Abstract
The fall armyworm (FAW), Spodoptera frugiperda, is a serious pest that threatens a range of important crops worldwide. It originated in America and rapidly dispersed throughout Africa and Asia in 2018. There are two subtypes, corn-strain (C-strain) and rice-strain (R-strain), that have different host plant preferences, and the individuals damaging maize in China were identified as C-strain. In the present study, we found FAW individuals damaging rice plants in the field of Guangdong Province, China. FAW larvae and male adults were collected, and the majority of FAWs were characterized as CO I R-strain Tpi C-strain, which is similar to the FAWs damaging maize in China. The FAW adults preferred laying eggs on maize plants more than on rice plants. Compared to those that were fed maize leaves, the FAW larvae were unable to survive when fed 4-week-old rice plants, whereas they could complete their life cycle on 2-week-old rice plants, for which the total survival rate was 8%. The pre-adult- and pupal-stage durations were prolonged, and the fecundity of adult females decreased. Thus, the FAWs found in paddy fields showed better fitness on maize than on rice in the laboratory. Owing to their low survival rate on rice plants, they were unlikely to damage paddy fields in large areas, but populations of FAWs in paddy fields should be monitored.
1. Introduction
The fall armyworm (FAW), Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), is a serious pest moth that has spread rapidly from tropical to subtropical areas worldwide. This polyphagous species is capable of feeding on more than 80 plant species, especially major crops including maize Zea mays L. and rice Oryza sativa L. [1]. Severe FAW infestations can be devastating, resulting in 21–53% yield loss in maize [2]. It is native to America’s tropical and subtropical areas, and since 2016, it has invaded western Africa and now appears in most African and Southeast Asian countries [3,4]. In December 2018, FAWs invaded southwestern China and then spread rapidly to 26 provinces, affecting maize field areas up to 1125.33 thousand hectares [5].
There are two FAW subpopulations that exhibit reproducible biases to host plants in the field [6]. The corn-strain (C-strain) is most common in maize and sorghum, while the rice-strain (R-strain) prefers rice, millet and grass species associated with pasture habitats [7,8]. The two strains also have different physiological features, such as different sex pheromone compositions, mating times, oviposition preference and susceptibility to Bacillus thuringiensis toxin Cry1F [9,10,11,12,13]. They are morphologically indistinguishable but can be identified using molecular markers that are associated with specific host plants [14]. The most commonly used genetic methods for population studies involve mitochondrial haplotypes, especially polymorphisms in the cytochrome oxidase subunit I gene (CO I) and the triosephosphate isomerase (Tpi) gene, which are highly conserved in noctuid moths and located on the Z sex chromosome [6,14].
Rice is one of the important staple crops, feeding more than half of the world’s population [15], and it has been reported as one of the key hosts of FAW in other areas, e.g., the southeastern states of the U.S. and Philippines [16,17]. China is the largest producer and consumer of rice in the world, with the highest yield production in the world [18]. Since FAWs invaded China in 2019, they have been found to feed sporadically on paddy fields in the Yunnan, Fujian, and Hubei and Provinces in China [19,20,21,22]. Based on the Tpi gene, the FAWs found on paddy field in Hubei Province were identified as C-strain [21]. Therefore, it is necessary to monitor the population of FAWs on paddy fields.
In this work, we found FAW larvae damaging paddy fields in Taishan City and in Yangjiang City, Guangdong Province, China. To determine whether this population of FAW newly invaded China, larvae and adults were collected from the paddy fields in both of the locations and subsequently transferred to the laboratory to confirm their species and strain using molecular markers based on the CO I and Tpi genes. Based on the host strain, to assess whether this local population poses a threat to rice, we evaluate the fitness of these strains on rice and maize by comparing the oviposition preference and survival rate of the two colonies on maize and rice leaves, and then age-stage, two-sex life table of the Taishan colony.
2. Materials and Methods
2.1. Insects and Plants
From the paddy field (O. sativa variety Yuehesimiao) in Yangjiang City, Guangdong Province, China (21.7523° N 111.6176° E), thirty-five 5th–7th instar FAW larvae were collected by hand in 20 December 2020. Six of them were used for host strain identification and the rest were used for further experiments.
From the paddy fields (O. sativa variety Xiangsimiao 126) in Taishan city, Guangdong Province, China (21.8254° N 112.5587° E), forty-seven 5th–7th instar FAW larvae were collected by hand, and 5 male adults were captured by sex pheromone traps set in paddy fields in 14 November 2021. The village where these paddy fields were located had no maize fields during that season. Three of the larvae and all male adults were used for host strain identification and the rest larvae were used for further experiments.
For the oviposition preference, survival rate, and developmental time experiments, seeds of the rice variety Xiangsimiao 126 were obtained from the Taishan Agricultural Sciences Research Institute, and the maize seed (Z. mays variety Zhengdan 958) was provided by Doneed Co., Ltd. (Beijing, China) Seeds were sown in a plastic pot (diam. 9 cm × 10 cm) filled with moist commercial mix composed of 45% vermiculite, 35% peat moss and 20% organic matter (Xianghui Agriculture Technology, Liuyang, China) and watered when needed.
All the experiments were conducted at 26 ± 1 °C, 70 ± 5% RH and 18:6 h L:D in incubators (QX-256, Jiangnan Instrument Factory, Ningbo, China).
2.2. Host Strain Identification
Total DNA was extracted from the collected FAW samples in accordance with the instructions of the manufacturer of the TIANamp Genomic DNA Kit (DP304, Tiangen, Beijing, China). The CO I gene and Tpi gene were amplified to confirm the species and, the host strain identify using 2× SanTaq PCR Master Mix (Sangon Biotech, Shanghai, China) according to the instructions. Primers JM76 (5′-GAGCTGAATTAGGGACTCC-3′) and JM77 (5′-ATCACCTCCACCTGCAGGATC-3′) were used to produce a 569-bp CO I fragment, and Tpi-F (5′-GGTGAAATCTCCCCTGCTATG-3′) and Tpi-R (5′-AATTTTATTACCTGCTGTGG-3′) were used to produce a 406-bp Tpi fragment [14]. PCR parameters were as follows: initial denaturation (95 °C for 10 min); 35 cycles of 95 °C for 1 min, 55 °C for 40 s, and 72 °C for 1 min; and a final extension of 10 min at 72 °C [4]. All PCR products were assessed by 1.5% agarose gel electrophoresis and subsequently sequenced by Sangon Biotech Co., Ltd. (Shanghai, China). Phylogenetic trees were generated via neighbor-joining (NJ) tree analysis and included CO I gene fragments of related Spodoptera species.
2.3. Oviposition Preference
The larvae collected from the paddy fields in Taishan and Yangjiang cities were transferred to the laboratory and individually fed on leaves collected from the same paddy field until the pupal stage in plastic vials (dia. 3 cm × 3 cm). After emergence, as treatments, three pairs of male and female adult moths from each city were transferred into cages (1 m × 1 m, 0.15 mm mesh) containing two 4-week-old rice plants and two 4-week-old maize plants for two-choice tests. The plants were randomly placed in four corners of the cage, and 10% sucrose water was provided as food in the middle of the cage bottom. There were 3 replicates and each cage was used as a replicate. After 48 h, the number of egg masses laid on each seedling was recorded.
2.4. Survival Rate and Developmental Time
The survival rate of FAWs collected from paddy fields of Taishan and Yangjiang cities were examined via the eggs collected in oviposition preference experiments. After hatching, the new larvae from each colony were placed individually in plastic vials (dia. 3 cm × 3 cm), and provided with 4-week-old section of rice or maize leaf (approximately 1 cm × 1 cm, leaf vein was removed) as treatment. The leaf sections were renewed every 2–3 days. For both colonies and treatments, the mortality and stage of the FAWs were recorded daily until the pupal stage. There were 70 larvae per treatment.
