Abstract
Elucidating the intricate interplay between microorganisms and crops, mainly maize (Zea mays), holds promise for devising agronomic interventions to augment yield, inducing defense mechanisms against pests and pathogens, and alleviating abiotic stresses. In pursuit of comprehending the composition of the maize root microbiomes across different genotypes, we conducted a thorough characterization of the root and rhizosphere microbiome of three indigenous Ecuadorian Andean maize cultivars, UCE-Amarillo, INIAP-122, and UCE-Pepa grown in a native soil, employing high-throughput DNA sequencing targeting the 16S rRNA gene. Root tissues and rhizosphere were collected from 18 plants from each cultivar, plus 18 bulk soil samples. Alpha diversity metrics, such as the Chao and Shannon indices, were employed to assess microbial richness and evenness, revealing a notably lower diversity within the root microbiome than the rhizosphere and bulk soil compartments. Furthermore, discernible plant genotype-driven microbiome assembly patterns were observed across all compartments. Taxonomic profiling unveiled a conspicuous absence of Actinobacteria and an elevated abundance of Proteobacteria within the root microbiome. Moreover, 34 amplicon sequence variants (ASVs) were present in all the root and rhizosphere samples (100% occupancy) from the three cultivars, making these the core microbiome of the indigenous Ecuadorian Andean maize. The highly consistent root and rhizosphere colonization across cultivars make these 34 ASVs great candidates as bioinoculants for maize in the Andean region of Ecuador. Target isolation and functional characterization of these microbes are necessary for future applications to enhance their resistance to biotic and abiotic stresses.
1. Introduction
Cereals are the basis of animal and human nutrition as they are the primary source of calories in the diet. Maize (Zea mays) is one of the most consumed cereals worldwide, producing approximately 197 M ha annually [1]. Maize also has several industrial applications for human food, animal feed, and industrial products such as alcohols (ethanol), bioplastics, adhesives, lubricating oils, rubbers, and chemical products. It is also used in industrial ingredients such as maltodextrin, dextrose, glucose syrup, and sorbitol, which are used in food, pharmaceuticals, cosmetics, and other products [2]. Furthermore, maize is consumed fresh or used as an ingredient in multiple local ancestral dishes in Latin America, especially in Mexico and the Andean region, where more than 50 maize cultivars are grown yearly [3]. Because of its importance, maize production needs to be guaranteed in the context of global climate change: reduced periods of rainfall, higher temperatures, and pest and disease increases [4].
The plant microbiome, especially the microbial communities associated with the roots, has been studied for its potential to increase yield and induce resistance and tolerance to various biotic and abiotic stresses. In recent years, studies on the maize microbiome have focused on analyzing the microbiome under conditions of heat stress [5], drought [6], the presence of heavy metals [7], agricultural management [8], and bioactive compounds [9]. The microbial community can influence stomata opening during drought, delaying wilting and potentially affecting water and nutrient retention within the rhizosphere [10]. Moreover, the root-associated microbiome (rhizobiome) aids in tolerating high concentrations of heavy metals, leading to enhanced maize yield in contaminated soils [11]. These findings suggest that the microbiome significantly affects plant physiology and growth under hostile conditions. However, these studies are focused on a reduced number of commercial cultivars in large producer countries, sidelining cultivars in more diverse regions with different soil types and climates, which is necessary to exploit the genetic potential of maize and microbial functional diversity.
This study aimed to characterize the rhizosphere and root endosphere bacterial microbiomes of three cultivars grown in the Ecuadorian Andes (Table 1). The bacterial microbiomes of these maize varieties differed in the three compartments described: bulk soil, rhizosphere, and root endosphere. The root endosphere had the lowest biodiversity. Taxonomic profiling revealed a conspicuous absence of Actinobacteria and an elevated abundance of Proteobacteria within the root microbiome. We also found consistently 34 bacterial taxa present in the rhizosphere and endosphere in the three cultivars, showing a robust bacterial core. Understanding the diversity of microorganisms and the core microbiome recruited by maize will help us to understand how specific taxa could be used in future studies to design synthetic biological communities for sustainable agriculture.

Table 1.
Phenotypic variables of three maize genotypes, under normal irrigation scheme.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
The cultivars UCE-Pepa, UCE-Amarillo, and INIAP-122 were obtained from the Faculty of Agricultural Sciences of the Universidad Central del Ecuador (Table 1). The seeds were sterilized with 2% sodium hypochlorite for 2 min. An amount of 50 g of seeds were placed in 500 mL plastic pots under greenhouse conditions and watered at field capacity every two days with a Hoagland growth solution. Eighteen plants per genotype (1 plant per pot) were grown for 15 days, and samples from each compartment (bulk soil, rhizosphere and root endosphere) were taken for bacterial microbiome analysis. The agricultural soil used as substrate was collected from the Malchinguí area (0°01′56.7″ N 78°20′47.3″ W). UCE-Pepa and UCE-Amarillo were used in this study because they demonstrated adaptation to drought with optimum yields. These genotypes were selected after six cycles of extensive breeding in different environments using the mass selection from families of half-siblings, obtaining favorable results for drought tolerance and ear height in Malchinguí, Pedro Moncayo canton in the province of Pichincha, Ecuador [12]. In Malchinguí, the annual rainfall average was less than 500 mm, making it an arid zone, allowing maize to adapt to these climatic conditions. These cultivars have genetic characteristics that enable them to maintain yields despite climatic conditions such as lack of irrigation and high temperatures [12]. On the other hand, the cultivar INIAP-122 (Table 1) was developed by the Instituto de Investigaciones Agropecuarias del Ecuador (INIAP), and it is a cultivar characterized by its earliness, small size, resistance to lodging, and good grain quality, as well as resistance to foliar diseases such as leaf blight and rust, caused by Helminthosporium sp. and Puccinia sp. and to ear rot caused by Fusarium moniliforme [13].
