Abstract
Soil cadmium (Cd) contamination poses a serious threat to ecosystems, and the application of phosphorus fertilizers can reduce Cd toxicity. However, the specific effects of different phosphorus fertilizers on the subcellular distribution and chemical morphology of Cd in eggplant grown in calcareous Cd-contaminated soil remain unclear. This study examined the impact of various types and levels of phosphate fertilizers on the subcellular distribution and chemical morphology of cadmium in eggplant seedlings using a two-factor analysis. The investigation was conducted via a pot experiment utilizing a two-factor analysis. The application of 0.35 g kg−1 dicalcium phosphate significantly decreased the Cd content in the subcellular distribution and induced notable alterations in the chemical morphology of Cd in eggplant roots. Specifically, the ethanol-extracted Cd state decreased by 65.45%, and the sodium chloride-extracted Cd state decreased by 64.65%. Conversely, Cd extracted by deionized water, acetic acid, hydrochloric acid, and the residue state increased by 6.20%, 4.01%, 20.87%, and 17.85%, respectively. The application of 0.35 g kg−1 dicalcium phosphate resulted in the most significant reduction in Cd content in eggplant and modification of subcellular Cd distribution and chemical morphology in roots.
1. Introduction
Unreasonable social development coupled with activities in the mining industry and the utilization of toxic and harmful heavy metal fertilizers and pesticides have resulted in cadmium becoming the most widespread heavy metal pollutant in soil. Cadmium (Cd) is highly toxic and mobile, capable of being absorbed by plants, leading to cadmium pollution issues in at least 280,000 hectares of agricultural soil in China [1,2,3]. Soil pollution research in China revealed that 19.4% of agricultural cropland experienced excessive heavy metal contamination, with Cd being the element displaying the highest soil heavy metal contamination, exceeding the standard by 7% [4,5]. Cd can transfer from the soil to the food chain and subsequently into the human body, leading to severe health issues. It has been classified as a Group 1 human carcinogen [6].
Remediation of soil Cd pollution is challenging, with a long treatment cycle requiring suitable technologies. Previous studies indicate that the application of phosphorus fertilizer is a crucial auxiliary measure for remediating heavy metal-contaminated soil [7]. Phosphorus (P), an essential element for plant growth and physiological functions, not only provides nutrients and participates in metabolic processes through various pathways [8,9,10] but also influences Cd activity in the soil, reducing its toxic effects [11,12,13]. Studies have demonstrated that applying 0.8% P fertilizer (monoammonium phosphate) significantly enhances the growth and yield of wheat while decreasing Cd accumulation in straw and seeds [14]. Similarly, the application of 109 mg P kg−1 P fertilizer (calcium superphosphate) diminishes Cd mobility in soil and Cd accumulation in lettuce [15]. P fertilizers can inhibit the uptake of Cd by plants and facilitate the formation of Cd-P precipitates in the soil. This process reduces the bioavailability of Cd, thereby mitigating damage to plants caused by Cd [16,17]. Furthermore, the bioactivity of Cd in plants is associated with its subcellular distribution and chemical morphology. Both factors can significantly impact the amount of free Cd2+ in plant cells, potentially influencing the transport and accumulation of Cd throughout the plant [18]. Research suggests that Cd primarily binds to the cell wall of tobacco leaves as phosphate or pectate, subsequently distributing to vesicles via organic nuclear protein binding [19]. Additionally, an increase in Cd content results in elevated Cd content in the cell wall of wheat [20]. Previous studies have focused on the use of P fertilizers to mitigate the toxic effects of Cd in soil, while the influence of P fertilizer application on the distribution of Cd in plant subcellular compartments and on the chemical morphology of Cd remains unclear.
