
Transcriptome Analysis of Potential Regulatory Genes under Chemical Doubling in Maize Haploids


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Doubling Methods
2.2. Construction of RNA-Seq Libraries and Transcriptome Sequencing
2.3. Read Processing and Identification of Differentially Expressed Genes
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Validation
2.5. Statistical Analysis
3. Results
3.1. Pollen Dispersal Efficiency of Maize Haploids after Colchicine Treatment
3.2. Quality Analysis of Maize Haploid Transcriptomes Sequencing
3.3. Identification of DEGs in B73 and ZNC442 Inoculated with Colchicine at Different Time Points
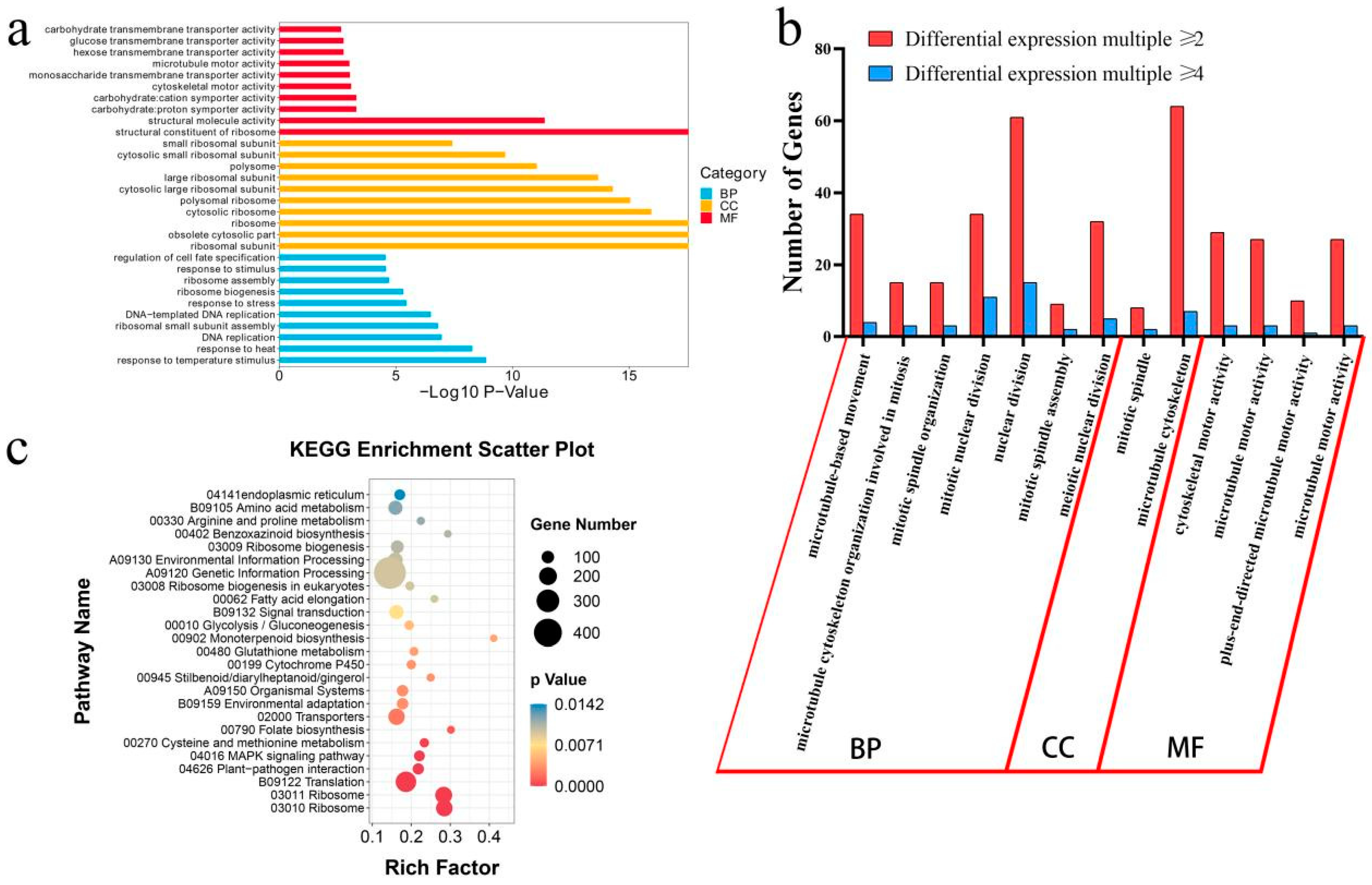
3.4. GO Enrichment Analysis of the Common DEGs in Different Treatment with Colchicine
3.5. KEGG Enrichment Analysis of the Common DEGs in Different Treatment with Colchicine
3.6. Comparative Analysis of Potential Double-Related Genes with GO and KEGG
3.7. qRT-PCR Confirms Gene Expression Profiles
4. Discussion
4.1. Haploid Doubling Methods and Efficiency
4.2. DEGs Enrichment Analysis by RNA-Seq
4.3. Candidate Genes Identified by Chemical Treatment for Chromosome Doubling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, W.R. An overview of the ways and roles of maize planting technology promotion. Mod. Agric. 2018, 10, 25–26. (In Chinese) [Google Scholar]
- Qin, X.; Feng, F.; Li, Y.; Xu, S.; Siddique, K.H.M.; Liao, Y.; Lübberstedt, T. Maize yield improvements in China: Past trends and future directions. Plant Breed. 2016, 135, 166–176. [Google Scholar] [CrossRef]
- Liu, Y.C.; Sun, Q.J. Progress in deep processing technology of corn husk, an industrial corn by-product. Agric. Prod. Process. 2017, 3, 72–75. (In Chinese) [Google Scholar] [CrossRef]
- Hui, G.Q.; Du, H.W.; Yang, S.H.; Liu, G.H.; Wang, Z.T.; Zhang, Y.R.; Zheng, Y.P.; Yan, J.B.; Zhang, M.T.; Li, J.S. Efficiency of doubling maize haploids by different herbicides. J. Crops 2012, 38, 416–422. (In Chinese) [Google Scholar] [CrossRef]
- Sauer, G.; Korner, R.; Hanisch, A.; Ries, A.; Nigg, E.A.; Sillje, H.H.W. Proteome Analysis of the Human Mitotic Spindle. Mol. Cell. Proteom. 2005, 4, 35–43. [Google Scholar] [CrossRef]
- Walczak, C.E.; Cai, S.; Khodjakov, A. Mechanisms of chromosome behaviour during mitosis. Nat. Rev. Mol. Cell Biol. 2010, 11, 91–102. [Google Scholar] [CrossRef]
- Wood, K.W.; Sakowicz, R.; Goldstein, L.S.; Cleveland, D.W. CENP-E is a plus end–directed kinetochore motor required for metaphase chromosome alignment. Cell 1997, 91, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; O'connell, C.B.; Khodjakov, A.; Walczak, C.E. Chromosome congression in the absence of kinetochore fibres. Nat. Cell Biol. 2009, 11, 832–838. [Google Scholar] [CrossRef]
- Röber, F.; Gordillo, G.; Geiger, H. In vivo haploid induction in maize. Performance of new inducers and significance of doubled haploid lines in hybrid breeding [Zea mays L.]. Maydica 2005, 50, 275–283. [Google Scholar]