2.5. Life Table of FAWs Feeding on Rice or Maize Leaves
FAW eggs used in life-table parameter estimation were obtained from the offspring of FAWs colony established from larvae in Taishan City, that fed on maize leaves, as described in Section 2.4. One hundred newly hatched larvae were provided with a section of rice or maize leaf (about 2 cm2) cut from 2-week-old seedlings by the procedure described the same as above until they entered the pupal stage. Once emerged, adults from each treatment were randomly paired and transferred to a clean container (dia. 9 cm × 10.5 cm). Paper towels were used as spawning substrates, and 10% sucrose water was supplied as food. The survival rate and quantity of eggs were recorded daily.
2.6. Data Analysis
The survival rate data were analyzed based on the Mantel–Cox test and Kaplan–Meier survival curves generated using SPSS 21 (IBM, Albany, NY, USA). The life table parameters were calculated according to two-sex life table theory [23,24,25,26] using TWOSEX-MSChart [27]. The oviposition preference data were compared via paired t-tests. The age-specific life table parameters and developmental duration of S. frugiperda were first tested for normality. For normally distributed data, we used one-way ANOVA to determine the effect of treatment, and we compared the means of treatments using Duncan’s multiple range test. For the nonnormally distributed data, we used a nonparametric method, the Kruskal–Wallis test, with Bonferroni corrections for multiple comparisons.
3. Results
3.1. Most of the FAW Larvae Were CO I R-Strain Tpi C-Strain
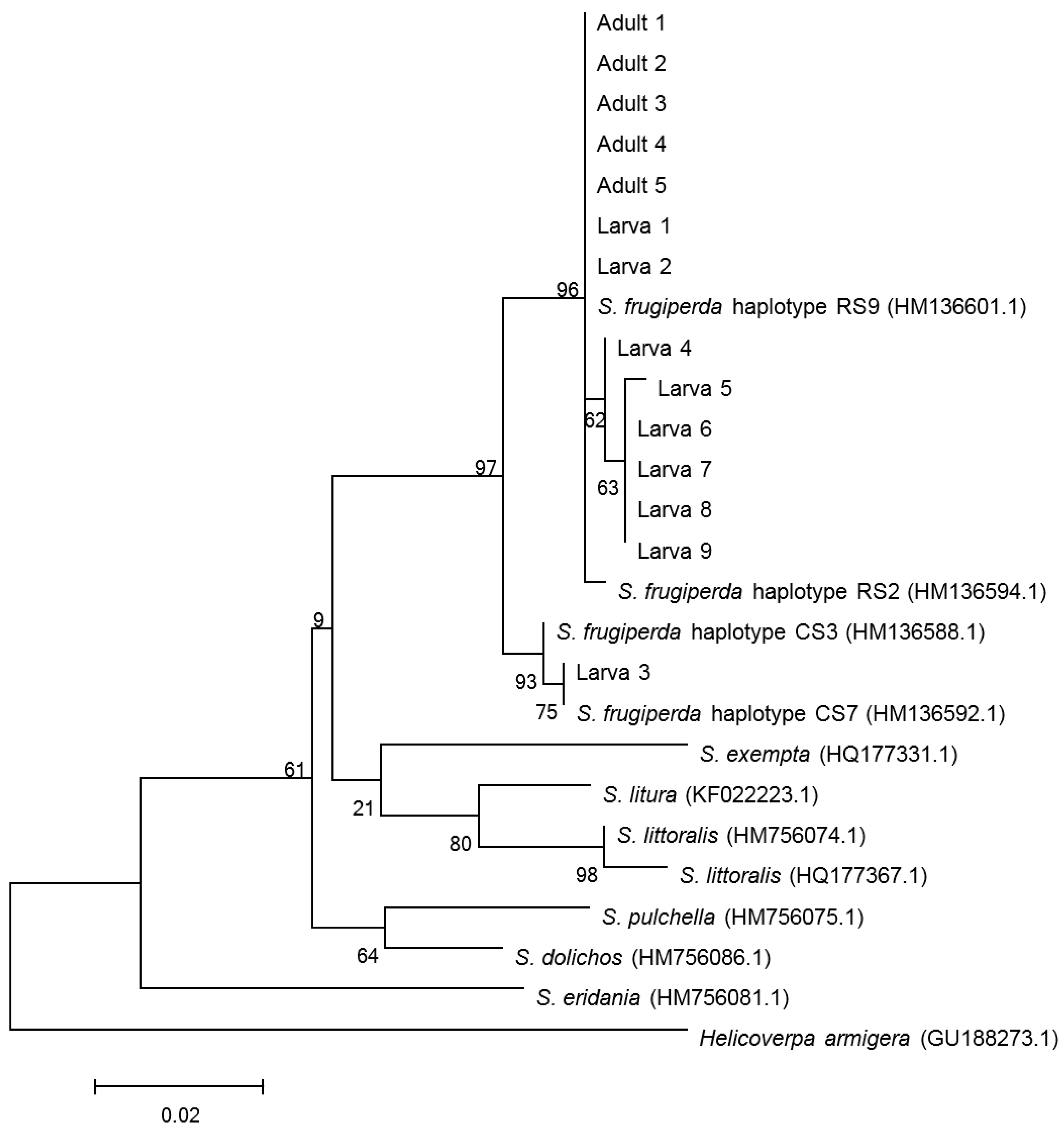
The CO I segment was used to confirm the species identification by comparing it with the barcode sequences in GenBank. Based on BLAST analyses of the sequences obtained from the NCBI GenBank database and phylogenetic tree (Figure 1), the larvae and adults collected from the paddy field in both Yangjiang City and Taishan City were identified as S. frugiperda, as indicated by 99–100% similarity in sequence data and coverage.
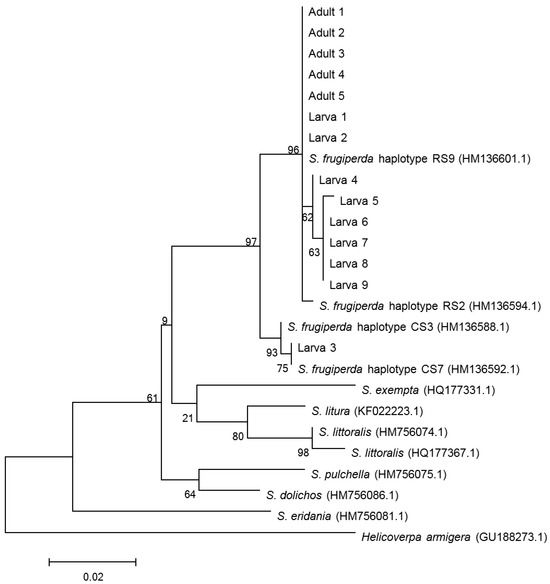
Figure 1.
Phylogenetic tree constructed by NJ analysis based on CO I gene fragments. Adults 1–5 indicate the samples of adult moths collected in Taishan City. Larva 1–3 indicate the samples of larvae collected in Taishan City and Larva 4–9 indicate the samples of larvae collected in Yangjiang City. The number in parentheses indicate the accession number of the sequence in GenBank.
Among all the samples, one sequence of samples collected in Taishan City clustered with the CO I C-strain haplotype (HM136588.1), and the others clustered with the CO I R-strain haplotype (HM136601.1); these findings are consistent with the results of comparing polymorphic sites of CO I gene fragments (Figure 2). Based on the polymorphism characteristics of the Tpi gene, all the tested larvae and adults collected from the paddy field were C-strain (Figure 3).