Numerous studies have investigated the application of P fertilizers to alleviate the toxic effects of Cd in acidic soils [21,22,23]. However, globally, over 30% of soils are calcareous and are found in arid or semi-arid regions, where Cd pollution is also prevalent [24]. Cd exhibits low solubility in calcareous soils and is highly prone to enrichment by plants, impacting humans through the food chain [25]. Currently, there is limited research on Cd pollution in calcareous soils. Although China possesses abundant P resources, there are various high-yield P fertilizers. Thus, addressing the challenge of utilizing P resources to mitigate Cd harm while ensuring normal agricultural production is an urgent matter. Most prior studies on remediating Cd pollution focused on a single P fertilizer, with fewer exploring the effects of multiple P fertilizer types on Cd pollution [16,17,26]. Eggplant is extensively cultivated globally and ranks as the third-largest crop in the Solanaceae family [27]. Eggplants exhibit a higher propensity for cadmium accumulation compared to other vegetables [28]. Therefore, in this study, we added a quantitative amount of Cd to calcareous soil to simulate Cd-polluted soil, applied four types of P fertilizers and implemented four P application levels, and used eggplant as the research object to analyze the effects of different P fertilizers on the subcellular distribution and chemical morphology of Cd in eggplant at seedling stage. The aim was to clarify the recommended types and application rates of P fertilizers for Cd-polluted soils and to offer theoretical support for enhancing phytoremediation efficiency. We formulated the following hypotheses: (1) applying P fertilizer can reduce Cd content in all parts of eggplant seedlings and increase eggplant biomass; (2) the application of P fertilizer, particularly at a rate of 0.35 g kg−1, can diminish the subcellular Cd content in all parts of the eggplant seedlings and alter the distribution proportions; and (3) various types of P fertilizer application can decrease the content of Cd in eggplant roots, thereby reducing the toxicity of Cd pollution to eggplant roots.
2. Material and Methods
2.1. Cd-Polluted Soil Preparation
The test soil, a calcareous brown soil, was sourced from Beizhang Village (37°25′ N, 112°33′ E), Taigu District, Jinzhong City, Shanxi Province, China. Soil samples from the 0–20 cm layer were obtained using a screw auger, naturally air-dried, and sieved through a 2 mm sieve after impurity removal. CdCl2 2.5 H2O was chosen as the Cd source to simulate Cd-polluted soil. It was applied to the soil as a solution containing 5 mg kg−1 of Cd. At this concentration, Cd can significantly decrease plant fertility indicators and inhibit growth [29]. The soil was cycled between wet and dry conditions three times, equilibrated for 2 months at room temperature, air-dried, sieved with a 0.85 mm sieve, and set aside [30]. The test soil exhibited the following physicochemical properties: pH 8.2, organic matter 17.68 g kg−1, total nitrogen (N) 0.76 g kg−1, total P 0.82 g kg−1, alkali-hydrolyzable N 75.57 mg kg−1, available P 13.24 mg kg−1, available potassium (K) 145.37 mg kg−1, and total Cd 0.21 mg kg−1.
2.2. Experimental Design
The experiment involved four types of P fertilizers (monoammonium phosphate (MAP), superphosphate (SSP), diammonium phosphate (DAP), and dicalcium phosphate (DCP)) and four phosphorus levels (0, 0.15, 0.25, and 0.35 g kg−1, denoted as P0, P1, P2, and P3, respectively). A two-factor completely randomized design was used to fill each plastic pot (32 cm diameter × 30 cm depth) with 10.0 kg of equilibrated Cd-polluted soil and then add different types and levels of P fertilizers. The experiment consisted of 13 treatments, each replicated three times, with a total of 39 pots of soil. In order to ensure sufficient supply of other large elements except P, N and K fertilizers were applied to each treatment; the N fertilizer was supplemented with urea to N 0.2 g kg−1, and the K fertilizer was supplemented with K2O to 0.15 g kg−1. The soil was mixed well in each plastic pot, water was added to 20% of the soil water content, and the soil was equilibrated in the experimental base of Shanxi Agricultural University for 15 days. In order to exclude interferences, the reagents used in the experiments were analytically pure and purchased from Tianjin Komeo Chemical Reagent Co. (Tianjin Komeo Chemical Reagent Co, Tianjin, China).
After being fully watered and left for 1 day on 25 June 2022, planting of eggplant seedlings was carried out. Eggplant seedlings were selected from those that had grown 2 to 3 true leaves, and the variety was Thistle Nong Ermin Eggplant, which was purchased in the local market. Two eggplant seedlings were planted in each pot of balanced soil and watered regularly every day to maintain the moisture content in the soil at about 70% of the field water holding capacity and to provide relevant protection against pests and diseases.