2.2. Bacterial Microbiome Analysis
2.2.1. Sampling and Sample Processing
Eighteen plants per genotype were used to characterize the bacterial microbiome. Per genotype, we obtained 18 samples of the root, 18 samples of the rhizosphere (soil that was attached to the root) per genotype, and 18 samples of bulk soil. Briefly, the root was separated from the leaf, shaken to remove excess soil, and washed with 10 mM MgSO4 until all soil was removed. Then, the root was frozen and freeze-dried. The rhizosphere and bulk soil were collected in 1.8 mL tubes and frozen at −20 °C until processing.
2.2.2. DNA Extraction and Sequencing
Bulk soil, rhizosphere, and root DNA were extracted with the DNeasy® Power Soil® Pro kit from Qiagen, Germany, following the manufacturer’s instructions. Sequencing was performed by BIOSEQUENCE S.A.S. (Quito, Ecuador) using the Illumina Miseq platform 2 × 250 v3 (Albany, NY, USA). The 16S gene for bacteria was amplified with primers 341F-805R. After removing primers, the resulting sequences were collapsed into amplicon sequence variants (ASVs) using the DADA2 version 1.8.1 [14]. Taxonomic assignment of each ASV was performed using the Naïve Bayes k-mer method, which was implemented using the MOTHUR package [14] and the Silva 132 database [15]. All samples were rarefied to 69,000 reads per sample before analysis. Amplicon sequencing data are available at the NCBI Sequence Read Archive (project PRJNA1175348).
2.2.3. Diversity Analysis
Statistical analysis was performed in R studio computer software (Version 3.3) [14]. Alpha diversity metrics (Shannon diversity index and Chao1 index) and the Bray–Curtis dissimilarity index were calculated using the Vegan package version 2.5–3 [16]. Beta diversity analysis (principal coordinate analysis) was based on the Bray–Curtis dissimilarity calculated from the rarefied abundance tables [14]. A stacked bar chart was made to visualize the microbial communities’ composition. All plots were created using the ggplot2 package [17]. The core bacterial microbiome for each cultivar was defined as all the ASV with a 100% occupancy (the ASV is present in all samples for each cultivar). A heatmap plotting the log counts of the common core ASV for all three varieties was created using the Pheatmap package.