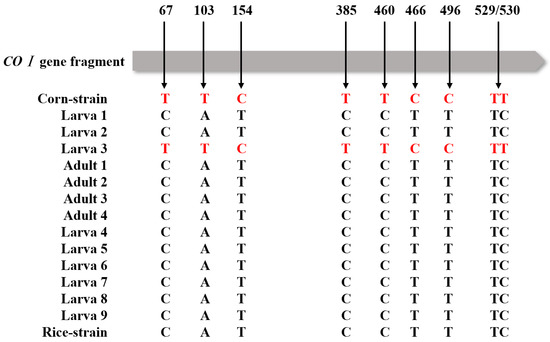
Figure 2.
Haplotype analysis based on CO I gene fragments; arrows indicate maize or rice strain-specific polymorphic loci. Adults 1–5 indicate the samples of adult moths collected in Taishan City. Larva 1–3 indicate the samples of larvae collected in Taishan City and Larva 4–9 indicate the samples of larvae collected in Yangjiang City.
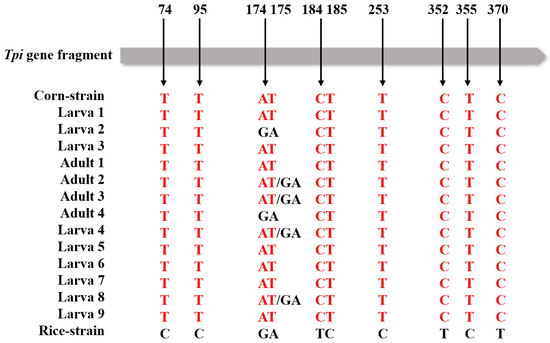
Figure 3.
Haplotype analysis based on Tpi gene fragments; arrows indicate corn or rice strain-specific polymorphic loci. Adults 1–5 indicate the samples of adult moths collected in Taishan City. Larva 1–3 indicate the samples of larvae collected in Taishan City and Larva 4–9 indicate the samples of larvae collected in Yangjiang City.
3.2. Oviposition Preference of FAW Collected from Paddy Fields
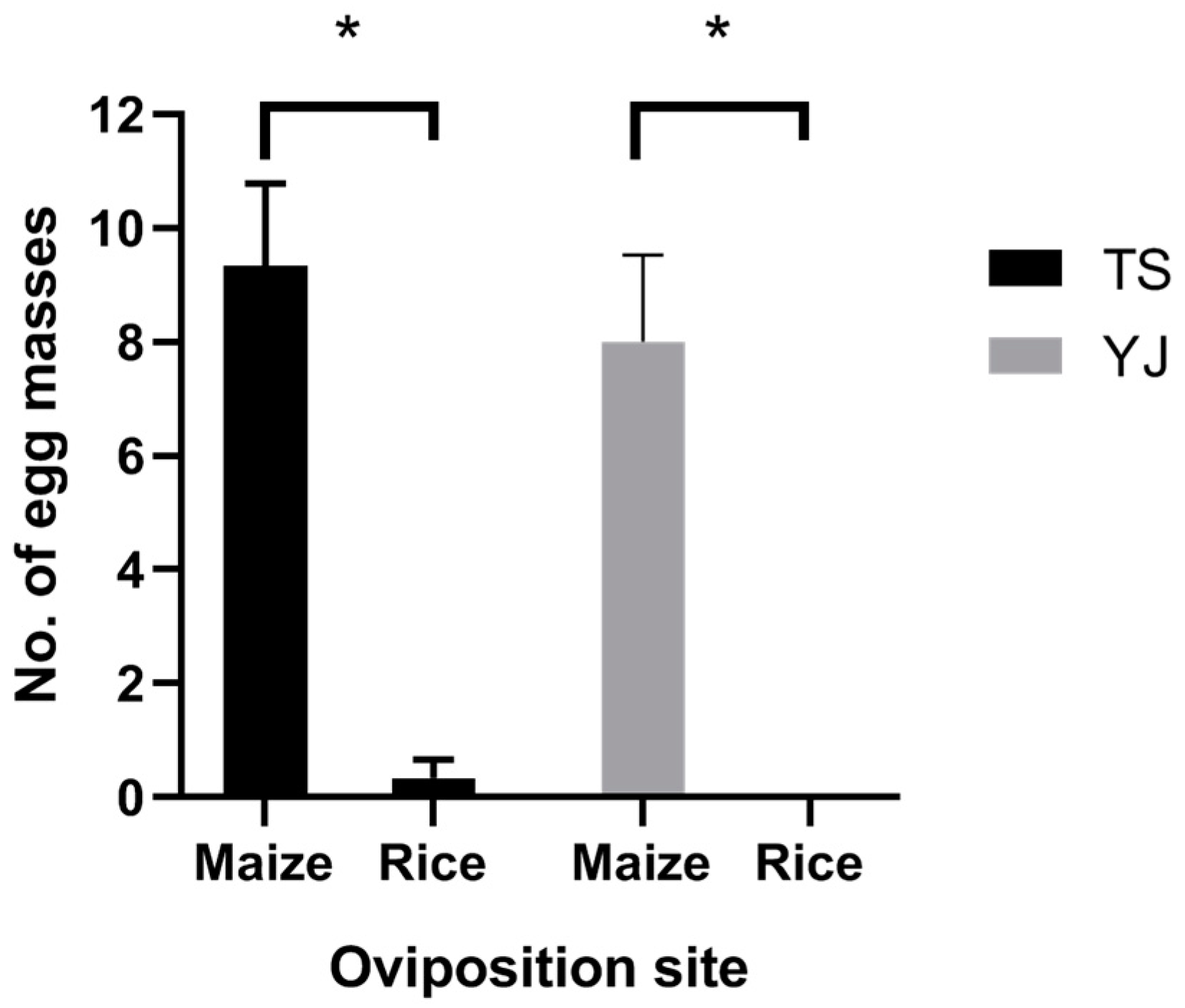
To our surprise, both FAW female adults from Taishan (TS) and Yangjiang (YJ) City significantly preferred maize seedlings for oviposition sites compared to rice seedlings according to the two-choice test (Figure 4, TS: t2 = 5.892, p = 0.028; YJ: t2 = 5.237, p = 0.035). The mean egg masses of TS female adults on the maize and rice seedlings were 9.67 ± 1.20 and 0.33 ± 0.33, respectively, and for YJ female adults, they were 8.00 ± 3.00 and 0.00 ± 0.00, respectively.
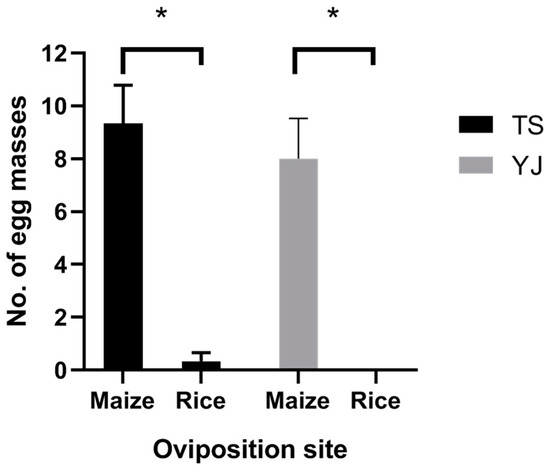
Figure 4.
