
Transcriptomics and Metabolomics Analysis Revealed the Ability of Microbacterium ginsengiterrae S4 to Enhance the Saline-Alkali Tolerance of Rice (Oryza sativa L.) Seedlings
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Determination of Physicochemical Indicators
2.3. RNA Extraction, cDNA Library Preparation, and Sequencing
2.4. Metabolite Extraction and UPLC-MS/MS Analysis
2.5. Bioinformatics Analysis of the Transcriptome and Metabolome
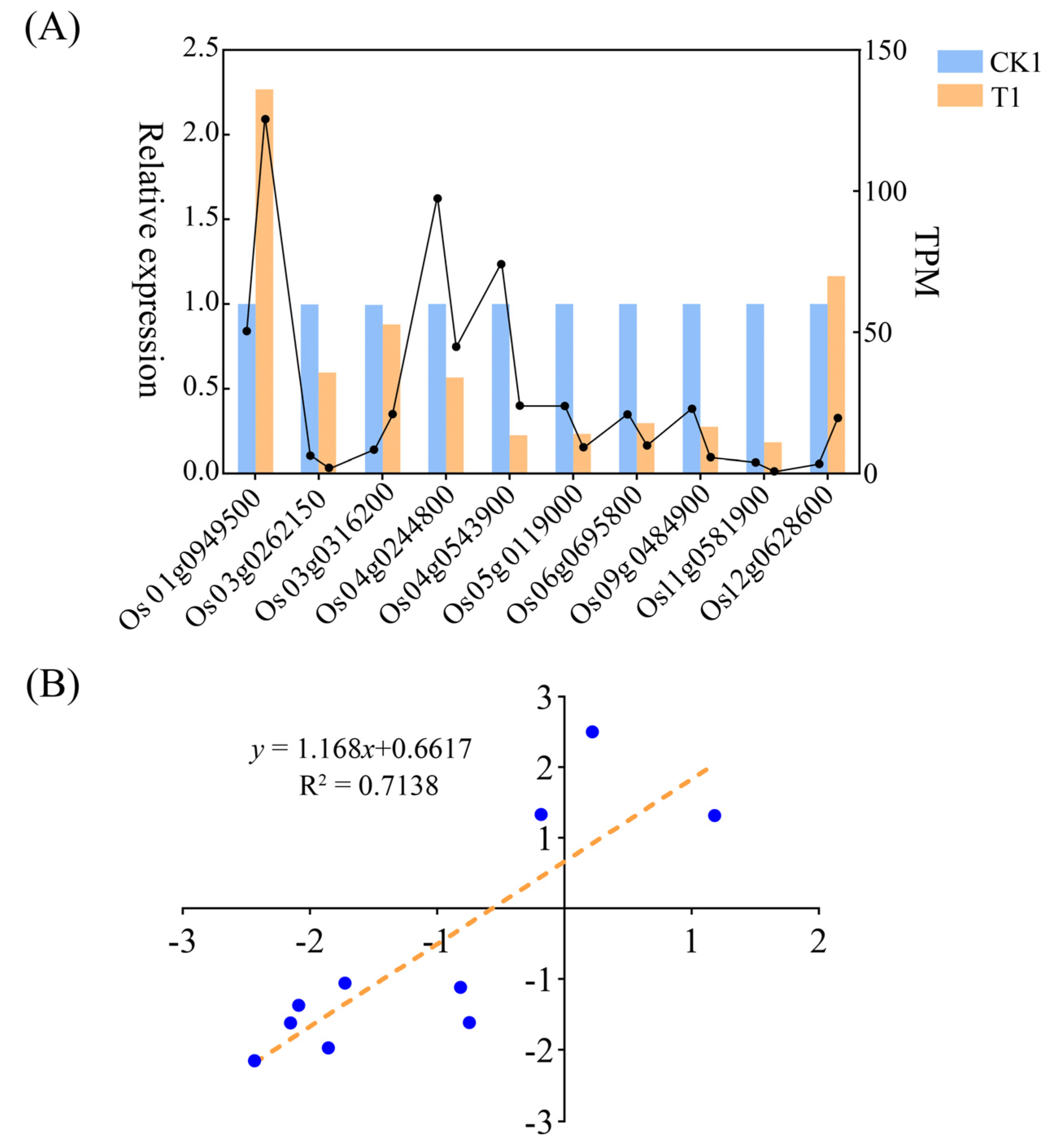
2.6. RT-qPCR Assays
3. Results
3.1. The S4 Bacteria Promotes the Growth of Rice Seedlings under Saline–Alkali Stress Conditions
3.2. The Impact of S4 Bacteria on the Physicochemical Indicators of Rice Seedlings under Saline–Alkali Conditions
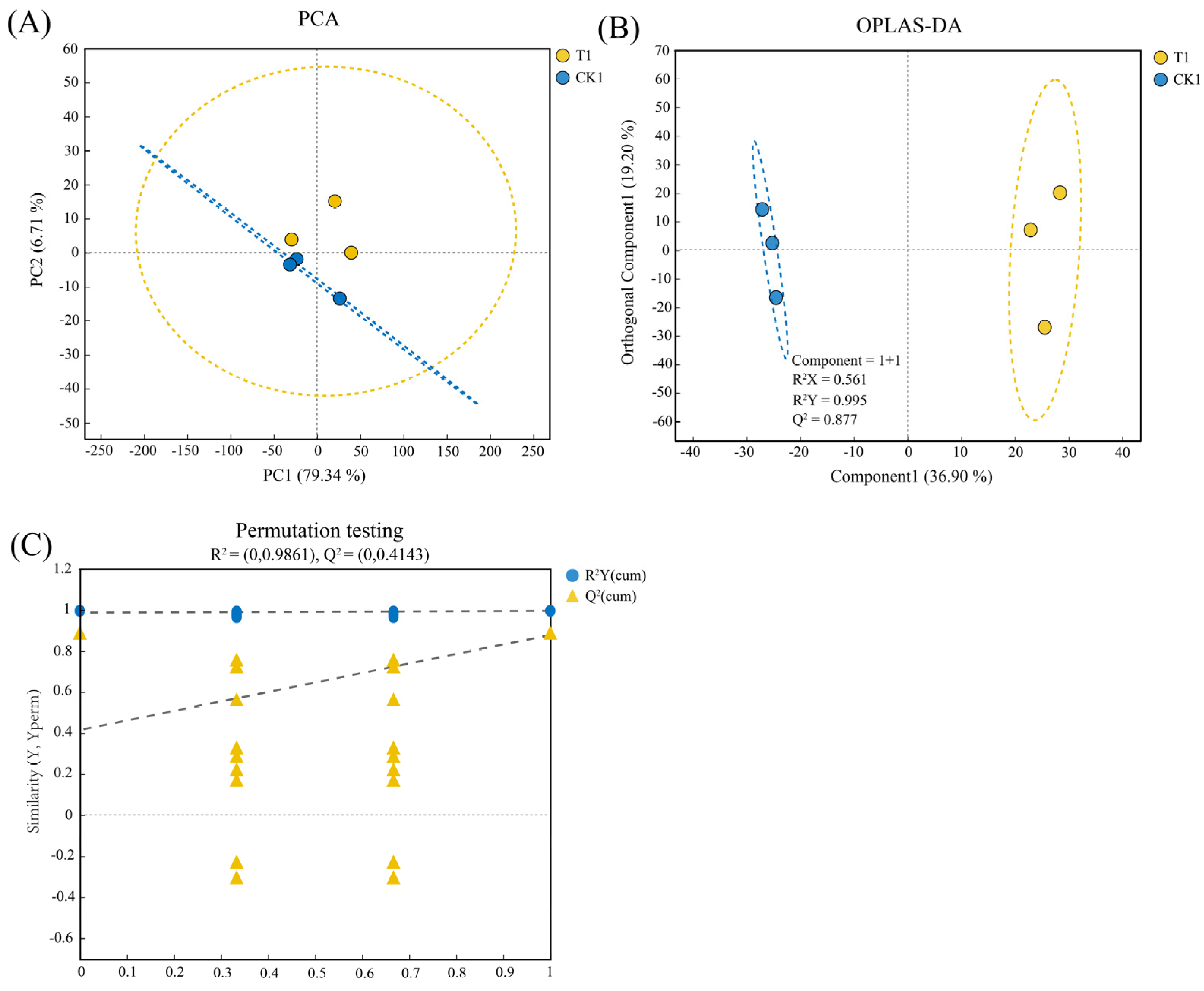
3.3. Comparative Analysis of Transcriptome and Metabolome Samples
3.4. Functional Prediction and Classification of DEGs and Differential Metabolites
3.5. Functional Enrichment Analysis of DEGs and Differential Metabolites
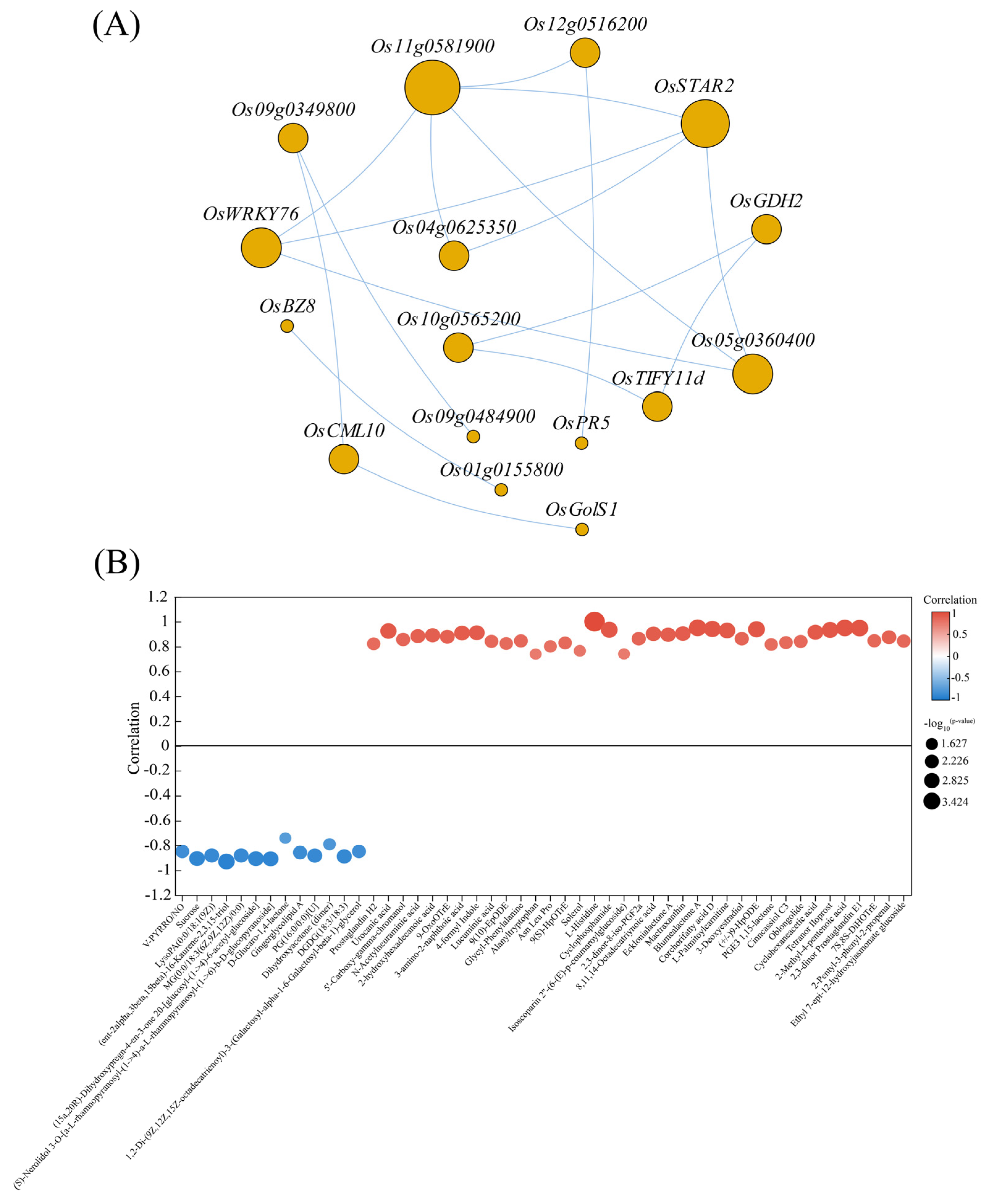
3.6. DEGs and Differential Metabolite Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Nouri, H.; Chavoshi Borujeni, S.; Nirola, R.; Hassanli, A.; Beecham, S.; Alaghmand, S.; Saint, C.; Mulcahy, D. Application of Green Remediation on Soil Salinity Treatment: A Review on Halophytoremediation. Process Saf. Environ. Prot. 2017, 107, 94–107. [Google Scholar] [CrossRef]
- Qadir, M.; Qureshi, A.S.; Cheraghi, S.A. Extent and Characterisation of Salt-affected Soils in Iran and Strategies for Their Amelioration and Management. Land Degrad. Develop. 2008, 19, 214–227. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, H.; Wang, T.; Chen, S.; Dai, S. Proteomics-based Investigation of Salt-responsive Mechanisms in Plant Roots. J. Proteom. 2013, 82, 230–253. [Google Scholar] [CrossRef]
- Li, R.; Shi, F.; Fukuda, K. Interactive Effects of Salt and Alkali Stresses on Seed Germination, Germination Recovery, and Seedling Growth of a Halophyte Spartina alterniflora (Poaceae). S. Afr. J. Bot. 2010, 76, 380–387. [Google Scholar] [CrossRef]