2.3. Measurement Methods
Eggplant seedling samples were collected 25 days after planting. Harvested seedlings underwent triple washing under tap water, followed by rinsing with deionized water to separate roots, stems, and leaves. These components were then dried in a 105 °C desiccator for 30 min, further dried in an 80 °C desiccator to achieve constant weight, and weighed to determine dry weight. Additionally, plant samples were crushed for subsequent use [31].
Quantities of 0.1 g of crushed plant samples were weighed, 8 mL of HNO3 and 2 mL of HClO4 were added by acid digestion and digested in a hot plate for 2 to 3 h until a white solution was formed, and whole Cd was determined by graphite furnace atomic absorption spectrometry (ZEEnit 650, Analytik, Jena, Germany) [32].
In the subcellular distribution assay of Cd in plants, 0.2 g of a plant sample was weighed. The pre-cooled extract (with 0.25 M sucrose, 50 mM Tris-HCl (pH 7.5), and 1 mM dithioerythritol) was added and ground thoroughly to form a homogenate. The resulting precipitate, obtained by filtering the homogenate, represented the cell wall fraction. The clear liquid was then subjected to centrifugation at 20,000× g for 45 min, and the resulting pellet was identified as the organelle fraction. The resulting supernatant was termed the soluble tissue. Cd content was determined separately using the method outlined above for Cd determination [33].
For the determination of the chemical morphology of cadmium [34], the Cd chemical morphologies in the roots of the eggplant seedlings were extracted using different solutions: (1) ethanol extraction state (FE): 80% ethanol for inorganic Cd extraction; (2) deionized water extraction state (FW): deionized water, for extracting water-soluble Cd(H2PO4)2 from organic acid complexes and Cd; (3) sodium chloride extraction state (FNaCl): 1 M NaCl for extracting Cd bound to pectic acid and proteins; (4) acetic acid extraction state (FHAC): 2% acetic acid (HAC) was used to extract insoluble CdHPO4, Cd3(PO4)2, and other Cd-P complexes; (5) hydrochloric acid extraction state (FHCL): 0.6 M HCl for extracting oxalic acid-bound Cd; and (6) residue state (FR): Cd in the residue. Fresh root tissues were homogenized in the extracts using a mortar and pestle, diluted at a ratio of 1:100 (w/v), and shaken at 25 °C for 22 h. Centrifugation and stirring were performed at 5000× g for 10 min. Supernatants from three replicates were combined and used in each of the five extract solutions, and the Cd content of the extract was determined using the method described above.
2.4. Calculations and Statistical Analysis
Data were processed in Excel 2010. One-way ANOVA (Duncan’s test, p < 0.05) was conducted using SPSS 23. Two-way ANOVA for the type and level of P fertilizer for each indicator was performed using SPSS 23, and significant differences were determined using the least significant difference (LSD) test. The correlation between the subcellular and root chemical morphology of Cd in each part of the eggplant was investigated using Pearson’s analysis. Origin 2024 software was employed for plotting. The bioconcentration factor (BCF) and the translocation factor (TF) for eggplant Cd were calculated based on dry weight using the following equation [35].
where represents Cd concentrations in roots, stems, and leaves and is Cd in soil (both in mg kg−1) [36].
where represents the Cd concentration in stems and leaves and is Cd in roots (both in mg kg−1) [36].