3. Results
3.1. Bacterial Microbial Communities of the Bulk Soil, Rhizosphere, and Root Differ Between Maize Genotypes
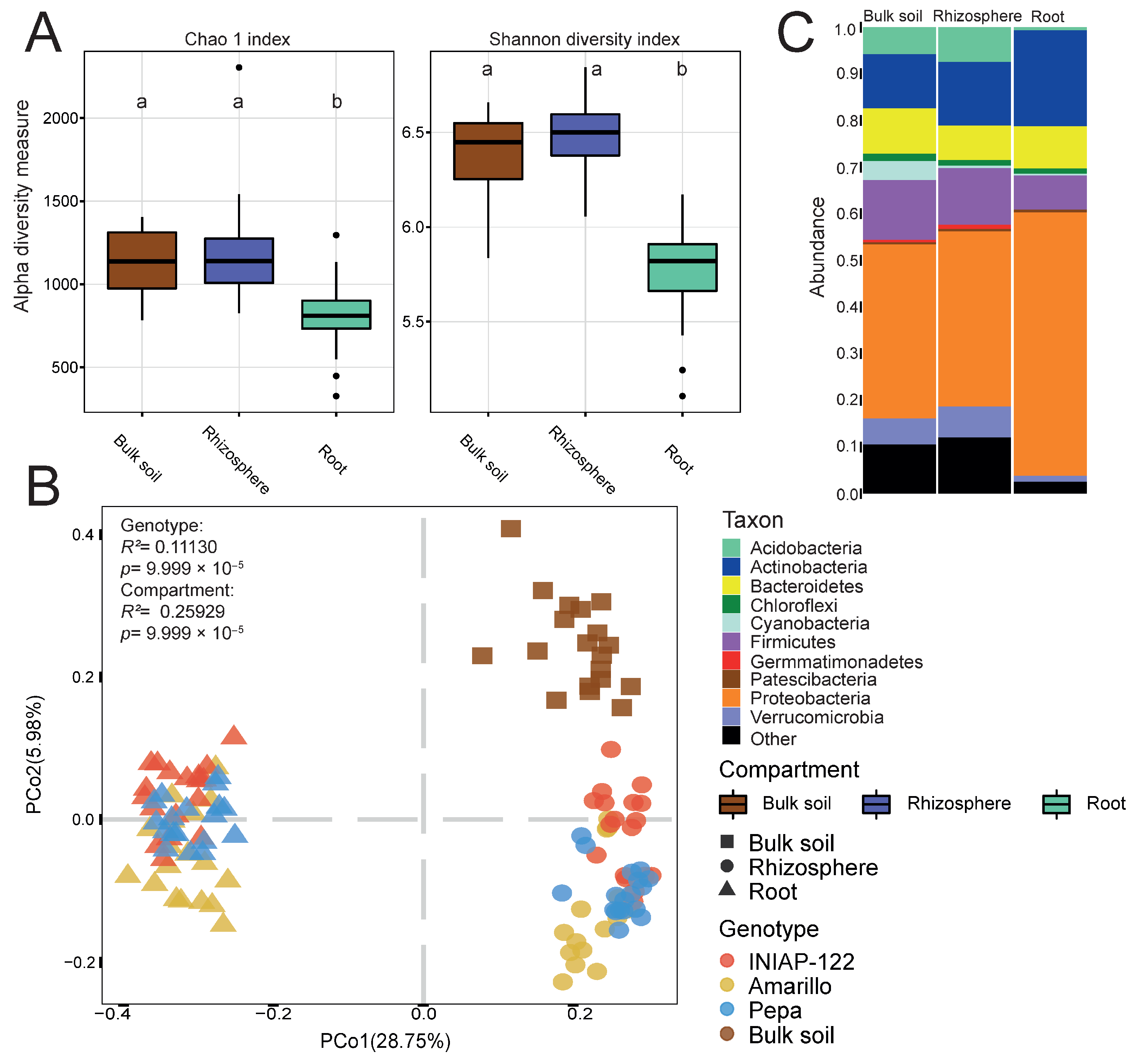
We observed differences in alpha diversity across each compartment, with the root exhibiting significantly lower diversity than the rhizosphere and the bulk soil (Figure 1A). Principal coordinate analysis (PCoA) based on dissimilarities (Figure 1B) revealed distinct assemblages between the root, rhizosphere, and bulk soil fractions. PCo1 accounted for 28.8% of the variance, distinguishing between the root and rhizosphere, while PCo2 (5.9% of the variance) represented the separation between the rhizosphere and soil (R2 = 0.26, p-value = 9.999 × 10−5). Furthermore, the plant genotype exerted a significant effect (R2 = 0.11, p-value = 9.999 × 10−5). Accordingly, with the reduced alpha diversity in the root, we observed a decreased number of phyla in the roots (Figure 1C). The communities in the root showed the absence of Acidobacteria, a higher relative abundance of Actinobacteria and Proteobacteria, and notably lower levels of Firmicutes. These findings indicate distinct bacterial microbiome compositions across the three root compartments in maize cultivars, potentially influenced by factors such as plant genetics. Additionally, these results provide insight into the previously unstudied microbiome composition of maize cultivars in this Andean soil.
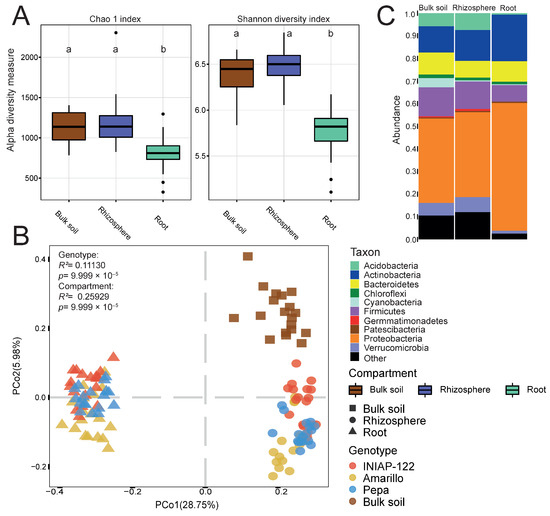
Figure 1.
Differential bacterial microbiome between compartments: root, rhizosphere, and bulk soil. (A) Alpha diversity indexes: Chao 1 and Shannon. (B) Principal coordinate analysis (PCoA) showing the effect of the compartments and plant genotype. p value determined by PERMANOVA is presented. (C) Bar graph representing the phylum relative abundance for each compartment. Different lowercase letters indicate statistically significant differences (p-value < 0.05) according to Tukey’s HSD test.