Oviposition preference of FAW females collected from rice fields in Taishan City (TS) and Yangjiang City (YJ) in Guangdong Province. Values = mean ± SE. *, p < 0.05. n = 3.
3.3. The Survival Rate of 1st-Generation FAW Larvae Feeding on Maize or Rice Leaves
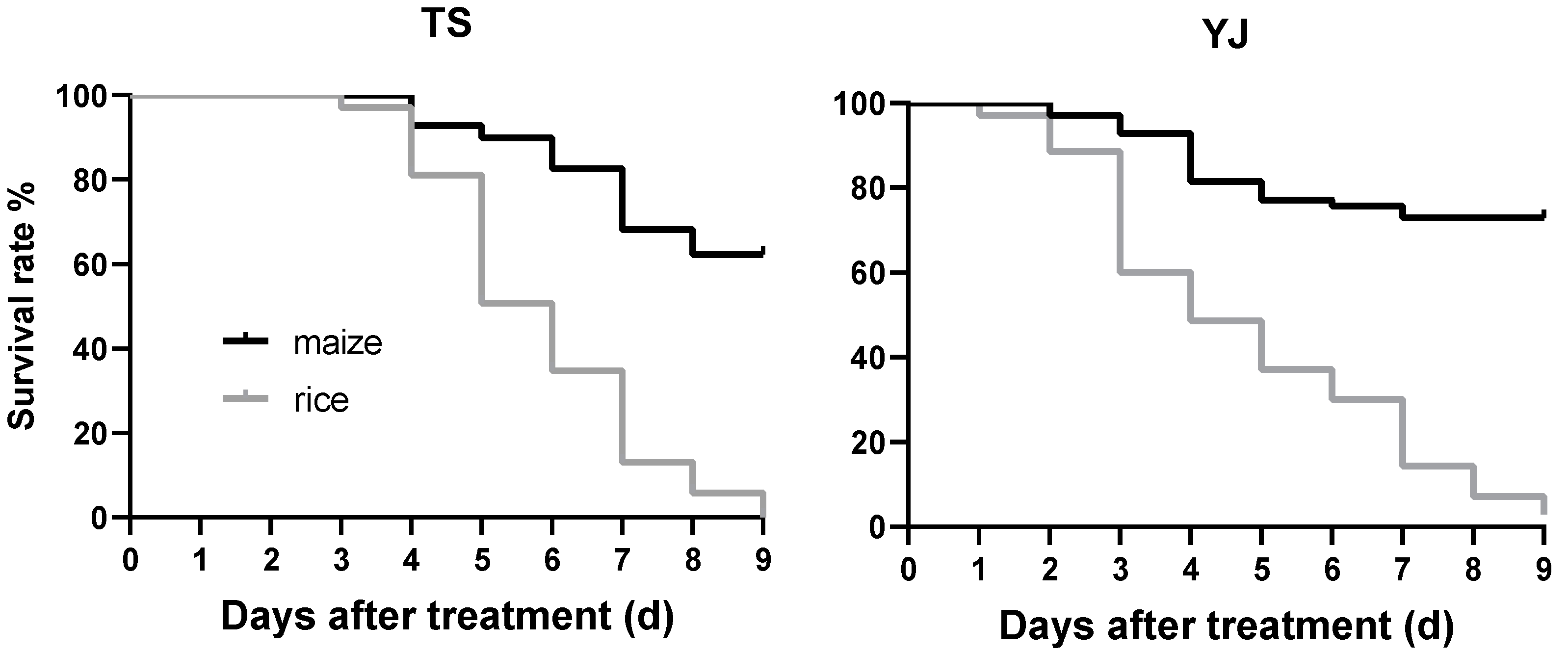
The Mantel-Cox test showed significant differences between the cumulative survival curves of FAW larvae feeding on maize and those feeding on rice leaves (Figure 5, TS: df = 1, χ2 = 70.47, p < 0.0001; YJ: df = 1, χ2 = 71.81, p < 0.0001). In TS colony, seventy FAW larvae were fed rice leaves and their survival rate decreased to 0% at 9 days after hatching. At 9 days, the survival rate of the larvae feeding on maize leaves was 62.3%. In YJ colony, the survival rate of the larvae at 9 days after hatching were 2.86% and 72.86% on rice and maize leaves, respectively.
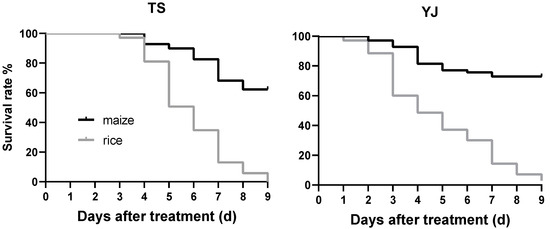
Figure 5.
Survival probability of FAW larvae collected in Taishan City (TS) and Yangjiang City (YJ) in Guangdong Province, feeding on maize or rice leaves. n = 70.
3.4. Life Table of 2nd-Generation FAW Feeding on Maize and Rice Leaves
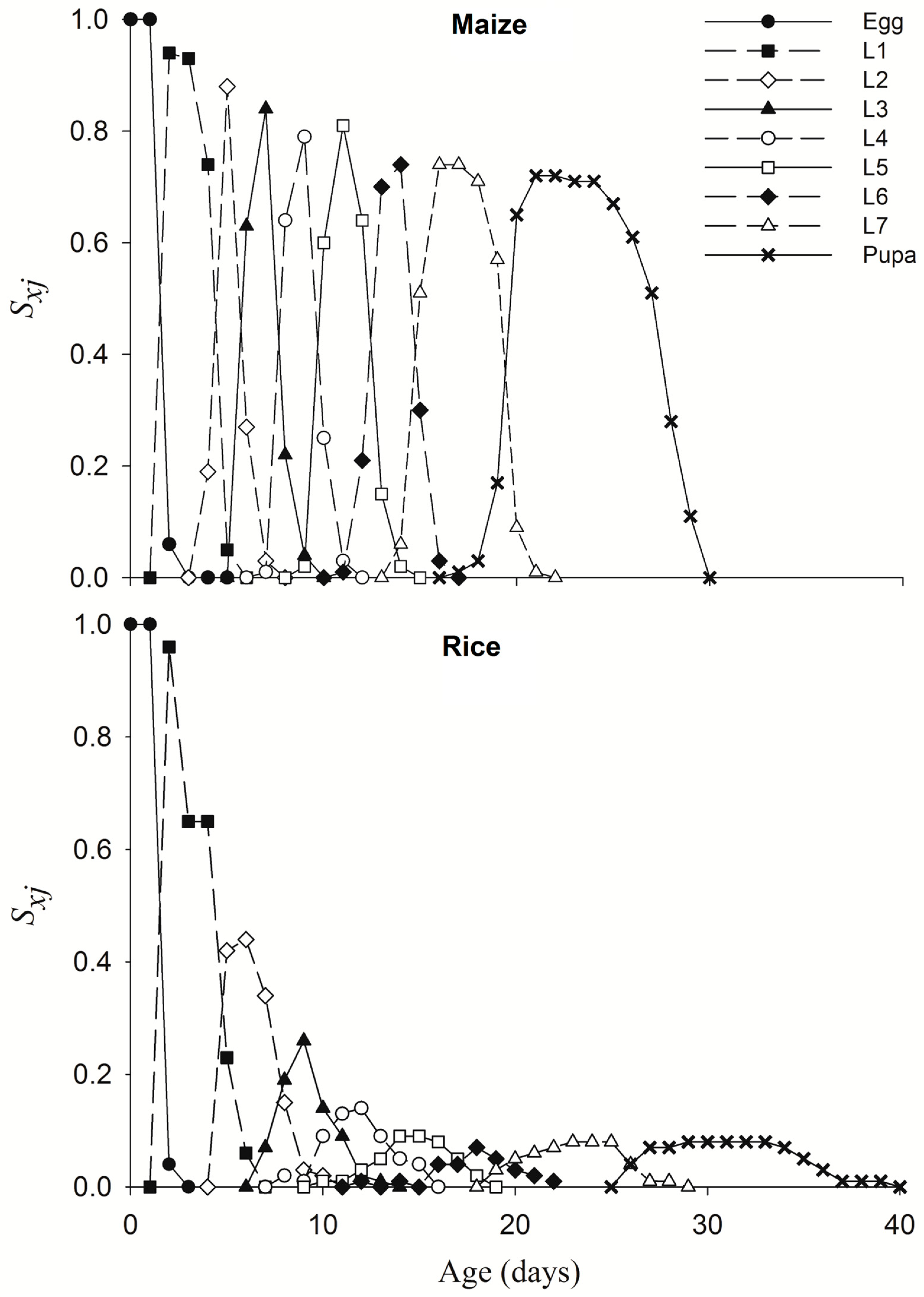
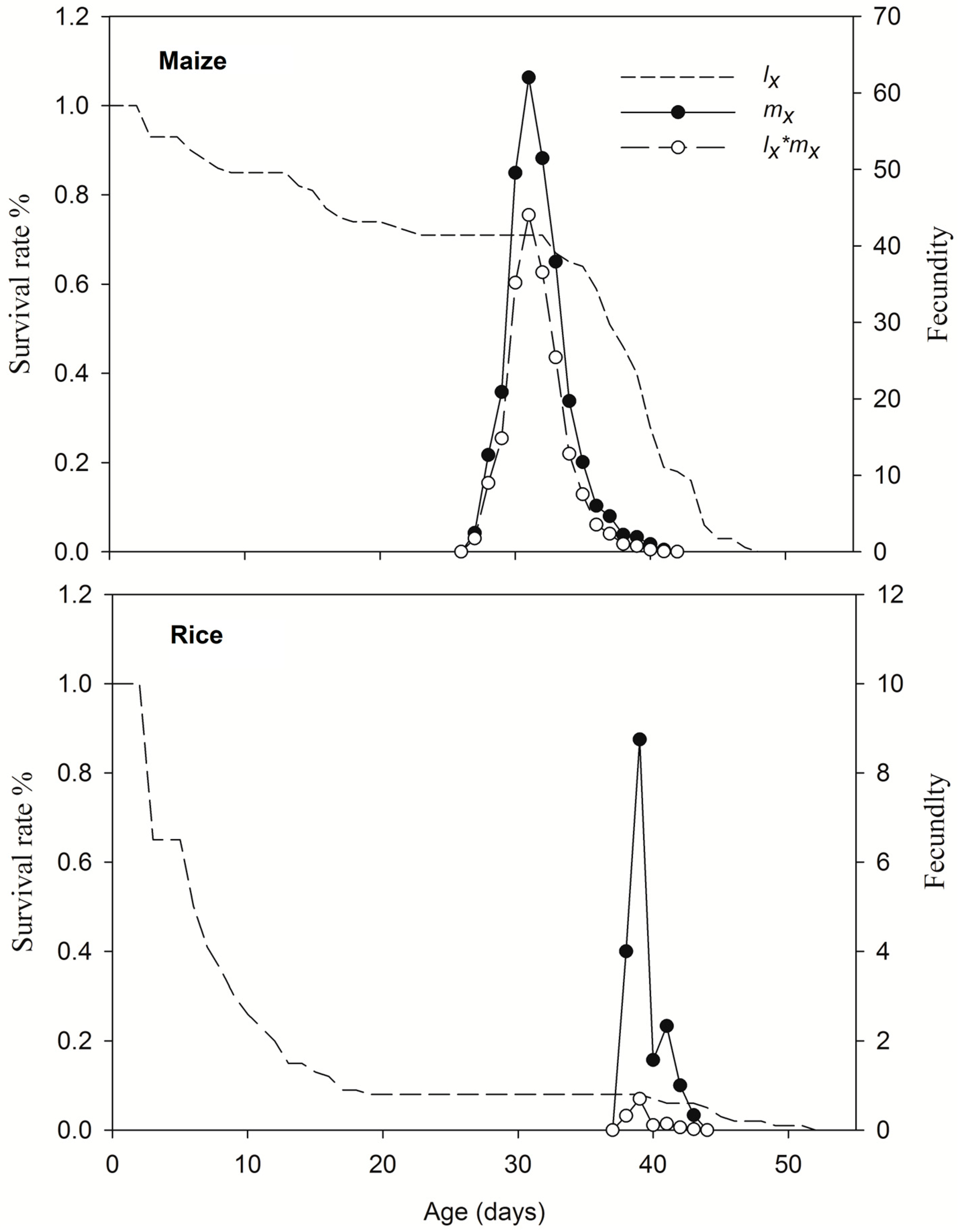