- Sugihara, N.; Higashigawa, T.; Aramoto, D.; Kato, A. Haploid plants carrying a sodium azide-induced mutation (fdr1) produce fertile pollen grains due to first division restitution (FDR) in maize (Zea mays L.). Theor. Appl. Genet. 2013, 126, 2931–2941. [Google Scholar] [CrossRef]
- Yang, J.W. Localisation and Cytological Mechanism of Natural Male Flower Doubling in Haploid Maize and QTL Analysis of Ear Traits. Ph.D. thesis, Henan Agricultural University, Zhengzhou, China, 2020. (In Chinese). [Google Scholar] [CrossRef]
- Ren, J.; Wu, P.; Tian, X.; Lübberstedt, T.; Chen, S. QTL mapping for haploid male fertility by a segregation distortion method and fine mapping of a key QTL qhmf4 in maize. Theor. Appl. Genet. 2017, 130, 1349–1359. [Google Scholar] [CrossRef]
- Wang, J.; Liu, D.; Guo, X.; Yang, W.; Wang, X.; Zhan, K.; Zhang, A. Variability of gene expression after polyhaploidization in wheat (Triticum aestivum L.). G3 Genes Genomes Genet. 2011, 1, 27–33. [Google Scholar] [CrossRef]
- Wang, S.; Wang, K.S.; Liu, Z.S.; Zhang, B.D.; Li, D.X. Analysis of the dynamics of maize inbred line selection in the United States. Foreign Agron. Misc. Crops 1994, 6, 10–13. (In Chinese) [Google Scholar]
- Peng, Y.C.; Ma, X.M. Fitness analysis of major quantitative traits in several commonly used maize inbred lines. Misc. Crops 2001, 4, 1–4. (In Chinese) [Google Scholar]
- Lan, H.; Xiang, Y.; Li, L.J.; Wei, R.J.; Xia, C. Selection and promotion of the new maize variety Chuandan 99. Maize Sci. 2023, 31, 25–29. (In Chinese) [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef]
- Tadepalli, S.; Stewart, G.C.; Nagaraja, T.; Narayanan, S.K. Leukotoxin operon and differential expressions of the leukotoxin gene in bovine Fusobacterium necrophorum subspecies. Anaerobe 2008, 14, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Vanstraelen, M.; Inzé, D.; Geelen, D. Mitosis-specific kinesins in Arabidopsis. Trends Plant Sci. 2006, 11, 67–175. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, H.C.; Xu, S.W.; Chen, S.J. Progress of haploid doubling technology in maize. Maize Sci. 2010, 18, 12–14+19. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, L.; Ci, J.B.; Cui, X.Y.; Zhang, Y.; Nan, N.; Li, J.Z.; Yang, W.G. Studies on haploid induction and doubling rate of maize under different ecological conditions. J. Jilin Agric. Univ. 2014, 36, 139–143. (In Chinese) [Google Scholar] [CrossRef]
- Liu, X.X.; Yang, W.; Ci, J.B.; Cui, X.Y.; Zhang, Y.; Yang, W.G. Effects of different base materials and colchicine concentrations on doubling rate of maize haploids. J. Jilin Agric. Univ. 2015, 37, 136–140. (In Chinese) [Google Scholar] [CrossRef]
- Deng, K.P.; Li, X.Y.; Jing, G.X.; Lu, B.; Xing, Z.; Zhao, R.G. Research on efficient haploid doubling method in glutinous maize. Seeds 2017, 36, 82–85. (In Chinese) [Google Scholar] [CrossRef]