3.2. Effect of Plant Genetics on Bacterial Microbiome Assembly in Rhizosphere and Root
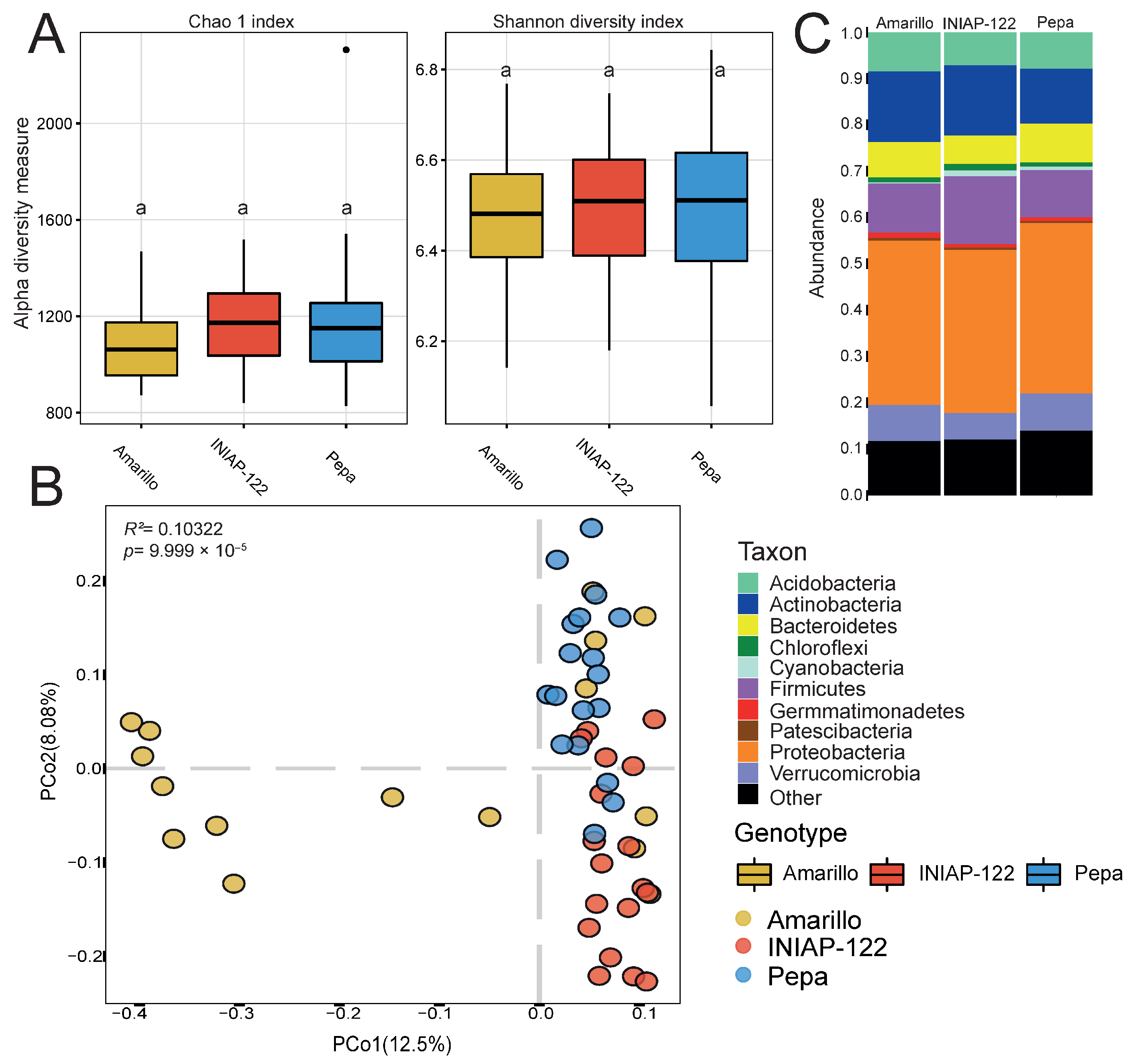
We observed that alpha diversity in the rhizosphere was not significantly different between cultivars (Figure 2A). However, in principal coordinate analysis (PCoA) based on dissimilarities (Figure 2B), we found that the bacterial communities were different and separated by genotype. The Amarillo cultivar exhibited the most significant variability among the three, while UCE-Pepa and INIAP-122 displayed more defined structures, albeit differing from each other. At the phylum level, INIAP-122 displayed a higher relative abundance of monoderm bacteria (Actinobacteria and Firmicutes) while the two landraces showed an increase in Bacteroidetes and Proteobacteria (Figure 2C). This finding underscores the influence of plant genetics on bacterial microbiome structure, highlighting a close relationship between the plant and surrounding microorganisms.
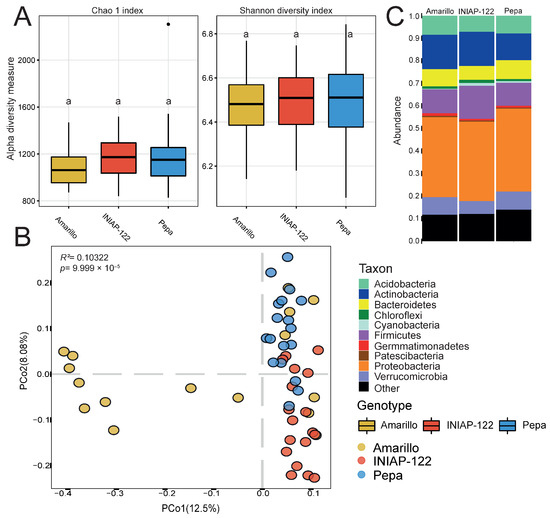
Figure 2.
Differential bacterial rhizosphere microbiome between maize cultivars. (A) Alpha diversity indexes: Chao 1 and Shannon. (B) Principal coordinate analysis (PCoA) showing the effect of the plant genotype. p value determined by PERMANOVA is presented. (C) Bar graph representing the phylum relative abundance for each cultivar. Different lowercase letters indicate statistically significant differences (p < 0.05) according to Tukey’s HSD test.
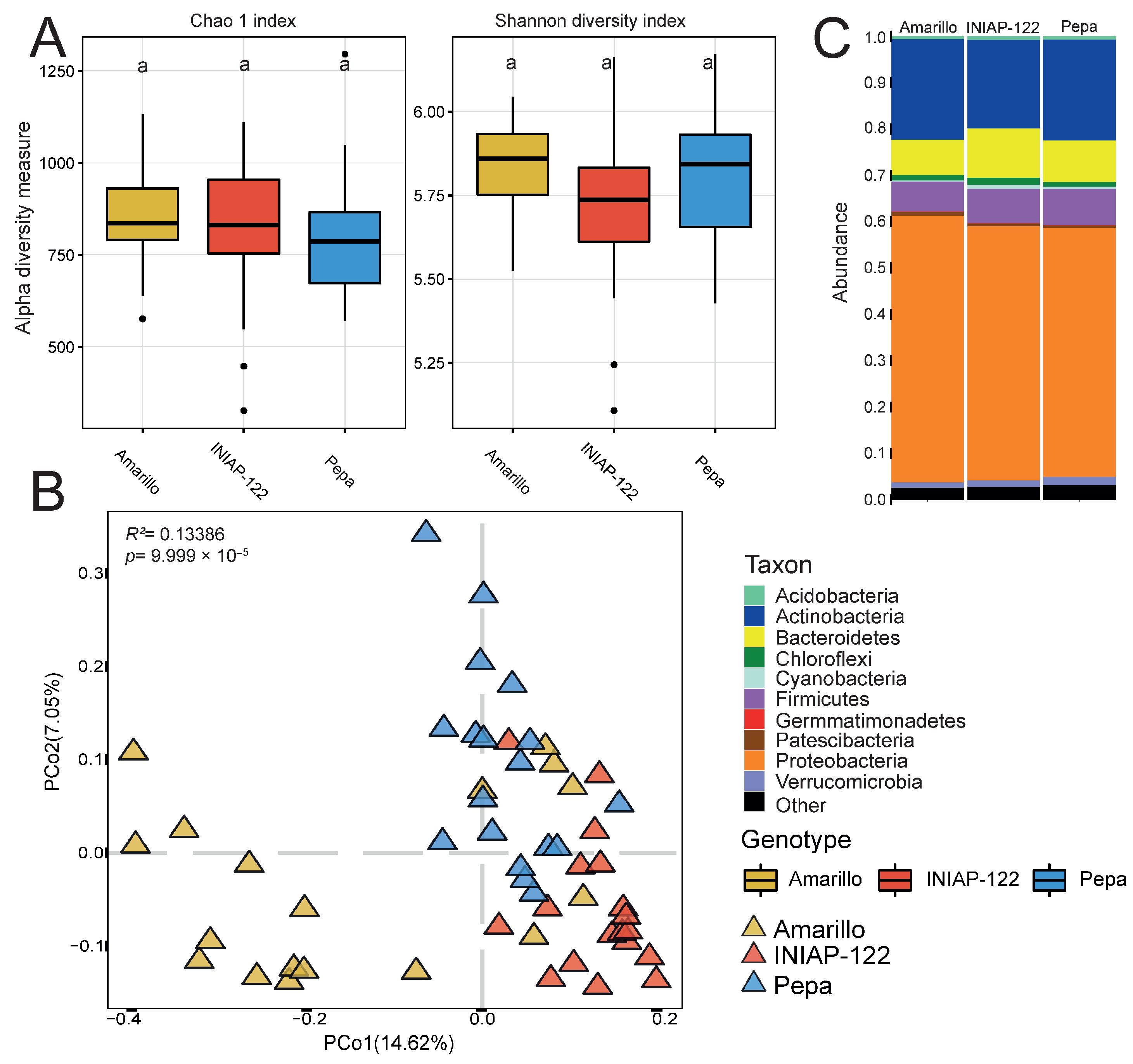
Similarly, root samples did not exhibit significant changes in alpha diversity among the three cultivars (Figure 3A). However, PCoA based on dissimilarities (Figure 3B) revealed a more similar structuring of the bacterial microbiome in UCE-Pepa and INIAP-122 compared to that of UCE-Amarillo. It is well-established that plants recruit microorganisms that provide the most benefits. In this case, all three varieties recruited the same taxa to the roots with minimal variations, suggesting a robust bacterial core microbiome.
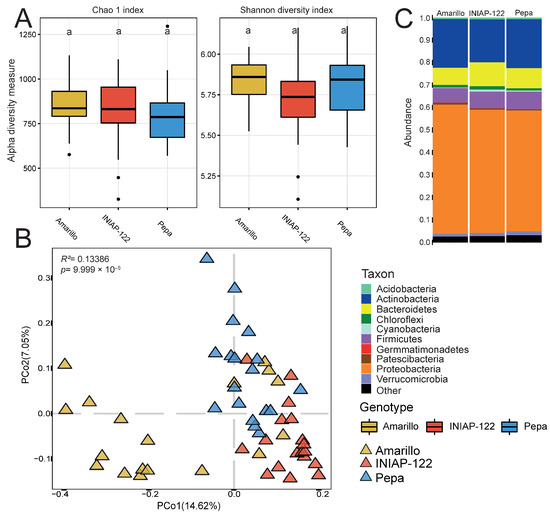
Figure 3.
