Abstract
The increasing demand for bioactive compounds from plants has spurred the exploration of innovative strategies to enhance their production. This review focuses on three advanced methodologies: elicitation, metabolic engineering, and using biosynthesized nanoparticles. Elicitation, through the application of abiotic and biotic stressors, stimulates plant defense mechanisms, and significantly boosts secondary metabolite production. Metabolic engineering further refines this process by genetically modifying key pathways to redirect resources towards the synthesis of valuable compounds. Additionally, biosynthesized metallic and metallic oxide nanoparticles offer a sustainable method to enhance plant growth and metabolite production. These nanoparticles act not only as elicitors, but also catalyze metabolic processes and improve the efficiency of bioactive compound synthesis. Despite significant advancements, challenges remain in optimizing these approaches for large-scale agricultural application, particularly in understanding the precise interactions between nanoparticles and plant systems. The integration of these strategies holds great potential for sustainable agriculture and the development of high-value phytochemicals. Future research should focus on unraveling the complex mechanisms behind these interactions and refining these methods for commercial use, ensuring their environmental and economic sustainability.
1. Introduction
The production of bioactive compounds in plants is crucial for various industries, including pharmaceuticals, nutraceuticals, and cosmetics. Recent advancements have introduced several innovative tools and techniques to enhance the yield and quality of these compounds [1,2]. The production of bioactive compounds in plants can be significantly enhanced by cyclodextrins [3], LED lighting [4], microbial cell factories [5], modern pretreatment methods [6], Temporary Immersion System (TIS) bioreactors [7], Plant Cell Suspension Culture (CSC) in bioreactors [8], and abiotic stress conditions [9], with all of them offering promising solutions to boost the yield and quality of these valuable compounds. These innovations not only improve efficiency, but also support sustainable and environmental friendly practices in the production of bioactive compounds.
The presence of these bioactive compounds is derived from the secondary metabolism activation of the plants. Plant secondary metabolites (PSMs) are organic compounds not directly involved in the primary growth, development, or reproduction of plants, but play crucial roles in plant defense, stress responses, and ecological interactions [9]. Plants produce or increase the concentration of secondary metabolites like terpenoids, alkaloids, and phenolic compounds in response to abiotic stresses such as high temperatures, drought, and UV radiation [10]. One of the objectives of researchers worldwide is to have a regular and high-quality plant production, regardless of all the factors that might alter the final product, such as climate change, water scarcity, and the increasing demand for biofuels, meat, and dairy, projecting a strain on agricultural production and food security [11,12,13].
It is important to know and understand how the combined effects of stresses are crucial for developing resilient plant varieties [14,15]. Uncontrolled stress affects several physiological and biochemical processes in plants, including photosynthesis, respiration, and nutrient uptake, leading to reduced growth and development [16,17].
The increasing demand for bioactive compounds has driven significant research into their production in plants through several methods, for example, by the elicitation technique, which is the process of stimulating the production of secondary metabolites and enhancing plant defense mechanisms through the application of biotic or abiotic agents known as elicitors [18]. Elicitation, in general, significantly boosts the production of secondary metabolites such as glucosinolates, aminobutyric acid, phenolic compounds, and tropane alkaloids in various plants, enhancing their functional properties and biological activities, such as antioxidant activity [19]. Another proposal of how to increase the bioactive compounds in the plants has been metabolic engineering, which offers another promising strategy by directly manipulating the biosynthetic pathways within plants [20]. Through this process, genetic modification, specific enzymes and pathways can be targeted to enhance the flow of precursors toward the desired bioactive compounds [21].
Nevertheless, these efforts have not been enough, and this is where nanotechnology can be applied as a powerful tool: as nanofertilizers, biostimulants, elicitors, eustressors, or as boosting tools to enhance natural plant defenses via bioactive metabolism [22,23,24]. The need for more efficient and sustainable approaches has led to the exploration of the use of nanoparticles (NPs) in elicitation and metabolic engineering processes [25,26]. The advent of nanotechnology has introduced a new dimension to this field, along with biosynthesized metallic and metallic oxide nanoparticles, derived through environmental friendly methods, showing great potential in enhancing plant growth and secondary metabolite production [27,28,29,30].
This review aims to explore the synergistic potential of elicitation, metabolic engineering, and biosynthesized nanoparticles in boosting bioactive compound production in plants, along with the key challenges in integrating these tools. While the benefits are well documented, further research is needed to fully understand their mechanisms to optimize their application for commercial use and to highlight their role in sustainable agriculture. The information selected to be part of this work followed the following criteria: (i) published in the past decade, (ii) review articles, to provide a broader understanding of existing knowledge and trends across relevant subfields, (iii) original research articles, with emphasis on experimental studies that present empirical data regarding the biosynthesis, application, and the effects of nanomaterials on plant systems; the (iv) keywords Elicitation, Green Synthesis, Nanomaterials, Bioactive Compounds, Plant Metabolism, Metallic Nanoparticles, Oxide Nanoparticles, Sustainability, Agricultural Applications, and Nanotechnology in Agriculture; and (v) where searches were conducted in reputable scientific databases such as PubMed, Scopus, and Web of Science.
2. Elicitation Techniques
Population growth, over-exploration of croplands, climate change, and the demand for healthy foods have caused agricultural practices to deviate from traditional technologies, making use of new strategies that guarantee optimal crop production that meets food needs, provides in terms of quality, and includes compounds that benefit the consumer, in addition to boosting the agricultural economy and being environmental friendly [31]. Among these new strategies, the use of the elicitation method is presented. The elements that generate these changes are known as elicitors, and can be classified into biotic (microbial extracts, fungal cell wall components, and plant hormones) [18] and abiotic (including physical factors such as UV light, temperature changes, drought, salinity, and chemical agents, for example, heavy metals, salts, and nanomaterials (NMs)) [19]. These kinds of stresses are used to obtain a positive response from the plant’s secondary metabolism, and in this specific case, the elicitor is known as a eustressor. The general idea of using a eustressor is to induce a positive response in the crop regarding yield, growth, and quality, tolerance to diseases, pests, and abiotic factors, and an increase in compounds of interest. Nevertheless, it is essential to understand how eustressors can modify plant secondary metabolism for optimizing general plant adaptation mechanisms [9], because the response varies between plants, the type of stress applied, and environmental conditions [32,33,34]. When the plant´s response is considered negative, the term distress is applied [23,33].
Biotic eustressors, which include biostimulants (such as phytohormones and microorganisms) and abiotic eustressors, such as UV light and acoustic waves, can improve the plant´s essential nutrient production, growth regulators, or protective compounds [35]. However, if the response is the activation of the immune system of the plant, it is called an elicitor regardless of the origin or type of stress factor [23,36,37].
The eliciting action of different eustressors induces a series of complex mechanisms where the plant gives a response that counteracts the effect and allows for an adaptive process to the stress factor through the perception of external signals that trigger a series of signaling, gene transcription, proteins, non-coding RNAs, and secondary metabolites that help the plant to survive [38] (Figure 1). Also, reactive oxygen species (ROS) are pivotal in the plant’s adaptive response to various stressors, functioning as key signaling molecules that trigger complex intracellular and systemic mechanisms. These processes involve intricate interactions with plant hormones, antioxidant systems, and even plant-associated microbiota [39].
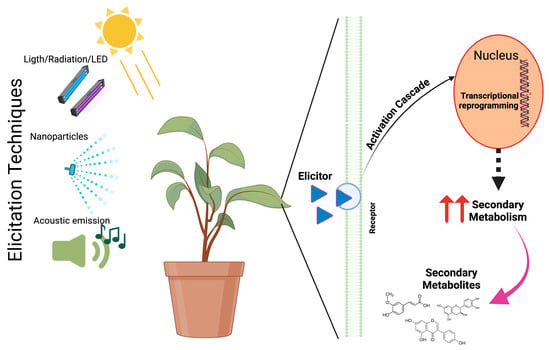
Figure 1.
Schematic representation of the action of elicitation techniques.
Among these strategies the plant uses to adapt and to enhance bioactive compound pro duction, the implementation of light, acoustic emissions, and nanoparticles is currently one example of innovative methods being used. It should be mentioned that although these strategies have yielded positive results in terms of the production of secondary metabolites, it is not yet possible to generalize the results [4]. Additionally, there are limitations regarding the application of these methodologies in open fields or in extensive crop areas, as well as in the interaction of some with the environment, where their interaction has not yet been clarified. However, the results obtained indicate that the application of these tools can promote sustainable agriculture [40].
Research on bioactive compounds focuses specially on secondary metabolites, which are compounds that, in the plant system, contribute to pollination, repel pests, stimulate communication between crops, and allow for adaptation to stress factors [41]. As has been mentioned, novel strategies seek to increase the production of these compounds. In recent years, emphasis has been placed on those factors that are easily to apply, regardless of the nature of the eustressor (biotic or abiotic). For example, this includes abiotic factors such as radiation (UV, LED) [4,42,43,44,45,46,47,48,49], electric fields [33,50], sound waves [32,51,52], and nanomaterials [53,54,55,56,57,58,59,60,61,62].
2.1. Physical and Chemical Factors for Secondary Metabolites Production
Plants produce different metabolites, and some of them are involved in plant development and growth, such as carbohydrates, lipids, and proteins, known as primary metabolites [32]. Other compounds are produced as a plant defense mechanism or as a response to stress conditions, and are called secondary metabolites [63]. Secondary metabolites include flavonoids, terpenoids, alkaloids, and phenols, and are compounds that can increase in concentration in the plant through elicitation techniques [64]. Elicitation, as has been mentioned, consists in exposing the plant to factors (biotic or abiotic stressors) which trigger a series of signal transduction pathways, generally leading to the production of active oxygen species (AOS), the synthesis of defense enzymes, and the activation of metabolite biosynthesis to increase secondary metabolite production [55,65]. In Table 1, some examples of eustressor effects are shown, depending on abiotic factors (physical or chemical).

Table 1.
Examples of eustressor effects classified by physical or chemical factor.
Elicitation by physical or chemical factors triggers a series of mechanisms that induce various signals in the plant, such as interaction between receptors and the triggering of signaling cascades, such as the mitogen-activated protein kinase (MAPK) pathway and the activation of NADPH oxidase, responsible for the production of reactive oxygen and nitrogen species, which in turn triggers a defense mechanism involving the expression of defense genes as well as the activation of enzymes related to the synthesis of secondary metabolites, such as the phenylalanine ammonia lyase (PAL) enzyme. The above has been studied and documented in the growth, development, physiology, and biochemistry of plants, providing advantages in terms of the production of secondary metabolites. However, the effect of these factors is still not fully understood, which represents a disadvantage given that the effects induced by the elicitors will depend on the species used, the type of elicitor, the frequency of the application or interaction, the level or dosage, and the specific characteristics of each elicitor used [83].
During plant growth and development, the plants are exposed to various physical factors, inducing a response at the molecular level; this response includes the activation of receptors and sensors that allow plants to detect signals and respond defensively to protect themselves, resulting in the synthesis of secondary metabolites [36,84].
For example, light/radiation plays a fundamental role in photosynthesis and the regulation of the production of secondary metabolites, in addition to the fact that plants have sensitive and precise detection capabilities for different wavelength ranges, including ultraviolet (UV) light and solar radiation [85,86]; therefore, plant development and growth are related to photomorphogenic development, which can be altered by light characteristics [87]. Also, light/radiation can induce oxidative damage and thus increase reactive oxygen species (ROS), consequently generating oxidative stress that induces signaling cascades such as the activation of the endogenous enzyme system or the activation of the pentose phosphate pathway involved in producing secondary metabolites [55]. Additionally, plants have developed adaptive and exploitative systems, both of which are to protect themselves from high doses of UV radiation and to maximize the absorption of usable wavelengths for the regular photosynthesis process [88,89,90].
Searching for a better way to control the amount and specificity of the wavelength delivered to the plant for the optimal development, growth, and production of secondary metabolites, implementation of light emitting diodes (LEDs) is increasing. The LED used for crops in closed, and controlled environments have demonstrated an eliciting action [49], improving the quality and quantity of horticultural productivity in the Mediterranean basin in the winter season [91,92], with specific effects on the concentration of carotenoids [46], polyphenols [47], and other compounds [93]. However, there is variability in the results in the vast diversity of plant species, so a universal model of LED lighting is not yet known. Further studies are needed to discover the eliciting role of LED lights and radiation in producing secondary metabolites in plants.
In general, magnetic fields are force fields created because of the movement of electric flow, and are expressed in Gauss (G) or Tesla (T). The magnetic fields (Vm−1) are an environmental factor that plants experience since they are part of the Earth´s magnetic field [94]. However, it has been shown that magnetic fields act as abiotic eustressors, and can induce membrane permeability, generating changes in electrical charges and the concentration of ions and free radicals. The free movement of ions through the plant´s membrane ion channels triggers metabolic responses that improves the physiological and biochemical response, for example, the increase in the germination rate and vigor of seedlings of many crops [95,96]. These responses increase the production of ROS and the initiation of oxidative stress, inducing the activation of antioxidant enzymes and the expression of their genes [97]. Studies indicate that the mechanisms induced by magnetic fields, such as the induction of phytohormone syntheses, generates positive effects on the germination, growth, and development of plants, in addition to mitigating the impact generated by other abiotic stress, such as irradiation, salinity, and temperature [78,80].
Although there are various studies on the effect of magnetic fields on plants, these have focused mainly on their use as a preparatory treatment for seeds before sowing. Magnetic fields have been shown to influence germination processes and seedling growth, in addition to being safe for the environment; for this reason, this type of treatment is an area of opportunity for agricultural practice. It offers advantages by being a non-destructive treatment, easy to apply, and friendly to the environment, in addition to increasing the germination rate and vigor of plants [95,96].
Another physical factor is sound, which is defined as a series of longitudinal waves of pressure that propagate through compressible media, such as air, liquids, and solids; its units of measurement are Hertz (Hz) or kilo Hertz (kHz). When the sound has a frequency and sufficient sound pressure, it is called a perceived sound and can be measured by means of acoustic emissions. The application of acoustic emissions at different frequencies, pressure levels, durations, and repetitions of exposure periods has been proven to have an influence on plant growth, germination, and development [98].
Acoustic emissions can trigger signaling and transduction cascades, such as stress-related gene expression and energy metabolism, and increase secondary metabolites [32]. Appel and Cocroft [99] demonstrated that the sounds emitted by insect chewing and their subsequent application to plants increase the production of secondary metabolites involved in the defense response. While Jeong et al. [100] improved natural protection in crops by applying a 100 dB amplification of a particular frequency range (0 to 15 kHz), Bhandawat et al. [51] demonstrated that specific frequencies induce a positive response at the genetic level to heat stress situations, implying a conditioning response and excellent resistance to future stress factors. Although studies indicate an eliciting effect on acoustic emissions, further research is needed on these effects on different crops. Because acoustic emissions applied on a larger scale should be monitored regarding environment modification, it has nevertheless been noticed that the noise spectra for water streams and rivers, water breaking the surface, and waterfalls can be measured in the range 0–500 Hz [82], which led the researchers of this area to try to propose new measurement methods to confirm that acoustic emissions modify plant defense mechanisms and their general growth development.
Additionally, there are areas of interest and importance to ecological innovation that have not been fully clarified, for example, finding and applying acoustic waves in a controlled manner to resemble some type of abiotic or biotic stress, and thereby make the plant more resistant [101]. In addition, we provide information regarding the proposed hypothesis that plants emit sounds, such as that proposed by Telewski [102], which indicates that plants use similar mechanisms to those used by mechanochemical enzymes such as myosins, which use the chemical energy of the hydrolysis of adenosine triphosphate (ATP) on actin and thereby induce mechanical vibrations within cells. In this context, there are several unknowns that must be resolved; however, the application of this strategy provides advantages by involving a clean methodological environment, not being expensive, and featuring positive results that have been observed when it is applied. On the other hand, there is still a lack of information on the impact of acoustic waves on the environment and the management of their application in large areas, since all the studies that have been carried out or developed have focused on small and controlled spaces.
Along with physical factors, there are the chemical factors, which are also used to enhance secondary metabolite production. The advances in nanotechnology through recent decades have allowed them to have a wide range of applications, including in agriculture. The structural and physicochemical characteristics of the nanomaterials allows them to interact at the plant´s intracellular level, triggering signaling cascades involved in growth, development, physiology, and biochemical mechanisms [55]. Nanoparticles have been used as elicitors to produce secondary metabolites. For example, the application of copper nanoparticles (Cu NPs) induces an increase in antioxidant capacity, as well as an increase in the production of compounds of interest, such as phenols, flavonoids, alkaloids, and saponins in a hydroponic culture of Bacopa monnieri L. [103]. The application of silver nanoparticles (AgNPs) in Celastrus paniculatus roots increased the levels of phytochemical celastrol, which is of interest for its therapeutic properties [104].
Additionally, it has been observed that nanomaterials not only act on the plant, but also interact with its ecosystem. For example, it has been reported that SiO2 NPs can reduce cadmium (Cd) uptake and arsenic (As) accumulation in both pot and field experiments, alleviating heavy metal element-induced oxidative stress in crops [105]. Additionally, SiO2 NPs can eliminate unwanted weeds and insects with less cost and energy, possessing great potential for agricultural sustainability [106]. On the other hand, different types of nanoparticles and their derivatives have attracted considerable attention for their potential antimicrobial activity and their possible application as agricultural fungicides. In this sense, strategies have been developed to produce green nanofungicides manufactured from biodegradable materials, including enzymes, nutrients, polysaccharides, plant extracts, biodegradable polymers, and microorganisms. Microorganisms such as bacteria, algae, yeasts, fungi, actinomycetes, and viruses are used in the bio-mediated synthesis of nanometals. Biofungicides based on nanotechnology can offer a cheap, efficient, and eco-friendly strategy for pest control in agriculture. Nanoformulations of biofungicides protect the timely degradation of the environment and have a significant impact on target species. Less environmental contamination and safe handling are advantages of nanofungicides [107]. Despite the advances that have been obtained in recent years, nanotechnology in agriculture is still in its developmental stage. The effect of nanomaterials on plants and the ecosystem surrounding them has yet to be fully discovered, and there is a wide scope for the development of precise nano-agrochemicals with respect to their containment in suitable carrier materials, manufacturing techniques, and modes of application, and, above all, the evaluation of the safety and toxicological implications of nanoproducts in the plant, its environment, and the crop and its final products.
2.2. Recent Studies and Advancements
Plants are a source of phytochemicals (primary and secondary metabolites) [108]. Among the secondary metabolites, we can find phenols, flavonoids, terpenoids, alkaloids, etc. These secondary metabolites are of human interest, since they can be used in diverse industries such as pharmaceuticals, food, medicine, and others [109]. One of the limitations of obtaining these compounds and meeting their demand is that secondary metabolites are available in small quantities, so it is necessary to look for strategies to increase their production in plants. As has been mentioned, elicitation is a strategy that has yielded promising results. However, the variability of the response between plants/vegetables/crops, type of elicitation, dosage, exposure time, and cultivation time, among each other factors, is seen as a challenge to establishing a universal elicitation model.
Nevertheless, some strategies have allowed for better control of the production of secondary metabolites in recent years. In this sense, plant cell cultures (in vitro techniques) greatly address these challenges, especially in propagating high-value commercial crops [110]. Despite some restrictions, such as challenges to expanding production, instability in biosynthesis, and inadequate yields in in vitro plant models, some advances have been found via the elicitation process with nanoparticles in in vitro plant systems. Al-Oubaidi and Kasid [111] demonstrated a statistically significant increase in the levels of phenols and flavonoids when supplementing the growth medium of Ciner arietinum callus with titanium dioxide nanoparticles (TiO2 NPs) at concentrations of 4.5 and 6 mg L−1. Kruszka et al. [112] demonstrated the potential of nanoparticles to stimulate the synthesis of secondary metabolites by treating cell suspension cultures of Hypericum perforatum L. with metal (Ag, Au, Cu, Pd) and metal oxide (CeO2, CuO, TiO2, ZnO) nanoparticles, thereby obtaining several types of secondary metabolites.
On the other hand, as well as in in vitro systems, and in vertical or containerized farming, LED elicitation in protocorm-like-body (PLB) cultures of a Dendrobium hybrid orchid indicated that pretreatment with white LEDs for more than three subculture cycles, and subsequent exposure with blue-red LEDs (1:1), increased flavonoid content. In contrast, high-intensity green LEDs enhanced phenol accumulation, and low-intensity white LEDs increased antioxidant activity [113]. Specific plants have empathetic and precise sensing abilities for various ranges of wavelengths. Thereby, light emitting diodes (LEDs) allow for the radiation of a narrow light spectrum to meet the requirements of plants for their growth and development. Lian et al. [114] demonstrated that LEDs serve as abiotic stress factors, inducing the synthesis of secondary metabolites [115,116].
To the best of our knowledge, there are still several areas to explore regarding the production of secondary metabolites through acoustic emissions elicitation in in vitro systems. But acoustic emission as an elicitation strategy has generated significant interest in recent years due to its innovation and environmental friendliness. In this sense, studies have been carried out on pollinator sounds and nectar production [117], buzz pollination [118], defenses against herbivores, and pollen elimination [119]. Caicedo-López et al. [52] demonstrated that it is possible to record the vibrations generated by plants under water stress, and that the application of these same acoustic emissions to plants without water stress activates the plant´s defense mechanisms and increases the concentration of capsaicin in a Capsicum annuum L. model. These examples indicate the importance of sounds for the plant, as well as their ecological role in it. Sound not only allows for the process of adaptation to stress factors, it allows for communication and anticipation of the situation, generating a language between plants and their awareness of the environment.
Although there are various studies that provide relevant information on the mechanisms that plants develop in situations of biotic and abiotic stress, and this has allowed for the application of physical and chemical factors such as elicitors to create positive changes such as increases in secondary metabolites, higher crop yields, increases in the germination and development processes of the plant, and resistance to pathogens, among other aspects, there are still many questions regarding the application doses, exposure times, types of application, crop responses, environment, toxicity, etc. Therefore, studies are necessary to establish a technological precedent for communication/interaction between elicitors, plants, and secondary metabolites.
3. Metabolic Engineering Approaches
Metabolic engineering aims to improve cellular activity by manipulating metabolism. In this sense, the application of metabolic engineering in agriculture aims to increase the production of secondary metabolites of interest. The strategies developed for this purpose consist in increasing the flow of carbon towards the route responsible for the metabolite of interest or blocking intermediate steps that give rise to alternative products. This is achieved by increasing the activity of a key enzyme or by inhibiting the enzymes responsible for the alternative products. Another strategy consists in blocking the enzymes responsible for the degradation process of the product or inhibiting enzymes that use it for the synthesis of other derivatives. In any of the strategies to be used, it is necessary to clarify the enzyme to be used, the key points where it will act, and the product of interest [120,121,122]. Although metabolic engineering uses recombinant DNA technology to modify enzymatic activity, and thereby obtain an increase or decrease in the activity of an enzyme responsible for the synthesis or degradation of a secondary metabolite of interest, it still needs to allow for enough basic metabolism to provide all the energy for basic needs, as well as the over production of secondary components. It is necessary that the process can be monitored and, in some cases, even quantifiable.
In this sense, the science of “omics” reflects the innovation in various technologies that lead to the study of life processes in their entirety. This science has potential application in multiple sectors, including the environmental, industrial, health, and agricultural sectors, mainly in crop improvement [90]. Within the science of “omics”, metabolomics presents a comprehensive quantitative evaluation of all metabolites of an organism under specific conditions. Metabolomics has several objectives, including investigating the causes of biological effects, such as interactions between plant pathogens and the environment, and the interplay between stress factors or elicitors and plants. To achieve these goals, tools for metabolite separation, detection, and quantification are employed.
