya, A. The trade-in-trade: Multifunctionalities, current market and challenges for arbuscular mycorrhizal fungal inoculants. Symbiosis 2023, 89, 259–272. [Google Scholar] [CrossRef]
- IJdo, M.; Cranenbrouck, S.; Declerck, S. Methods for large-scale production of AM fungi: Past, present, and future. Mycorrhiza 2011, 21, 1–16. [Google Scholar] [CrossRef]
- Sayeed Akhtar, M.; Nor, S.; Abdullah, A. Mass production techniques of arbuscular mycorrhizal fungi: Major advantages and disadvantages: A review. Biosci. Biotechnol. Res. Asia 2014, 11, 1199–1204. [Google Scholar] [CrossRef]
- Chourasiya, D.; Gajghate, R.; Prakash, A.; Sharma, M.P. Low-cost technologies for AMF inoculum production using various agro-wastes and other by-products. In Arbuscular Mycorrhizal Fungi in Sustainable Agriculture: Inoculum Production and Application; Springer Nature: Singapore, 2024; pp. 145–165. [Google Scholar]
- Bencherif, K.; Laruelle, F.; Tisserant, B.; Dalpé, Y.; Lounés-Hadj Sahraoui, A. Engineering approach for production of arbuscular mycorrhizal inoculum adapted to saline soil management. Stresses 2023, 3, 404–423. [Google Scholar] [CrossRef]
- Genre, A.; Bonfante, P. Building a mycorrhizal cell: How to reach compatibility between plants and arbuscular mycorrhizal fungi. J. Plant Interact. 2005, 1, 3–13. [Google Scholar] [CrossRef]
- Pepe, A.; Giovannetti, M.; Sbrana, C. Lifespan and functionality of mycorrhizal fungal mycelium are uncoupled from host plant lifespan. Sci. Rep. 2018, 8, 10235. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Lahrmann, U.; Langen, G. Broad compatibility in fungal root symbioses. Curr. Opin. Plant Biol. 2014, 20, 135–145. [Google Scholar] [CrossRef] [PubMed]
- INVAM International Culture Colletion of (Vesicular) Arbuscular Micorrhizal Fungi. Available online: https://invam.ku.edu/ (accessed on 16 September 2024).
- Kiddee, S.; Yuttavanichakul, W.; Boonkerd, N.; Teaumroong, N.; Saito, K.; Tittabutr, P. Secretion compounds from Brevibacillus Sp. SUT47 promote spore Propagation of Acaulospora Tuberculata colonizing maize roots (Zea mays L. cultivar Suwan 5). ScienceAsia 2020, 46, 634–638. [Google Scholar] [CrossRef]
- Bezrutczyk, M.; Hartwig, T.; Horschman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired phloem loading in zmsweet13a, b, c sucrose transporter triple knock-out mutants in Zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.W.; Barta, D.J.; Ignatius, R.W.; Martin, T.S. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and the opportunities of modern LED systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Xu, Y.; Chang, Y.; Chen, G.; Lin, H. The Research on LED supplementary lighting system for plants. Optik 2016, 127, 7193–7201. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Green-light supplementation for enhanced lettuce growth under red- and blue-light-emitting diodes. HortScience 2004, 39, 1617–1622. [Google Scholar] [CrossRef]
- Ohtake, N.; Ishikura, M.; Suzuki, H.; Yamori, W.; Goto, E. Continuous irradiation with alternating red and blue light enhances plant growth while keeping nutritional quality in lettuce. HortScience 2018, 53, 1804–1809. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Urbonavičiūtė, A.; Šabajevienė, G.; Duchovskis, P. The effect of red and blue light component on the growth and development of frigo strawberries. Zemdirb. Agric. 2010, 97, 99–104. [Google Scholar]
- He, D.; Kozai, T.; Niu, G.; Zhang, X. Light-emitting diodes for horticulture. In Light-Emitting Diodes Materials, Processes, Devices and Applications; Springer: Cham, Switzerland, 2019; pp. 513–547. [Google Scholar]