Eggs of the 1st-generation FAWs feeding on maize leaves were further subjected to life table observation. FAW larvae and pupa from 2nd generation of TS colony feeding on rice had smaller size than that on maize leaf (Figure S1). Larvae feeding on different crop leaves significantly affected the 1st–7th-stage duration, pupa duration, female pre-oviposition period, adult longevity and fecundity of FAW (Table 1). Compared with that of maize leaves, the pre-adult-stage duration of rice leaf feeding was prolonged by an average of 8.18 days, the pupal-stage duration was prolonged by 1.26 days, and adult female FAW on maize leaves laid 602.23 more eggs (Table 1, Figure 6). The overall survival rates on maize and rice leaves were 71% and 4%, respectively, and the survival rate of FAW larvae on rice leave decreased rapidly to 65% on 3 days after hatching (Figure 6). The death of last adult on maize and rice leaves occurred at 48 and 51 days, respectively (Figure 7).

Table 1.
Age-specific life table of FAW larvae feeding on corn or rice leaves. The values are presented as the means ± SEs.
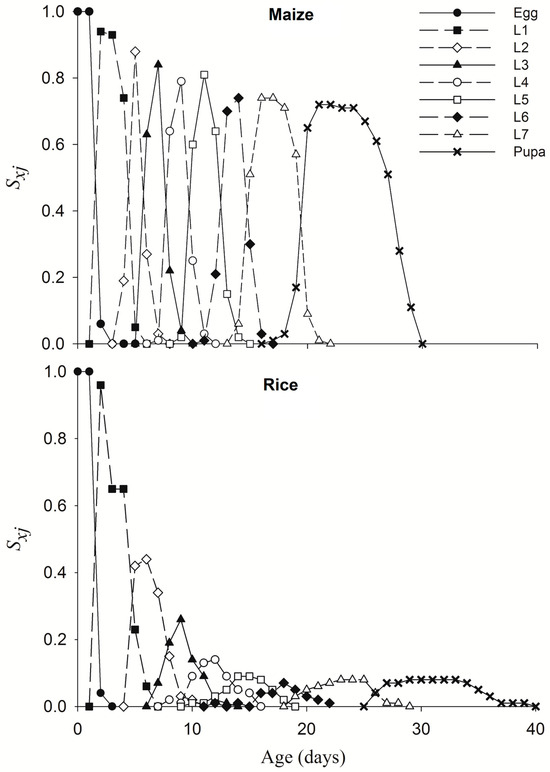
Figure 6.
Age-stage survival rate (sxj) of FAWs fed maize or rice leaves.