Differential bacterial root microbiome between maize cultivars. (A) Alpha diversity indexes: Chao 1 and Shannon. (B) Principal coordinate analysis (PCoA) showing the effect of the plant genotype. p value determined by PERMANOVA is presented. (C) Bar graph representing the phylum relative abundance for each cultivar. Different lowercase letters indicate statistically significant differences (p < 0.05) according to Tukey’s HSD test.
3.3. The Bacterial Core Microbiome Between Andean Maize Cultivars
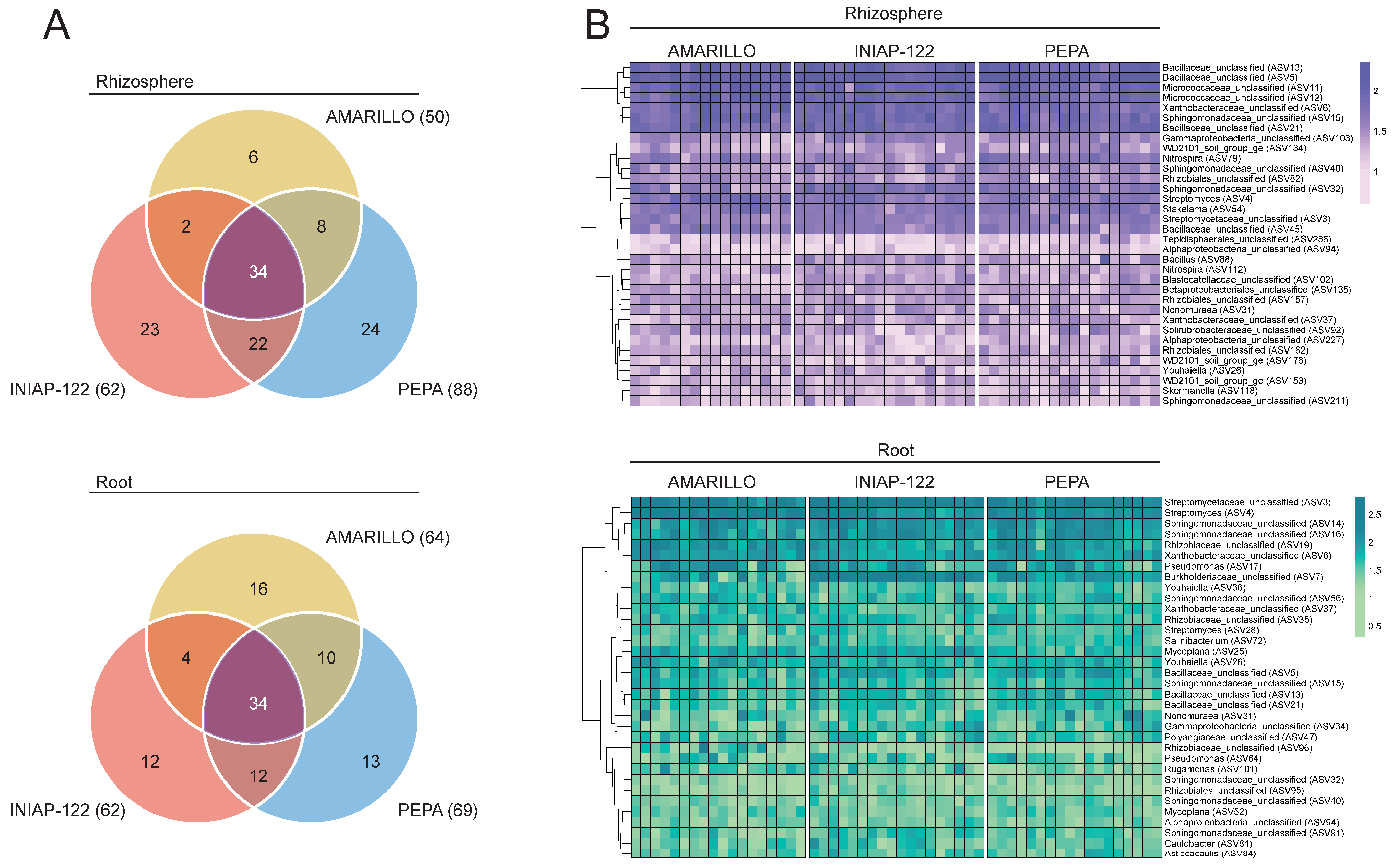
To establish the bacterial core microbiome, we identify which amplicon sequence variants (ASVs) were present in all the analyzed samples for each cultivar (100% occupancy). Every cultivar had its own core microbiome formed by 50 to 88 ASVs for the rhizosphere and 62 to 69 ASVs for the root. For the rhizosphere, INIAP-122 and UCE-Pepa shared a large number of ASVs (56 in total), and UCE-Amarillo had the lowest cultivar-specific ASVs (six). On the other hand, the shared ASVs and the cultivar-specific ASVs were more evenly distributed across cultivars in the core root microbiome. Finally, we identified 34 ASVs in the rhizosphere and 34 ASVs in the root endosphere that were common for the three cultivars (i.e., the bacterial core microbiome) (Figure 4A). Many ASVs comprising this bacterial core microbiome were classified in the Bacillaceae, Micrococcaceae, Xanthobacteraceae, and Sphingomonadaceae families (Figure 4B). The consistent abundance (counts) of the core ASVs across individual samples indicated that these microbes are robust colonizers of the rhizosphere and the root of the maize cultivars regardless of the genotype (Figure 4B).
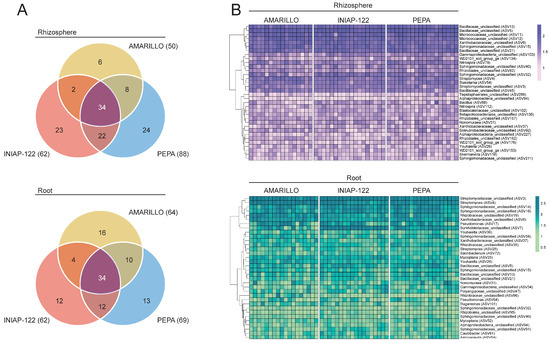
Figure 4.
Bacterial core microbiome present in Amarillo, INIAP-122, and Pepa maize. (A) The Venn diagram indicates 34 ASV present in 100% of the analyzed samples of the three cultivars. (B) Heatmap of the count (log10) of the core ASVs (rows) for each individual sample (columns) for the three cultivars and the two compartments (rhizosphere and root endosphere). Log10(count) is shown as deep purple (rhizosphere) and deep green (root).