- Xu, B.; Cao, L.; Zhang, Z.; Li, X.; Zhao, X.; Wang, X.; Wang, Y.; Wu, B.; Zhou, W.; Lin, C.; et al. Physiological Effects of Combined NaCl and NaHCO3 Stress on the Seedlings of Two Maple Species. Front. Plant Sci. 2023, 14, 1209999. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, A.L.; Muneer, S.; Kim, Y.; Al-Rawahi, A.; Al-Harrasi, A. Silicon and Salinity: Crosstalk in Crop-Mediated Stress Tolerance Mechanisms. Front. Plant Sci. 2019, 10, 1429. [Google Scholar] [CrossRef]
- Vaishnav, A.; Shukla, A.K.; Sharma, A.; Kumar, R.; Choudhary, D.K. Endophytic Bacteria in Plant Salt Stress Tolerance: Current and Future Prospects. J. Plant Growth Regul. 2018, 38, 650–668. [Google Scholar] [CrossRef]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Khan, M.I.; Shahid, N.; Aaliya, K. Bottlenecks in Commercialisation and Future Prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Gray, E.J.; Smith, D.L. Intracellular and Extracellular PGPR: Commonalities and Distinctions in the Plant–bacterium Signaling Processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Oleńska, E.; Małek, W.; Wójcik, M.; Święcicka, I.; Thijs, S.; Vangronsveld, J. Beneficial Features of Plant Growth-promoting Rhizobacteria for Improving Plant Growth and Health in Challenging Conditions: A Methodical Review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O. Rhizosphere Microbiome Modulators: Contributions of Nitrogen Fixing Bacteria towards Sustainable Agriculture. Int. J. Environ. Res. Public Health 2018, 15, 574. [Google Scholar] [CrossRef]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially Mediated Plant Salt Tolerance and Microbiome-based Solutions for Saline Agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Olaniyan, F.T.; Alori, E.T.; Adekiya, A.O.; Ayorinde, B.B.; Daramola, F.Y.; Osemwegie, O.O.; Babalola, O.O. The Use of Soil Microbial Potassium Solubilizers in Potassium Nutrient Availability in Soil and Its Dynamics. Ann. Microbiol. 2022, 72, 45. [Google Scholar] [CrossRef]