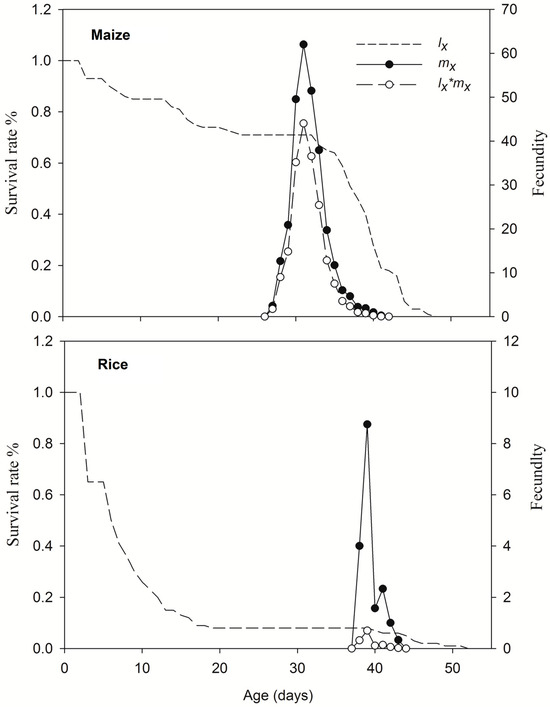
Figure 7.
Survival rate (lx), age-specific fecundity (mx) and age-stage specific reproduction (lx * mx) of FAWs fed maize or rice leaves.
The different crop feedings had effects on the net reproduction rate (R0), mean generation time (T), intrinsic rate of increase (r) and finite rate of increase (λ) (Table 2). The r and λ values of FAW populations were >0 and >1, respectively, for both maize and rice leaves, indicating that FAWs were able to survive on the two plant leaves. The net reproduction rate (R0) of FAWs feeding on maize leaves was greater than that of those feeding on rice leaves, as was the intrinsic rate of increase (r). The mean generation time (T) was prolonged when feeding on rice leaves and was extended by 8.02 days. The finite rate of increase (λ) decreased from 1.1778 to 1.0075 d−1. Thus, feeding on maize was more conducive to the development and reproduction of FAW of TS colony.
Table 2.
Population parameters of FAW feeding on maize or rice leaves. The values are presented as the means ± SEs.
4. Discussion
The association between strains and host plants is not absolute in FAWs [28,29]. Therefore, despite the population of FAWs that was identified as C-strain and mainly damaged maize in China, the risk of attack on rice still exists [4], as the FAWs was indeed found to damage rice in south of China [17,18,19,20]. In this study, we found that of the 14 tested FAW samples collected in paddy fields of Taishan and Yangjiang city, thirteen of them were interstrain hybrids of the C-strain and R-strain (CO I R-strain Tpi C-strain, Figure 1, Figure 2 and Figure 3). The result of a high proportion of hybrid individuals was consistent with the FAW populations found on maize in China [30,31], which suggested the individuals found on rice in our study came from the same populations that damaging maize in China. This finding raises our concerned about this FAW colony would have an outbreak on paddy fields. Therefore, further experiments were carried out to compare their fitness on maize and rice, and results showed that the individuals collected on rice plants preferred laying eggs on maize plants rather than rice and had a higher survival rate on maize than rice. Life table observations showed that comparing with maize, feeding on rice prolonged development-stage and adult-stage durations of FAWs and the fecundity of adult females decreased, which indicating the FAW population presented on rice had better fitness on maize than rice.
The C-strain and R-strain are capable of cross-hybridization in the field [29]. Based on the host plant and polymorphism characteristics of the Tpi gene, FAW individuals invading China have been identified as C-strain [4]. However, most FAWs in China were found to be R-strain based by polymorphisms in the COI gene but were C-strain based on the Tpi gene [30,31]. Zhang et al. (2019) [31] compared the population genetic characteristics of 318 samples of FAWs from 131 cities in China and reported that less than 4% of the total samples were C-strain based on the mitochondrial COI gene, whereas the haplotype characteristics of the Tpi gene indicated that all the samples were “C-strain”. In addition, the frequencies of hybrids were comparable to those observed in previous studies of FAWs in America, Africa and Australia [1,3,32,33]. For example, the agreement of both mitochondrial (COI gene) and nuclear (Tpi gene) markers as ‘corn-strain’ haplotypes occurred in 2 samples of the 48 samples in northern Australia [3]. These findings suggested that these populations were generated from an interstrain hybrid of an R-strain female and a C-strain male, which raised concerns about the potential risk of FAWs to rice [30]. The consistent preponderance of the CO I R-strain Tpi C-strain discordant type suggests that a directional bias in interstrain mating favoring CO I R-strain females may be a significant contributor to the distribution of fall armyworm genotypes found in the wild [32].
It has been reported that the R-strain FAW exhibit strong preference towards oviposition on grass and C-strain females oviposited indiscriminately on host plants and artificial structures [34], whereas Ingber et al., (2021) [13] suggested that maize is the preferred oviposition host in both of the R-strain and C-strain in America. As for the hybrid population of FAW that invaded China, they have been reported mainly damages maize in field and oviposition choice of female adults preferred maize over other host plants [35]. However, the FAWs female adult they used for oviposition preference came from the colony that reared on artificial diet or maize. In our case, FAWs were found damaging rice plants in fields, and female adults preferred laying eggs on maize rather than on rice plants as well (Figure 4). Therefore, diet of rice plant of FAWs larvae did not altered oviposition preference of female adults, which was also observed for the 1st and 2nd generations of FAWs when larvae continually reared on rice plants [36].
Survival rate, developmental duration and fecundity are important measures of host fitness [37]. The population of FAWs established in China can survive on rice plants under laboratory conditions; for example, Hafeez et al. (2021) [36] continuously reared FAW larvae on rice plants (cv. Liangyou-887) for 20 generations, and Guo et al. (2022) [38] maintained the FAW colony on rice plants (cv. Taichung Native 1) for at least 13 generations. We found that FAW larvae were unable to survive when fed 4-week-old rice plants, whereas they could complete their life cycle on 2-week-old rice plants, with the total survival rate was 8% (Figure 5 and Figure 6, Table 1). During the experiments, we found that rice leaf at 4-week-old had harder and rougher surface than the leaf at 2-week-old, which might be responsible for the lower survival rate of FAW larvae on 4-week-old rice leaf. Moreover, our findings showed that the population damaging rice had even lower survival rates than that of previous studies [36,38], which may be due to the resistance of different rice varieties to FAWs.
When under no-choice experiments in laboratory, FAW can complete its life cycle on a wide range of crops, such as maize Z. mays, kidney bean Phaseolus vulgaris, soybean Glycine max, oilseed rape Brassica napus, sunflower Helianthus annuus, peanut Arachis hypogaea, buckwheat Fagopyrum esculentum, and ginger Zingiber officinale [39,40,41]. A life table study is crucial to understand population dynamics and forecast future population levels. In our life table study, the pre-adult- and pupal-stage durations were prolonged, and the fecundity of adult females decreased when fed on rice (Figure 6 and Figure 7). The result was consistent with FAW individuals damaging maize in China that they exhibited better fitness on maize than rice in the laboratory [36,38], and the authors revealed that rice-fed FAWs exhibited stronger feeding preferences, greater larval survival and greater pupal weight on maize plants than on rice plants. Our results indicated that maize is more suitable than rice for the FAW populations presented on rice. FAW larvae normally complete six larval instar stages, but they may complete between six and seven larval instar stages depending on temperature and host plant availability [42]. Compared with the curves for the age-stage survival rate of various FAW instar larvae on maize at 26 °C [43], the FAWs collected from the paddy field had seven instars and the pre-adult-stage duration were prolonged (Table 1). Seventh instar larvae were also found when FAW feeding on soybean and sunflower plants [41]. The result could be explained by when FAW was continually fed on rice plants, compared with FAW that fed on maize, the detoxification genes were highly expressed [36,44], which indicating the rice-fed FAW was under pressure and metabolic energy is diverted to detoxification rather than being used for growth and development [45].