Metabolomics and other techniques such as genetic markers provide more comprehensive information on various eliciting factors. Genetic markers provide information on various molecular aspects of the plant, such as genes associated with stress responses. To identify these markers, several DNA-based molecular markers, such as simple sequence repeats (SSR), restricted fragment length polymorphism (RFLP), amplified fragment length polymorphisms (AFLPs), single nucleotide polymorphisms (SNPs), intersequence simple sequence repeats (ISSRs), and others, are used for the detection of germplasm to evaluate stress tolerance and map genes associated with stress response [123]. Sinaga and Marpaung [124] assessed the potential of genetic markers as a tool to study abiotic stress and its relationship with physiological changes in pineapple, providing a comprehensive understanding of the genetic response to abiotic stress and metabolomic expression, and thereby provide alternatives to improve agricultural productivity and the development of stress-resistant varieties.
Chromatographic techniques are essential tools in metabolomics associated with separation and include gas chromatography (GC), high-performance liquid chromatography (HPLC), and capillary electrophoresis (CE). In being sensitive techniques, it is possible to obtain information regarding the identification and quantification of metabolites by detecting them based on their physical properties (mass, absorbance, volatility, size, charge, and hydrophobicity, among others), as well as by using tools such as infrared, UV-visible, fluorescence, nuclear magnetic resonance (NMR), gas chromatography (GC), liquid chromatography (LC), and size exclusion [90]. In this sense, metabolomics contributes to the identification of metabolites by generating metabolic fingerprints and metabolic profiles that can help to identify resistance to pathogens due to overexposure to types of fungicide [125], stress factors (water) [126], traceability, in terms of origin and cultivars [127], fluctuations in metabolites in different physiological states [128], and food safety [129,130], among others.
Elicitation processes can contribute to the inhibition or activation of enzymes involved in metabolic pathways related to the production of secondary metabolites. It is indicated that UV radiation on plant callus causes an increase in nitric oxide (NO) synthesis, nitric oxide synthase, and phenylalanine ammonia lyase activities, leading to flavonoid formation and increased flavonoid levels [115]. In low-temperature conditions, like exposure to −5 °C for 4 h, tea plants (Camellia sinensis) accumulate more glycosylated sesquiterpene and nerolidol glucoside. Their storage was expedited by the enzyme plant glycosyltransferase (UGT91Q2) [131]. Bacteria-derived elicitors are also used to increase the production of secondary metabolites. The root culture of Scopolia parviflora when treated with bacterial extract showed production of scopolamine due to the inhibitory effect of elicitors on gene (H6H-Hyoscyamine 6β-hydroxylase) expression [132]. According to Zhao et al. [133], elicitation with yeast polysaccharide (YPS) effectively increased the hairy root growth and flavonoid (rutin and quercetin) production of Fagopyrum tataricum in a concentration-dependent manner by the stimulation of the phenylpropanoid pathway. The extract of mycelium and the polysaccharide fraction derived from the endophytic root fungus Trichoderma atroviride D16 was used as a biotic elicitor in hairy roots of Salvia miltiorrhiza. The polysaccharide fraction was found to be more active for promoting hairy root growth (increased ~60% higher than the control) and stimulating the biosynthesis of tanshinone (aditerpene) by influencing the expression of genes related to the secondary metabolites’ biosynthetic pathway related to microbe-associated molecular patterns (MAMPs) and damage-associated molecular patterns (DAMPs) [134]. Li et al. [135] observed the positive co-relationship between the accumulation of tanshinone (a diterpene) and the expression of selected genes of the tanshinone biosynthetic pathway, such as 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR), isopentenyl diphosphate isomerase (IPPI), and Geranyl Geranyl diphosphate synthase (GGPPS), in hairy root culture during yeast extract elicitation.
On the other hand, a study demonstrated that accumulation of eight flavonoids, i.e., rutin, neohesperidin, buddleoside, liquiritigenin, quercetin, isorhamnetin, kaempferol, and isoliquiritigenin, in purified chitosan-elicited hairy roots was elevated due to the upregulation of chalcone synthase and flavonoid 3′-hydroxylase genes of the flavonoid biosynthetic pathway [136]. Other elicitors such as methyl jasmonate and salicilic acid, at different time points (6, 12, 24, and 48 h), influenced the accumulation of morphinan alkaloids (thebaine, morphine, and codeine) and the relative expression level of six main regulatory genes of the morphinan pathway (COR, SalAT, SalR, T6ODM, CODM, and Salsyn) [137].
Metabolic engineering, in other words, consists in the process of optimizing native metabolic pathways and regulatory networks, or assembling heterologous metabolic pathways to produce specific molecules. This kind of engineering uses molecular, genetic, and combinatorial approaches. Modifications may include the overexpression of limiting steps, the introduction of heterologous genes, and/or the elimination of competitive pathways [129,138]. These modifications may contribute to the production of secondary metabolites in plants.
4. Biosynthesized Metallic and Metallic Oxide Nanoparticles in Plant Metabolite Production
4.1. Nanotechnology and Nanomaterial (NM) Classification
The excessive increase in food demand due to population growth has caused the agricultural sector to enter a state of stress concerning crop production [139]. Additionally, climate change, environmental pollution, resource scarcity, significant post-harvest losses, and increased demands for water and energy have further impaired the efficiency of crop yields and food distribution [140]. For these reasons, it is imperative to develop new strategies for food production that ensure quality and safety while promoting more efficient and environmentally friendly methods [141]. Nanotechnology has emerged as a continuously evolving field and is extensively researched for its potential applications in agriculture, offering solutions to various agricultural and environmental challenges [61,142]. Nanotechnology involves the manipulation of matter on a molecular scale, forming structures ranging from 1 to 100 nm [143].
Numerous studies have explored how nanomaterials can enhance crop production and protection in agroecosystems. They can increase food yield and quality, detect and manage diseases, monitor plant growth, and minimize crop losses and waste, thereby bolstering global food security [144]. These materials possess innovative properties due to their small size, large surface area, crystalline arrangement, composition, and reactivity, which confer unique properties from those of the bulk material [142,144,145,146]. NMs are classified based on the synthesis process, the origin of the materials, and their structure or dimension [147]. According to their dimensionality, they can be divided into four classes: (i) 0D, with all of their dimensions at the nanoscale, (>100 nm) (nanoparticles, quantum dots); (ii) 1D, with two dimensions at the nanoscale and one dimension not at the nanoscale (nanotubes, nanorods, nanofibers, nanowires); (iii) 2D, with two dimensions larger than the nanoscale (thin films); and (iv) 3D, with three dimensions larger than 100 nm, but with components below 100 nm in size (nanocomposites, bundles, multi-nanolayer-type structures) [145,147,148,149,150,151]. Moreover, NMs can also be classified based on their composition or nature into categories such as carbon-based materials, metal-based materials, and composite-based materials (Figure 2) [152].
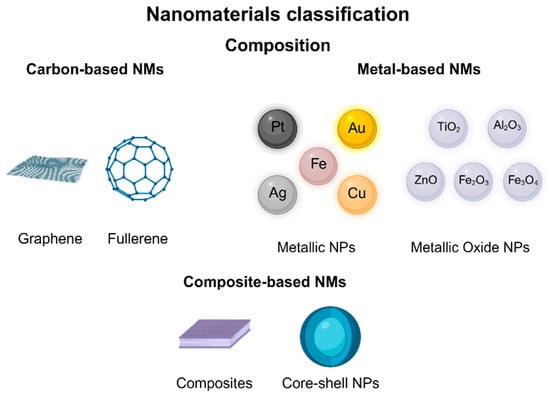
Figure 2.
Nanomaterial classification by composition.
Nanoparticles (NPs) possess unique properties that make them excellent candidates for use in the agricultural sector due to their capacity to be translocated in different parts of the plant and their capacity for ion release and dissolution. Nanoparticles that lack carbon atoms in their structure are known as inorganic nanoparticles, and they are typically defined as those composed of metals or metal oxides [148].
4.2. Metallic and Metallic Oxide Nanoparticles
Metallic nanoparticles (MNPs) exhibit unique properties due to quantum confinement, which alters their electronic and optical characteristics [139]. Combined with their high surface-to-volume ratio, MNPs display enhanced interaction with ultraviolet (UV) and visible light, particularly through the phenomenon known as surface plasmon resonance [146,153], providing valuable information about nanoparticle size, shape, and surface characteristics. Metals such as aluminum (Al), cadmium (Cd), cobalt (Co), titanium (Ti), copper (Cu), gold (Au), iron (Fe), lead (Pb), silver (Ag), and zinc (Zn) are frequently used to obtain these NPs. Typically ranging in size from 10 to 100 nm, these MNPs can be synthesized through top-down or bottom-up methods, resulting in distinctive surface characteristics such as porosity, surface charge, and reactivity [145,148,149,153,154].
Metallic nanoparticles have been used in applications in agriculture (Figure 3), such as antimicrobial agents to inhibit bacterial diseases in crops [57], fertilizers [154], or even as sensors to identify the presence of plant diseases [148,155]. They can also be used to provide essential microelements for plants. In some studies, Zn NPs and Fe NPs have shown effects on the photosynthetic process and growth parameters, depending on the doses used [141,156,157]. Furthermore, Mg, Cu, and Zn NPs have been investigated as antifungal agents [158]; Ca NPs have improved resilience to salinity and plant development [159], and K NPs have been used as nanofertilizers and growth promoters [160].
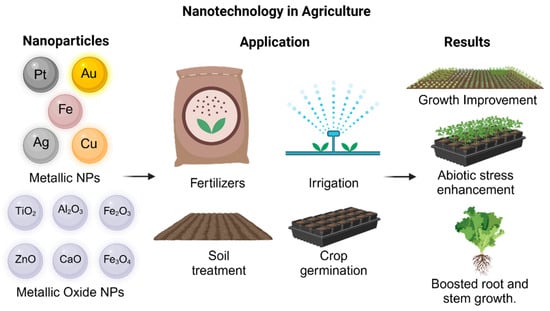
Figure 3.
Metallic and metallic oxide NP application in agriculture.
Some of the most commonly used metal oxide nanoparticles in the agricultural sector (Figure 3) are CaO NPs, which activate enzymes and participate in membrane transport metabolism, nitrate absorption, biomass ratio, and photosynthetic rate [161]; ZnO NPs alleviate zinc deficiency and are easily absorbed by plants due to their small size and large surface area [162]. TiO2 NPs improve soil salinity and increase leaf length, promote cell division, and induce changes in phytohormonal levels by increasing the activity of certain enzymes, promoting nitrate absorption and the transformation of inorganic nitrogen to organic nitrogen, mobilizing nutrients, improving tolerance to biotic and abiotic stress, improving nitrogen assimilation, and changing the response to sunlight [60,67]. SiO2 NPs generate some resistance against pathogens and abiotic stress factors, and they induce biochemical and physiological processes, improving plant growth and yield. Also, they increase lignification, biomass, flowering, nutrient and water absorption, chlorophyll content, and antioxidant enzymes, improving photosynthetic efficiency. Along with the fact that they can be applied as nanopesticides, nano-herbicides, nanofertilizers, and priming agents [163,164], studies have shown that Fe oxide NPs enhance plant physiological processes, including the stimulation of antioxidant enzyme activity and reduction in oxidative stress [165,166,167]. Specifically, Fe3O4 has been shown to promote growth, photosynthesis, respiration, and the content of photosynthetic pigments in leaves, while Fe2O3 induces seed germination, enhances growth, and increases iron accumulation in grains [166,167]. These nanoparticles also boost biomass, chlorophyll content, and ROS production, thereby activating enzymatic defense mechanisms under stress conditions [157,168], and they can replace traditional Fe fertilizers and improve nutrient absorption [169,170,171,172]. Additionally, iron oxide NPs reduce cadmium concentration and promote root and shoot elongation under metal stress by stimulating chlorophyll biosynthesis and redox processes, thereby improving photosynthetic performance [173]. Both Fe2O3 and Fe3O4 have proven effective in enhancing germination rates, plant biomass, and pigmentation, reinforcing their role in mitigating oxidative stress and promoting plant health [173].
Nevertheless, some potential risks associated with the long-term use of metallic and metal oxide nanoparticles in agriculture, particularly concerning soil health, can be observed, for example, NPs can alter the soil physical properties by increasing hydraulic conductivity and soil porosity, and reducing the distance between soil particles [174]. Also, these NPs can affect soil chemical properties, including pH, cation exchange capacity (CEC), electrical conductivity (EC), redox potential (Eh), and soil organic matter (SOM) content [175]. Some research studies about metallic and metallic oxide NPs have shown their toxicity to soil microbes, reducing the population of beneficial bacteria and fungi, which are crucial for nutrient cycling and soil fertility [176]. NPs can accumulate in plant tissues, with varying degrees of translocation depending on the type of NP and plant species. This can lead to phytotoxic effects, including reduced root and shoot biomass, and impaired chlorophyll formation [177,178].
Synthesis Methods of Metallic and Metallic Oxide Nanoparticles
Nanomaterials (NMs) can be synthesized using several methods, which are generally classified into two primary approaches: the top-down approach and the bottom-up approach [139]. The top-down approach involves breaking down bulk material into smaller particles through physical processes such as etching, crushing, ball milling, chemical vapor deposition (CVD), lithography, physical vapor deposition (PVD), and grinding [145,152]. On the other hand, the bottom-up approach involves synthesizing nanoparticles starting from the atomic level and then assembling them into the desired material. Methods within this category include nanoparticle formation from the sol–gel method, chemical reduction, precipitation, hydrothermal synthesis, atomic layer deposition, and green synthesis. A general overview of these methods can be observed in Figure 4 and Figure 5 [139,150,179,180,181,182,183,184].
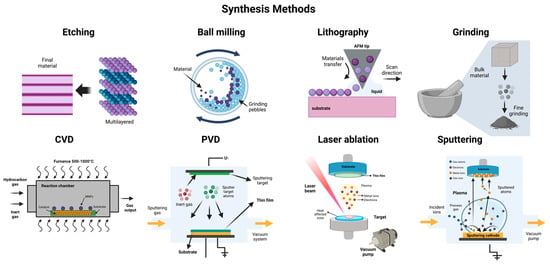
Figure 4.
Different top-down synthesis method for metallic and metallic oxide nanoparticles.
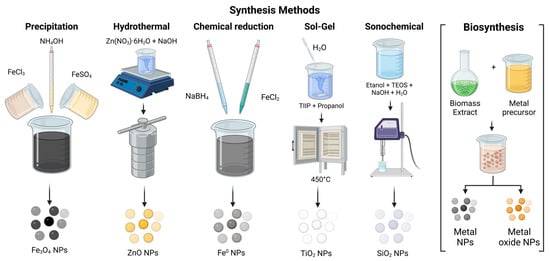
Figure 5.
Different bottom-up synthesis methods for metallic and metallic oxide nanoparticles.
Bottom-up techniques can yield nanomaterials with uniform size, shape, and distribution by precisely controlling the variables of the chemical synthesis process, such as temperature, concentration, stirring, pressure, and the type of reagents used as precursors [145,152] (Figure 5).
As nanotechnology evolves, so do the synthesis routes to obtain nanoparticles. Due to chemical methods using toxic chemicals [142], research has been conducted to synthesizing nanoparticles from a variety of biological entities, such as microbes or plants [143]. Biosynthesis is emerging to develop environmentally friendly and sustainable processes, simplifying operations in chemical production [139,154,185,186,187].
Among the microbes that aid in biosynthesis, various types of bacteria, fungi, algae, and yeasts can be found [188]. On the other hand, plant-mediated biosynthesis depends on the extract obtained, where the diversity of the extracts is greater due to the unique conditions required to apply these chemical compounds and their antioxidant activities [185,186]. The reducing agents derived from these natural substances guide metal reduction and promote stabilization [188]. These agents include terpenes, phenols, flavonoids, amines, amides, proteins, pigments, alkaloids, steroids, etc. [189], and their effectiveness depends on their redox number.
The production of metallic NPs using bioactive extracts involves three phases: (i) the activation phase, where metal ions are reduced and nucleated; (ii) the growth phase, where NPs increase in size and proximity, thereby enhancing thermodynamic stability through the control of parameters such as temperature, and pH being the one responsible of the corona effect; and (iii) the final phase, determined by the composition of the herbal extract [139,142,152].
Although bacterial-mediated synthesis occurs in high abundance, grows rapidly, is easily accessible, and involves unique metal-binding proteins, it has a lower metabolic capacity, and the bioreductive agents produced are from unicellular microorganisms. [53,139,144,188].
Fungal-mediated synthesis is more effective for large-scale nanoparticle production due to its higher metabolic capacity and the greater quantities of bioreductive agents produced by multicellular microorganisms [190]. Plant extracts are increasingly being explored as stabilizing and reducing agents due to their natural abundance and eco-friendly properties [191,192,193,194].
Depending on the synthesis method selected and the selected biomasses, the physicochemical properties, functionality, and stability of the NPs are obtained [142], such as size, dispersion, morphology, and crystalline structure [152,195]. To obtain metallic or metallic oxide nanoparticles by means of biosynthesis, different types of biomasses, such as those listed in Table 2, can be used.
Table 2.
Types of biomasses used as bioreductive agents.
Overall, further studies and toxicological testing are necessary to ensure the safety of these synthesis methods. The green synthesis of nanoparticles through the utilization of plant extracts holds significant potential for applications in both environmental and biomedical fields. This method not only promotes eco-friendly practices, but also aligns with the growing demand for safer and more sustainable technological advancements [56,155,196,200,201,202,203,204,205,206,207].
There are plenty of studies in the literature dealing with different biomass sources as precursors for biosynthesis; nevertheless, plants have shown that it is cost-effective, as it utilizes readily available natural resources and does not require expensive equipment or high-energy inputs [28,208,209]. Different parts of the plants, including leaves, stems, barks, fruits, and flowers, can be used for nanoparticle synthesis, offering a wide range of options for different applications [209]. Some metal-based nanoparticles that have been obtained from plant extracts are presented in Table 3.
Table 3.
Plant extracts as bioreductive agents.
4.3. Mechanisms of Action, Applications and Case Studies
The primary objective of elicitation is to enhance the production of secondary metabolites through the interaction of the elicitor with biochemical pathways, which typically has adverse effects on biomass development, such as changes in morphology [233]. Nonetheless, elicitation can improve certain plant quality traits, including aroma, taste, and color. It is important to note that responses to elicitation are highly dependent on the plant model, the concentration of the elicitor, and the duration of stimulation. Consequently, the effects of elicitation strategies cannot be universally generalized, as studies have demonstrated that the dose–response relationship is not always linear [53]. There are no established guidelines or models for predicting the dose–response relationship in this context. More research to determine the optimal concentrations and exposure times for different NPs and plants should be conducted. Innovative elicitation techniques employing abiotic factors encompass a diverse range of methods, including electromagnetic radiation, acoustic waves, volatile compounds, nutrient deprivation, soil pollutants, and nanomaterials. These novel approaches aim to stimulate secondary metabolite production in plants by leveraging non-biological stressors, thereby enhancing the efficacy and scope of elicitation strategies in various agricultural and biotechnological applications [53,233].
Nanoparticles possess the capability to target specific cellular organelles within plants, enabling the precise release of herbicides, nanopesticides, and fertilizers. Additionally, they can facilitate gene manipulation and expression in targeted plant cells, thereby enhancing plant metabolism. This targeted delivery system improves the efficiency and effectiveness of agricultural treatments and biotechnological interventions, promoting healthier and more productive crops while minimizing environmental impact [234,235].
Nanotechnology and nanomaterials have paved the way for innovative methods to minimize nutrient losses in fertilization using nanofertilizers or nano-encapsulated nutrients [236,237]. These advanced approaches enable enhanced targeting by releasing nutrients on demand as plants grow, ensuring optimal nutrient availability and uptake [155]. This technology not only increases the efficiency of fertilization but also reduces environmental impact by preventing excess nutrient runoff and promoting sustainable agricultural practices [62].
Sometimes the size of structural components in plants enables NP translocation to be efficiently distributed throughout plant organs [59]. Similarly, the size of cellular components is comparable to that of nanoparticles, allowing them to easily permeate cells. However, the potential adverse biological effects of nanoparticles have been widely studied, highlighting the need for careful consideration and regulation in their application [53]. Despite these concerns, the unique properties of nanoparticles offer significant advantages in the targeted delivery and improved efficacy of agricultural interventions [238].
4.3.1. Absorption Pathways of Nanoparticles in Plants
NMs can be absorbed by plant roots primarily through the flow of water and nutrients in the soil. This absorption is significantly influenced by the size, shape, and surface properties of the NMs [239]. The efficiency of uptake and subsequent transport within the plant depends on these characteristics, which determine the interaction of NMs with root cells and their movement through the plant’s vascular system. Understanding these factors is crucial for optimizing the application of nanotechnology in agriculture to enhance nutrient delivery and plant health [34].
In the rhizosphere, the region of soil surrounding the roots, NMs can interact with soil particles and microorganisms. These interactions can significantly influence the mobility and availability of NMs for root absorption; nevertheless, most root studies with NMs have shown that they remain external. The complex interplay between NMs, soil components, and microbial communities can alter the physical and chemical properties of NMs, affecting their uptake efficiency and potential benefits or risks to plant health [34,240]. Understanding these interactions is essential for developing effective and safe applications of nanotechnology in agriculture [57].
NMs penetrate the cell wall and plasma membrane of root epidermal cells through various mechanisms, including endocytosis and passive diffusion. These processes enable NMs to enter the plant cells, where they can interact with intracellular components and pathways. Endocytosis involves the active engulfment of NMs by the cell membrane, forming vesicles that transport them into the cell [241]. Passive diffusion by cell channels, on the other hand, allows NMs to move across the cell membrane based on concentration gradients. Both mechanisms play a crucial role in the internalization and subsequent transport of NMs within plant tissues [53].
NPs can be transported within the plant through the vascular system, utilizing both the xylem and the phloem. In the xylem, nanoparticles move from the roots to the leaves, driven by the process of transpiration [242]. Through the phloem, although less common, nanoparticles can be translocated from the leaves to other parts of the plant [58].
Foliar absorption occurs when nanoparticles are applied directly onto the leaves through spraying. These nanoparticles can penetrate the leaf cuticle and enter the mesophyll cells [243]. Stomata, small openings on the leaf surface, also facilitate nanoparticle entry, granting access to the internal cells. In some cases, nanoparticles can cross the cuticle channels directly without passing through stomata, particularly if they are small and possess properties that enhanced penetration [244].
Intracellularly, nanoparticles can be absorbed by various mechanisms. Endocytosis is a key process wherein cells internalize nanoparticles by forming vesicles from the plasma membrane, involving proteins such as clathrin and caveolin [163]. NPs can also diffuse passively through cell membranes if they are very small and lipophilic [244]. Additionally, some nanoparticles may be actively transported across the cell membrane using specific ions and small-molecule transporters. The efficiency of these processes depends on factors such as the size, surface reactivity, and concentration of the nanoparticles, and the specific characteristics of the plant [245,246].