- Wen, K.; Li, L.; Liu, Y.Q.; Chen, S.J. A preliminary report on efficient biological induction of maize haploids and its doubling method. J. China Agric. Univ. 2006, 5, 17–20. (In Chinese) [Google Scholar]
- Tao, R.; Hou, J.X.; Liu, H.X.; Sang, J.; Wang, X.Q.; Yue, Y.H.; Sun, C.B. Haploid induction and doubling in Glutinous maize. Maize Sci. 2020, 28, 27–31. (In Chinese) [Google Scholar] [CrossRef]
- Qi, Z.Y.; Yang, H.; Qiu, Z.G.; Zhang, Y.Q.; Li, S.J.; Yuan, L.; Jin, C. Study on the effect of different treatments on doubling efficiency of maize haploids. South. Agric. 2013, 7, 61–63+74. (In Chinese) [Google Scholar] [CrossRef]
- Song, G.L. Research on Efficient Hybrid Induction and Doubling Method of Maize Haploid. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2011. (In Chinese). [Google Scholar]
- Gibbons, I. Dynein ATPases as microtubule motors. J. Biol. Chem. 1988, 263, 15837–15840. [Google Scholar] [CrossRef] [PubMed]
- Pfarr, C.; Coue, M.; Grissom, P.; Hays, T.; Porter, M.; McIntosh, J. Cytoplasmic dynein is localized to kinetochores during mitosis. Nature 1990, 345, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Steuer, E.; Wordeman, L.; Schroer, T.; Sheetz, M. Localization of cytoplasmic dynein to mitotic spindles and kinetochores. Nature 1990, 345, 266–268. [Google Scholar] [CrossRef]
- Hyman, A.A.; Mitchison, T.J. Two different microtubule-based motor activities with opposite polarities in kinetochores. Nature 1991, 351, 206–211. [Google Scholar] [CrossRef]
- Vallee, R. Movement on two fronts. Nature 1991, 351, 187–188. [Google Scholar] [CrossRef]
- Vaisberg, E.A.; Koonce, M.P.; McIntosh, J.R. Cytoplasmic dynein plays a role in mammalian mitotic spindle formation. J. Cell Biol. 1993, 123, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Heald, R.; Tournebize, R.; Blank, T.; Sandaltzopoulos, R.; Becker, P.; Hyman, A.; Karsenti, E. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nature 1996, 382, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.A.; McAinsh, A. Prime movers: The mechanochemistry of mitotic kinesins. Nat. Rev. Mol. Cell Biol. 2014, 15, 257–271. [Google Scholar] [CrossRef]
- Ali, I.; Yang, W.-C. The functions of kinesin and kinesin-related proteins in eukaryotes. Cell Adhes. Migr. 2020, 14, 139–152. [Google Scholar] [CrossRef]
- Pandey, H.; Popov, M.; Goldstein-Levitin, A.; Gheber, L. Mechanisms by which kinesin-5 motors perform their multiple intracellular functions. Int. J. Mol. Sci. 2021, 22, 6420. [Google Scholar] [CrossRef]
- Mazumdar, M.; Misteli, T. Chromokinesins: Multitalented players in mitosis. Trends Cell Biol. 2005, 15, 349–355. [Google Scholar] [CrossRef]
- Walczak, C.E.; Heald, R. Mechanisms of mitotic spindle assembly and function. Int. Rev. Cytol. 2008, 265, 111–158. [Google Scholar] [CrossRef]
- Tokai, N.; Fujimoto-Nishiyama, A.; Toyoshima, Y.; Yonemura, S.; Tsukita, S.; Inoue, J.; Yamamota, T. Kid, a novel kinesin-like DNA binding protein, is localized to chromosomes and the mitotic spindle. EMBO J. 1996, 15, 457–467. [Google Scholar] [CrossRef]