- Viršile, A.; Olle, M.; Duchovskis, P. LED lighting in horticulture. In Light Emitting Diodes for Agriculture: Smart Lighting; Springer: Berlin/Heidelberg, Germany, 2017; pp. 113–147. [Google Scholar]
- Xu, J.; Guo, Z.; Jiang, X.; Ahammed, G.J.; Zhou, Y. Light regulation of horticultural crop nutrient uptake and utilization. Hortic. Plant J. 2021, 7, 367–379. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.P.; Wang, J.Z.; Fu, W. Growth, photosynthesis, and nutrient uptake at different light intensities and temperatures in lettuce. HortScience 2019, 54, 1925–1933. [Google Scholar] [CrossRef]
- Cruz, A.F. Effect of light-emitting diodes on arbuscular mycorrhizal fungi associated with bahiagrass (Paspalum notatum Flügge) and millet [Pennisetum glaucum (L.) R. Br]. Bioagro 2016, 28, 163–170. [Google Scholar]
- Tenzin, U.W.; Noirungsee, N.; Runsaeng, P.; Noppradit, P.; Klinnawee, L. Dry-season soil and co-cultivated host plants enhanced propagation of arbuscular mycorrhizal fungal spores from sand dune vegetation in trap culture. J. Fungi 2022, 8, 1061. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Saito, K. Role of cell wall polyphosphates in phosphorus transfer at the arbuscular interface in mycorrhizas. Front. Plant Sci. 2021, 12, 725939. [Google Scholar] [CrossRef]
- Songsaeng, A.; Tittabutr, P.; Umnajkitikorn, K.; Boonkerd, N.; Wongdee, J.; Songwattana, P.; Piromyou, P.; Greetatorn, T.; Girdthai, T.; Teaumroong, N. Application of light-emitting diodes with plant growth-promoting rhizobacteria and arbuscular mycorrhiza fungi for tomato seedling production. Agronomy 2022, 12, 2458. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un systeme radiculaire. Recherche de methodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Daniels, B.A.; Skipper, H. Methods for the Recovery and quantitative estimation of propagules from soil. In Methods & Principles of Mycorrhizal Research, American Phytopathological Society; American Phytopathological Society: Saint Paul, MN, USA, 1982; pp. 29–35. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Kiddee, S.; Wongdee, J.; Piromyou, P.; Songwattana, P.; Greetatorn, T.; Boonkerd, N.; Teaumroong, N.; Saito, K.; Tittabutr, P. Unveiling the tripartite synergistic interaction of plant-arbuscular mycorrhizal fungus symbiosis by endophytic Bacillus velezensis S141 in Lotus japonicus. Symbiosis 2024, 92, 355–367. [Google Scholar] [CrossRef]
- Sugimura, Y.; Kawahara, A.; Maruyama, H.; Ezawa, T. Plant foraging strategies driven by distinct genetic modules: Cross-ecosystem transcriptomics approach. Front. Plant Sci. 2022, 13, 903539. [Google Scholar] [CrossRef]
- Lu, Y.; Li, Y.; Zhang, J.; Xiao, Y.; Yue, Y.; Duan, L.; Zhang, M.; Li, Z. Overexpression of Arabidopsis molybdenum cofactor sulfurase gene confers drought tolerance in maize (Zea mays L.). PLoS ONE 2013, 8, e52126. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Ezawa, T.; Saito, K. Polyphosphate polymerizing and depolymerizing activity of VTC4 protein in an arbuscular mycorrhizal fungus. Soil Sci. Plant Nutr. 2022, 68, 256–267. [Google Scholar] [CrossRef]
- Kitazaki, K.; Fukushima, A.; Nakabayashi, R.; Okazaki, Y.; Kobayashi, M.; Mori, T.; Nishizawa, T.; Reyes-Chin-Wo, S.; Michelmore, R.W.; Saito, K.; et al. Metabolic reprogramming in leaf lettuce grown under different light quality and intensity conditions using narrow-band LEDs. Sci. Rep. 2018, 8, 7914. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Ottosen, C.O.; Rosenqvist, E. Spectral effects of LEDs on chlorophyll fluorescence and pigmentation in Phalaenopsis ‘Vivien’and ‘Purple Star’. Physiol. Plant. 2015, 154, 314–327. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, A.; Cheng, Z.M. (Max) Effects of light emitting diode lights on plant growth, development and traits a meta-analysis. Hortic. Plant J. 2021, 7, 552–564. [Google Scholar] [CrossRef]