4.3.2. Nanoparticles to Enhance Metabolite Synthesis
NMs can act as elicitors that enhance the production of secondary metabolites in plants. These metabolites, depending on their chemistry, while not essential for plant growth and development, play crucial roles in defense against abiotic and biotic stress. NMs can induce changes in the biosynthesis of these compounds through various mechanisms [233]. NMs interact directly with plant cells, triggering stress responses that lead to the production of secondary metabolites, such as phenolic compounds, flavonoids, and alkaloids [34,58,199]. These interactions can induce changes in cellular signaling and activate specific metabolic pathways, resulting in the altered expression of genes related to the biosynthesis of secondary metabolites [58]. Specific nanoparticles have differentiated effects:
- Silver nanoparticles (Ag NPs) have demonstrated the ability to increase the production of phenolic compounds and flavonoids in plants such as fenugreek, thereby enhancing their antioxidant capacity and pharmacological properties [170,199]. Research has shown that Ag NPs can interact with plant cells, inducing oxidative stress and activating defense mechanisms [247]. This activation leads to the upregulation of genes involved in the biosynthesis of phenolic compounds and flavonoids, which are key secondary metabolites with significant antioxidant properties [59,248]. Ag NPs enhanced tolerance to saline stress by reducing the salt ion content in plants and improving antioxidant enzyme activity, resulting in increased yield and photosynthetic activity [249].
- Titanium dioxide nanoparticles (TiO2 NPs) can enhance the synthesis of secondary metabolites under stress conditions, such as UV radiation, by increasing the production of antioxidants and other protective compounds [34,250]. When plants are exposed to TiO2 NPs, these nanoparticles can interact with cellular components, leading to the generation of reactive oxygen species (ROS). This controlled oxidative stress can act as a signal that triggers the plant’s defense mechanisms, resulting in the upregulation of pathways involved in the production of secondary metabolites [250]. Under UV radiation, TiO2 NPs can further amplify the plant’s stress response, promoting the synthesis of antioxidants and other protective compounds. These secondary metabolites, such as phenolics and flavonoids, play crucial roles in mitigating oxidative damage and enhancing the plant’s overall resilience to environmental stress [251,252]. In maize crops, hydroponic exposure to TiO2 NPs increased cadmium absorption and phytotoxicity, resulting in reduced chlorophyll content and dry weight in plants. However, the foliar application of TiO2 NPs helped reduce cadmium content in the shoots, thereby mitigating phytotoxicity [253].
- Iron oxide nanoparticles (Fe3O4 NPs) in hydroponic cultures have demonstrated the ability to enhance plant productivity and growth, which is associated with an increase in the synthesis of secondary metabolites. In hydroponic systems, the introduction of Fe3O4 NPs can improve nutrient availability and uptake by plants. These nanoparticles can interact with root systems, enhancing the absorption of essential nutrients and thereby promoting overall plant health and vigor. This improved nutrient status can stimulate metabolic activities within the plant, leading to increased growth rates and higher biomass production [152]. Fe3O4 NPs can induce stress responses that activate the biosynthesis of secondary metabolites. These metabolites, such as phenolic compounds, flavonoids, and other antioxidants, play vital roles in plant defense and adaptation to environmental challenges. The presence of Fe3O4 NPs can enhance the production of these compounds, contributing to the plant’s resilience and overall quality [169,171,254]. The exposure of alfalfa crops to iron nanoparticles (Fe NPs) resulted in increased root length and chlorophyll content due to interactions with hydroxyl radicals that loosened the cell wall [255].
- Cerium oxide nanoparticles (CeO2 NPs) in soybean cultivated under varying soil moisture conditions, both fresh and dry weight increased without significantly affecting the total chlorophyll content. CeO2 NPs may enhance photosynthetic efficiency by improving water and nutrient availability, leading to better growth and biomass production. The impact of nanoparticles on photosynthesis was dependent on soil moisture levels. The findings highlight the importance of considering environmental factors such as soil moisture when assessing the effects of nanoparticles on plant growth and photosynthesis [256].
- The foliar application of zinc oxide nanoparticles (ZnO NPs) in cowpea and okra crops under salinity conditions improved growth parameters compared to plants not treated with nanoparticles. This improvement is attributed to the release of zinc, a vital micronutrient involved in various plant metabolic processes. The improved growth parameters observed in ZnO-treated cowpea and okra plants include increased biomass, better root and shoot development, and higher overall vigor. This indicates that ZnO nanoparticles can mitigate the adverse effects of salinity on plant growth [172,257].
The use of nanomaterials (NMs) as elicitors can significantly enhance the nutritional and pharmacological quality of crops, making agricultural products more valuable and beneficial for human health, as long the NM safety has been proved, and they are not interfering with the target secondary components’ purity [258]. By increasing the production of secondary metabolites, plants can become more resilient to various stress conditions, including drought, salinity, and pathogen attacks. In the presence of biotic stress, such as pathogen attacks, the increased synthesis of secondary metabolites can strengthen the plant’s immune response, providing better protection against diseases [245]. This makes crops more robust and less dependent on chemical pesticides, contributing to more sustainable agricultural practices. Overall, the strategic use of NMs as elicitors offers a promising approach to improve crop quality and resilience. This not only enhances the nutritional and pharmacological value of agricultural products, but also supports sustainable farming practices by reducing the need for chemical inputs and improving plant health and productivity [53,244].
NPs have emerged as significant agents in enhancing the production of bioactive compounds in plants [154]. These compounds, which include flavonoids, phenolic acids, alkaloids, and carotenoids, are valuable for their applications in medicine, pharmacology, cosmetics, agriculture, and the food industry [259,260]. NPs can stimulate specific metabolic pathways, such as shikimate-phenylpropanoid biosynthesis and tryptophan and galactose metabolisms, which are crucial to producing secondary metabolites [233,261]. NPs interact with cellular signaling pathways by inducing oxidative stress, which triggers a cascade of cellular events, including cell cycle arrest, apoptosis by ROS, inflammation, and the induction of antioxidant enzymes [262] (Figure 6). These interactions have significant implications for both beneficial and potentially harmful biological effects.
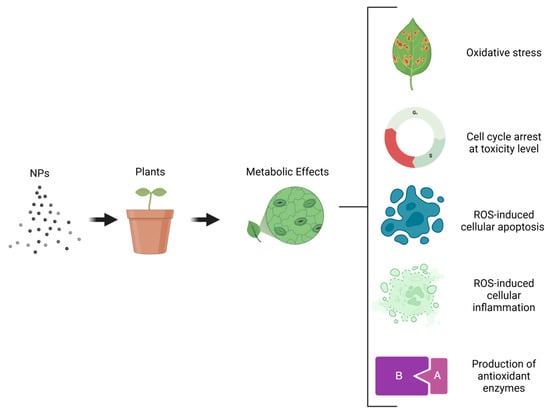
Figure 6.
Metabolic effects induced by NPs in plants cells.
NPs can regulate pathways related to stress response, cell proliferation, and metabolism, leading to the production of bioactive compounds [189]. Certain NPs, such as iron-based (Fe-based) and cerium oxide (CeO2) NPs, have been shown to enhance photosynthesis and promote plant growth, indirectly supporting the production of bioactive compounds by improving overall plant health and metabolic efficiency [263,264]. Also, Exposure to NPs causes shifts in metabolic pathways, such as the transition from sugar metabolism to amino acid metabolism, which can lead to the production of different bioactive compounds [265,266]. Bioactive compound production could be induced by enhancing the plant’s defense mechanisms and overall metabolic activity. This stimulation is critical for improving the plant’s resilience to environmental stressors and promoting healthy growth [267,268]. Metabolic responses to NPs can indeed be tissue-specific, with roots and leaves exhibiting different patterns of metabolic reprogramming. This tissue specificity can significantly influence the type and quantity of bioactive compounds produced in plants [266,269]. The application of NPs in agriculture holds promise for improving plant health and productivity, making it a valuable tool in modern agricultural biotechnology [53,269].
Nanoparticle-induced hormesis in plants highlights how low concentrations of nanoparticles can enhance growth and stimulate the production of crucial secondary metabolites, such as flavonoids and phenolics, while high doses become toxic [267,269,270]. The NOAEL ratio assesses the safe range for nanoparticle application, indicating the threshold where nanoparticles stimulate growth without adverse effects, allowing for controlled use in enhancing plant defense and adaptation [271].
4.3.3. Negative Impact of Nanoparticles on Plant Cells
NPs are increasingly used in various industries due to their unique properties, such as their large specific surface area and high reactivity [139]. However, their widespread use raises concerns about their potential negative impacts on the environment and human health in general. But, specifically, the concern is on plant cells and overall plant health, as their interactions with plant tissues may lead to unintended stress or toxicity [272,273,274]. This bioaccumulation poses risks not only to plant health but also to the food chain, as these NPs can enter the human diet through contaminated crops [169,273,275].
NPs can induce oxidative stress in plants, depending on the location and accumulation, leading to the production of ROS, which can cause membrane damage and ion leakage, ultimately inhibiting plant growth. This oxidative stress disrupts cellular homeostasis, negatively affecting key physiological processes [170,241]. Also, the oxidative stress caused by NPs may damage DNA, proteins, and cellular structures, leading to the activation of cell cycle checkpoints. If DNA damage is detected, cells may enter cell cycle arrest to prevent the propagation of genetic errors, allowing time for repair mechanisms to be activated [246,260,276]. Exposure to metal and metal oxide NPs has the potential to disrupt key metabolic processes in plants, including nucleic acid synthesis, enzyme activity, and the metabolism of carbon and nitrogen [170]. These disruptions can interfere with essential physiological functions, ultimately leading to a reduction in biomass accumulation and negatively impacting overall plant growth and development. The effects of such exposure underscore the need for a thorough understanding of nanoparticle interactions within plant systems to mitigate any adverse outcomes [277,278].
NP exposure in plants has the potential to alter gene expression profiles in plants, affecting critical processes such as plant defense and root development by interfering with pathways like salicylic acid signaling [279]. This exposure can inhibit root hair formation, impairing nutrient and water uptake while also weakening the plant’s defense mechanisms, making it more vulnerable to microbial pathogens. These disruptions highlight the need for careful assessment of nanoparticle effects on plant physiology to avoid compromising crop health and productivity [277,280]. Depending on the dose and place of accumulation in the plant, NPs can interfere with the plant cell wall and vascular system, disrupting the internalization and transport of essential nutrients [244,280]. This interference can result in phytotoxic effects, negatively impacting plant growth, metabolism, and overall health. Such disruptions may alter nutrient uptake and distribution, leading to nutrient deficiencies and impaired physiological functions [241,277].
The negative impacts of NPs on plant cells are multifaceted, encompassing the induction of oxidative stress, the inhibition of root development, the disruption of gene expression, and interference with the plant’s vascular system [244,258]. These effects can result in reduced plant growth, compromised defense mechanisms, and overall phytotoxicity. Understanding these interactions is essential for accurately assessing the environmental risks posed by NPs and for developing effective strategies to mitigate their adverse impacts on plant health and ecosystem stability [145,281].
4.4. Toxicological Concerns, Risk Mitigation, and Guidelines for Safety Applications
Nanotechnology has revolutionized numerous sectors, including agriculture, by leveraging the unique properties of nanomaterials, such as their high surface area, reactivity, and ability to interact with biological systems at the molecular level [272]. However, the increasing use of NPs raises significant concerns about their potential toxicity and environmental impact. As NPs accumulate in soil and water, they may pose risks to plant health, disrupt ecosystems, and affect the food chain, necessitating thorough research into their long-term effects and the development of guidelines to ensure their safe and sustainable application in agriculture and other industries [282].
To mitigate the risks associated with NP use in agriculture, several strategies have been employed. Before the application of NPs, a thorough risk assessment must be conducted to evaluate their potential environmental and health impacts [283]. This assessment should include an in-depth analysis of nanoparticle bioavailability, their chemical and physical transformation in the environment, and their interactions within the soil–plant system, and variability studies [275]. The application of NPs should be carefully controlled in terms of concentration and exposure duration to minimize potential toxic effects. By using lower concentrations and optimizing exposure methods, such as targeted delivery systems, the risk of phytotoxicity can be reduced while still harnessing the beneficial properties of NPs [170]. Establishing comprehensive regulatory guidelines is essential for ensuring the safe use of NPs [282]. These guidelines should include thorough risk assessment protocols, defined permissible exposure limits, and specific safety measures for handling, applying, and disposing of NPs. Additionally, continuous monitoring and evaluation of their environmental and health impacts are crucial to adapt and refine these strategies, ensuring responsible and sustainable implementation of nanotechnology in agriculture [284,285]. Conducting detailed anatomical and physiological studies is essential for understanding the specific effects of NPs on plant systems. This includes performing histological analyses to examine tissue and cell structure alterations, metal bio-uptake studies to track the accumulation and distribution of NPs within plant tissues, and assessments of growth hormones and transcription factors to determine how NPs influence key regulatory pathways in plant development and stress response [285,286].
By implementing effective risk mitigation strategies and adhering to strict regulatory guidelines, the safe use of NPs in plants can be ensured. This approach will help balance the advantages of nanotechnology with the protection of environmental sustainability and food safety, fostering a responsible and sustainable integration of NPs into agriculture [59,272].
Nevertheless, this topic has been discussed in recent decades without any significant results, and this is related to the following: (i) there is no harmonization of the terms “nanotechnology”, “nanomaterial”, “nanoparticle”, and “nanostructured material” and whether some of them have families of “nanoforms”, according to nanomaterial regulations in the EU created by Nanotechnology Industries Association; (ii) nanomaterial toxicity evaluation should be performed according to the application field, such as, food, cosmetics, and health; however, there are no standards to compare against the results obtained. The measurements are performed comparing the bulk materials with the nanomaterials; (iii) the a lack of specific test for NM toxicity, because those used nowadays cannot predict the dispersion, agglomeration, and precipitation of NMs during measurements; and (iv) most underdeveloped countries around the world do not have national policies about nanotechnology and nanoscience, with the reason why being that some of those countries are not willing to provide the economical resources to try to solve some of the issues mentioned before, because in general, these society do not know what nanotechnology and nanoscience are.
Finally, in 2020, to make REACH (registration, evaluation, authorization, and restriction of chemicals) enforceable for nanomaterials, in a first step, the Malta Initiative focused on projects in which OECD (Organization for Economic Co-operation and Development) Test Guidelines (TGs) were amended or developed for nanoscale substances. Other legislative areas, such as European regulations on biocides, cosmetics, and plastic food packaging, also refer to some extent to the OECD TGs when it comes to safety assessments for nanomaterials, and thus also benefit from the amendments. This initiative is expected to be released in the first stage in the second half of 2025 (https://malta-initiative.org, accessed on 7 November 2024).
5. Integration of Elicitation, Metabolic Engineering, and Biosynthesized Nanoparticles to Bioactive Compound Production in Plants
The integration of elicitation, metabolic engineering, and biosynthesized nanoparticles represents a cutting-edge approach to enhancing the production of bioactive compounds in plants. Each of these techniques contributes uniquely to this goal, and their combination can lead to synergistic effects, resulting in significantly improved yields of desired phytochemicals [287]. The unique properties of nanoparticles to enhance the production of valuable metabolites, optimize metabolic pathways, and develop innovative therapeutic strategies represent a promising frontier in biotechnology [288].
As has been presented, elicitation involves the use of biotic or abiotic factors to stimulate the plant’s defense mechanisms, leading to the increased production of secondary metabolites. These metabolites often include valuable bioactive compounds such as alkaloids, flavonoids, and terpenoids, which are crucial for pharmaceuticals, nutraceuticals, and other industries [287]. Metabolic engineering involves the genetic modification of plants to enhance or redirect their metabolic pathways. This can be done to either overproduce existing bioactive compounds or to produce novel compounds that the plant does not naturally synthesize, or by overexpressing key enzymes to enhance the activity of specific enzymes involved in the biosynthetic pathway of a target compound, or even by genetic modification [288,289].
Nanoparticles synthesized via biological methods offer a biocompatible and environmentally friendly approach [290] to enhancing plant growth and secondary metabolite production. These nanoparticles can influence plant metabolism in several ways: (i) by nanoparticle uptake, the NPs can act as elicitors themselves, triggering defense responses similar to traditional biotic or abiotic elicitors; (ii) in nanoparticle-mediated delivery, the NPs can be used to deliver elicitors, genes, or other molecules directly into plant cells, enhancing the efficiency and effectiveness of metabolic engineering and elicitation strategies, for example, SiO2; and (iii) by catalytic activity, where some metallic nanoparticles can catalyze reactions within the plant cells, potentially enhancing the biosynthesis of targeted metabolites [291].
By integrating elicitation, metabolic engineering, and biosynthesized nanoparticles, researchers can develop highly efficient systems for boosting the production of bioactive compounds in plants. For example, NPs can be engineered to carry specific elicitors, which are then delivered into plant tissues to induce the production of desired compounds. Some have proposed using NPs as gene delivery vehicles, after they have been modified through metabolic engineering directly into plant cells, improving the efficiency of genetic modification. Also, combining elicitation and NPs simultaneously triggers plant defense mechanisms and enhances the uptake of nutrients or other growth-promoting factors [292,293,294,295]. The integration of these methods is particularly promising for agriculture and biotechnology industries aiming to produce high-value compounds more efficiently and sustainably. It offers the potential to (i) increase the yield of pharmaceuticals and nutraceuticals, (ii) reduce the need for chemical inputs in agriculture, (iii) enhance the stress resistance and overall health of plants, and (iv)contribute to sustainable agricultural practices using green synthesized nanomaterials [296].
The literature concerning the integration of the elicitation strategy with metabolic engineering and biosynthesized NPs as a strategy for bioactive compound production is limited, and several research gaps require further investigation. For example, elicitation and gene overexpression, transcription factor regulation, plant polyploidization, precursor feeding, and plant growth methods, such as factory farming, have been studied as effective methods to improve the synthesis of secondary metabolites in medicinal plants; nevertheless, there is no evidence in these studies of the integration of NPs for the same purpose, but only as a single strategy [297].
Other studies integrated transcriptomics, proteomics, and metabolomics with systems biology, and could identify novel genes and biosynthetic pathways. This integration aids in metabolic engineering by providing targets for gene manipulation to enhance secondary metabolite production [298]. Enhancing the catalytic properties of rate-limiting enzymes through site-directed mutagenesis and fusing them with other enzymes can significantly improve productivity. This approach, combined with metabolic engineering, and CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) modifications [299], can synergistically boost the production of target compounds [300]. On the other hand, as has been presented, nanoparticles can mimic natural elicitors like phytohormones and microbial extracts, enhancing specialized metabolic pathways and gene regulation in plant cell cultures. This approach can be tailored by designing NPs with specific physicochemical properties to target desired metabolites [287].
Integrating elicitation, metabolic engineering, and biosynthesized NPs involves overcoming challenges related to cytotoxicity, metabolic complexity, targeted delivery, bioactivity maintenance, scalability, and regulatory concerns [290]. For example, (i) NPs can exhibit cytotoxic effects, which pose a significant challenge for their safe application in biological systems. Ensuring that NPs are biocompatible while maintaining their functional properties is crucial [287,290]; (ii) the complexity and dynamic nature of metabolic pathways in cells make it difficult to achieve a consistent and high-yield production of desired metabolites, because it depends on the dose and bioaccumulation. Dynamic control systems and the fine-tuning of metabolic pathways are required to address these issues [288,301]; (iii) transitioning from exploratory research to commercially viable strategies requires scalable and cost-effective production methods. This includes the development of robust and reproducible processes for NP synthesis and functionalization [287]. Addressing these challenges through innovative approaches and thorough research is essential for advancing the field and achieving practical applications.
6. Future Directions and Applications
NPs have the potential to enhance the production of plants’ specialized metabolites. Also, the application of elicitation and metabolic engineering has shown outstanding results in this area. Combining NPs along with elicitation and metabolic engineering seems to be a powerful tool for generating a new sustainable agricultural practice. However, further research is required to identify suitable nanomaterials and to understand their uptake, translocation, internalization, and elicitation mechanisms comprehensively [54]. Extensive studies are needed to determine which types of NPs (e.g., metal oxides, carbon-based, or organic NPs) are most effective in enhancing the production of specific secondary metabolites [59,163,233]. The size, shape, and surface properties (such as charge) of NPs play crucial roles in their interaction with plant cells, and these factors must be optimized for different plant species and desired outcomes [139,302].
While abiotic elicitors can be effective in enhancing plant resistance and secondary metabolite production, their use is associated with several disadvantages. These include the inhibition of photosynthesis, complex and unpredictable metabolic responses, increased susceptibility to other stressors, negative effects on plant biomass, and genotype-dependent variability [84,303,304]. Regarding metabolic engineering in plants, it can lead to significant negative effects, including reduced concentrations of important phenolic compounds, hidden metabolic constraints, and complex responses to combined abiotic stresses. These challenges highlight the need for a comprehensive understanding of plant metabolism and the development of more sophisticated engineering strategies to mitigate adverse impacts [128,305,306].
In general, depending on the application of the NPs as elicitors or the use of other abiotic/biotic elicitors, and some methods revied form the metabolic engineering, research should also delve into changes in mechanisms at the cellular level, including endocytosis and passive diffusion, to determine how NPs interact with intracellular components, for instance [245]. Understanding the mechanisms by which NPs elicit the production of specialized metabolites is critical, due to plants being stationary and having evolved mechanisms by which they can protect themselves. This includes studying how NPs induce stress responses, activate specific signaling pathways, and influence gene expression related to secondary metabolite biosynthesis [103,260,307], because in the end, the tools mentioned in this work are related to and focused on the purpose of increasing bioactive compound production in plants.
NPs in agriculture have the potential to combat nutrient deficiencies, promote stress tolerance, and improve crop yield and quality. However, their use may also pose adverse effects on the environment, health, and the food chain [308]. The release of NPs into the environment can affect soil health and microbial communities [309]. Their persistence and bioaccumulation can lead to long-term ecological consequences [154,170]. Exposure to NPs through the food chain or direct contact can pose health risks. The toxicity of NPs depends on their composition, size, and concentration, and their long-term effects are still not fully understood [60]. NPs can enter the food chain, potentially affecting the health of consumers and wildlife [246]. The bioavailability and toxicity of NPs in edible plant parts needs careful assessment to ensure food safety [310,311].
Utilizing plants for nanoparticle synthesis leverages inexpensive and readily available plant materials. This approach reduces the overall production costs compared to traditional chemical and physical methods. However, concerns remain about the potential health impacts caused by nanotoxicity upon their release, as recently mentioned [273]. Plants provide a green synthesis route for nanoparticles, utilizing natural reducing and stabilizing agents found in plant extracts [196]. Nanoparticles synthesized using plants are often biocompatible and biodegradable, minimizing environmental pollution and reducing the risk of long-term ecological damage [281]. This process eliminates the need for the hazardous chemicals and high-energy consumption typically associated with conventional nanoparticle synthesis methods [200,312].
Nanoparticles can be engineered to act as fertilizers, pesticides, and herbicides directly (as vectors) in the plant tissues or root zones, improving the precision and effectiveness of these inputs [141,313]. Also, NPs can provide a controlled release of nutrients and agrochemicals, ensuring a steady supply over time and reducing the frequency of applications needed [150]. In agriculture, these nanoparticles can enhance crop growth, protect against pests and diseases, and improve nutrient uptake without the harmful environmental impact of conventional agrochemicals [58]. Biocompatible nanoparticles synthesized using plant extracts can be used for drug delivery, imaging, and diagnostics, offering a safer alternative to chemically synthesized nanoparticles [314,315]. The use of safe and stable nanoparticles in industrial processes can lead to the development of more sustainable and less polluting technologies [281].
To fully harness the potential of plant-mediated synthesis, the standardization of protocols and characterization techniques is necessary to ensure consistency and reproducibility. Research is needed to address challenges related to scaling up the synthesis process for industrial production while maintaining safety and efficacy [145]. Comprehensive toxicological studies are essential to evaluate the long-term safety of plant-synthesized nanoparticles, particularly for biomedical and food applications.