3. Results
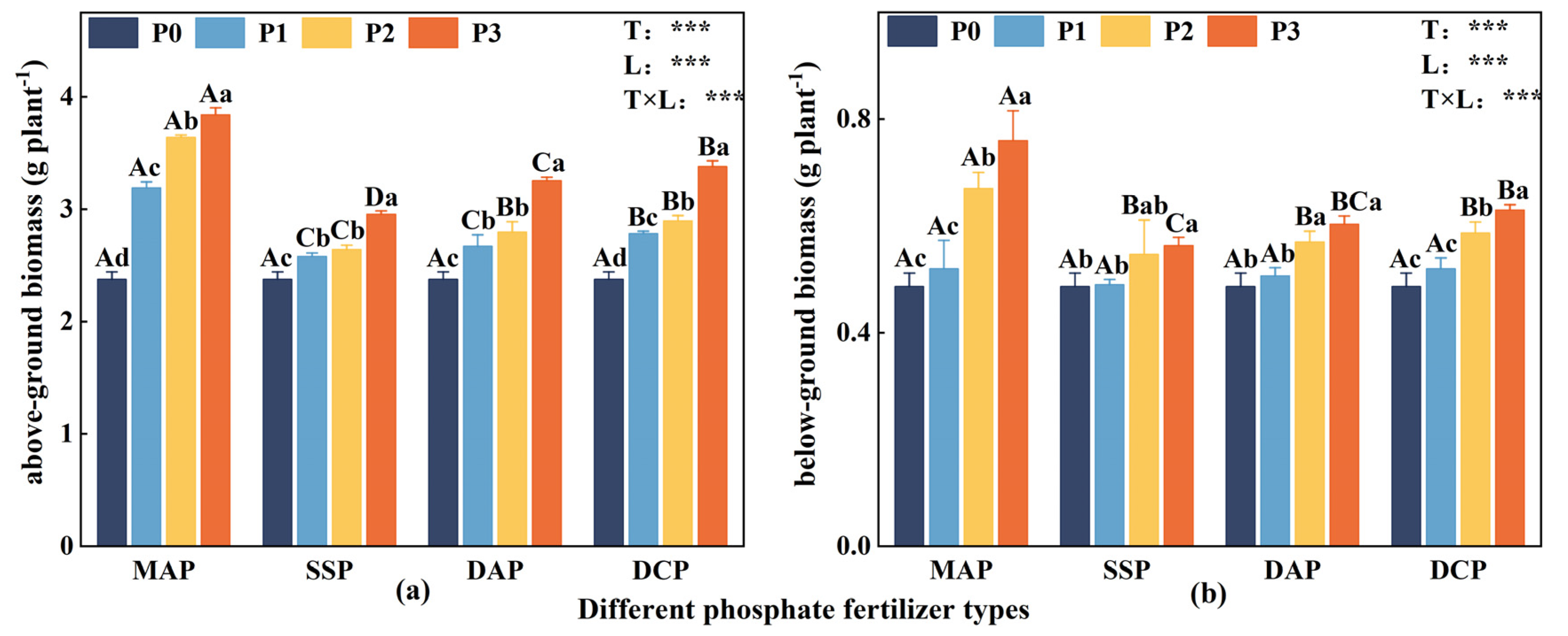
3.1. Eggplant Seedling Biomass
Two-way ANOVA indicated significant effects (p < 0.05) of P fertilizer type, P fertilizer level, and their interaction on both above- and belowground biomass of eggplant at the seedling stage under Cd stress. In the Cd-polluted environment, both above- and belowground biomass were lower in the no-P treatment (P0). The biomass showed a significant increase with the application of P fertilizer (Figure 1). Among the various P fertilizer types, the impact of applied MAP on both above- and belowground biomass was the most significant (p < 0.05), followed by DCP. When comparing different P fertilizer levels, it was observed that the above- and belowground biomass exhibited the highest increase at the P3 level, and the various P fertilizer types were ranked as follows: MAP > DCP > DAP > SSP. In comparison with P0, the aboveground biomass of MAP, DCP, DAP, and SSP increased significantly by 61.6%, 24.4%, 36.9%, and 42.2% at the P3 level; moreover, belowground biomass exhibited the most significant increase with the application of MAP at the P3 level.
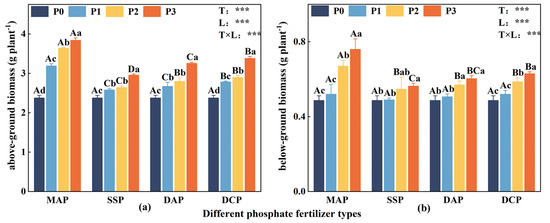
Figure 1.