- Chen, Y.; Ye, J.; Kong, Q. Potassium-Solubilizing Activity of Bacillus aryabhattai SK1-7 and Its Growth-Promoting Effect on Populus alba L. Forests 2020, 11, 1348. [Google Scholar] [CrossRef]
- Prittesh, P.; Avnika, P.; Kinjal, P.; Jinal, H.N.; Sakthivel, K.; Amaresan, N. Amelioration Effect of Salt-tolerant Plant Growth-promoting Bacteria on Growth and Physiological Properties of Rice (Oryza sativa) under Salt-stressed Conditions. Arch. Microbiol. 2020, 202, 2419–2428. [Google Scholar] [CrossRef]
- Kumar, A.; Patel, J.S.; Meena, V.S.; Srivastava, R. Recent Advances of PGPR Based Approaches for Stress Tolerance in Plants for Sustainable Agriculture. Biocatal. Agric. Biotechnol. 2019, 20, 101271. [Google Scholar] [CrossRef]
- Yuan, C.; Mou, C.; Wu, W.; Guo, Y. Effect of Different Fertilization Treatments on Indole-3-acetic Acid Producing Bacteria in Soil. J. Soil. Sediment. 2011, 11, 322–329. [Google Scholar] [CrossRef]
- Ganapati, R.K.; Naveed, S.A.; Zafar, S.; Wang, W.; Xu, J. Saline-Alkali Tolerance in Rice: Physiological Response, Molecular Mechanism, and QTL Identification and Application to Breeding. Rice Sci. 2022, 29, 412–434. [Google Scholar] [CrossRef]
- Theocharis, A.; Bordiec, S.; Fernandez, O.; Paquis, S.; Dhondt-Cordelier, S.; Baillieul, F.; Clément, C.; Barka, E.A. Burkholderia phytofirmans PsJN primes Vitis vinifera L. and Confers a Better Tolerance to Low Nonfreezing Temperatures. Mol. Plant Microbe Interact. 2012, 25, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, Z.; Liaquat, M.; Nazir, A.; Liaqat, R.; Iftikhar, H.; Anwar, W.; Itrat, N. Potential of Plant Growth Promoting Rhizobacteria to Mitigate Chromium Contamination. Environ. Technol. Inno. 2022, 28, 102826. [Google Scholar] [CrossRef]
- Diagne, N.; Ndour, M.; Djighaly, P.I.; Ngom, D.; Ngom, M.C.; Ndong, G.; Svistoonoff, S.; Cherif-Silini, H. Effect of Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi (AMF) on Salt Stress Tolerance of Casuarina obesa (Miq.). Front. Sustain. Food Syst. 2020, 4, 601004. [Google Scholar] [CrossRef]