4. Discussion
Exploring the bacterial microbiome within three Ecuadorian Andean maize cultivars, specifically adapted to endure drought conditions, has unveiled a nuanced divergence in microbial composition across distinct root compartments. This divergence is accompanied by microbial taxa previously documented as potential instigators of tolerance mechanisms within plants [5,18]. For instance, a survey of root and rhizosphere bacterial microbiome of 129 accessions of locally adapted maize in Mexico showed that the plants recruit specific bacteria from the genus Massilia to lessen nitrogen deficiency stress; even the mere inoculation with isolates from this genus was capable of inducing root and shoot growth and adaptation to low nitrogen conditions [19]. Such observations suggest a tailored recruitment strategy employed by these maize cultivars to harness specific microorganisms’ beneficial attributes in alleviating plant stress.
Using a threshold of 100% occupancy (i.e., ASVs present in all 18 samples per compartment per genotype), we found a core microbiome comprising 34 ASVs for the rhizosphere and 34 ASVs for the root endosphere. Most core ASVs were classified into the Bacillaceae, Sphingomonadaceae, Streptomycetaceae, and Rhizobiaceae families. These four families have been reported as part of the core microbiome of maize in previous studies using different genotypes in different locations (reviewed in [20]). Conversely, we did not find ASVs from the Nocardioidaceae or Paenibacillaceae as part of the core; further experiments are necessary to identify if these missing taxa are related to the genotypes or the soil used in this study. It is worth noting that, in the Andean region and worldwide, maize is commonly produced in an agroecological cereal-legume cropping system, and the bacteria shared between these two plant species enhance productivity and resilience [21]. Members of the core microbiome from the Streptomycetaceae and Rhizobiaceae families have been identified as key components in the interaction of maize with peanuts [22] or beans [23].