Aboveground biomass (a) and belowground biomass (b) of eggplant at seedling stage under different P fertilizer application rates. A, B, C, and D represent significant differences between types at the same level of P fertilizer, while a, b, c, and d represent significant differences between levels of the same P fertilizer type. T and L indicate the significance levels of two-factor ANOVA for P fertilizer type and P fertilizer level, respectively. *** p < 0.05 [35].
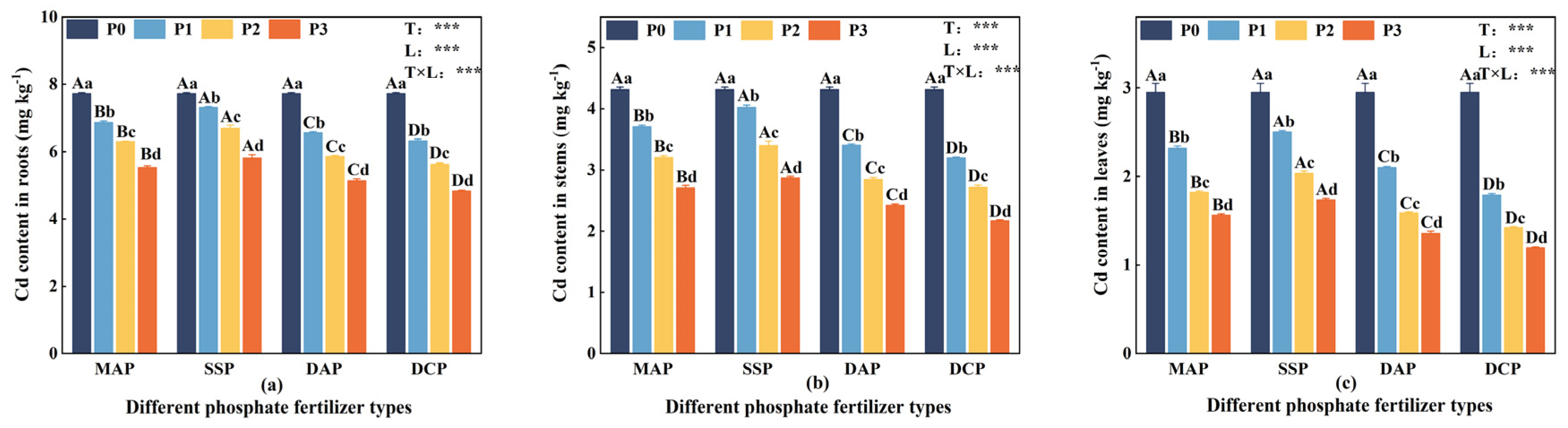
3.2. Cd Content in Eggplant Seedlings
Two-way ANOVA indicated that the type and level of P fertilizer, along with their interaction, had significant effects (p < 0.05) on the Cd content in the roots, stems, and leaves of eggplant at the seedling stage under Cd stress (Figure 2). The Cd content in roots, stems, and leaves was higher under P0 conditions, gradually decreasing with the application of P fertilizer. Among different P fertilizer types, the Cd content in roots, stems, and leaves followed the order DCP < DAP < MAP < SSP, with DCP significantly reducing Cd content, while SSP had a lesser impact. Across different levels of P fertilizer, the reduction in Cd content in each part became more pronounced with increasing levels, especially at the P3 level. DCP, when compared to SSP at the P3 level, significantly reduced Cd content in the roots, stems, and leaves of eggplant seedlings by 16.75%, 24.36%, and 31.17%, respectively. At the P3 level, DCP exhibited a substantial reduction in Cd content in the roots, stems, and leaves of eggplant seedlings by 59.82%, 98.54%, and 147.1%, respectively, compared to the P0 level.
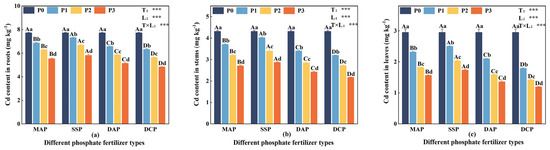
Figure 2.