- Gupta, A.; Mishra, R.; Rai, S.; Bano, A.; Pathak, N.; Fujita, M.; Kumar, M.; Hasanuzzaman, M. Mechanistic Insights of Plant Growth Promoting Bacteria Mediated Drought and Salt Stress Tolerance in Plants for Sustainable Agriculture. Int. J. Mol. Sci. 2022, 23, 3741. [Google Scholar] [CrossRef]
- Anderson, J.; Mitchell-Olds, T. Ecological Genetics and Genomics of Plant Defenses: Evidence and Approaches. Funct. Ecol. 2011, 25, 312–324. [Google Scholar] [CrossRef]
- Yan, S.; Bhawal, R.; Yin, Z.; Thannhauser, T.; Zhang, S. Recent Advances in Proteomics and Metabolomics in Plants. Mol. Hortic. 2022, 2, 17. [Google Scholar] [CrossRef]
- Pandey, A.; Khan, M.K.; Hamurcu, M.; Brestic, M.; Topal, A.; Gezgin, S. Insight into the Root Transcriptome of a Boron-Tolerant Triticum zhukovskyi Genotype Grown under Boron Toxicity. Agronomy 2022, 12, 2421. [Google Scholar] [CrossRef]
- Zou, Y.; Shi, Y.; Liu, F.; Li, E.; Yang, Q.; Chen, R.; Li, Q. The Synergistic Lipogenesis Inhibition and Molecular Mechanism of Polyphenols and Polysaccharides from Mulberry Leaf. Food Biosci. 2023, 57, 103522. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of A Transcriptome from RNA-seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A One-stop, Comprehensive Bioinformatic Platform for Multiomics Analyses. iMeta 2022, 1, 12. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.Z.; Huang, J.J.; Ding, Y.; Wu, J.M.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A Web Server for Annotation and Identification of Enriched Pathways and Diseases. Nucleic Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Maksup, S.; Supaibulwatana, K.; Selvaraj, G. High-Quality Reference Genes for Quantifying the Transcriptional Responses of Oryza sativa L. (ssp. indica and japonica) to Abiotic Stress Conditions. Chin. Sci. Bull. 2013, 58, 1919–1930. [Google Scholar] [CrossRef]