Additionally, a previous study that compared the microbiome changes during maize domestication found that landraces and modern hybrid genotypes have the same alpha diversity; however, they had different community compositions, and the core microbiome members were differentially abundant across the domestication gradient. Interestingly, our study found that the bacterial core ASVs were consistently recruited in the local cultivars UCE-Pepa, UCE-Amarillo, and the commercial cultivar INIAP-122, showing that these bacteria are more reliable in colonizing different genotypes, making these ASVs good candidates as bioinoculants [24]. It is important to isolate these microbes and validate their functionality, as shown in previous research that proved that core microbes could enhance resistance and tolerance to abiotic and biotic stresses in maize [25,26,27]. Going one step further, using microorganisms and synthetic communities (SynComs) from native soils, such the one used in this study, might have had higher success in improving plant health and yield [28].
The present findings provide a deeper understanding of the structural organization of the microbiome across diverse maize cultivars endemic to the Andean region. Notably, a discernible dichotomy between the microbial communities inhabiting the root system and those inhabiting the rhizosphere and surrounding soil is evident [29]. Building upon existing research that underscores the multifactorial nature of microbiome assembly, our findings reinforce that plant compartments are important modulators of bacterial microbiomes [30,31].
5. Conclusions
The bacterial root microbiome of Andean maize varieties differs significantly from the rhizosphere and lacks specific phyla that are found in the root. This discrepancy underscores how plant genetics and soil history shape microbial communities differently in each compartment. The unique microbial composition of the root, distinct from the rhizosphere, highlights the selective processes that are influenced by factors such as host genetics. We found a consistent 34 ASVs in the endosphere core microbiome of Andean maize in Ecuador. Understanding these dynamics is crucial for unraveling the complex interactions between plants and microbes and their impact on agricultural ecosystems. The adaptation of maize to the dry zone of Malchinguí might have impacted the selection and recruitment of the described soil bacteria. Consequently, the microbiome present in this soil may be vital to understanding stress tolerance using microbes in the future.
Author Contributions
A.L.-R., H.J.A.-B. and D.X.R.-V. designed the experiments. M.A.S.-G., N.B.-M., A.L.-R. and A.F.L.-G. performed the experiments. M.A.S.-G. and D.X.R.-V. analyzed the data. L.F.U.-P. and B.A.G.-P. made the DNA sequencing. M.A.S.-G. wrote the manuscript. D.X.R.-V., H.J.A.-B., S.P.-A. and A.L.-R. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by USFQ grants, Poli Grant 003, and collaboration grants. The company BIOSEQUENCE Ecuador supported it the sequencing part.
Data Availability Statement
Amplicon sequencing data are available at the NCBI Sequence Read Archive (project PRJNA1175348). This research and collections have been performed under the permit MAAE-DBI-CM-2022-0223 given by the Ministry of Environment of Ecuador.
Conflicts of Interest
Author Brigitte A. Garzón-Proaño and Luis Felipe Urresta were employed by the company BIOSEQUENCE S.A.S. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Correction Statement
This article has been republished with a minor correction to the Data Availability Statement. This change does not affect the scientific content of the article.
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global Maize Production, Consumption and Trade: Trends and R&D Implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar]
- Kaul, J.; Jain, K.; Olakh, D. An Overview on Role of Yellow Maize in Food, Feed and Nutrition Security. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 3037–3048. [Google Scholar] [CrossRef]
- Perales, H.; Golicher, D. Mapping the Diversity of Maize Races in Mexico. PLoS ONE 2014, 9, e114657. [Google Scholar] [CrossRef] [PubMed]
- Tokatlidis, I.S. Adapting Maize Crop to Climate Change. Agron. Sustain. Dev. 2013, 33, 63–79. [Google Scholar] [CrossRef]
- Tiziani, R.; Miras-Moreno, B.; Malacrinò, A.; Vescio, R.; Lucini, L.; Mimmo, T.; Cesco, S.; Sorgonà, A. Drought, Heat, and Their Combination Impact the Root Exudation Patterns and Rhizosphere Microbiome in Maize Roots. Environ. Exp. Bot. 2022, 203, 105071. [Google Scholar] [CrossRef]
- Swift, J.F.; Kolp, M.R.; Carmichael, A.; Ford, N.E.; Hansen, P.M.; Sikes, B.A.; Kleiner, M.; Wagner, M.R. Environmental Legacy Effects Impact Maize Growth and Microbiome Assembly under Drought Stress. BioRxiv 2024, 2023.04.11.536405. Available online: http://biorxiv.org/content/early/2024/01/08/2023.04.11.536405.abstract (accessed on 27 August 2024).