The use of nanoparticles in agriculture presents significant opportunities for enhancing crop production, reducing the use for harmful agrochemicals, and promoting sustainable farming practices [316]. Nevertheless, the adoption of nanotechnology in agriculture also brings forth several regulatory challenges that need to be addressed to ensure environmental safety (water and soil quality), human health, and cost–benefit effectiveness [58,317]. For instance, there are concerns about the environmental behavior of nanopesticides and their potential harmful effects on non-target organisms and beneficial insects [318]. As the open field application of nanopesticides can cause ecological safety issues, there is an urgent need for stringent regulatory frameworks to ensure their safe use [319], but the information available is in the early stages of development, even though the birth of nanotechnology as a novel science occurred more than a few decades ago. In agriculture, there is a critical need for elaborate investigations into the mechanistic applications and agroecological toxicity of engineered nanomaterials to develop safe and effective regulatory frameworks [272].
Addressing the regulatory challenges associated with the use of nanoparticles in agriculture, there are more questions than answers, (i) there is a need for comprehensive studies on the fate, mobility, and toxicity of NPs in soil and their interactions with soil constituents [61], (ii) we need to develop controlled release and stimuli-responsive systems for NPs to minimize environmental exposure and enhance the efficiency of agrochemicals [317], (iii) and more studies about precision in the delivery of NPs are needed to understand how it can be performed to reduce the required dosage and mitigate potential negative impacts on non-target organisms [319]. (iv) Establishing a robust regulatory framework that includes standardized testing protocols for the synthesis, application, and environmental impact of NPs is crucial [60], (v) regulatory associations need to develop guidelines for the safe use of NPs, considering their unique properties and potential risks [319], (vi) increasing public awareness and understanding of the benefits and risks associated with NPs in agriculture can help in gaining public acceptance and support for regulatory measures [318], and (vii) transparent communication about the safety and efficacy of NP-based products is essential to address public concerns and promote informed decision-making [320].
There is a significant gap in specific regulatory frameworks for formulations containing nanomaterials, which complicates the approval process due to the unique properties of NPs that require additional safety, quality, and efficacy testing. The absence of a clear international definition and consistent regulatory guidelines for NMs creates uncertainty and hinders the development and acceptance of nanoproducts [321,322]. The scientific community and governmental figures have called for the harmonization and approximation of international regulatory practices to ensure a consistent assessment of the risk–benefit balance of nanomaterials, considering both human and environmental health. The development of standardized protocols and universal methodologies for characterizing nanomaterials is essential to overcome conflicting safety studies and to establish reliable safety regulations [323,324,325,326]. The complexity and dynamic behavior of advanced nanomaterials present new challenges for safety and sustainability assessments, which existing regulatory frameworks may not fully address [323]. Approaches such as Safe Innovation and Safe- and Sustainable-by-Design, along with standardized protocols and the TRAAC framework (Transparency, Reliability, Accessibility, Applicability and Completeness), are essential steps towards achieving comprehensive and reliable regulatory oversight for nanomaterials; but, still, more work and more involvement from people is needed [325,326].
7. Conclusions
The integration of elicitation, metabolic engineering, and biosynthesized nanoparticles represents a promising frontier in enhancing bioactive compound production in plants. This approach leverages the unique properties of nanoparticles to augment the efficiency of elicitation and metabolic engineering strategies, offering a sustainable alternative to traditional methods. The use of biosynthesized metallic and metallic oxide nanoparticles, derived through environmentally friendly methods, adds an additional layer of eco-compatibility to these processes, which is crucial, given the growing demand for green technology in agriculture and biotechnology.
However, the application of these advanced tools presents challenges. The variability in plant responses to different elicitors and nanoparticles, coupled with the complexity of metabolic pathways, makes it difficult to establish a universal approach that is effective across all plant species and conditions. Furthermore, while nanoparticles offer significant advantages in terms of precision and efficiency, their potential environmental and health impacts require careful consideration. The interactions between nanoparticles and plant systems are still not fully understood, particularly regarding their long-term effects on plant physiology and the surrounding ecosystem.
Future research should focus on unraveling the mechanisms underlying nanoparticle uptake, translocation, and interaction within plant cells. Additionally, there is a need to develop standardized protocols for nanoparticle application in agriculture, ensuring that their use is both safe and effective. As these technologies continue to evolve, they hold the potential for revolutionizing the production of bioactive compounds in plants, contributing to sustainable agricultural practices and the development of high-value products for the pharmaceutical, nutraceutical, and food industries. However, this progress must be balanced with a rigorous assessment of the risks and benefits, ensuring that the adoption of these technologies does not compromise environmental and human health.
This review emphasizes the need for further research into the mechanisms of nanoparticle uptake, translocation, and their long-term effects on both plant and environmental health, about the diverse mechanisms that can be modified in plants by elicitation and metabolic engineering for their integration for sustainable agriculture and enhanced bioactive compound production, which seems to hold significant promise.
Author Contributions
Conceptualization, L.A.M.-C., A.A.F.-P., and K.E.E.; investigation, L.A.M.-C., M.Y.H.-R., A.A.F.-P., and K.E.E.; resources, A.A.F.-P. and K.E.E.; writing—original draft preparation, L.A.M.-C., M.Y.H.-R., A.A.F.-P., and K.E.E.; writing—review and editing, K.E.E.; visualization, A.A.F.-P. and K.E.E.; supervision, A.A.F.-P. and K.E.E.; funding acquisition, L.A.M.-C., A.A.F.-P., and K.E.E. All authors have read and agreed to the published version of the manuscript.
Funding
FIN202413; FIN202417; FIN202420 (FONFIVE-2024 Universidad Autónoma de Querétaro).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Mariana Y. Hernández-Ramírez thanks FONFIVE-2024 for the scholarship granted.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Barone, R.P.; Knittel, D.K.; Ooka, J.K.; Porter, L.N.; Smith, N.T.; Owens, D.K. The production of plant natural products beneficial to humanity by metabolic engineering. Curr. Plant Biol. 2020, 24, 100121. [Google Scholar] [CrossRef]
- Awere, C.O.; Rakkammal, K.; Mwaura, M.M.; Anadebe, V.C.; Ramesh, M. Hairy-root technology: A metabolic engineering tool and specialized metabolite pathway elucidation and production of secondary metabolites. A review. Results Eng. 2024, 23, 102697. [Google Scholar] [CrossRef]
- Cardillo, A.B.; Perassolo, M.; Giulietti, A.M.; Rodriguez Talou, J. Cyclodextrins: A tool in plant cell and organ culture bioprocesses for the production of secondary metabolites. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 1–19. [Google Scholar] [CrossRef]
- Livadariu, O.; Maximilian, C.; Rahmanifar, B.; Cornea, C.P. LED Technology Applied to Plant Development for Promoting the Accumulation of Bioactive Compounds: A Review. Plants 2023, 12, 1075. [Google Scholar] [CrossRef]
- de Bruijn, W.J.C.; Levisson, M.; Beekwilder, J.; van Berkel, W.J.H.; Vincken, J.P. Plant Aromatic Prenyltransferases: Tools for Microbial Cell Factories. Trends Biotechnol. 2020, 38, 917–934. [Google Scholar] [CrossRef]
- Krakowska-Sieprawska, A.; Kiełbasa, A.; Rafińska, K.; Ligor, M.; Buszewski, B. Modern Methods of Pre-Treatment of Plant Material for the Extraction of Bioactive Compounds. Molecules 2022, 27, 730. [Google Scholar] [CrossRef]
- De Carlo, A.; Tarraf, W.; Lambardi, M.; Benelli, C. Temporary Immersion System for Production of Biomass and Bioactive Compounds from Medicinal Plants. Agronomy 2021, 11, 2414. [Google Scholar] [CrossRef]
- Motolinía-Alcántara, E.A.; Castillo-Araiza, C.O.; Rodríguez-Monroy, M.; Román-Guerrero, A.; Cruz-Sosa, F. Engineering Considerations to Produce Bioactive Compounds from Plant Cell Suspension Culture in Bioreactors. Plants 2021, 10, 2762. [Google Scholar] [CrossRef]
- Yeshi, K.; Crayn, D.; Ritmejerytė, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27, 313. [Google Scholar] [CrossRef] [PubMed]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental Factors Regulate Plant Secondary Metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef] [PubMed]
- Rosegrant, M.W.; Tokgoz, S.; Bhandary, P. The New Normal? A Tighter Global Agricultural Supply and Demand Relation and Its Implications for Food Security. Am. J. Agric. Econ. 2013, 95, 303–309. [Google Scholar] [CrossRef]
- Sheikh Mohammad Fakhrul, I.; Zahurul, K. World’s Demand for Food and Water: The Consequences of Climate Change. In Desalination; Mohammad Hossein Davood Abadi, F., Vahid, V., Amir Hooshang, T., Eds.; IntechOpen: Rijeka, Croatia, 2019; Chapter 4. [Google Scholar]
- Tian, X.; Engel, B.A.; Qian, H.; Hua, E.; Sun, S.; Wang, Y. Will reaching the maximum achievable yield potential meet future global food demand? J. Clean. Prod. 2021, 294, 126285. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Francini, A.; Sebastiani, L. Abiotic Stress Effects on Performance of Horticultural Crops. Horticulturae 2019, 5, 67. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef]
- Alcalde, M.A.; Perez-Matas, E.; Escrich, A.; Cusido, R.M.; Palazon, J.; Bonfill, M. Biotic Elicitors in Adventitious and Hairy Root Cultures: A Review from 2010 to 2022. Molecules 2022, 27, 5253. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Oh, D.-H. Elicitation: A new perspective into plant chemo-diversity and functional property. Crit. Rev. Food Sci. Nutr. 2023, 63, 4522–4540. [Google Scholar] [CrossRef]
- Huang, L.; Ho, C.-T.; Wang, Y. Biosynthetic pathways and metabolic engineering of spice flavors. Crit. Rev. Food Sci. Nutr. 2021, 61, 2047–2060. [Google Scholar] [CrossRef]
- Ullah, N.; Shahzad, K.; Wang, M. The Role of Metabolic Engineering Technologies for the Production of Fatty Acids in Yeast. Biology 2021, 10, 632. [Google Scholar] [CrossRef]
- Cai, L.; Cai, L.; Jia, H.; Liu, C.; Wang, D.; Sun, X. Foliar exposure of Fe3O4 nanoparticles on Nicotiana benthamiana: Evidence for nanoparticles uptake, plant growth promoter and defense response elicitor against plant virus. J. Hazard. Mater. 2020, 393, 122415. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Hernández, M.C.; Parola-Contreras, I.; Montoya-Gómez, L.M.; Torres-Pacheco, I.; Schwarz, D.; Guevara-González, R.G. Eustressors: Chemical and physical stress factors used to enhance vegetables production. Sci. Hortic. 2019, 250, 223–229. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.d.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.d.L.; Torres-Pacheco, I. Plant Hormesis Management with Biostimulants of Biotic Origin in Agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef]
- Vega-Vásquez, P.; Mosier, N.S.; Irudayaraj, J. Nanoscale Drug Delivery Systems: From Medicine to Agriculture. Front. Bioeng. Biotechnol. 2020, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Avellan, A.; Yun, J.; Morais, B.P.; Clement, E.T.; Rodrigues, S.M.; Lowry, G.V. Critical Review: Role of Inorganic Nanoparticle Properties on Their Foliar Uptake and in Planta Translocation. Environ. Sci. Technol. 2021, 55, 13417–13431. [Google Scholar] [CrossRef] [PubMed]
- Alabdallah, N.M.; Hasan, M.M.; Hammami, I.; Alghamdi, A.I.; Alshehri, D.; Alatawi, H.A. Green Synthesized Metal Oxide Nanoparticles Mediate Growth Regulation and Physiology of Crop Plants under Drought Stress. Plants 2021, 10, 1730. [Google Scholar] [CrossRef]
- Soni, V.; Raizada, P.; Singh, P.; Cuong, H.N.; Rangabhashiyam, S.; Saini, A.; Saini, R.V.; Le, Q.V.; Nadda, A.K.; Le, T.-T.; et al. Sustainable and green trends in using plant extracts for the synthesis of biogenic metal nanoparticles toward environmental and pharmaceutical advances: A review. Environ. Res. 2021, 202, 111622. [Google Scholar] [CrossRef]
- Cuong, H.N.; Pansambal, S.; Ghotekar, S.; Oza, R.; Thanh Hai, N.T.; Viet, N.M.; Nguyen, V.-H. New frontiers in the plant extract mediated biosynthesis of copper oxide (CuO) nanoparticles and their potential applications: A review. Environ. Res. 2022, 203, 111858. [Google Scholar] [CrossRef]
- Saravanan, A.; Kumar, P.S.; Karishma, S.; Vo, D.-V.N.; Jeevanantham, S.; Yaashikaa, P.R.; George, C.S. A review on biosynthesis of metal nanoparticles and its environmental applications. Chemosphere 2021, 264, 128580. [Google Scholar] [CrossRef]
- Zain, M.; Ma, H.; Ur Rahman, S.; Nuruzzaman, M.; Chaudhary, S.; Azeem, I.; Mehmood, F.; Duan, A.; Sun, C. Nanotechnology in precision agriculture: Advancing towards sustainable crop production. Plant Physiol. Biochem. 2024, 206, 108244. [Google Scholar] [CrossRef]
- Alvarado, A.M.; Aguirre-Becerra, H.; Vázquez-Hernández, M.C.; Magaña-Lopez, E.; Parola-Contreras, I.; Caicedo-Lopez, L.H.; Contreras-Medina, L.M.; Garcia-Trejo, J.F.; Guevara-Gonzalez, R.G.; Feregrino-Perez, A.A. Influence of Elicitors and Eustressors on the Production of Plant Secondary Metabolites. In Natural Bio-Active Compounds: Volume 1: Production and Applications; Akhtar, M.S., Swamy, M.K., Sinniah, U.R., Eds.; Springer: Singapore, 2019; pp. 333–388. [Google Scholar]
- Godínez-Mendoza, P.L.; Hurtado-Zuñiga, A.; Siboney-Montante, V.; Guzman-Cruz, R.; Guevara-González, R.G. Eustressors to Improve Plant Secondary Metabolites Production: Insect Frass and Physical Factors as Examples Applied in Agriculture and Horticulture. In Molecular Dynamics of Plant Stress and Its Management; Shahid, M., Gaur, R., Eds.; Springer Nature: Singapore, 2024; pp. 25–49. [Google Scholar]
- Páramo, L.; Feregrino-Pérez, A.A.; Vega-González, M.; Escobar-Alarcón, L.; Esquivel, K. Medicago sativa L. Plant Response against Possible Eustressors (Fe, Ag, Cu)-TiO2: Evaluation of Physiological Parameters, Total Phenol Content, and Flavonoid Quantification. Plants 2023, 12, 659. [Google Scholar] [CrossRef] [PubMed]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2015, 5, 804. [Google Scholar] [CrossRef] [PubMed]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Long, T.A. Many needles in a haystack: Cell-type specific abiotic stress responses. Curr. Opin. Plant Biol. 2011, 14, 325–331. [Google Scholar] [CrossRef]
- Berrios, L.; Rentsch, J.D. Linking Reactive Oxygen Species (ROS) to Abiotic and Biotic Feedbacks in Plant Microbiomes: The Dose Makes the Poison. Int. J. Mol. Sci. 2022, 23, 4402. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, Y.Z.; Abdel-Rahman, S.S.A.; Soliman, W.S.; Salaheldin, S. Growth, yield, and quality of roselle (Hibiscus sabdariffa L.) plants as affected by nano zinc and bio-stimulant treatments. Hortic. Environ. Biotechnol. 2021, 62, 879–890. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an Effective Strategy for the Biotechnological Production of Bioactive High-Added Value Compounds in Plant Cell Factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Duarte-Sierra, A.; Munzoor Hasan, S.M.; Angers, P.; Arul, J. UV-B radiation hormesis in broccoli florets: Glucosinolates and hydroxy-cinnamates are enhanced by UV-B in florets during storage. Postharvest Biol. Technol. 2020, 168, 111278. [Google Scholar] [CrossRef]
- Neugart, S.; Bumke-Vogt, C. Flavonoid Glycosides in Brassica Species Respond to UV-B Depending on Exposure Time and Adaptation Time. Molecules 2021, 26, 494. [Google Scholar] [CrossRef]