- Hao, X.; Papadopoulos, A.P. Effects of supplemental lighting and cover materials on growth, photosynthesis, biomass partitioning, early yield and quality of greenhouse cucumber. Sci. Hortic. 1999, 80, 1–18. [Google Scholar] [CrossRef]
- Jones, M.A. Using light to improve commercial value. Hortic. Res. 2018, 5, 47. [Google Scholar] [CrossRef]
- Cavagnaro, T.R.; Jackson, L.E.; Six, J.; Ferris, H.; Goyal, S.; Asami, D.; Scow, K.M. Arbuscular mycorrhizas, microbial communities, nutrient availability, and soil aggregates in organic tomato production. Plant Soil 2006, 282, 209–225. [Google Scholar] [CrossRef]
- Lupo, M.; Bashir, M.A.; Silvestri, C.; Brunori, E.; Pica, A.L.; Cristofori, V. LED lighting effects on plant growth and quality of Pyrus communis L. propagated in vitro. Agronomy 2022, 12, 2531. [Google Scholar] [CrossRef]
- Stamford, J.D.; Stevens, J.; Mullineaux, P.M.; Lawson, T. LED Lighting: A Grower’s Guide to Light Spectra. HortScience 2023, 58, 180–196. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Kang, C.; Kaiser, E.; Kuang, Y.; Yang, Q.; Li, T. Red/Blue light ratios induce morphology and physiology alterations differently in cucumber and tomato. Sci. Hortic. 2021, 281, 109995. [Google Scholar] [CrossRef]
- Matsumura, A.; Horii, S.; Ishii, T. Effects of arbuscular mycorrhizal fungi and intercropping with bahiagrass on growth and anti-oxidative enzyme activity of radish. J. Jpn. Soc. Hortic. Sci. 2007, 76, 224–229. [Google Scholar] [CrossRef][Green Version]
- Yu, Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2018, 17, 25–36. [Google Scholar] [CrossRef]
- Petrillo, E.; Herz, M.A.G.; Barta, A.; Kalyna, M.; Kornblihtt, A.R. Let there be light: Regulation of gene expression in plants. RNA Biol. 2014, 11, 1215–1220. [Google Scholar] [CrossRef]
- Fellbaum, C.R.; Gachomo, E.W.; Beesetty, Y.; Choudhari, S.; Strahan, G.D.; Pfeffer, P.E.; Kiers, E.T.; Bücking, H. Carbon availability triggers fungal nitrogen uptake and transport in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 2666–2671. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the arbuscular mycorrhiza market: From arbuscules to common mycorrhizal networks. N. Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef]
- Thompson, J.P. Soilless culture of vesicular–arbuscular mycorrhizae of cereals: Effects of nutrient concentration and nitrogen source. Can. J. Bot. 1986, 64, 2282–2294. [Google Scholar] [CrossRef]
- Harris, S.D. Branching of Fungal Hyphae: Branching of fungal hyphae: Regulation, mechanisms and comparison with other branching systems. Mycologia 2008, 100, 823–832. [Google Scholar] [CrossRef]
- Sugimura, Y.; Saito, K. Transcriptional profiling of arbuscular mycorrhizal roots exposed to high levels of phosphate reveals the repression of cell cycle-related genes and secreted protein genes in Rhizophagus irregularis. Mycorrhiza 2017, 27, 139–146. [Google Scholar] [CrossRef]
- Ezawa, T.; Saito, K. How do arbuscular mycorrhizal fungi handle phosphate? New insight into fine-tuning of phosphate metabolism. New Phytol. 2018, 220, 1116–1121. [Google Scholar] [CrossRef]
- Kobae, Y.; Kawachi, M.; Saito, K.; Kikuchi, Y.; Ezawa, T.; Maeshima, M.; Hata, S.; Fujiwara, T. Up-regulation of genes involved in N-acetylglucosamine uptake and metabolism suggests a recycling mode of chitin in intraradical mycelium of arbuscular mycorrhizal fungi. Mycorrhiza 2015, 25, 411–417. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).