- AL-Huqail, A.A.; El-Bondkly, A.M.A. Improvement of Zea mays L. Growth Parameters under Chromium and Arsenic Stress by the Heavy Metal-Resistant Streptomyces sp. NRC21696. Int. J. Environ. Sci. Technol. 2022, 19, 5301–5522. [Google Scholar] [CrossRef]
- Pooniya, V.; Zhiipao, R.R.; Biswakarma, N.; Kumar, D.; Shivay, Y.S.; Babu, S.; Das, K.; Choudhary, A.K.; Swarnalakshmi, K.; Choudhary, R.L.; et al. Conservation Agriculture Based Integrated Crop Management Sustains Productivity and Economic Profitability along with Soil Properties of the Maize-Wheat Rotation. Sci. Rep. 2022, 12, 1962. [Google Scholar] [CrossRef]
- Murphy, K.M.; Edwards, J.; Louie, K.B.; Bowen, B.P.; Sundaresan, V.; Northen, T.R.; Zerbe, P. Bioactive Diterpenoids Impact the Composition of the Root-Associated Microbiome in Maize (Zea mays). Sci. Rep. 2021, 11, 333. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Fan, W.; Deng, J.; Shao, L.; Jiang, S.; Xiao, T.; Sun, W.; Xiao, E. The Rhizosphere Microbiome Improves the Adaptive Capabilities of Plants under High Soil Cadmium Conditions. Front. Plant Sci. 2022, 13, 914103. [Google Scholar] [CrossRef] [PubMed]
- Coral Valenzuela, J.V.; Andrade Bolaños, H.J.; Pumisacho Gualoto, M.M.; Caicedo Chávez, J.D.; Salazar Vizuete, D.R. Caracterización Morfológica y Agronómica de dos Genotipos de Maíz (Zea mays L.) en la Zona Media de la Parroquia Malchinguí. ACI Av. En Cienc. E Ing. 2019, 11, 21–33. [Google Scholar] [CrossRef]
- Silva, E.; Dobronsky, J.; Heredia, J. INIAP-122 Chaucho Mejorado: Variedad de Maíz Amarillo Harinoso Semi-Precoz para la Provincia de Imbabura; INIAP: Quito, Ecuador, 1997. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Copenhagen, Denmark, 2018. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. 2012. Available online: https://CRAN.R-project.org/package=vegan (accessed on 30 August 2024).
- Xu, S.; Zhan, L.; Tang, W.; Wang, Q.; Dai, Z.; Zhou, L.; Feng, T.; Chen, M.; Wu, T.; Hu, E.; et al. MicrobiotaProcess: A Comprehensive R Package for Deep Mining Microbiome. Innovation 2023, 4, 100388. [Google Scholar] [CrossRef]
- Walters, D.R.; Scholes, J.D.; Goss, M.J.; Brown, J.F.; Bevan, M.W. Exploring the Maize Microbiome for Novel Insights into Plant-Microbe Interactions and Drought Tolerance. Plant Cell Environ. 2018, 41, 2877–2890. [Google Scholar]
- He, X.; Wang, D.; Jiang, Y.; Li, M.; Delgado-Baquerizo, M.; McLaughlin, C.; Marcon, C.; Guo, L.; Baer, M.; Moya, Y.A.T.; et al. Heritable microbiome variation is correlated with source environment in locally adapted maize varieties. Nat. Plants 2024, 10, 598–617. [Google Scholar] [CrossRef]
- Singh, R.; Goodwin, S.B. Exploring the Corn Microbiome: A Detailed Review on Current Knowledge, Techniques, and Future Directions. PhytoFrontiers™ 2022, 2, 158–175. [Google Scholar] [CrossRef]
- Jalloh, A.A.; Khamis, F.M.; Yusuf, A.A.; Subramanian, S.; Mutyambai, D.M. Long-term push–pull cropping system shifts soil and maize-root microbiome diversity paving way to resilient farming system. BMC Microbiol. 2024, 24, 92. [Google Scholar] [CrossRef]
- Wang, N.; Wang, T.; Chen, Y.; Wang, M.; Lu, Q.; Wang, K.; Dou, Z.; Chi, Z.; Qiu, W.; Dai, J.; et al. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. Nat. Commun. 2024, 15, 839. [Google Scholar] [CrossRef]
- Lanzavecchia, G.; Frascarelli, G.; Rocchetti, L.; Bellucci, E.; Bitocchi, E.; Di Vittori, V.; Sillo, F.; Ferraris, I.; Carta, G.; Delledonne, M.; et al. Genotype combinations drive variability in the microbiome configuration of the rhizosphere of maize/bean intercropping system. Int. J. Mol. Sci. 2024, 25, 1288. [Google Scholar] [CrossRef]
- Poppeliers, S.W.; Sánchez-Gil, J.J.; de Jonge, R. Microbes to Support Plant Health: Understanding Bioinoculant Success in Complex Conditions. Curr. Opin. Microbiol. 2023, 73, 102286. [Google Scholar] [CrossRef] [PubMed]
- Beirinckx, S.; Viaene, T.; Haegeman, A.; Debode, J.; Amery, F.; Vandenabeele, S.; Nelissen, H.; Inzé, D.; Tito, R.; Raes, J.; et al. Tapping into the Maize Root Microbiome to Identify Bacteria That Promote Growth under Chilling Conditions. Microbiome 2020, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wei, Q.; Wu, H.; Chen, X.; Xiao, C.; Ye, Y.; Liu, C.; Yu, H.; Guo, Y.; Sun, W.; et al. Bacillus species are core microbiota of resistant maize cultivars that induce host metabolic defense against corn stalk rot. Microbiome 2024, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, M.; Huang, S.; Li, L.; Gao, Q.; Wang, Y.; Zhang, S.; Huang, S.; Yuan, L.; Wen, Y.; et al. A highly conserved core bacterial microbiota with nitrogen-fixation capacity inhabits the xylem sap in maize plants. Nat. Commun. 2022, 13, 3361. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, D.; Li, F.; Dong, Y.; Jin, Z.; Liao, Y.; Li, X.; Peng, S.; Delgado-Baquerizo, M.; Li, X. Superiority of native soil core microbiomes in supporting plant growth. Nat. Commun. 2024, 15, 6599. [Google Scholar] [CrossRef]
- Tkacz, A.; Cheema, J.; Chandra, G.; Grant, A.; Poole, P.S. Stability and Succession of the Rhizosphere Microbiota Depends Upon Plant Type and Soil Composition. ISME J. 2015, 9, 2349–2359. [Google Scholar] [CrossRef]
- Leopold, D.R.; Busby, P.E. Host Genotype and Colonist Arrival Order Jointly Govern Plant Microbiome Composition and Function. Curr. Biol. 2020, 30, 3260–3266.e5. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere Microbiome Assemblage Is Affected by Plant Development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef]


















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).