- Wang, P.; Xu, Z.; Zhang, Y.; Ma, Y.; Yang, J.; Zhou, F.; Gao, Y.; Li, G.; Hu, X. Over-expression of Spermidine Synthase 2 (SlSPDS2) in Tomato Plants Improves Saline-alkali Stress Tolerance by Increasing Endogenous Polyamines Content to Regulate Antioxidant Enzyme System and Ionic Homeostasis. Plant Physiol. Biochem. 2022, 192, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Liu, J.; Li, Q.; Liu, J.; Bai, M.; Li, X.; Zhu, Q.; Cui, Y.; Rasheed, A. Effects of Exogenous Brassinolide and AM Fungi on Growth, Photosynthetic Characteristics and Antioxidant Enzyme System of Leymus chinensis under Salt and Alkali Stress. Not. Bot. Horti. Agrobo. 2023, 51, 13378. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, F.Q.; Ren, H.; Liu, J.; Mu, W.; Wang, Y. Pseudomonas Monteilii PN1: A Great Potential P-nitrophenol Degrader with Plant Growth Promoting Traits under Drought and Saline–alkali Stresses. Biotechnol. Lett. 2019, 41, 801–811. [Google Scholar] [CrossRef]
- Xun, F.; Xie, B.; Liu, S.; Guo, C. Effect of Plant Growth-promoting Bacteria (PGPR) and Arbuscular Mycorrhizal Fungi (AMF) Inoculation on Oats in Saline-alkali Soil Contaminated by Petroleum to Enhance Phytoremediation. Environ. Sci. Pollut. Res. Int. 2014, 22, 598–608. [Google Scholar] [CrossRef]
- Singh, J.; Das, S.; Kapuganti, J.G.; Ranjan, A.; Foyer, C.H.; Thakur, J.K. Physiological Implications of SWEETs in Plants and Their Potential Applications in Improving Source-sink Relationships for Enhanced Yield. Plant Biotechnol. J. 2022, 21, 1528–1541. [Google Scholar] [CrossRef] [PubMed]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and Its Tolerance Strategies: A Review. Environ. Sci. Poll. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Xu, C.; Tang, X.; Shao, H.; Wang, H. Salinity Tolerance Mechanism of Economic Halophytes from Physiological to Molecular Hierarchy for Improving Food 1uality. Curr. Genom. 2016, 17, 207–214. [Google Scholar] [CrossRef]
- Sytar, O.; Barki, S.; Zivcak, M.; Brestic, M. The Involvement of Different Secondary Metabolites in Salinity Tolerance of Crops. In Salinity Responses and Tolerance in Plants; Kumat, V., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 21–48. [Google Scholar] [CrossRef]