Behavior and physiology of FAW could change when there are no other suitable hosts [36,40,46]. The adaptations of herbivorous insects to host plants involves key morphological, behavioral and physiological changes [47,48]. Although FAWs from paddy fields exhibited low fitness when feeding on rice, their fitness for rice could increase rapidly over a few generations, with phenotypic and genetic changes in traits related to host plant adaptation. For example, when larvae continued to be reared on rice, adult females preferred laying eggs on maize in the 1st and 2nd generations and from the 4th generation, adult females showed significant orientation to rice as compared to corn [36]. After we found FAW damaging rice in 2021, insecticides and pheromone traps were used to control the local population of FAW, and until recently, no FAW-damaged rice was found again at those sites. Future study could focus on comparing the behavioral and physiological differences of FAW presented on rice and maize.
5. Conclusions
Our results suggest that the presence of FAW on rice was likely due to the absence of other suitable hosts. The FAWs might be at risk for endangering rice if they have no choice among host plants to provide food for a few generations, so populations of FAWs in paddy fields should be closely tracked.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14102344/s1, Figure S1: The TS colony of FAW maintained on rice seedlings (A), the FAW larvae feeding on maize or rice leaf at 24 days after hatching in life table observation (B) and the pupa of FAW when fed on maize or rice leaf at the larval stage in life table observation (C).
Author Contributions
Conceptualization, P.L. and L.Z.; Methodology, P.L. and L.Z.; Software, X.P. and H.S.; Validation, X.P.; Formal analysis, P.L.; Investigation, P.L.; Resources, D.S.; Data curation, D.S.; Writing—original draft, P.L.; Writing—review & editing, L.Z. and J.Z.; Visualization, H.S.; Supervision, J.Z.; Project administration, Q.Y.; Funding acquisition, Q.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Innovation Foundation Program of Guangdong Academy of Agricultural Sciences (No. 202206) and the Guangzhou Science and Technology Project (No. 202206010150 and No. 2023A04J0804).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nagoshi, R.N.; Goergen, G.; Tounou, K.A.; Agboka, K.; Koffi, D.; Meagher, R.L. Analysis of Strain Distribution, Migratory Potential, and Invasion History of Fall Armyworm Populations in Northern Sub-Saharan Africa. Sci. Rep. 2018, 8, 3710. [Google Scholar] [CrossRef] [PubMed]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.J.W.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest. Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Piggott, M.P.; Tadle, F.P.J.; Patel, S.; Gomez, K.C.; Thistleton, B. Corn-strain or Rice-strain? Detection of Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), in northern Australia. Int. J. Trop. Insect Sci. 2021, 41, 2607–2615. [Google Scholar] [CrossRef]
- Sun, X.; Hu, C.; Jia, H.; Wu, Q.; Shen, X.; Zhao, S.; Jiang, Y.; Wu, K. Case Study on the First Immigration of Fall Armyworm, Spodoptera frugiperda Invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Huang, Y.; Dong, Y.; Huang, W.; Ren, B.; Deng, Q.; Shi, Y.; Bai, J.; Ren, Y.; Geng, Y.; Ma, H. Overwintering Distribution of Fall Armyworm (Spodoptera frugiperda) in Yunnan, China, and Influencing Environmental Factors. Insects 2020, 11, 805. [Google Scholar] [CrossRef]
- Nagoshi, R.N. The Fall Armyworm Triose Phosphate Isomerase (Tpi) Gene as a Marker of Strain Identity and Interstrain Mating. Ann. Entomol. Soc. Am. 2010, 103, 283–292. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Silvie, P.; Meagher, R.L.; Lopez, J.; Machado, V. Identification and Comparison of Fall Armyworm (Lepidoptera: Noctuidae) Host Strains in Brazil, Texas, and Florida. Ann. Entomol. Soc. Am. 2007, 100, 394–402. [Google Scholar] [CrossRef]
- Murúa, M.G.; Nagoshi, R.N.; DosSantos, D.A.; Hay-Roe, M.M.; Meagher, R.L.; Vilardi, J.C. Demonstration Using Field Collections that Argentina Fall Armyworm Populations Exhibit Strain-specific Host Plant Preferences. J. Econ. Entomol. 2015, 108, 2305–2315. [Google Scholar] [CrossRef]
- Groot, A.T.; Marr, M.; Schöfl, G.; Lorenz, S.; Svatos, A.; Heckel, D.G. Host Strain Specific Sex Pheromone Variation in Spodoptera frugiperda. Front. Zool. 2008, 5, 20. [Google Scholar] [CrossRef]
- Schöfl, G.; Heckel, D.G.; Groot, A.T. Time-shifted Reproductive Behaviours among Fall Armyworm (Noctuidae: Spodoptera frugiperda) Host Strains: Evidence for Differing Modes of Inheritance. J. Evol. Biol. 2009, 22, 1447–1459. [Google Scholar] [CrossRef]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of Field-evolved Resistance to Bacillus thuringiensis-derived Cry1F δ-endotoxin in Spodoptera frugiperda Populations from Argentina. Pest. Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.A.; Mason, C.E.; Flexner, L. Cry1 Bt Susceptibilities of Fall Armyworm (Lepidoptera: Noctuidae) Host Strains. J. Econ. Entomol. 2018, 111, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.A.; McDonald, J.H.; Mason, C.E.; Flexner, L. Oviposition Preferences, Bt Susceptibilities, and Tissue Feeding of Fall Armyworm (Lepidoptera: Noctuidae) Host Strains. Pest. Manag. Sci. 2021, 77, 4091–4099. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Koffi, D.; Agboka, K.; Tounou, K.A.; Banerjee, R.; Jurat-Fuentes, J.L.; Meagher, R.L. Comparative Molecular Analyses of Invasive Fall Armyworm in Togo Reveal Strong Similarities to Populations from the Eastern United States and the Greater Antilles. PLoS ONE 2017, 12, e0181982. [Google Scholar]
- Wang, Y.; Xue, Y.; Li, J. Towards Molecular Breeding and Improvement of Rice in China. Trends Plant Sci. 2005, 10, 610–614. [Google Scholar] [CrossRef]
- Pantoja, A.; Smith, C.M.; Robinson, J.F. Effects of the Fall Armyworm (Lepidoptera: Noctuidae) on Rice Yields. J. Econ. Entomol. 1986, 79, 1324–1329. [Google Scholar] [CrossRef]
- Valdez, E.; Joshi, R.; Rillon, G.; Donayre, D.K.; Martin, E.; Dela Cruz, K.; Sandoval, F.; Faheem, M.; Sivapragasam, A. Rice: A New Host of Fall Armyworm Spodoptera frugiperda (J.E. Smith) and Its Strains in the Philippines. Insect Environ. 2023, 26, 129–136. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. 2024. Available online: https://www.fao.org/faostat/ (accessed on 18 September 2024).