- Sidibé, A.; Charles, M.T.; Lucier, J.-F.; Xu, Y.; Beaulieu, C. Preharvest UV-C Hormesis Induces Key Genes Associated With Homeostasis, Growth and Defense in Lettuce Inoculated with Xanthomonas campestris pv. vitians. Front. Plant Sci. 2022, 12, 793989. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.T.D.; Puthur, J.T. UV-B priming enhances specific secondary metabolites in Oryza sativa (L.) empowering to encounter diverse abiotic stresses. Plant Growth Regul. 2020, 92, 169–180. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Sakalauskienė, S.; Samuolienė, G.; Jankauskienė, J.; Viršilė, A.; Novičkovas, A.; Sirtautas, R.; Miliauskienė, J.; Vaštakaitė, V.; Dabašinskas, L.; et al. The effects of LED illumination spectra and intensity on carotenoid content in Brassicaceae microgreens. Food Chem. 2015, 173, 600–606. [Google Scholar] [CrossRef]
- Loi, M.; Villani, A.; Paciolla, F.; Mulè, G.; Paciolla, C. Challenges and Opportunities of Light-Emitting Diode (LED) as Key to Modulate Antioxidant Compounds in Plants. A Review. Antioxidants 2021, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef]
- Sena, S.; Kumari, S.; Kumar, V.; Husen, A. Light emitting diode (LED) lights for the improvement of plant performance and production: A comprehensive review. Curr. Res. Biotechnol. 2024, 7, 100184. [Google Scholar] [CrossRef]
- Liu, Z.; Tian, L.; Chen, M.; Zhang, L.; Lu, Q.; Wei, J.; Duan, X. Hormesis Responses of Growth and Photosynthetic Characteristics in Lonicera japonica Thunb. to Cadmium Stress: Whether Electric Field Can Improve or Not? Plants 2023, 12, 933. [Google Scholar] [CrossRef]
- Bhandawat, A.; Jayaswall, K.; Sharma, H.; Roy, J. Sound as a stimulus in associative learning for heat stress in Arabidopsis. Commun. Integr. Biol. 2020, 13, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Caicedo-Lopez, L.H.; Guevara-Gonzalez, R.G.; Andrade, J.E.; Esquivel-Delgado, A.; Perez-Matzumoto, A.E.; Torres-Pacheco, I.; Contreras-Medina, L.M. Effect of hydric stress-related acoustic emission on transcriptional and biochemical changes associated with a water deficit in Capsicum annuum L. Plant Physiol. Biochem. 2021, 165, 251–264. [Google Scholar] [CrossRef]
- Aguirre-Becerra, H.; Feregrino-Pérez, A.A.; Esquivel, K.; Perez-Garcia, C.E.; Vazquez-Hernandez, M.C.; Mariana-Alvarado, A. Nanomaterials as an alternative to increase plant resistance to abiotic stresses. Front. Plant Sci. 2022, 13, 1023636. [Google Scholar] [CrossRef]
- Anjum, S.; Anjum, I.; Hano, C.; Kousar, S. Advances in nanomaterials as novel elicitors of pharmacologically active plant specialized metabolites: Current status and future outlooks. RSC Adv. 2019, 9, 40404–40423. [Google Scholar] [CrossRef] [PubMed]
- Humbal, A.; Pathak, B. Influence of exogenous elicitors on the production of secondary metabolite in plants: A review (“VSI: Secondary metabolites”). Plant Stress. 2023, 8, 100166. [Google Scholar] [CrossRef]
- Kaningini, A.G.; Nelwamondo, A.M.; Azizi, S.; Maaza, M.; Mohale, K.C. Metal Nanoparticles in Agriculture: A Review of Possible Use. Coatings 2022, 12, 1586. [Google Scholar] [CrossRef]
- Kirova, E.; Geneva, M.; Petrova, M.; Miladinova-Georgieva, K.; Sichanova, M. Employment of nanoparticles for improvement of plant growth and development. Botanica 2022, 28, 113–132. [Google Scholar] [CrossRef]
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in Agroindustry: Applications, Toxicity, Challenges, and Trends. Nanomaterials 2020, 10, 1654. [Google Scholar] [CrossRef]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in sustainable agriculture: Recent developments, challenges, and perspectives. Front. Microbiol. 2017, 8, 1014. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control. Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.u.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-Biotechnology in Agriculture: Use of Nanomaterials to Promote Plant Growth and Stress Tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. Int. J. Mol. Sci. 2023, 24, 3266. [Google Scholar] [CrossRef]
- Dasari, R.; Gopu, C.; Vankudoth, S.; Dharavath, S.; Taduri, S. Enhancement of production of pharmaceutically important anti-cancerous compound; cucurbitacin E via elicitation and precursor feeding of in vitro culture of Citrullus colocynthis (L.) Schard. Vegetos 2020, 33, 323–334. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Singh, M.; Srivastava, M.; Kumar, A.; Pandey, K.D. 12—Biosynthesis of nanoparticles and applications in agriculture. In Role of Plant Growth Promoting Microorganisms in Sustainable Agriculture and Nanotechnology; Kumar, A., Singh, A.K., Choudhary, K.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 199–217. [Google Scholar]
- Uddin, M.; Bhat, U.H.; Singh, S.; Singh, S.; Chishti, A.S.; Khan, M.M.A. Combined application of SiO2 and TiO2 nanoparticles enhances growth characters, physiological attributes and essential oil production of Coleus aromatics Benth. Heliyon 2023, 9, e21646. [Google Scholar] [CrossRef]
- Castro-González, C.G.; Sánchez-Segura, L.; Gómez-Merino, F.C.; Bello-Bello, J.J. Exposure of stevia (Stevia rebaudiana B.) to silver nanoparticles in vitro: Transport and accumulation. Sci. Rep. 2019, 9, 10372. [Google Scholar] [CrossRef]
- Nokandeh, S.; Ramezani, M.; Gerami, M. The physiological and biochemical responses to engineered green graphene/metal nanocomposites in Stevia rebaudiana. J. Plant Biochem. Biotechnol. 2021, 30, 579–585. [Google Scholar] [CrossRef]
- Magaña-López, E.; Palos-Barba, V.; Zuverza-Mena, N.; Vázquez-Hernández, M.C.; White, J.C.; Nava-Mendoza, R.; Feregrino-Pérez, A.A.; Torres-Pacheco, I.; Guevara-González, R.G. Nanostructured mesoporous silica materials induce hormesis on chili pepper (Capsicum annuum L.) under greenhouse conditions. Heliyon 2022, 8, e09049. [Google Scholar] [CrossRef]
- Guzmán-Báez, G.A.; Trejo-Téllez, L.I.; Ramírez-Olvera, S.M.; Salinas-Ruíz, J.; Bello-Bello, J.J.; Alcántar-González, G.; Hidalgo-Contreras, J.V.; Gómez-Merino, F.C. Silver Nanoparticles Increase Nitrogen, Phosphorus, and Potassium Concentrations in Leaves and Stimulate Root Length and Number of Roots in Tomato Seedlings in a Hormetic Manner. Dose-Response 2021, 19, 15593258211044576. [Google Scholar] [CrossRef]
- Chahardoli, A.; Sharifan, H.; Karimi, N.; Kakavand, S.N. Uptake, translocation, phytotoxicity, and hormetic effects of titanium dioxide nanoparticles (TiO2NPs) in Nigella arvensis L. Sci. Total Environ. 2022, 806, 151222. [Google Scholar] [CrossRef]
- Yadegari, M. Study of phytohormones effects on UV-B stress seeds of thyme species. J. Herb. Drugs 2017, 8, 109–115. [Google Scholar] [CrossRef]
- Volkova, P.Y.; Clement, G.; Makarenko, E.S.; Kazakova, E.A.; Bitarishvili, S.V.; Lychenkova, M.A. Metabolic Profiling of γ-Irradiated Barley Plants Identifies Reallocation of Nitrogen Metabolism and Metabolic Stress Response. Dose-Response 2020, 18, 1559325820914186. [Google Scholar] [CrossRef]
- Orlando, M.; Trivellini, A.; Puccinelli, M.; Ferrante, A.; Incrocci, L.; Mensuali-Sodi, A. Increasing the functional quality of Crocus sativus L. by-product (tepals) by controlling spectral composition. Hortic. Environ. Biotechnol. 2022, 63, 363–373. [Google Scholar] [CrossRef]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Zare Mehrjerdi, M.; Dianati Daylami, S.; Serek, M.; Woltering, E.; Li, T. Blue Light Improves Vase Life of Carnation Cut Flowers Through Its Effect on the Antioxidant Defense System. Front. Plant Sci. 2020, 11, 511. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Dobránszki, J. Magnetic fields: How is plant growth and development impacted? Protoplasma 2016, 253, 231–248. [Google Scholar] [CrossRef]
- Sarraf, M.; Kataria, S.; Taimourya, H.; Santos, L.O.; Menegatti, R.D.; Jain, M.; Ihtisham, M.; Liu, S. Magnetic Field (MF) Applications in Plants: An Overview. Plants 2020, 9, 1139. [Google Scholar] [CrossRef]
- Shabrangy, A. Using Magnetic Fields to Enhance the Seed Germination, Growth, and Yield of Plants. In Plant Functional Genomics: Methods and Protocols; Maghuly, F., Ed.; Springer US: New York, NY, USA, 2024; Volume 2, pp. 375–395. [Google Scholar]
- Afzal, I.; Saleem, S.; Skalicky, M.; Javed, T.; Bakhtavar, M.A.; ul Haq, Z.; Kamran, M.; Shahid, M.; Sohail Saddiq, M.; Afzal, A.; et al. Magnetic Field Treatments Improves Sunflower Yield by Inducing Physiological and Biochemical Modulations in Seeds. Molecules 2021, 26, 2022. [Google Scholar] [CrossRef]
- Paponov, I.A.; Fliegmann, J.; Narayana, R.; Maffei, M.E. Differential root and shoot magnetoresponses in Arabidopsis thaliana. Sci. Rep. 2021, 11, 9195. [Google Scholar] [CrossRef]
- Rodrigo-Moreno, A.; Bazihizina, N.; Azzarello, E.; Masi, E.; Tran, D.; Bouteau, F.; Baluska, F.; Mancuso, S. Root phonotropism: Early signalling events following sound perception in Arabidopsis roots. Plant Sci. 2017, 264, 9–15. [Google Scholar] [CrossRef]
- Javed, R.; Khan, B.; Sharafat, U.; Bilal, M.; Galagedara, L.; Abbey, L.; Cheema, M. Dynamic interplay of metal and metal oxide nanoparticles with plants: Influencing factors, action mechanisms, and assessment of stimulatory and inhibitory effects. Ecotoxicol. Environ. Saf. 2024, 271, 115992. [Google Scholar] [CrossRef]
- Venegas-Molina, J.; Proietti, S.; Pollier, J.; Orozco-Freire, W.; Ramirez-Villacis, D.; Leon-Reyes, A. Induced tolerance to abiotic and biotic stresses of broccoli and Arabidopsis after treatment with elicitor molecules. Sci. Rep. 2020, 10, 10319. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. The interplay between light and jasmonate signalling during defence and development. J. Exp. Bot. 2011, 62, 4087–4100. [Google Scholar] [CrossRef]
- Adil, M.; Ren, X.; Jeong, B.R. Light elicited growth, antioxidant enzymes activities and production of medicinal compounds in callus culture of Cnidium officinale Makino. J. Photochem. Photobiol. B Biol. 2019, 196, 111509. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, V.; Pellegrino, A.; Muleo, R.; Forgione, I. Light and Plant Growth Regulators on In Vitro Proliferation. Plants 2022, 11, 844. [Google Scholar] [CrossRef]
- Hectors, K.; Prinsen, E.; De Coen, W.; Jansen, M.A.K.; Guisez, Y. Arabidopsis thaliana plants acclimated to low dose rates of ultraviolet B radiation show specific changes in morphology and gene expression in the absence of stress symptoms. New Phytol. 2007, 175, 255–270. [Google Scholar] [CrossRef]
- Aksakal, O.; Tabay, D.; Esringu, A.; Icoglu Aksakal, F.; Esim, N. Effect of proline on biochemical and molecular mechanisms in lettuce (Lactuca sativa L.) exposed to UV-B radiation. Photochem. Photobiol. Sci. 2017, 16, 246–254. [Google Scholar] [CrossRef]
- Yadav, D.; Tanveer, A.; Malviya, N.; Yadav, S. Chapter 1—Overview and Principles of Bioengineering: The Drivers of Omics Technologies. In Omics Technologies and Bio-Engineering; Barh, D., Azevedo, V., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 3–23. [Google Scholar]
- Appolloni, E.; Paucek, I.; Pennisi, G.; Manfrini, L.; Gabarrell, X.; Gianquinto, G.; Orsini, F. Winter Greenhouse Tomato Cultivation: Matching Leaf Pruning and Supplementary Lighting for Improved Yield and Precocity. Agronomy 2023, 13, 671. [Google Scholar] [CrossRef]
- Tiwari, V.; Kamara, I.; Ratner, K.; Many, Y.; Lukyanov, V.; Ziv, C.; Gilad, Z.; Esquira, I.; Charuvi, D. Daytime or Edge-of-Daytime Intra-Canopy Illumination Improves the Fruit Set of Bell Pepper at Passive Conditions in the Winter. Plants 2022, 11, 424. [Google Scholar] [CrossRef]
- Liu, W.; Zha, L.; Zhang, Y. Growth and Nutrient Element Content of Hydroponic Lettuce are Modified by LED Continuous Lighting of Different Intensities and Spectral Qualities. Agronomy 2020, 10, 1678. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef]
- Podleśny, J.; Podleśna, A.; Gładyszewska, B.; Bojarszczuk, J. Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings. Agronomy 2021, 11, 494. [Google Scholar] [CrossRef]
- Sun, P.; Zheng, F.; Wang, K.; Zhong, M.; Wu, D.; Zhu, H. Electro- and Magneto-Modulated Ion Transport through Graphene Oxide Membranes. Sci. Rep. 2014, 4, 6798. [Google Scholar] [CrossRef]
- Rajabbeigi, E.; Ghanati, F.; Abdolmaleki, P.; Payez, A. Antioxidant capacity of parsley cells (Petroselinum crispum L.) in relation to iron-induced ferritin levels and static magnetic field. Electromagn. Biol. Med. 2013, 32, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Del Stabile, F.; Marsili, V.; Forti, L.; Arru, L. Is There a Role for Sound in Plants? Plants 2022, 11, 2391. [Google Scholar] [CrossRef]
- Appel, H.M.; Cocroft, R.B. Plants respond to leaf vibrations caused by insect herbivore chewing. Oecologia 2014, 175, 1257–1266. [Google Scholar] [CrossRef]
- Jeong, M.-J.; Cho, J.-I.; Park, S.-H.; Kim, K.-H.; Lee, S.; Kwon, T.-R.; Park, S.-C.; Siddiqui, Z.S. Sound frequencies induce drought tolerance in rice plant. Pak. J. Bot. 2014, 46, 2015–2020. [Google Scholar]
- Fernandez-Jaramillo, A.A.; Duarte-Galvan, C.; Garcia-Mier, L.; Jimenez-Garcia, S.N.; Contreras-Medina, L.M. Effects of acoustic waves on plants: An agricultural, ecological, molecular and biochemical perspective. Sci. Hortic. 2018, 235, 340–348. [Google Scholar] [CrossRef]
- Telewski, F.W. A unified hypothesis of mechanoperception in plants. Am. J. Bot. 2006, 93, 1466–1476. [Google Scholar] [CrossRef]
- Lala, S. Nanoparticles as elicitors and harvesters of economically important secondary metabolites in higher plants: A review. IET Nanobiotechnol. 2021, 15, 28. [Google Scholar] [CrossRef]
- Moola, A.K.; Senthil Kumar, T.; Ranjitha Kumari, B.D. Enhancement of Celastrol compound by silver nanoparticles and acetosyringone in Celastrus paniculatus Willd. through adventitious and hairy root culture. J. Plant Biochem. Biotechnol. 2022, 31, 429–434. [Google Scholar] [CrossRef]
- Tian, L.; Shen, J.; Sun, G.; Wang, B.; Ji, R.; Zhao, L. Foliar Application of SiO(2) Nanoparticles Alters Soil Metabolite Profiles and Microbial Community Composition in the Pakchoi (Brassica chinensis L.) Rhizosphere Grown in Contaminated Mine Soil. Environ. Sci. Technol. 2020, 54, 13137–13146. [Google Scholar] [CrossRef]
- Xia, L.; Huang, H.; Feng, W.; Chen, Y. Silica nanoparticles boost plant resistance against pathogens. Sci. Bull. 2021, 66, 1151–1153. [Google Scholar] [CrossRef]
- Miteu, G.D.; Emmanuel, A.A.; Addeh, I.; Ojeokun, O.; Olayinka, T.; Godwin, J.S.; Adeyemo, O.I.; Benneth, E.O. Nanoscience and technology as a pivot for sustainable agriculture and its One Health approach awareness. Sci. One Health 2023, 2, 100020. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Berni, R.; Muñoz-Sanchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.A.; Cantini, C.; Cai, G.; Hausman, J.-F.; et al. Production of Plant Secondary Metabolites: Examples, Tips and Suggestions for Biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef]
- Jadid, N.; Widodo, A.F.; Ermavitalini, D.; Sa’adah, N.N.; Gunawan, S.; Nisa, C. The medicinal Umbelliferae plant Fennel (Foeniculum vulgare Mill.): Cultivation, traditional uses, phytopharmacological properties, and application in animal husbandry. Arab. J. Chem. 2023, 16, 104541. [Google Scholar] [CrossRef]
- Jadid, N.; Ramadani, M.R.N.; Widodo, A.F.; Sa’adah, N.N.; Ermavitalini, D.; Rahmawati, M.; Sari, S.A.; Soleha, I.D.; Mas’ud, F. In silico characterization of GbPAL, GbCHS, GbDFR and GbANS structural genes involved in the biosynthesis of flavonoids in Gynura bicolor DC. S. Afr. J. Bot. 2024, 165, 428–442. [Google Scholar] [CrossRef]
- Rohela, G.K.; Saini, P.; Aziz, D.; Rafiq, S.; Jogam, P.; Zhang, B. Nanoparticles as elicitors and stimulators for plant tissue culture, transgenics, and genome editing: A comprehensive review. Ind. Crops Prod. 2024, 222, 120097. [Google Scholar] [CrossRef]
- Kruszka, D.; Selvakesavan, R.K.; Kachlicki, P.; Franklin, G. Untargeted metabolomics analysis reveals the elicitation of important secondary metabolites upon treatment with various metal and metal oxide nanoparticles in Hypericum perforatum L. cell suspension cultures. Ind. Crops Prod. 2022, 178, 114561. [Google Scholar] [CrossRef]
- Yeow, L.C.; Chew, B.L.; Sreeramanan, S. Elevation of secondary metabolites production through light-emitting diodes (LEDs) illumination in protocorm-like bodies (PLBs) of Dendrobium hybrid orchid rich in phytochemicals with therapeutic effects. Biotechnol. Rep. 2020, 27, e00497. [Google Scholar] [CrossRef] [PubMed]
- Lian, T.T.; Moe, M.M.; Kim, Y.J.; Bang, K.S. Effects of Different Colored LEDs on the Enhancement of Biologically Active Ingredients in Callus Cultures of Gynura procumbens (Lour.) Merr. Molecules 2019, 24, 4336. [Google Scholar] [CrossRef] [PubMed]
- Poornananda, M.N.; Jameel, M.A.K. Abiotic and Biotic Elicitors–Role in Secondary Metabolites Production through In Vitro Culture of Medicinal Plants. In Abiotic and Biotic Stress in Plants; Arun, K.S., Chitra, S., Eds.; IntechOpen: Rijeka, Croatia, 2016; Chapter 10. [Google Scholar]