- Zhong, A.; Tan, F.-Q.; Yang, W.-X. Chromokinesin: Kinesin superfamily regulating cell division through chromosome and spindle. Gene 2016, 589, 43–48. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Reads No. | Bases (bp) | CleanData (bp) | Q30 (bp) | Q30 (%) | N (%) |
|---|---|---|---|---|---|---|
| Z_Y_6.2_1 | 50,317,580 | 7,597,954,580 | 7,232,153,758 | 7,159,455,508 | 94.22 | 0.000727 |
| Z_Y_6.2_2 | 53,547,544 | 8,085,679,144 | 7,700,032,996 | 7,577,260,127 | 93.71 | 0.00071 |
| Z_Y_6.2_3 | 52,561,604 | 7,936,802,204 | 7,554,463,560 | 7,452,697,706 | 93.9 | 0.000719 |
| Z_Y_10_1 | 50,164,896 | 7,574,899,296 | 7,207,362,276 | 7,162,091,078 | 94.55 | 0.000724 |
| Z_Y_10_2 | 47,767,530 | 7,212,897,030 | 6,858,606,032 | 6,781,139,650 | 94.01 | 0.000716 |
| Z_Y_10_3 | 53,007,968 | 8,004,203,168 | 7,615,212,068 | 7,570,813,834 | 94.58 | 0.00072 |
| Z_Y_ck_1 | 48,438,328 | 7,314,187,528 | 6,959,239,982 | 6,925,222,508 | 94.68 | 0.000724 |
| Z_Y_ck_2 | 44,036,844 | 6,649,563,444 | 6,326,827,520 | 6,251,640,160 | 94.01 | 0.000724 |
| Z_Y_ck_3 | 49,099,690 | 7,414,053,190 | 7,066,275,124 | 7,019,775,977 | 94.68 | 0.000726 |
| B_Y_6.2_1 | 56,845,106 | 8,583,611,006 | 8,167,464,972 | 8,087,517,555 | 94.22 | 0.00072 |
| B_Y_6.2_2 | 53,973,586 | 8,150,011,486 | 7,761,899,810 | 7,678,790,837 | 94.21 | 0.000719 |
| B_Y_6.2_3 | 57,158,714 | 8,630,965,814 | 8,188,484,474 | 8,160,629,443 | 94.55 | 0.000726 |
| B_Y_10_1 | 56,008,830 | 8,457,333,330 | 8,051,888,666 | 7,973,175,410 | 94.27 | 0.000723 |
| B_Y_10_2 | 51,722,322 | 7,810,070,622 | 7,416,724,078 | 7,327,715,065 | 93.82 | 0.000721 |
| B_Y_10_3 | 54,808,624 | 8,276,102,224 | 7,860,312,852 | 7,777,963,949 | 93.98 | 0.000726 |
| B_Y_ck_1 | 50,640,186 | 7,646,668,086 | 7,280,916,188 | 7,196,531,116 | 94.11 | 0.000724 |
| B_Y_ck_2 | 48,312,288 | 7,295,155,488 | 6,946,188,448 | 6,879,618,096 | 94.3 | 0.000717 |
| B_Y_ck_3 | 45,038,272 | 6,800,779,072 | 6,473,769,546 | 6,412,670,907 | 94.29 | 0.000731 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhan, P.; Pu, R.; Xiang, W.; Meng, X.; Yang, S.; Hu, G.; Zhao, S.; Han, J.; Xia, C.; et al. Transcriptome Analysis of Potential Regulatory Genes under Chemical Doubling in Maize Haploids. Agronomy 2024, 14, 624. https://doi.org/10.3390/agronomy14030624
Li Y, Zhan P, Pu R, Xiang W, Meng X, Yang S, Hu G, Zhao S, Han J, Xia C, et al. Transcriptome Analysis of Potential Regulatory Genes under Chemical Doubling in Maize Haploids. Agronomy. 2024; 14(3):624. https://doi.org/10.3390/agronomy14030624
Chicago/Turabian StyleLi, Youqiang, Penglin Zhan, Rumin Pu, Wenqi Xiang, Xin Meng, Shiqi Yang, Gaojiao Hu, Shuang Zhao, Jialong Han, Chao Xia, and et al. 2024. "Transcriptome Analysis of Potential Regulatory Genes under Chemical Doubling in Maize Haploids" Agronomy 14, no. 3: 624. https://doi.org/10.3390/agronomy14030624