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on Law of Diffusion Damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Yang, J.; Tao, Y.; Liu, Q.; Zheng, Z.; Zhou, H. Report of Fall Armyworm Infesting Rice Seedlings in Wuxue, Hubei Province. China Plant Prot. 2020, 40, 44–45. (In Chinese) [Google Scholar]
- Zhang, H. First Report and Biotype Identification of Fall Armyworm Damaging Rice Seedlings in Yunxiao, Fujian Province. China Plant Prot. 2020, 40, 41–43+53. (In Chinese) [Google Scholar]
- Li, Y.H.; Zhou, W.W.; Wu, H.F.; Shen, Y.F.; Zheng, K.M.; Chen, Y.P.; Lu, M.H. Occurrence and Damage Dynamics of Spodoptera frugiperda in Yunnan Province from 2019 to 2021. China Plant Prot. 2024, 44, 43–48. (In Chinese) [Google Scholar]
- Chi, H.; Liu, H. Two New Methods for the Study of Insect Population Ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table Analysis Incorporating Both Sexes and Variable Development Rates Among Individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-Stage, Two-Sex Life Tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and Its Host Myzus persicae (Sulzer) (Homoptera: Aphididae) with Mathematical Proof of the Relationship between Female Fecundity and the Net Reproductive Rate. Env. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.; Lee, C.; Chi, H. Population and Damage Projection of Spodoptera litura (F.) on Peanuts (Arachis hypogaea L.) under Different Conditions Using the Age-Stage, Two-Sex Life Table. Pest. Manag. Sci. 2014, 70, 805–813, Erratum in Pest. Manag. Sci. 2014, 70, 1936. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2016.
- Juárez, M.L.; Schöfl, G.; Vera, M.T.; Vilardi, J.C.; Murúa, M.G.; Willink, E.; Hänniger, S.; Heckel, D.G.; Groot, A.T. Population Structure of Spodoptera frugiperda Maize and Rice Host Forms in South America: Are They Host Strains? Entomol. Exp. Appl. 2014, 3, 182–199. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Fleischer, S.; Eagher, R.L. Demonstration and Quantification of Restricted Mating Between Fall Armyworm Host Strains in Field Collections by SNP Comparisons. J. Econ. Entomol. 2017, 110, 2568–2575. [Google Scholar] [CrossRef]
- Zhang, L.; Jin, M.; Zhang, D.; Jiang, Y.; Liu, J.; Wu, K.; Xiao, Y. Molecular Identification of Invasive Fall Armyworm Spodoptera frugiperda in Yunnan Province. Plant Prot. 2019, 45, 19–24+56. (In Chinese) [Google Scholar]
- Zhang, L.; Liu, B.; Jiang, Y.; Liu, J.; Wu, K.; Xiao, Y. Molecular Characterization Analysis of Fall Armyworm Populations in China. Plant Prot. 2019, 45, 20–27. (In Chinese) [Google Scholar]
- Jacobs, A.; Vuuren, A.V.; Rong, I.H. Characterization of the Fall Armyworm (Spodoptera frugiperda J.E. Smith) (Lepidoptera: Noctuidae) from South Africa. Afr. Entomol. 2018, 26, 45–49. [Google Scholar] [CrossRef]
- Nagoshi, R.N. Evidence that A Major Subpopulation of Fall Armyworm Found in the Western Hemisphere is Rare or Absent in Africa, which may Limit the Range of Crops at Risk of Infestation. PLoS ONE 2019, 14, e0208966. [Google Scholar] [CrossRef] [PubMed]
- Meagher, R.L.; Nagoshi, R.N.; Stuhl, C.J. Oviposition Choice of Two Fall Armyworm (Lepidoptera: Noctuidae) Host Strains. J. Insect Behav. 2011, 24, 337–347. [Google Scholar] [CrossRef]
- Wang, W.W.; He, P.Y.; Liu, T.X.; Jing, X.F.; Zhang, S.Z. Comparative Studies of Ovipositional Preference, Larval Feeding Selectivity, and Nutritional Indices of Spodoptera frugiperda (Lepidoptera: Noctuidae) on 6 Crops. J. Econ. Entomol. 2023, 116, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.; Li, X.; Ullah, F.; Zhang, Z.; Zhang, J.; Huang, J.; Khan, M.M.; Chen, L.; Ren, X.; Zhou, S.; et al. Behavioral and Physiological Plasticity Provides Insights into Molecular Based Adaptation Mechanism to Strain Shift in Spodoptera frugiperda. Int. J. Mol. Sci. 2021, 22, 10284. [Google Scholar] [CrossRef]
- Moreau, J.; Benrey, B.; Thiéry, D. Grape Variety Affects Larval Performance and also Female Reproductive Performance of the European Grapevine Moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2006, 96, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Jin, R.; Guo, Z.; Cai, T.; Zhang, Y.; Gao, J.; Huang, G.; Wan, H.; He, S.; Xie, Y.; et al. Insecticide Susceptibility and Mechanism of Spodoptera frugiperda on Different Host Plants. J. Agric. Food Chem. 2022, 70, 11367–11376. [Google Scholar] [CrossRef]
- Li, D.Y.; Zhi, J.R.; Zhang, T.; Ye, J.; Liang, Y. Effects of Different Host Plants on the Development and Reproduction of Spodoptera frugiperda. J. Environ. Entomol. 2020, 42, 311–317. [Google Scholar]
- Xie, W.; Zhi, J.; Ye, J.; Zhou, Y.; Li, C.; Liang, Y.; Yue, W.; Li, D.; Zeng, G.; Hu, C. Age-stage, Two-sex Life Table Analysis of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) Reared on Maize and Kidney Bean. Chem. Biol. Technol. Agric. 2021, 8, 44. [Google Scholar] [CrossRef]
- He, L.; Wu, Q.; Gao, X.; WU, K. Population Life Tables for the Invasive Fall Armyworm, Spodoptera frugiperda Fed on Major Oil Crops Planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Luginbill, P. The Fall Army Worm; US Department of Agriculture: Washington, DC, USA, 1928; pp. 2–7.
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of Larval Performance and Oviposition Preference of Spodoptera frugiperda among Three Host Plants: Potential Risks to Potato and Tobacco Crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef]
- Han, W.K.; Tang, F.X.; Yan, Y.Y.; Wang, Y.; Liu, Z.W. Plasticity of the Gene Transcriptional Level and Microbiota in the Gut Contributes to the Adaptability of the Fall Armyworm to Rice Plants. J. Agric. Food Chem. 2021, 47, 18546–18556. [Google Scholar] [CrossRef] [PubMed]
- Simelane, D.O.; Steinkraus, D.C.; Kring, T.J. Predation Rate and Development of Coccinella septempunctata L. Influenced by Neozygites fresenii-Infected Cotton Aphid Prey. Biol. Control 2008, 44, 128–135. [Google Scholar] [CrossRef]
- Sotelo-Cardona, P.; Chuang, W.P.; Lin, M.Y.; Chiang, M.Y.; Ramasamy, S. Oviposition Preference Not Necessarily Predicts Offspring Performance in the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) on Vegetable Crops. Sci. Rep. 2021, 11, 15885. [Google Scholar]
- Orsucci, M.; Audiot, P.; Dorkeld, F.; Pommier, A.; Vabre, M.; Gschloessl, B.; Rialle, S.; Severac, D.; Bourguet, D.; Streiff, R. Larval Transcriptomic Response to Host Plants in Two Related Phytophagous Lepidopteran Species: Implications for Host Specialization and Species Divergence. BMC Genom. 2018, 19, 265. [Google Scholar] [CrossRef]
- Cantón, P.E.; Bonning, B.C. Transcription and Activity of Digestive Enzymes of Nezara viridula Maintained on Different Plant Diets. Front. Physiol. 2019, 10, 1553. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).