- Golovatskaya, I.F.; Karnachuk, R.A. Role of green light in physiological activity of plants. Russ. J. Plant Physiol. 2015, 62, 727–740. [Google Scholar] [CrossRef]
- Veits, M.; Khait, I.; Obolski, U.; Zinger, E.; Boonman, A.; Goldshtein, A.; Saban, K.; Seltzer, R.; Ben-Dor, U.; Estlein, P.; et al. Flowers respond to pollinator sound within minutes by increasing nectar sugar concentration. Ecol. Lett. 2019, 22, 1483–1492. [Google Scholar] [CrossRef]
- De Luca, P.A.; Vallejo-Marín, M. What’s the ‘buzz’ about? The ecology and evolutionary significance of buzz-pollination. Curr. Opin. Plant Biol. 2013, 16, 429–435. [Google Scholar] [CrossRef]
- Appel, H.; Cocroft, R. Plant ecoacoustics: A sensory ecology approach. Trends Ecol. Evol. 2023, 38, 623–630. [Google Scholar] [CrossRef]
- Mann, V.; Harker, M.; Pecker, I.; Hirschberg, J. Metabolic engineering of astaxanthin production in tobacco flowers. Nat. Biotechnol. 2000, 18, 888–892. [Google Scholar] [CrossRef]
- Sato, F.; Hashimoto, T.; Hachiya, A.; Tamura, K.; Choi, K.B.; Morishige, T.; Fujimoto, H.; Yamada, Y. Metabolic engineering of plant alkaloid biosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 367–372. [Google Scholar] [CrossRef]
- Vom Endt, D.; Kijne, J.W.; Memelink, J. Transcription factors controlling plant secondary metabolism: What regulates the regulators? Phytochemistry 2002, 61, 107–114. [Google Scholar] [CrossRef]
- Chugh, V.; Kaur, D.; Purwar, S.; Kaushik, P.; Sharma, V.; Kumar, H.; Rai, A.; Singh, C.M.; Kamaluddin; Dubey, R.B. Applications of Molecular Markers for Developing Abiotic-Stress-Resilient Oilseed Crops. Life 2023, 13, 88. [Google Scholar] [CrossRef]
- Sinaga, A.O.Y.; Marpaung, D.S.S. Abiotic stress-induced gene expression in pineapple as a potential genetic marker. Adv. Agrochem. 2024, 3, 133–142. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, Z.; Hu, Z.; Wang, T.; Teng, L.; Dai, T.; Liu, P.; Hao, J.; Liu, X. Utilizing metabolomic approach to study the mode of action of fungicides and corresponding resistance in plant pathogens. Adv. Agrochem. 2024, 3, 197–205. [Google Scholar] [CrossRef]
- Padilla, Y.G.; Miras-Moreno, B.; Gisbert-Mullor, R.; Lucini, L.; López-Galarza, S.; Calatayud, Á. Leaves and roots metabolomic signatures underlying rootstock-mediated water stress tolerance in grafted pepper plants. Plant Stress 2024, 13, 100542. [Google Scholar] [CrossRef]
- Gong, X.; Li, X.; Xia, Y.; Xu, J.; Li, Q.; Zhang, C.; Li, M. Effects of phytochemicals from plant-based functional foods on hyperlipidemia and their underpinning mechanisms. Trends Food Sci. Technol. 2020, 103, 304–320. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Gómez-Cadenas, A.; Mittler, R. Plant responses to climate change: Metabolic changes under combined abiotic stresses. J. Exp. Bot. 2022, 73, 3339–3354. [Google Scholar] [CrossRef]
- Oms-Oliu, G.; Odriozola-Serrano, I.; Martín-Belloso, O. Metabolomics for assessing safety and quality of plant-derived food. Food Res. Int. 2013, 54, 1172–1183. [Google Scholar] [CrossRef]
- Oh, S.; Lu, C. Vertical farming—Smart urban agriculture for enhancing resilience and sustainability in food security. J. Hortic. Sci. Biotechnol. 2023, 98, 133–140. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef]
- Naik, P.M.; Al-Khayri, J.M. Impact of Abiotic Elicitors on In vitro Production of Plant Secondary Metabolites: A Review. J. Adv. Res. Biotech. 2016, 1, 7. [Google Scholar]
- Zhao, J.L.; Zou, L.; Zhang, C.Q.; Li, Y.Y.; Peng, L.X.; Xiang, D.B.; Zhao, G. Efficient production of flavonoids in Fagopyrum tataricum hairy root cultures with yeast polysaccharide elicitation and medium renewal process. Pharmacogn. Mag. 2014, 10, 234–240. [Google Scholar] [CrossRef]
- Ming, Q.; Su, C.; Zheng, C.; Jia, M.; Zhang, Q.; Zhang, H.; Rahman, K.; Han, T.; Qin, L. Elicitors from the endophytic fungus Trichoderma atroviride promote Salvia miltiorrhiza hairy root growth and tanshinone biosynthesis. J. Exp. Bot. 2013, 64, 5687–5694. [Google Scholar] [CrossRef]
- Li, B.; Wang, B.; Li, H.; Peng, L.; Ru, M.; Liang, Z.; Yan, X.; Zhu, Y. Establishment of Salvia castanea Diels f. tomentosa Stib. hairy root cultures and the promotion of tanshinone accumulation and gene expression with Ag+, methyl jasmonate, and yeast extract elicitation. Protoplasma 2016, 253, 87–100. [Google Scholar] [CrossRef]
- Jiao, J.; Gai, Q.-Y.; Wang, X.; Qin, Q.-P.; Wang, Z.-Y.; Liu, J.; Fu, Y.-J. Chitosan elicitation of Isatis tinctoria L. hairy root cultures for enhancing flavonoid productivity and gene expression and related antioxidant activity. Ind. Crops Prod. 2018, 124, 28–35. [Google Scholar] [CrossRef]
- Hashemi, S.M.; Naghavi, M.R. Production and gene expression of morphinan alkaloids in hairy root culture of Papaver orientale L. using abiotic elicitors. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 125, 31–41. [Google Scholar] [CrossRef]
- Liu, R.; Bassalo, M.C.; Zeitoun, R.I.; Gill, R.T. Genome scale engineering techniques for metabolic engineering. Metab. Eng. 2015, 32, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, K.; Martínez-Chávez, L.A.; Rosales-Pérez, A.; Hernández-Rangel, R. Chapter 1 Nanomaterials: Classification, synthesis methods, and physicochemical characterization. In Engineered Nanoparticles in Agriculture; Walter de Gruyter GmbH: Berlin, Germany, 2023; pp. 1–58. [Google Scholar] [CrossRef]
- Bratovcic, A.; Hikal, W.M.; Mehdizadeh, M.; Hussein, A.H.S.A.A.; Omidi, A.; Adetunji, C.O.; Omorefosa, O.O.; Bera, A. Application of Nanotechnology in Agroecosystems: Nanoparticles for Improving Agricultural Production. Rev. Agric. Sci. 2023, 11, 291–309. [Google Scholar] [CrossRef] [PubMed]
- Santás-Miguel, V.; Arias-Estévez, M.; Rodríguez-Seijo, A.; Arenas-Lago, D. Use of metal nanoparticles in agriculture. A review on the effects on plant germination. Environ. Pollut. 2023, 334, 122222. [Google Scholar] [CrossRef]
- Ndaba, B.; Roopnarain, A.; Rama, H.; Maaza, M. Biosynthesized metallic nanoparticles as fertilizers: An emerging precision agriculture strategy. J. Integr. Agric. 2022, 21, 1225–1242. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef]
- Vargas-Ortiz, J.R.; Gonzalez, C.; Esquivel, K. Magnetic Iron Nanoparticles: Synthesis, Surface Enhancements, and Biological Challenges. Processes 2022, 10, 2282. [Google Scholar] [CrossRef]
- Saleh, T.A. Nanomaterials: Classification, properties, and environmental toxicities. Environ. Technol. Innov. 2020, 20, 101067. [Google Scholar] [CrossRef]
- Baig, N.; Kammakakam, I.; Falath, W.; Kammakakam, I. Nanomaterials: A review of synthesis methods, properties, recent progress, and challenges. Mater. Adv. 2021, 2, 1821–1871. [Google Scholar] [CrossRef]
- Singh, V.; Yadav, P.; Mishra, V. Recent Advances on Classification, Properties, Synthesis, and Characterization of Nanomaterials. In Green Synthesis of Nanomaterials for Bioenergy Applications; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 83–97. [Google Scholar] [CrossRef]
- Khan, Y.; Sadia, H.; Ali Shah, S.Z.; Khan, M.N.; Shah, A.A.; Ullah, N.; Ullah, M.F.; Bibi, H.; Bafakeeh, O.T.; Khedher, N.B.; et al. Classification, Synthetic, and Characterization Approaches to Nanoparticles, and Their Applications in Various Fields of Nanotechnology: A Review. Catalysts 2022, 12, 1386. [Google Scholar] [CrossRef]
- Sudha, P.N.; Sangeetha, K.; Vijayalakshmi, K.; Barhoum, A. Nanomaterials history, classification, unique properties, production and market. In Emerging Applications of Nanoparticles and Architecture Nanostructures; Elsevier: Amsterdam, The Netherlands, 2018; pp. 341–384. [Google Scholar] [CrossRef]
- Mekuye, B.; Abera, B. Nanomaterials: An overview of synthesis, classification, characterization, and applications. Nano Sel. 2023, 4, 486–501. [Google Scholar] [CrossRef]
- Hassanien, H.; Darweesh, M. Nanomaterials: Classification and Properties-Part I. J. Nanosci. 2018, 1, 1–11. [Google Scholar]
- García-Ovando, A.E.; Ramírez Piña, J.E.; Esquivel Naranjo, E.U.; Cervantes Chávez, J.A.; Esquivel, K. Biosynthesized nanoparticles and implications by their use in crops: Effects over physiology, action mechanisms, plant stress responses and toxicity. Plant Stress 2022, 6, 109. [Google Scholar] [CrossRef]
- Asha, A.B.; Narain, R. Chapter 15—Nanomaterials properties. In Polymer Science and Nanotechnology; Narain, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 343–359. [Google Scholar]
- Karunakaran, G.; Sudha, K.G.; Ali, S.; Cho, E.-B. Biosynthesis of Nanoparticles from Various Biological Sources and Its Biomedical Applications. Molecules 2023, 28, 4527. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Shariq, M.; Asif, M.; Siddiqui, M.A.; Malan, P.; Ahmad, F. Green Nanotechnology: Plant-Mediated Nanoparticle Synthesis and Application. Nanomaterials 2022, 12, 673. [Google Scholar] [CrossRef]
- Rani, N.; Kumari, K.; Sangwan, P.; Barala, P.; Yadav, J.; Vijeta; Rahul; Hooda, V. Nano-Iron and Nano-Zinc Induced Growth and Metabolic Changes in Vigna radiata. Sustainability 2022, 14, 8251. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Zia ur Rehman, M.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Cruz-Luna, A.R.; Cruz-Martínez, H.; Vásquez-López, A.; Medina, D.I. Metal nanoparticles as novel antifungal agents for sustainable agriculture: Current advances and future directions. J. Fungi 2021, 7, 1033. [Google Scholar] [CrossRef]
- Gandhi, N.; Shruthi, Y.; Sirisha, G.; Anusha, C.R. Facile and Eco-Friendly Method for Synthesis of Calcium Oxide (CaO) Nanoparticles and its Potential Application in Agriculture. Haya Saudi J. Life Sci. 2021, 6, 89–103. [Google Scholar]
- Sheoran, P.; Goel, S.; Boora, R.; Kumari, S.; Yashveer, S.; Grewal, S. Biogenic synthesis of potassium nanoparticles and their evaluation as a growth promoter in wheat. Plant Gene 2021, 27, 310. [Google Scholar] [CrossRef]
- Roy, A.; Gauri, S.S.; Bhattacharya, M.; Bhattacharya, J. Antimicrobial activity of CaO nanoparticles. J. Biomed. Nanotechnol. 2013, 9, 1570–1578. [Google Scholar] [CrossRef]
- Wang, X.; Sun, W.; Zhang, S.; Sharifan, H.; Ma, X. Elucidating the Effects of Cerium Oxide Nanoparticles and Zinc Oxide Nanoparticles on Arsenic Uptake and Speciation in Rice (Oryza sativa) in a Hydroponic System. Environ. Sci. Technol. 2018, 52, 10040–10047. [Google Scholar] [CrossRef] [PubMed]
- Chhipa, H. Nanofertilizers and nanopesticides for agriculture. Environ. Chem. Lett. 2017, 15, 15–22. [Google Scholar] [CrossRef]
- Ferrusquía-Jiménez, N.I.; González-Arias, B.; Rosales, A.; Esquivel, K.; Escamilla-Silva, E.M.; Ortega-Torres, A.E.; Guevara-González, R.G. Elicitation of Bacillus cereus-Amazcala (B.c-A) with SiO2 Nanoparticles Improves Its Role as a Plant Growth-Promoting Bacteria (PGPB) in Chili Pepper Plants. Plants 2022, 11, 3445. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Ali, S.; Rizwan, M.; Rehman, M.Z.u.; Qayyum, M.F.; Wang, H.; Rinklebe, J. Responses of wheat (Triticum aestivum) plants grown in a Cd contaminated soil to the application of iron oxide nanoparticles. Ecotoxicol. Environ. Saf. 2019, 173, 156–164. [Google Scholar] [CrossRef]
- Manzoor, N.; Ahmed, T.; Noman, M.; Shahid, M.; Nazir, M.M.; Ali, L.; Alnusaire, T.S.; Li, B.; Schulin, R.; Wang, G. Iron oxide nanoparticles ameliorated the cadmium and salinity stresses in wheat plants, facilitating photosynthetic pigments and restricting cadmium uptake. Sci. Total Environ. 2021, 769, 145221. [Google Scholar] [CrossRef]
- Feng, Y.; Kreslavski, V.D.; Shmarev, A.N.; Ivanov, A.A.; Zharmukhamedov, S.K.; Kosobryukhov, A.; Yu, M.; Allakhverdiev, S.I.; Shabala, S. Effects of Iron Oxide Nanoparticles (Fe3O4) on Growth, Photosynthesis, Antioxidant Activity and Distribution of Mineral Elements in Wheat (Triticum aestivum) Plants. Plants 2022, 11, 1894. [Google Scholar] [CrossRef]
- Sundaria, N.; Singh, M.; Upreti, P.; Chauhan, R.P.; Jaiswal, J.P.; Kumar, A. Seed Priming with Iron Oxide Nanoparticles Triggers Iron Acquisition and Biofortification in Wheat (Triticum aestivum L.) Grains. J. Plant Growth Regul. 2019, 38, 122–131. [Google Scholar] [CrossRef]
- Li, J.; Hu, J.; Ma, C.; Wang, Y.; Wu, C.; Huang, J.; Xing, B. Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 2016, 159, 326–334. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of metal and metal oxide nanoparticles on plant: A critical review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T.; et al. Iron oxide nanoparticles as a potential iron fertilizer for peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef]
- Tariverdizadeh, N.; Mohebodini, M.; Chamani, E.; Ebadi, A. Iron and zinc oxide nanoparticles: An efficient elicitor to enhance trigonelline alkaloid production in hairy roots of fenugreek. Ind. Crops Prod. 2021, 162, 113240. [Google Scholar] [CrossRef]
- Yousaf, N.; Ishfaq, M.; Qureshi, H.A.; Saleem, A.; Yang, H.; Sardar, M.F.; Zou, C. Characterization of Root and Foliar-Applied Iron Oxide Nanoparticles (α-Fe2O3, γ-Fe2O3, Fe3O4, and Bulk Fe3O4) in Improving Maize (Zea mays L.) Performance. Nanomaterials 2023, 13, 3036. [Google Scholar] [CrossRef] [PubMed]
- Suazo-Hernández, J.; Arancibia-Miranda, N.; Mlih, R.; Cáceres-Jensen, L.; Bolan, N.; Mora, M.d.l.L. Impact on Some Soil Physical and Chemical Properties Caused by Metal and Metallic Oxide Engineered Nanoparticles: A Review. Nanomaterials 2023, 13, 572. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Dou, F.; Li, C.; Ma, X.; Ma, L.Q. Impacts of metallic nanoparticles and transformed products on soil health. Crit. Rev. Environ. Sci. Technol. 2021, 51, 973–1002. [Google Scholar] [CrossRef]
- Ameen, F.; Alsamhary, K.; Alabdullatif, J.A.; Alnadhari, S. A review on metal-based nanoparticles and their toxicity to beneficial soil bacteria and fungi. Ecotoxicol. Environ. Saf. 2021, 213, 112027. [Google Scholar] [CrossRef]
- Ahmed, B.; Rizvi, A.; Syed, A.; Jailani, A.; Elgorban, A.M.; Khan, M.S.; Al-Shwaiman, H.A.; Lee, J. Differential bioaccumulations and ecotoxicological impacts of metal-oxide nanoparticles, bulk materials, and metal-ions in cucumbers grown in sandy clay loam soil. Environ. Pollut. 2021, 289, 117854. [Google Scholar] [CrossRef]
- Peng, C.; Tong, H.; Shen, C.; Sun, L.; Yuan, P.; He, M.; Shi, J. Bioavailability and translocation of metal oxide nanoparticles in the soil-rice plant system. Sci. Total Environ. 2020, 713, 136662. [Google Scholar] [CrossRef]
- Lee, H.; Song, M.Y.; Jurng, J.; Park, Y.K. The synthesis and coating process of TiO2 nanoparticles using CVD process. Powder Technol. 2011, 214, 64–68. [Google Scholar] [CrossRef]
- Rane, A.V.; Kanny, K.; Abitha, V.K.; Thomas, S. Methods for Synthesis of Nanoparticles and Fabrication of Nanocomposites. Synthesis of Inorganic Nanomaterials; Woodhead Publishing: Sawston, UK, 2018; pp. 121–139. [Google Scholar] [CrossRef]
- Ealias, A.M.; Saravanakumar, M.P. A review on the classification, characterisation, synthesis of nanoparticles and their application. IOP Conf. Ser. Mater. Sci. Eng. 2017, 263, 032019. [Google Scholar] [CrossRef]
- Paramasivam, G.; Palem, V.V.; Sundaram, T.; Sundaram, V.; Kishore, S.C.; Bellucci, S. Nanomaterials: Synthesis and Applications in Theranostics. Nanomaterials 2021, 11, 3228. [Google Scholar] [CrossRef]
- Azar, S.M.A.; Mousa, A.A. 8—Mechanical and physical methods for the metal oxide powders production. In Metal Oxide Powder Technologies; Al-Douri, Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 169–187. [Google Scholar]
- Kalyani, N.T.; Dhoble, S.J. 1—Introduction to nano materials. In Quantum Dots; Thejo Kalyani, N., Dhoble, S.J., Michalska-Domańska, M., Vengadaesvaran, B., Nagabhushana, H., Arof, A.K., Eds.; Woodhead Publishing: Sawston, UK, 2023; pp. 3–40. [Google Scholar]
- Chen, X.; Gao, C.; Guo, L.; Hu, G.; Luo, Q.; Liu, J.; Nielsen, J.; Chen, J.; Liu, L. DCEO Biotechnology: Tools to Design, Construct, Evaluate, and Optimize the Metabolic Pathway for Biosynthesis of Chemicals. Chem. Rev. 2018, 118, 4–72. [Google Scholar] [CrossRef] [PubMed]
- Bhati, M. Biogenic synthesis of metallic nanoparticles: Principles and applications. Mater. Today Proc. 2021, 81, 882–887. [Google Scholar] [CrossRef]
- Vijayaraghavan, K.; Ashokkumar, T. Plant-mediated biosynthesis of metallic nanoparticles: A review of literature, factors affecting synthesis, characterization techniques and applications. J. Environ. Chem. Eng. 2017, 5, 4866–4883. [Google Scholar] [CrossRef]
- Khanna, P.; Kaur, A.; Goyal, D. Algae-based metallic nanoparticles: Synthesis, characterization and applications. J. Microbiol. Methods 2019, 163, 105656. [Google Scholar] [CrossRef]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles alter secondary metabolism in plants via ROS burst. Front. Plant Sci. 2017, 8, 832. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Shabasy, R.M.; Khalifa, S.A.M.; Saeed, A.; Shah, A.; Shah, R.; Iftikhar, F.J.; Abdel-Daim, M.M.; Omri, A.; Hajrahand, N.H.; et al. Metal nanoparticles fabricated by green chemistry using natural extracts: Biosynthesis, mechanisms, and applications. RSC Adv. 2019, 9, 24539–24559. [Google Scholar] [CrossRef]