- Harborne, J.B. Classes and Functions of Secondary Products from Plants. Chem. Plants 1999, 1, 1–25. [Google Scholar] [CrossRef]
- Yan, G.; Shi, Y.; Mu, C.; Wang, J. Differences in Organic Solute and Metabolites of Leymus chinensis in Response to Different Intensities of Salt and Alkali Stress. Plants 2023, 12, 1916. [Google Scholar] [CrossRef]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.S.; Watts, A.; Singh, B.P.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ABC Transporters in Maintaining Plant Homeostasis under Abiotic and Biotic Stresses. Physiol. Plantarum 2020, 171, 785–801. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the Molecular Mechanisms Mediating Plant Salt-stress Responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of Osmoprotectants in Improving Salinity and Drought Tolerance in Plants: A Review. Rev. Environ. Sci. Bio-Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Dasgupta, K.; Khadilkar, A.S.; Sulpice, R.; Pant, B.; Scheible, W.R.; Fisahn, J.; Stitt, M.; Ayre, B.G. Expression of Sucrose Transporter cDNAs Specifically in Companion Cells Enhances Phloem Loading and Long-distance Transport of Sucrose but Leads to An Inhibition of Growth and The Perception of A Phosphate Limitation. Plant Physiol. 2014, 165, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A New Insight of Salt Stress Signaling in Plant. Mol. Cell. 2016, 39, 447–459. [Google Scholar] [CrossRef]
- Taheri, P.; Tarighi, S. Riboflavin Induces Resistance in Rice Against Rhizoctonia Solani via Jasmonate-mediated Priming of Phenylpropanoid Pathway. J. Plant Physiol. 2010, 167, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Eichler, M.; Lavi, R.; Shainberg, A.; Lubart, R. Flavins are Source of Visible-light-induced Free Radical Formation in Cells. Las. Surg. Med. 2005, 37, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Jin, X.; Yang, Y.; Lin, Z.; Zhang, Y. The Regulatory Role of Riboflavin in The Drought Tolerance of Tobacco Plants Depends on ROS Production. Plant Growth Regul. 2015, 72, 269–277. [Google Scholar] [CrossRef]