- Aqeel Salim, A.; Bidin, N.; Bakhtiar, H.; Krishna Ghoshal, S.; Al Azawi, M.; Krishnan, G. Optical and structure characterization of cinnamon nanoparticles synthesized by pulse laser ablation in liquid (PLAL). J. Phys. Conf. Ser. 2018, 1027, 012002. [Google Scholar] [CrossRef]
- Nyabadza, A.; Vazquez, M.; Brabazon, D. A Review of Bimetallic and Monometallic Nanoparticle Synthesis via Laser Ablation in Liquid. Crystals 2023, 13, 253. [Google Scholar] [CrossRef]
- Fazio, E.; Gökce, B.; De Giacomo, A.; Meneghetti, M.; Compagnini, G.; Tommasini, M.; Waag, F.; Lucotti, A.; Zanchi, C.G.; Ossi, P.M.; et al. Nanoparticles Engineering by Pulsed Laser Ablation in Liquids: Concepts and Applications. Nanomaterials 2020, 10, 2317. [Google Scholar] [CrossRef]
- Khairani, I.Y.; Mínguez-Vega, G.; Doñate-Buendía, C.; Gökce, B. Green nanoparticle synthesis at scale: A perspective on overcoming the limits of pulsed laser ablation in liquids for high-throughput production. Phys. Chem. Chem. Phys. 2023, 25, 19380–19408. [Google Scholar] [CrossRef]
- Jamkhande, P.G.; Ghule, N.W.; Bamer, A.H.; Kalaskar, M.G. Metal nanoparticles synthesis: An overview on methods of preparation, advantages and disadvantages, and applications. J. Drug Deliv. Sci. Technol. 2019, 53, 101174. [Google Scholar] [CrossRef]
- Mittal, A.K.; Chisti, Y.; Banerjee, U.C. Synthesis of metallic nanoparticles using plant extracts. Biotechnol. Adv. 2013, 31, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, S.; Kodam, K.; Dhaygude, M.; Hudlikar, M. Novel route for rapid biosynthesis of lead nanoparticles using aqueous extract of Jatropha curcas L. latex. Mater. Lett. 2011, 65, 3170–3172. [Google Scholar] [CrossRef]
- Laokula, P.; Klinkaewnaronga, J.; Phokha, S.; Seraphin, S. Indium oxide (In2O3) nanoparticles using Aloe vera plant extract: Synthesis and optical properties. Optoelectron. Adv. Mater. Rapid Commun. 2008, 2, 101174. [Google Scholar]
- Torres-Limiñana, J.; Feregrino-Pérez, A.A.; Vega-González, M.; Escobar-Alarcón, L.; Cervantes-Chávez, J.A.; Esquivel, K. Green Synthesis via Eucalyptus globulus L. Extract of Ag-TiO2 Catalyst: Antimicrobial Activity Evaluation toward Water Disinfection Process. Nanomaterials 2022, 12, 1944. [Google Scholar] [CrossRef]
- Jadoun, S.; Arif, R.; Jangid, N.K.; Meena, R.K. Green synthesis of nanoparticles using plant extracts: A review. Environ. Chem. Lett. 2021, 19, 355–374. [Google Scholar] [CrossRef]
- Ahamed, M.; Majeed Khan, M.A.; Siddiqui, M.K.J.; Alsalhi, M.S.; Alrokayan, S.A. Green synthesis, characterization and evaluation of biocompatibility of silver nanoparticles. Phys. E Low-Dimens. Syst. Nanostructures 2011, 43, 1266–1271. [Google Scholar] [CrossRef]
- Bankar, A.; Joshi, B.; Kumar, A.R.; Zinjarde, S. Banana peel extract mediated novel route for the synthesis of silver nanoparticles. Colloids Surf. A Physicochem. Eng. Asp. 2010, 368, 58–63. [Google Scholar] [CrossRef]
- Bar, H.; Bhui, D.K.; Sahoo, G.P.; Sarkar, P.; De, S.P.; Misra, A. Green synthesis of silver nanoparticles using latex of Jatropha curcas. Colloids Surf. A Physicochem. Eng. Asp. 2009, 339, 134–139. [Google Scholar] [CrossRef]
- Dwivedi, A.D.; Gopal, K. Biosynthesis of silver and gold nanoparticles using Chenopodium album leaf extract. Colloids Surf. A Physicochem. Eng. Asp. 2010, 369, 27–33. [Google Scholar] [CrossRef]
- Thatyana, M.; Dube, N.P.; Kemboi, D.; Manicum, A.-L.E.; Mokgalaka-Fleischmann, N.S.; Tembu, J.V. Advances in Phytonanotechnology: A Plant-Mediated Green Synthesis of Metal Nanoparticles Using Phyllanthus Plant Extracts and Their Antimicrobial and Anticancer Applications. Nanomaterials 2023, 13, 2616. [Google Scholar] [CrossRef] [PubMed]
- Korbekandi, H.; Iravani, S.; Abbasi, S. Production of nanoparticles using organisms. Crit. Rev. Biotechnol. 2009, 29, 279–306. [Google Scholar] [CrossRef] [PubMed]
- Sangar, S.; Sharma, S.; Vats, V.K.; Mehta, S.K.; Singh, K. Biosynthesis of silver nanocrystals, their kinetic profile from nucleation to growth and optical sensing of mercuric ions. J. Clean. Prod. 2019, 228, 294–302. [Google Scholar] [CrossRef]
- Vanlalveni, C.; Lallianrawna, S.; Biswas, A.; Selvaraj, M.; Changmai, B.; Rokhum, S.L. Green synthesis of silver nanoparticles using plant extracts and their antimicrobial activities: A review of recent literature. RSC Adv. 2021, 11, 2804–2837. [Google Scholar] [CrossRef] [PubMed]
- Antunes Filho, S.; dos Santos, M.S.; dos Santos, O.A.L.; Backx, B.P.; Soran, M.-L.; Opriş, O.; Lung, I.; Stegarescu, A.; Bououdina, M. Biosynthesis of Nanoparticles Using Plant Extracts and Essential Oils. Molecules 2023, 28, 3060. [Google Scholar] [CrossRef]
- El-Kemary, M.; Zahran, M.; Khalifa, S.A.M.; El-Seedi, H.R. Spectral characterisation of the silver nanoparticles biosynthesised using Ambrosia maritima plant. Micro Nano Lett. 2016, 11, 311–314. [Google Scholar] [CrossRef]
- Kora, A.J.; Rastogi, L. Green synthesis of palladium nanoparticles using gum ghatti (Anogeissus latifolia) and its application as an antioxidant and catalyst. Arab. J. Chem. 2018, 11, 1097–1106. [Google Scholar] [CrossRef]
- Zheng, B.; Kong, T.; Jing, X.; Odoom-Wubah, T.; Li, X.; Sun, D.; Lu, F.; Zheng, Y.; Huang, J.; Li, Q. Plant-mediated synthesis of platinum nanoparticles and its bioreductive mechanism. J. Colloid. Interface Sci. 2013, 396, 138–145. [Google Scholar] [CrossRef]
- Mishra, D.; Chitara, M.K.; Negi, S.; Pal Singh, J.; Kumar, R.; Chaturvedi, P. Biosynthesis of Zinc Oxide Nanoparticles via Leaf Extracts of Catharanthus roseus (L.) G. Don and Their Application in Improving Seed Germination Potential and Seedling Vigor of Eleusine coracana (L.) Gaertn. Adv. Agric. 2023, 2023, 7412714. [Google Scholar] [CrossRef]
- Luty-Błocho, M.; Cyndrowska, J.; Rutkowski, B.; Hessel, V. Synthesis of Gold Clusters and Nanoparticles Using Cinnamon Extract—A Mechanism and Kinetics Study. Molecules 2024, 29, 1426. [Google Scholar] [CrossRef]
- Vidyasagar; Patel, R.R.; Singh, S.K.; Dehari, D.; Nath, G.; Singh, M. Facile green synthesis of silver nanoparticles derived from the medicinal plant Clerodendrum serratum and its biological activity against Mycobacterium species. Heliyon 2024, 10, e31116. [Google Scholar] [CrossRef] [PubMed]
- Bahattab, O.; Khan, I.; Bawazeer, S.; Rauf, A.; Qureshi, M.N.; Al-Awthan, Y.S.; Muhammad, N.; Khan, A.; Akram, M.; Islam, M.N.; et al. Synthesis and biological activities of alcohol extract of black cumin seeds (Bunium persicum)-based gold nanoparticles and their catalytic applications. Green. Process. Synth. 2021, 10, 440–455. [Google Scholar] [CrossRef]
- Gnanasekaran, R.; Yuvaraj, D.; Reddy, G.K.; Shangar, S.N.; Vijayakumar, V.; Iyyappan, J. Zinc oxide nanoparticles from leaf extract of Eclipta prostrata: Biosynthesis and characterization towards potential agent against film forming bacteria in metal working fluids. Environ. Chem. Ecotoxicol. 2024, 6, 206–215. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, W.; Tan, X.; Wang, Z.; Li, Y.; Li, W. Extract of Ginkgo biloba leaves mediated biosynthesis of catalytically active and recyclable silver nanoparticles. Colloids Surf. A Physicochem. Eng. Asp. 2019, 563, 31–36. [Google Scholar] [CrossRef]
- Aravind, M.; Amalanathan, M.; Mary, M.S.M. Synthesis of TiO2 nanoparticles by chemical and green synthesis methods and their multifaceted properties. SN Appl. Sci. 2021, 3, 409. [Google Scholar] [CrossRef]
- Patil, M.P.; Bayaraa, E.; Subedi, P.; Piad, L.L.A.; Tarte, N.H.; Kim, G.-D. Biogenic synthesis, characterization of gold nanoparticles using Lonicera japonica and their anticancer activity on HeLa cells. J. Drug Deliv. Sci. Technol. 2019, 51, 83–90. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Current status of plant metabolite-based fabrication of copper/copper oxide nanoparticles and their applications: A review. Biomater. Res. 2020, 24, 11. [Google Scholar] [CrossRef] [PubMed]
- Samari, F.; Salehipoor, H.; Eftekhar, E.; Yousefinejad, S. Low-temperature biosynthesis of silver nanoparticles using mango leaf extract: Catalytic effect, antioxidant properties, anticancer activity and application for colorimetric sensing. New J. Chem. 2018, 42, 15905–15916. [Google Scholar] [CrossRef]
- Sundrarajan, M.; Bama, K.; Bhavani, M.; Jegatheeswaran, S.; Ambika, S.; Sangili, A.; Nithya, P.; Sumathi, R. Obtaining titanium dioxide nanoparticles with spherical shape and antimicrobial properties using M. citrifolia leaves extract by hydrothermal method. J. Photochem. Photobiol. B Biol. 2017, 171, 117–124. [Google Scholar] [CrossRef]
- Ngom, I.; Ndiaye, N.M.; Fall, A.; Bakayoko, M.; Ngom, B.D.; Maaza, M. On the Use of Moringa Oleifera Leaves Extract for the Biosynthesis of NiO and ZnO Nanoparticles. MRS Adv. 2020, 5, 1145–1155. [Google Scholar] [CrossRef]
- Selvaraj, R.; Nagendran, V.; Varadavenkatesan, T.; Goveas, L.C.; Vinayagam, R. Stable silver nanoparticles synthesis using Tabebuia aurea leaf extract for efficient water treatment: A sustainable approach to environmental remediation. Chem. Eng. Res. Des. 2024, 208, 456–463. [Google Scholar] [CrossRef]
- Vincent, J.; Lau, K.S.; Evyan, Y.C.-Y.; Chin, S.X.; Sillanpää, M.; Chia, C.H. Biogenic Synthesis of Copper-Based Nanomaterials Using Plant Extracts and Their Applications: Current and Future Directions. Nanomaterials 2022, 12, 3312. [Google Scholar] [CrossRef] [PubMed]
- Bolade, O.P.; Williams, A.B.; Benson, N.U. Green synthesis of iron-based nanomaterials for environmental remediation: A review. Environ. Nanotechnol. Monit. Manag. 2020, 13, 100279. [Google Scholar] [CrossRef]
- Bhuiyan, M.S.H.; Miah, M.Y.; Paul, S.C.; Aka, T.D.; Saha, O.; Rahaman, M.M.; Sharif, M.J.I.; Habiba, O.; Ashaduzzaman, M. Green synthesis of iron oxide nanoparticle using Carica papaya leaf extract: Application for photocatalytic degradation of remazol yellow RR dye and antibacterial activity. Heliyon 2020, 6, e04603. [Google Scholar] [CrossRef]
- Patiño-Ruiz, D.; Sánchez-Botero, L.; Tejeda-Benitez, L.; Hinestroza, J.; Herrera, A. Green synthesis of iron oxide nanoparticles using Cymbopogon citratus extract and sodium carbonate salt: Nanotoxicological considerations for potential environmental applications. Environ. Nanotechnol. Monit. Manag. 2020, 14, 100377. [Google Scholar] [CrossRef]
- Huang, Y.; Haw, C.Y.; Zheng, Z.; Kang, J.; Zheng, J.-C.; Wang, H.-Q. Biosynthesis of Zinc Oxide Nanomaterials from Plant Extracts and Future Green Prospects: A Topical Review. Adv. Sustain. Syst. 2021, 5, 2000266. [Google Scholar] [CrossRef]
- Hatipoğlu, A.; Baran, A.; Keskin, C.; Baran, M.F.; Eftekhari, A.; Omarova, S.; Janas, D.; Khalilov, R.; Adican, M.T.; Kandemir, S.İ. Green synthesis of silver nanoparticles based on the Raphanus sativus leaf aqueous extract and their toxicological/microbiological activities. Environ. Sci. Pollut. Res. 2023. [Google Scholar] [CrossRef]
- Raut, R.W.; Mendhulkar, V.D.; Kashid, S.B. Photosensitized synthesis of silver nanoparticles using Withania somnifera leaf powder and silver nitrate. J. Photochem. Photobiol. B Biol. 2014, 132, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Montejo, S.d.J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as novel elicitors to improve bioactive compounds in plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A.; Rao, R.A.K. A review on biosynthesis of silver nanoparticles and their biocidal properties. J. Nanobiotechnol. 2018, 16, 14. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Mathur, P.; Roy, S. Nanosilica facilitates silica uptake, growth and stress tolerance in plants. Plant Physiol. Biochem. 2020, 157, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Semenova, N.A.; Burmistrov, D.E.; Shumeyko, S.A.; Gudkov, S.V. Fertilizers Based on Nanoparticles as Sources of Macro- and Microelements for Plant Crop Growth: A Review. Agronomy 2024, 14, 1646. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Fatemi, M.; Mollania, N.; Momeni-Moghaddam, M.; Sadeghifar, F. Extracellular biosynthesis of magnetic iron oxide nanoparticles by Bacillus cereus strain HMH1: Characterization and in vitro cytotoxicity analysis on MCF-7 and 3T3 cell lines. J. Biotechnol. 2018, 270, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Burman, U.; Kumar, P. Plant Response to Engineered Nanoparticles. Nanomater. Plants Algae Microorg. 2018, 1, 103–118. [Google Scholar] [CrossRef]
- López-Moreno, M.L.; De La Rosa, G.; Hernández-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. XAS Corroboration of the Uptake and Storage of CeO2 Nanoparticles and Assessment of their Differential Toxicity in Four Edible Plant Species. J. Agric. Food Chem. 2010, 58, 3689. [Google Scholar] [CrossRef]
- Gwinn, M.R.; Vallyathan, V. Nanoparticles: Health effects—Pros and cons. Environ. Health Perspect. 2006, 114, 1818–1825. [Google Scholar] [CrossRef]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, Uptake, and Translocation of Engineered Nanomaterials in Vascular plants. Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Shweta; Singh, S.; Singh, S.; Pandey, R.; Singh, V.P.; Sharma, N.C.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. An overview on manufactured nanoparticles in plants: Uptake, translocation, accumulation and phytotoxicity. Plant Physiol. Biochem. PPB 2017, 110, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. Impacts of Silver Nanoparticles on Plants: A Focus on the Phytotoxicity and Underlying Mechanism. Int. J. Mol. Sci. 2019, 20, 1003. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Meza, D.Y.; Guevara-González, R.G.; Esquivel, K.; Carbajal-Valenzuela, I.; Avila-Quezada, G.D. Green silver nanoparticles display protection against Clavibacter michiganensis subsp. michiganensis in tomato plants (Solanum lycopersicum L.). Plant Stress 2023, 10, 100256. [Google Scholar] [CrossRef]
- Müller, A.; Behsnilian, D.; Walz, E.; Gräf, V.; Hogekamp, L.; Greiner, R. Effect of culture medium on the extracellular synthesis of silver nanoparticles using Klebsiella pneumoniae, Escherichia coli and Pseudomonas jessinii. Biocatal. Agric. Biotechnol. 2016, 6, 107–115. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Raza, M.A.; Rizwan, M.; Tariq, R.; Ali, S.; Huang, L. Silver nanoparticles improved the plant growth and reduced the sodium and chlorine accumulation in pearl millet: A life cycle study. Environ. Sci. Pollut. Res. 2021, 28, 13712–13724. [Google Scholar] [CrossRef]
- Cordoba, A.; Hernández, R.; Viveros-Palma, I.; Mendoza, S.; Guevara-González, R.G.; Feregrino-Pérez, A.A.; Esquivel, K. Effect on plant growth parameters and secondary metabolite content of lettuce (Lactuca sativa L.), coriander (Coriandrum sativum), and chili pepper (Capsicum annuum L.) watered with disinfected water by Ag-TiO2 nanoparticles. Environ. Sci. Pollut. Res. 2021, 28, 37130–37141. [Google Scholar] [CrossRef]
- Younes, N.A.; Hassan, H.S.; Elkady, M.F.; Hamed, A.M.; Dawood, M.F.A. Impact of synthesized metal oxide nanomaterials on seedlings production of three Solanaceae crops. Heliyon 2020, 6, e03188. [Google Scholar] [CrossRef]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Zhao, L.; Wu, J.; Xiong, H.; Bao, Y.; Zeb, A.; Tang, J.; Liu, W. Foliar spray of TiO2 nanoparticles prevails over root application in reducing Cd accumulation and mitigating Cd-induced phytotoxicity in maize (Zea mays L.). Chemosphere 2020, 239, 124794. [Google Scholar] [CrossRef]
- Alidoust, D.; Isoda, A. Effect of γFe2O3 nanoparticles on photosynthetic characteristic of soybean (Glycine max (L.) Merr.): Foliar spray versus soil amendment. Acta Physiol. Plant. 2013, 35, 3365–3375. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, D.; Seo, S.M.; Kim, D. Physiological effects of zero-valent iron nanoparticles in rhizosphere on edible crop, Medicago sativa (Alfalfa), grown in soil. Ecotoxicology 2019, 28, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Rossi, L.; Stowers, C.; Zhang, W.; Lombardini, L.; Ma, X. The impact of cerium oxide nanoparticles on the physiology of soybean (Glycine max (L.) Merr.) under different soil moisture conditions. Environ. Sci. Pollut. Res. 2018, 25, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Mohammad Alabdallah, N.; Saeed Alzahrani, H. Impact of ZnO Nanoparticles on Growth of Cowpea and Okra Plants under Salt Stress Conditions. Biosci. Biotechnol. Res. Asia 2020, 17, 329–340. [Google Scholar] [CrossRef]
- Aslani, F.; Bagheri, S.; Muhd Julkapli, N.; Juraimi, A.S.; Hashemi, F.S.G.; Baghdadi, A. Effects of Engineered Nanomaterials on Plants Growth: An Overview. Sci. World J. 2014, 2014, 641759. [Google Scholar] [CrossRef]
- Salem, M.Z.M.; El-Hefny, M.; Ali, H.M.; Abdel-Megeed, A.; El-Settawy, A.A.A.; Böhm, M.; Mansour, M.M.A.; Salem, A.Z.M. Plants-derived bioactives: Novel utilization as antimicrobial, antioxidant and phytoreducing agents for the biosynthesis of metallic nanoparticles. Microb. Pathog. 2021, 158, 105107. [Google Scholar] [CrossRef]
- Wen, Y.; Liao, Y.; Tang, Y.; Zhang, H.; Zhang, J.; Liao, Z. Metabolic Effects of Elicitors on the Biosynthesis of Tropane Alkaloids in Medicinal Plants. Plants 2023, 12, 3050. [Google Scholar] [CrossRef]
- Ke, M.; Qu, Q.; Peijnenburg, W.J.G.M.; Li, X.; Zhang, M.; Zhang, Z.; Lu, T.; Pan, X.; Qian, H. Phytotoxic effects of silver nanoparticles and silver ions to Arabidopsis thaliana as revealed by analysis of molecular responses and of metabolic pathways. Sci. Total Environ. 2018, 644, 1070–1079. [Google Scholar] [CrossRef]
- Marano, F.; Hussain, S.; Rodrigues-Lima, F.; Baeza-Squiban, A.; Boland, S. Nanoparticles: Molecular targets and cell signalling. Arch. Toxicol. 2011, 85, 733–741. [Google Scholar] [CrossRef]
- Li, P.; Wang, A.; Du, W.; Mao, L.; Wei, Z.; Wang, S.; Yuan, H.; Ji, R.; Zhao, L. Insight into the interaction between Fe-based nanomaterials and maize (Zea mays) plants at metabolic level. Sci. Total Environ. 2020, 738, 139795. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, L.; Zhao, X.; Zhao, S.; Gu, X.; Du, W.; Wei, H.; Ji, R.; Zhao, L. Metabolomics Reveals the “Invisible” Responses of Spinach Plants Exposed to CeO2 Nanoparticles. Environ. Sci. Technol. 2019, 53, 6007–6017. [Google Scholar] [CrossRef]
- Singh, A.; Tiwari, S.; Pandey, J.; Lata, C.; Singh, I.K. Role of nanoparticles in crop improvement and abiotic stress management. J. Biotechnol. 2021, 337, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Ribeiro, T.P.; Santos, C.; Pinto, D.C.G.A.; Silva, A.M.S. TiO2 nanoparticles induced sugar impairments and metabolic pathway shift towards amino acid metabolism in wheat. J. Hazard. Mater. 2020, 399, 122982. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Ghorbanpour, M.; Kariman, K. Manganese oxide nanoparticle-induced changes in growth, redox reactions and elicitation of antioxidant metabolites in deadly nightshade (Atropa belladonna L.). Ind. Crops Prod. 2018, 126, 403–414. [Google Scholar] [CrossRef]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Salehi, H.; Miras-Moreno, B.; Chehregani Rad, A.; Pii, Y.; Mimmo, T.; Cesco, S.; Lucini, L. Relatively Low Dosages of CeO2 Nanoparticles in the Solid Medium Induce Adjustments in the Secondary Metabolism and Ionomic Balance of Bean (Phaseolus vulgaris L.) Roots and Leaves. J. Agric. Food Chem. 2020, 68, 67–76. [Google Scholar] [CrossRef]
- Zhang, Y.; Qi, G.; Yao, L.; Huang, L.; Wang, J.; Gao, W. Effects of Metal Nanoparticles and Other Preparative Materials in the Environment on Plants: From the Perspective of Improving Secondary Metabolites. J. Agric. Food Chem. 2022, 70, 916–933. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.; Iavicoli, I.; Calabrese, E.J. The two faces of nanomaterials: A quantification of hormesis in algae and plants. Environ. Int. 2019, 131, 105044. [Google Scholar] [CrossRef]
- Wahab, A.; Muhammad, M.; Ullah, S.; Abdi, G.; Shah, G.M.; Zaman, W.; Ayaz, A. Agriculture and environmental management through nanotechnology: Eco-friendly nanomaterial synthesis for soil-plant systems, food safety, and sustainability. Sci. Total Environ. 2024, 926, 171862. [Google Scholar] [CrossRef]
- Rai, P.K.; Kumar, V.; Lee, S.S.; Raza, N.; Kim, K.H.; Ok, Y.S.; Tsang, D.C.W. Nanoparticle-plant interaction: Implications in energy, environment, and agriculture. Environ. Int. 2018, 119, 1–19. [Google Scholar] [CrossRef]
- Mura, S.; Greppi, G.; Irudayaraj, J. Latest Developments of Nanotoxicology in Plants; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 125–151. [Google Scholar]
- Ahmed, B.; Rizvi, A.; Ali, K.; Lee, J.; Zaidi, A.; Khan, M.S.; Musarrat, J. Nanoparticles in the soil–plant system: A review. Environ. Chem. Lett. 2021, 19, 1545–1609. [Google Scholar] [CrossRef]
- Siddiqui, M.A.; Saquib, Q.; Ahamed, M.; Farshori, N.N.; Ahmad, J.; Wahab, R.; Khan, S.T.; Alhadlaq, H.A.; Musarrat, J.; Al-Khedhairy, A.A.; et al. Molybdenum nanoparticles-induced cytotoxicity, oxidative stress, G2/M arrest, and DNA damage in mouse skin fibroblast cells (L929). Colloids Surf. B Biointerfaces 2015, 125, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef]
- Iannone, M.F.; Groppa, M.D.; de Sousa, M.E.; Fernández van Raap, M.B.; Benavides, M.P. Impact of magnetite iron oxide nanoparticles on wheat (Triticum aestivum L.) development: Evaluation of oxidative damage. Environ. Exp. Bot. 2016, 131, 77–88. [Google Scholar] [CrossRef]
- García-Sánchez, S.; Bernales, I.; Cristobal, S. Early response to nanoparticles in the Arabidopsis transcriptome compromises plant defence and root-hair development through salicylic acid signalling. BMC Genom. 2015, 16, 341. [Google Scholar] [CrossRef]
- Kurczyńska, E.; Godel-Jędrychowska, K.; Sala, K.; Milewska-Hendel, A. Nanoparticles—Plant Interaction: What We Know, Where We Are? Appl. Sci. 2021, 11, 5473. [Google Scholar] [CrossRef]
- Mohammadinejad, R.; Karimi, S.; Iravani, S.; Varma, R.S. Plant-derived nanostructures: Types and applications. Green. Chem. 2015, 18, 20–52. [Google Scholar] [CrossRef]
- Kaphle, A.; Navya, P.N.; Umapathi, A.; Daima, H.K. Nanomaterials for agriculture, food and environment: Applications, toxicity and regulation. Environ. Chem. Lett. 2017, 16, 43–58. [Google Scholar] [CrossRef]
- Djibril Sekou, K.; Patel, H. A Review on the interaction between Nanoparticles and Toxic metals in Soil: Meta-analysis of their effects on soil, plants and human health. Soil. Sediment. Contam. Int. J. 2023, 32, 417–447. [Google Scholar] [CrossRef]
- Rodrigues, S.M.; Demokritou, P.; Dokoozlian, N.; Hendren, C.O.; Karn, B.; Mauter, M.S.; Sadik, O.A.; Safarpour, M.; Unrine, J.M.; Viers, J.; et al. Nanotechnology for sustainable food production: Promising opportunities and scientific challenges. Environ. Sci. Nano 2017, 4, 767–781. [Google Scholar] [CrossRef]
- El-Khawaga, A.M.; Zidan, A.; El-Mageed, A.I.A.A. Preparation methods of different nanomaterials for various potential applications: A review. J. Mol. Struct. 2023, 1281, 135148. [Google Scholar] [CrossRef]
- Pokhrel, L.R.; Dubey, B. Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 2013, 452–453, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.S.; Lenka, S.K.; Cahill, D.M.; Rookes, J.E. Designer nanoparticles for plant cell culture systems: Mechanisms of elicitation and harnessing of specialized metabolites. BioEssays 2021, 43, 2100081. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhou, J.; Wang, Y.; Wang, Y. Nano on micro: Tuning microbial metabolisms by nano-based artificial mediators to enhance and expand production of biochemicals. Curr. Opin. Biotechnol. 2020, 64, 161–168. [Google Scholar] [CrossRef]
- Kuska, J.; O’Reilly, E. Engineered biosynthetic pathways and biocatalytic cascades for sustainable synthesis. Curr. Opin. Chem. Biol. 2020, 58, 146–154. [Google Scholar] [CrossRef]
- Matur, M.; Madhyastha, H.; Shruthi, T.S.; Madhyastha, R.; Srinivas, S.P.; Navya, P.N.; Daima, H.K. Engineering bioactive surfaces on nanoparticles and their biological interactions. Sci. Rep. 2020, 10, 19713. [Google Scholar] [CrossRef]
- Zhang, D.; Li, X.; Xie, X.; Zheng, W.; Li, A.; Liu, Y.; Liu, X.; Zhang, R.; Deng, C.; Cheng, J.; et al. Exploring the Biological Effect of Biosynthesized Au–Pd Core–Shell Nanoparticles through an Untargeted Metabolomics Approach. ACS Appl. Mater. Interfaces 2021, 13, 59633–59648. [Google Scholar] [CrossRef]
- Ahmar, S.; Mahmood, T.; Fiaz, S.; Mora-Poblete, F.; Shafique, M.S.; Chattha, M.S.; Jung, K.-H. Advantage of Nanotechnology-Based Genome Editing System and Its Application in Crop Improvement. Front. Plant Sci. 2021, 12, 663849. [Google Scholar] [CrossRef]
- Lv, Z.; Jiang, R.; Chen, J.; Chen, W. Nanoparticle-mediated gene transformation strategies for plant genetic engineering. Plant J. 2020, 104, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Jat, S.K.; Bhattacharya, J.; Sharma, M.K. Nanomaterial based gene delivery: A promising method for plant genome engineering. J. Mater. Chem. B 2020, 8, 4165–4175. [Google Scholar] [CrossRef]
- Wu, K.; Xu, C.; Li, T.; Ma, H.; Gong, J.; Li, X.; Sun, X.; Hu, X. Application of Nanotechnology in Plant Genetic Engineering. Int. J. Mol. Sci. 2023, 24, 14836. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Dogan, I.; Hocaoglu-Ozyigit, A.; Yalcin, B.; Erdogan, A.; Yalcin, I.E.; Cabi, E.; Kaya, Y. Production of secondary metabolites using tissue culture-based biotechnological applications. Front. Plant Sci. 2023, 14, 1132555. [Google Scholar] [CrossRef] [PubMed]
- Borah, A.; Singh, S.; Chattopadhyay, R.; Kaur, J.; Bari, V.K. Integration of CRISPR/Cas9 with multi-omics technologies to engineer secondary metabolite productions in medicinal plant: Challenges and Prospects. Funct. Integr. Genom. 2024, 24, 207. [Google Scholar] [CrossRef] [PubMed]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, Y.; Yin, H. Recent advances in chemical modifications of guide RNA, mRNA and donor template for CRISPR-mediated genome editing. Adv. Drug Deliv. Rev. 2021, 168, 246–258. [Google Scholar] [CrossRef]
- Chen, X.; Shukal, S.; Zhang, C. Integrating Enzyme and Metabolic Engineering Tools for Enhanced α-Ionone Production. J. Agric. Food Chem. 2019, 67, 13451–13459. [Google Scholar] [CrossRef]
- Hartline, C.J.; Schmitz, A.C.; Han, Y.; Zhang, F. Dynamic control in metabolic engineering: Theories, tools, and applications. Metab. Eng. 2021, 63, 126–140. [Google Scholar] [CrossRef]
- Mukherjee, A.; Majumdar, S.; Servin, A.D.; Pagano, L.; Dhankher, O.P.; White, J.C. Carbon nanomaterials in agriculture: A critical review. Front. Plant Sci. 2016, 7, 172. [Google Scholar] [CrossRef]
- Iula, G.; Miras-Moreno, B.; Rouphael, Y.; Lucini, L.; Trevisan, M. The Complex Metabolomics Crosstalk Triggered by Four Molecular Elicitors in Tomato. Plants 2022, 11, 678. [Google Scholar] [CrossRef]
- García-Pérez, P.; Miras-Moreno, B.; Lucini, L.; Gallego, P.P. The metabolomics reveals intraspecies variability of bioactive compounds in elicited suspension cell cultures of three Bryophyllum species. Ind. Crops Prod. 2021, 163, 113322. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, J.; Li, Y.; Luo, G.; Li, L.; Yuan, H.; Mur, L.A.J.; Guo, S. Negative effects of the simulated nitrogen deposition on plant phenolic metabolism: A meta-analysis. Sci. Total Environ. 2020, 719, 137442. [Google Scholar] [CrossRef]
- Lynch, J.H.; Huang, X.Q.; Dudareva, N. Silent constraints: The hidden challenges faced in plant metabolic engineering. Curr. Opin. Biotechnol. 2021, 69, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Al-Sudani, W.K.K.; Al-Shammari, R.S.S.; Abed, M.S.; Al-Saedi, J.H.; Mernea, M.; Lungu, I.I.; Dumitrache, F.; Mihailescu, D.F. The Impact of ZnO and Fe2O3 Nanoparticles on Sunflower Seed Germination, Phenolic Content and Antiglycation Potential. Plants 2024, 13, 1724. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Desoky, E.S.M.; Babalghith, A.O.; El-Tahan, A.M.; Ibrahim, O.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; et al. Role of Nanoparticles in Enhancing Crop Tolerance to Abiotic Stress: A Comprehensive Review. Front. Plant Sci. 2022, 13, 946717. [Google Scholar] [CrossRef]
- Ahmed, A.; He, P.; He, P.; Wu, Y.; He, Y.; Munir, S. Environmental effect of agriculture-related manufactured nano-objects on soil microbial communities. Environ. Int. 2023, 173, 107819. [Google Scholar] [CrossRef]
- Dwivedi, S.; Saquib, Q.; Al-Khedhairy, A.A.; Musarrat, J. Understanding the Role of Nanomaterials in Agriculture. In Microbial Inoculants in Sustainable Agricultural Productivity: Vol. 2: Functional Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 271–288. [Google Scholar] [CrossRef]
- Avila-Quezada, G.D.; Ingle, A.P.; Golińska, P.; Rai, M. Strategic applications of nano-fertilizers for sustainable agriculture: Benefits and bottlenecks. Nanotechnol. Rev. 2022, 11, 2123–2140. [Google Scholar] [CrossRef]
- kazemi, S.; Hosseingholian, A.; Gohari, S.D.; Feirahi, F.; Moammeri, F.; Mesbahian, G.; Moghaddam, Z.S.; Ren, Q. Recent advances in green synthesized nanoparticles: From production to application. Mater. Today Sustain. 2023, 24, 100500. [Google Scholar] [CrossRef]
- Shang, Y.; Kamrul Hasan, M.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of Nanotechnology in Plant Growth and Crop Protection: A Review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef]
- Mujaddidi, N.; Nisa, S.; Al Ayoubi, S.; Bibi, Y.; Khan, S.; Sabir, M.; Zia, M.; Ahmad, S.; Qayyum, A. Pharmacological properties of biogenically synthesized silver nanoparticles using endophyte Bacillus cereus extract of Berberis lyceum against oxidative stress and pathogenic multidrug-resistant bacteria. Saudi J. Biol. Sci. 2021, 28, 6432–6440. [Google Scholar] [CrossRef] [PubMed]
- Datkhile, K.D.; Patil, S.R.; Durgawale, P.P.; Patil, M.N.; Jagdale, N.J.; Deshmukh, V.N.; More, A.L. Biogenic Silver Nanoparticles Synthesized Using Mexican Poppy Plant Inhibits Cell Growth in Cancer Cells through Activation of Intrinsic Apoptosis Pathway. Nano Biomed. Eng. 2020, 12, 241–252. [Google Scholar] [CrossRef]
- Khan, A.K.; Kousar, S.; Tungmunnithum, D.; Hano, C.; Abbasi, B.H.; Anjum, S. Nano-Elicitation as an Effective and Emerging Strategy for In Vitro Production of Industrially Important Flavonoids. Appl. Sci. 2021, 11, 1694. [Google Scholar] [CrossRef]
- Chaud, M.; Souto, E.B.; Zielinska, A.; Severino, P.; Batain, F.; Oliveira-Junior, J.; Alves, T. Nanopesticides in Agriculture: Benefits and Challenge in Agricultural Productivity, Toxicological Risks to Human Health and Environment. Toxics 2021, 9, 131. [Google Scholar] [CrossRef]
- Yin, J.; Su, X.; Yan, S.; Shen, J. Multifunctional Nanoparticles and Nanopesticides in Agricultural Application. Nanomaterials 2023, 13, 1255. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Singh, A.; Minkina, T.; Rawat, S.; Mandzhieva, S.; Sushkova, S.; Shuvaeva, V.; Nazarenko, O.; Rajput, P.; Ko-mariah; et al. Nano-Enabled Products: Challenges and Opportunities for Sustainable Agriculture. Plants 2021, 10, 2727. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Ahmed, T.; Ijaz, U.; Hameed, A.; Shahid, M.; Azizullah; Li, D.; Song, F. Microbe-oriented nanoparticles as phytomedicines for plant health management: An emerging paradigm to achieve global food security. Crit. Rev. Food Sci. Nutr. 2023, 63, 7489–7509. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.I.; Villacis-Aguirre, C.A.; López-Aguilar, K.V.; Santiago Padilla, L.; Altamirano, C.; Toledo, J.R.; Santiago Vispo, N. The Hitchhiker’s Guide to Human Therapeutic Nanoparticle Development. Pharmaceutics 2022, 14, 247. [Google Scholar] [CrossRef]
- Foulkes, R.; Man, E.; Thind, J.; Yeung, S.; Joy, A.; Hoskins, C. The regulation of nanomaterials and nanomedicines for clinical application: Current and future perspectives. Biomater. Sci. 2020, 8, 4653–4664. [Google Scholar] [CrossRef]
- Gottardo, S.; Mech, A.; Drbohlavová, J.; Małyska, A.; Bøwadt, S.; Riego Sintes, J.; Rauscher, H. Towards safe and sustainable innovation in nanotechnology: State-of-play for smart nanomaterials. NanoImpact 2021, 21, 100297. [Google Scholar] [CrossRef]
- Shandilya, N.; Marcoulaki, E.; Barruetabeña, L.; Llopis, I.R.; Noorlander, C.; Jiménez, A.S.; Oudart, Y.; Puelles, R.C.; Pérez-Fernández, M.; Falk, A.; et al. Perspective on a risk-based roadmap towards the implementation of the safe innovation approach for industry. NanoImpact 2020, 20, 100258. [Google Scholar] [CrossRef]
- Marcoulaki, E.; López de Ipiña, J.M.; Vercauteren, S.; Bouillard, J.; Himly, M.; Lynch, I.; Witters, H.; Shandilya, N.; van Duuren-Stuurman, B.; Kunz, V.; et al. Blueprint for a self-sustained European Centre for service provision in safe and sustainable innovation for nanotechnology. NanoImpact 2021, 23, 100337. [Google Scholar] [CrossRef]
- Shandilya, N.; Barreau, M.-S.; Suarez-Merino, B.; Porcari, A.; Pimponi, D.; Jensen, K.A.; Fransman, W.; Franken, R. TRAAC framework to improve regulatory acceptance and wider usability of tools and methods for safe innovation and sustainability of manufactured nanomaterials. NanoImpact 2023, 30, 100461. [Google Scholar] [CrossRef]
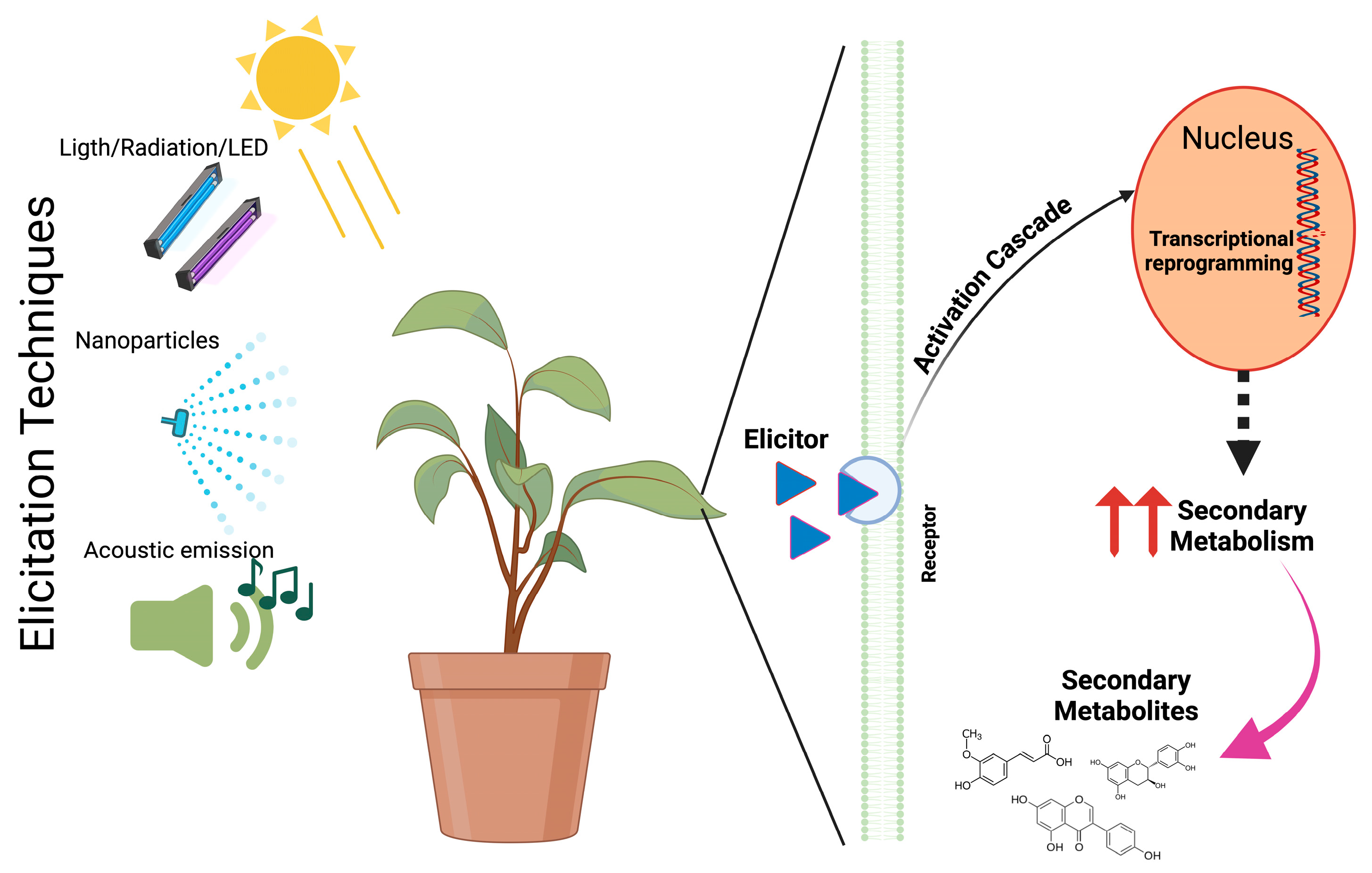
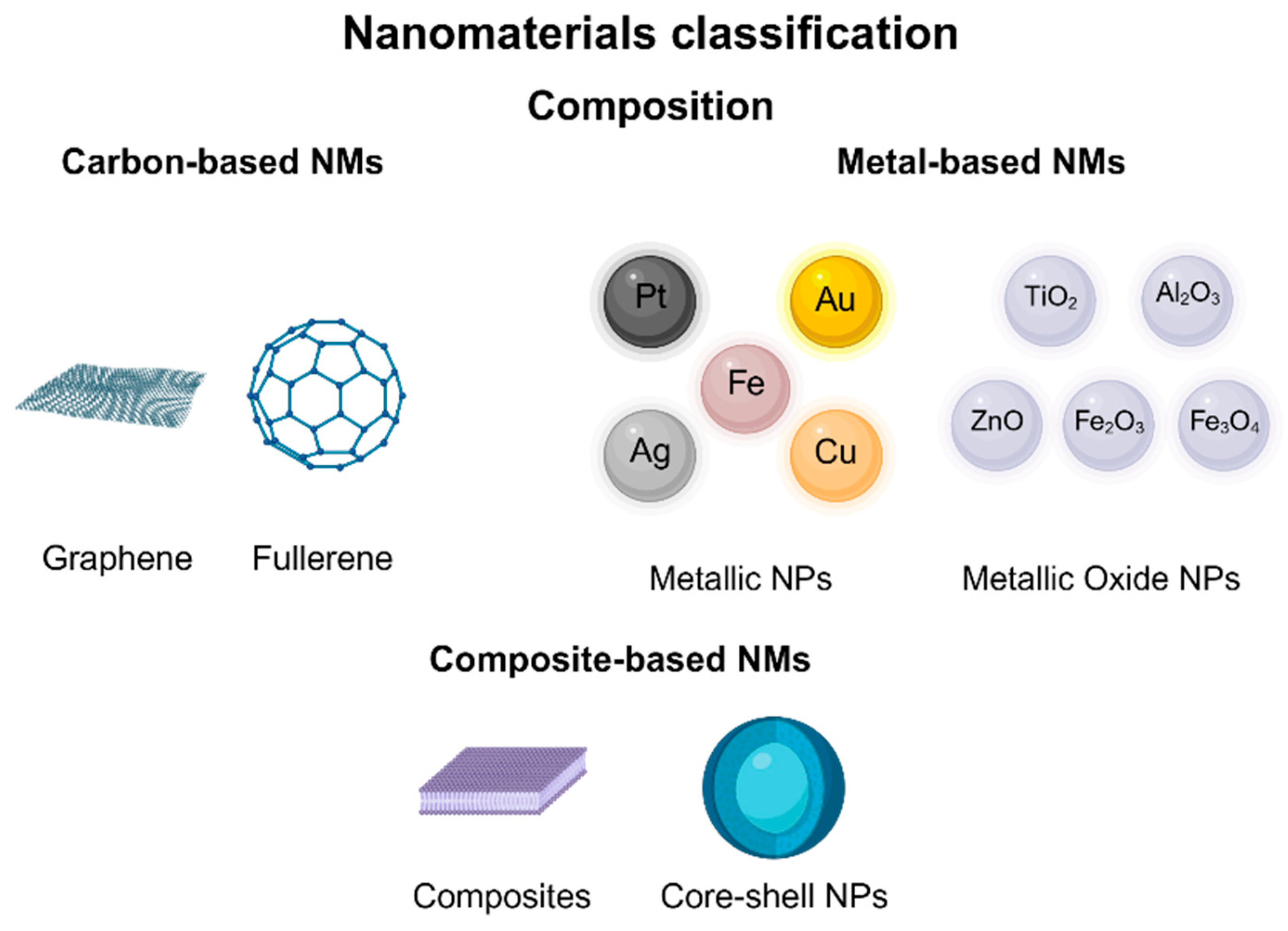
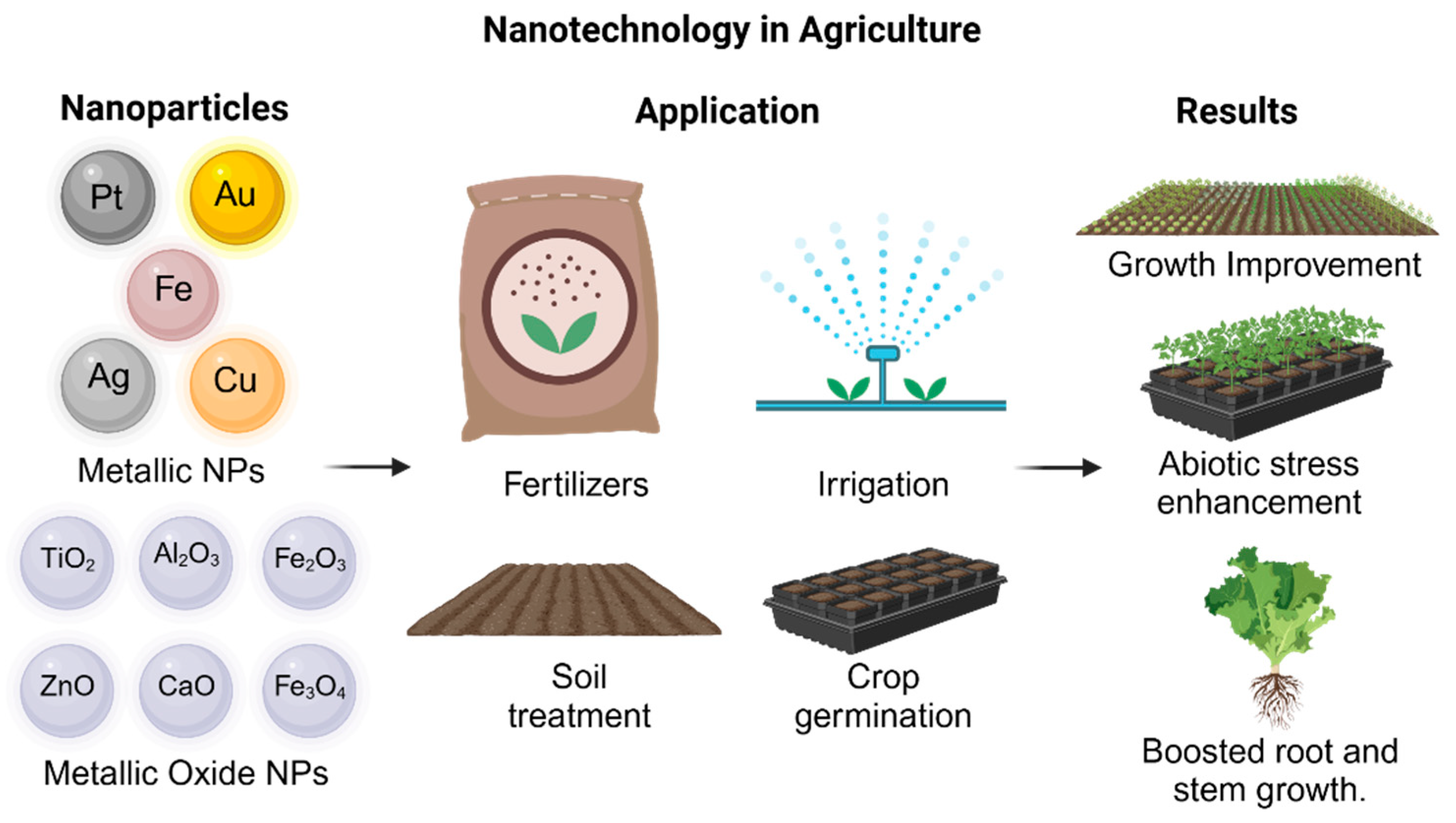
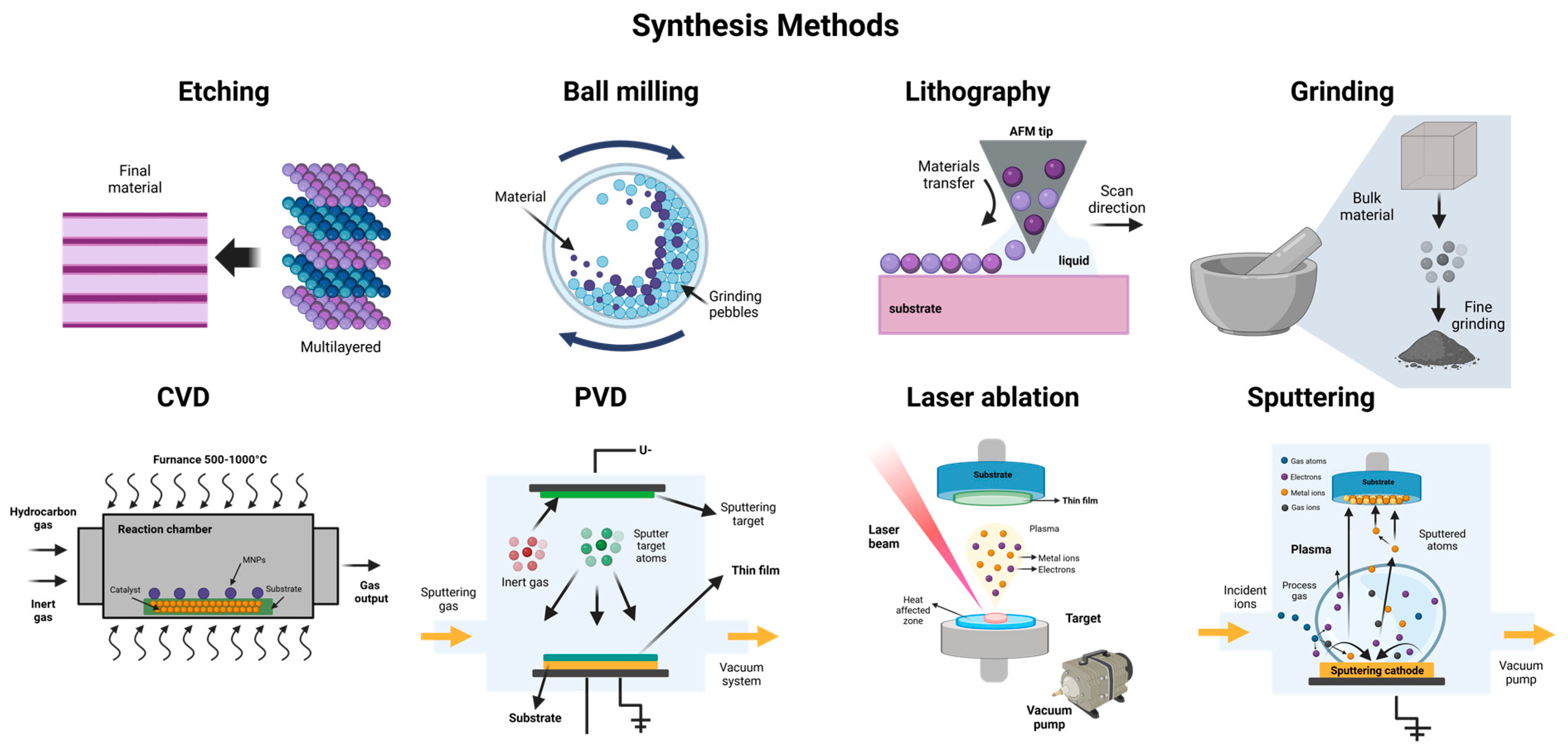
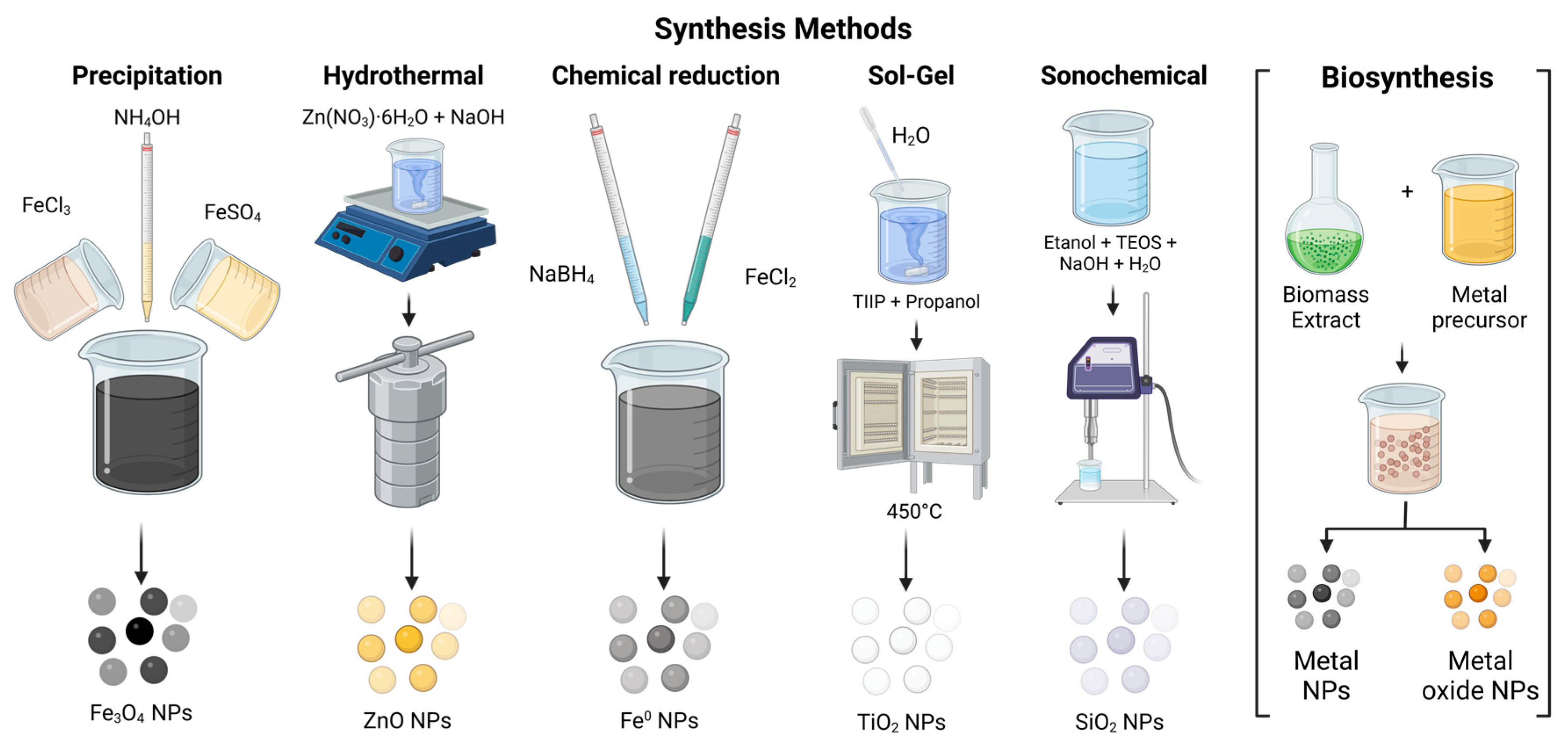
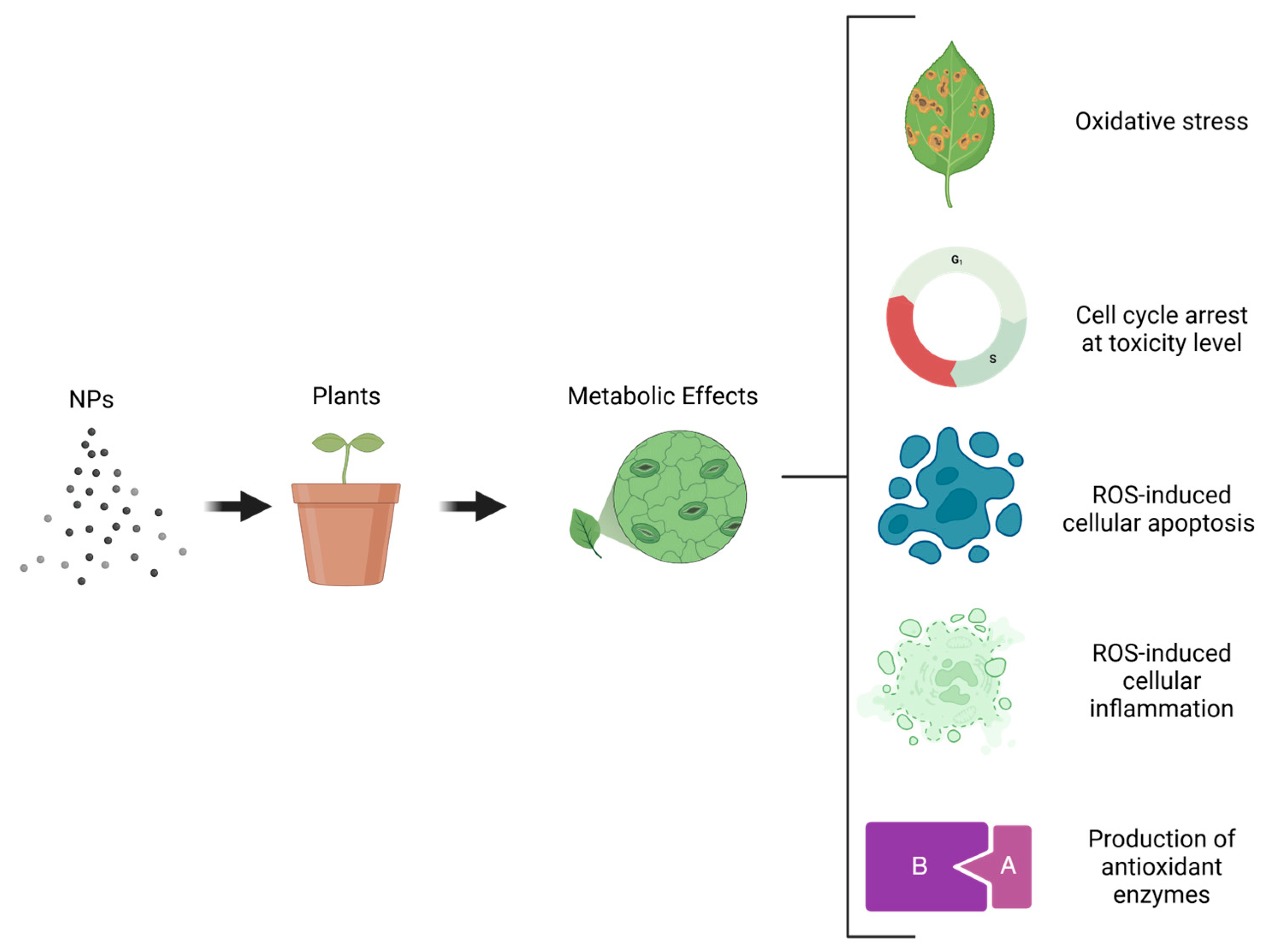
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).