- Abiko, T.; Obara, M.; Ushioda, A.; Hayakawa, T.; Hodges, M.; Yamaya, T. Localization of NAD-isocitrate Dehydrogenase and Glutamate Dehydrogenase in Rice Roots: Candidates for Providing Carbon Skeletons to NADH-glutamate Synthase. Plant Cell Physiol. 2005, 46, 1724–1734. [Google Scholar] [CrossRef]
- Huang, C.F.; Yamaji, N.; Mitani, N.; Yano, M.; Nagamura, Y.; Ma, J.F. A Bacterial-type ABC Transporter is Involved in Aluminum Tolerance in Rice. Plant Cell 2009, 21, 655–667. [Google Scholar] [CrossRef]
- Xu, J.M.; Lou, H.Q.; Jin, J.F.; Chen, W.W.; Wan, J.X.; Fan, W.; Yang, J.L. A Half-type ABC Transporter FeSTAR1 Regulates Al Resistance Possibly via UDP-glucose-based Hemicellulose Metabolism and Al Binding. Plant Soil 2018, 432, 303–314. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant Action of A Plant-derived Protein Hydrolysate Produced through Enzymatic Hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef]
- Krämer, U.; Cotter-Howells, J.D.; Charnock, J.M.; Baker, A.J.; Smith, J.A. Free Histidine as A Metal Chelator in Plants that Accumulate Nickel. Nature 1996, 379, 635–638. [Google Scholar] [CrossRef]
- Ji, H.; Yang, G.; Zhang, X.; Zhong, Q.; Qi, Y.; Wu, K.; Shen, T. Regulation of Salt Tolerance in The Roots of Zea mays by L-histidine through Transcriptome Analysis. Front. Plant Sci. 2022, 13, 1049954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, R.; Huang, K.; Huang, S.; Wang, H.; Wei, Z.; Li, Z.; Bian, M.; Jiang, W.; Wu, T.; et al. The OsWRKY63-OsWRKY76-OsDREB1B Module Regulates Chilling Tolerance in Rice. Plant J. 2022, 112, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, R.; Huang, K.; Wei, Z.; Guo, B.; Huang, S.; Li, Z.; Jiang, W.; Wu, T.; Du, X. OsWRKY76 Positively Regulates Drought Stress via OsbHLH148-mediated Jasmonate Signaling in Rice. Front. Plant Sci. 2023, 14, 1168723. [Google Scholar] [CrossRef] [PubMed]
- David, Y.C.; Ylagan, J.B.; Gonzales, H.A.; Chan, J.P.; Mondragon, J.S.; Tavera, M.; Redillas, M.C. Volatile Organic Compound Profiling of Capsicum annuum var. Longum Grown under Different Concentrations of Nitrogen. Hell. Plant Prot. J. 2021, 14, 77–88. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude | Longitude | Plants | Mass Fraction of Salt (%, g/g) | pH |
|---|---|---|---|---|---|
| Shapotou, Zhongwei, Ningxia | 37°28′ | 105°26′ | Oryza sativa | 0.32 | 8.3 |
| Suaeda glauca | 0.45 | 8.4 | |||
| Zea mays | 0.41 | 8.5 | |||
| Pingluo County, Ningxia | 39°14′ | 106°45′ | Helianthus annuus | 0.43 | 8.4 |
| Oryza sativa | 0.42 | 8.2 | |||
| Triticum aestivum | 0.45 | 8.3 | |||
| Yinchuan City, Ningxia | 38°20′ | 106°16′ | Triticum aestivum | 0.34 | 8.3 |
| Ipomoea aquatica | 0.30 | 8.2 | |||
| Sorghum dochna | 0.35 | 8.3 |
| Treatments | Plant Height (cm) | Root Length (cm) |
|---|---|---|
| CK0 | 17.77 ± 0.67 a | 8.67 ± 0.44 a |
| T0 | 18 ± 0.55 a | 8.43 ± 0.74 a |
| CK1 | 6.73 ± 0.62 c | 1.3 ± 0.15 c |
| T1 | 9.87 ± 0.38 b | 3.7 ± 0.15 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, H.; Qi, Y.; Zhang, X.; Yang, G. Transcriptomics and Metabolomics Analysis Revealed the Ability of Microbacterium ginsengiterrae S4 to Enhance the Saline-Alkali Tolerance of Rice (Oryza sativa L.) Seedlings. Agronomy 2024, 14, 649. https://doi.org/10.3390/agronomy14040649
Ji H, Qi Y, Zhang X, Yang G. Transcriptomics and Metabolomics Analysis Revealed the Ability of Microbacterium ginsengiterrae S4 to Enhance the Saline-Alkali Tolerance of Rice (Oryza sativa L.) Seedlings. Agronomy. 2024; 14(4):649. https://doi.org/10.3390/agronomy14040649
Chicago/Turabian StyleJi, Hongfei, Yuxi Qi, Xiu Zhang, and Guoping Yang. 2024. "Transcriptomics and Metabolomics Analysis Revealed the Ability of Microbacterium ginsengiterrae S4 to Enhance the Saline-Alkali Tolerance of Rice (Oryza sativa L.) Seedlings" Agronomy 14, no. 4: 649. https://doi.org/10.3390/agronomy14040649