Cd content in eggplant at seedling stage with different P fertilizers. (a) Cd content in roots. (b) Cd content in stems. (c) Cd content in leaves. A, B, C, and D represent significant differences between types at the same level of P fertilizer, while a, b, c, and d represent significant differences between levels of the same P fertilizer type. T and L indicate the significance levels of two-factor ANOVA for P fertilizer type and P fertilizer level, respectively. *** p < 0.05.
3.3. Bioconcentration and Translocation Factors for Eggplant Seedlings
The application of various types and levels of P fertilizers exhibited significant differences (p < 0.01) in the bioconcentration factor (BCF) and translocation factor (TF) across all parts of the eggplant (Table 1). BCF and TF displayed a decreasing trend with P fertilizer application. Roots exhibited a stronger enrichment capacity, while leaves showed a weaker capacity, indicating that Cd pollution primarily accumulated in roots. Root-to-leaf translocation capacity was weaker than root-to-stem translocation, suggesting that translocation from roots to aboveground sites mainly occurred in the stem. The BCFs and TFs of all eggplant parts for different P fertilizer types followed the order DCP < DAP < MAP ≤ SSP, with DCP showing the most significant decrease and SSP the slowest. Across different P fertilizer levels, the reduction in BCF and TF became more pronounced with increasing levels, reaching maximum significance at the P3 level. In comparison to the P0 level, the application of DCP at the P3 level significantly reduced BCF in roots, stems, and leaves by 18.93%, 34.74%, and 47.56%, respectively, and TFstem/root and TFleaf/root by 19.5% and 35.32%.

Table 1.
BCFs and TFs of Cd for eggplant seedlings with different P fertilizer applications.
3.4. Subcellular Distribution of Cd in Various Parts of Eggplant
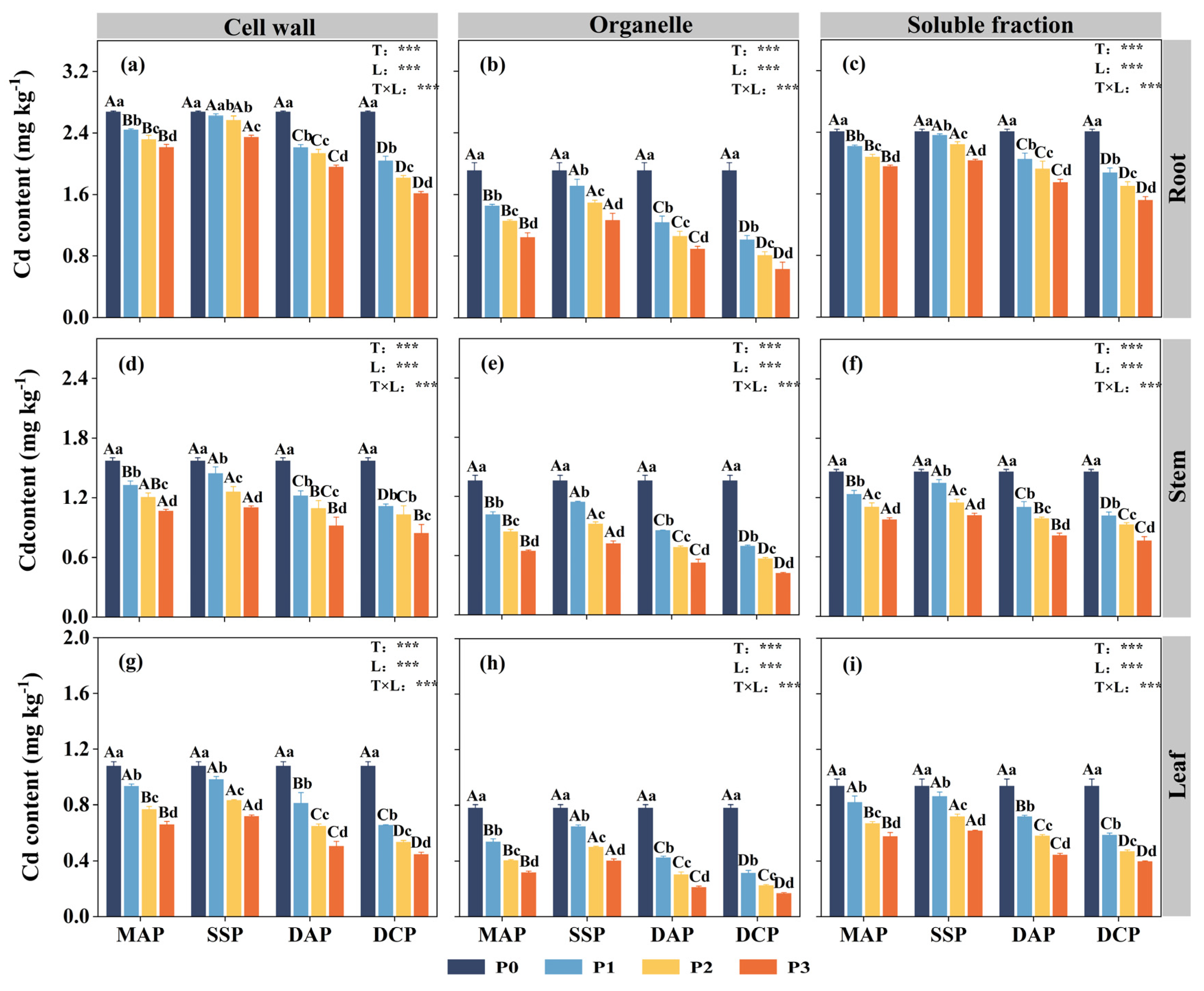
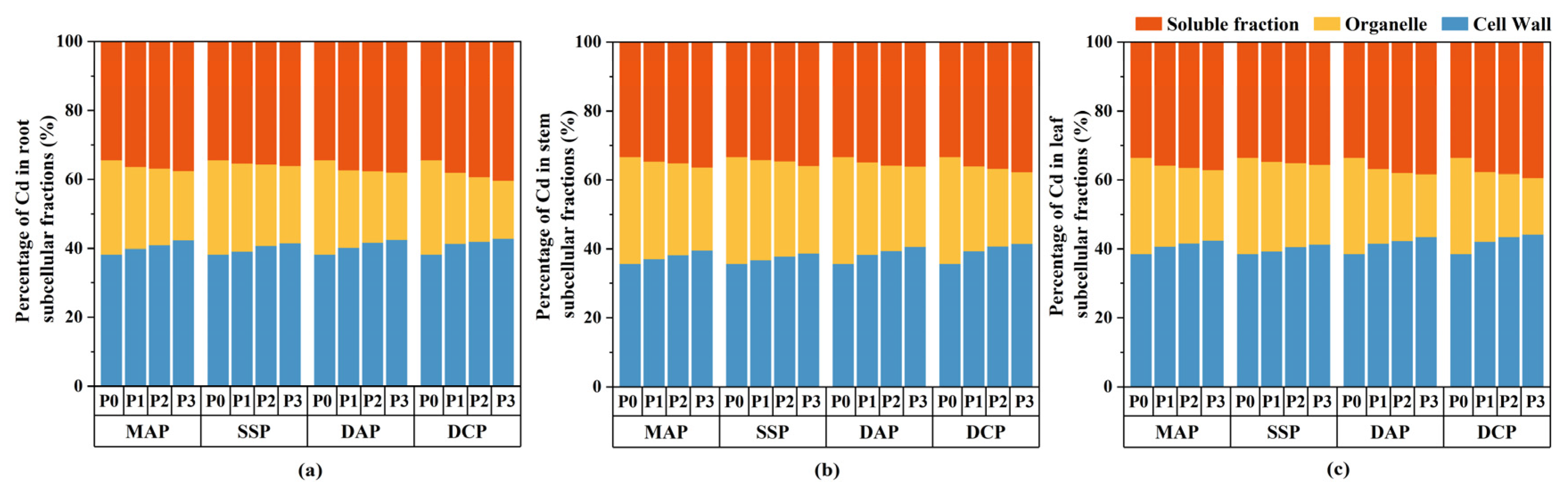
Two-way ANOVA indicated significant effects (p < 0.05) of P fertilizer type, P fertilizer level, and their interaction on the subcellular distribution of Cd in the roots, stems, and leaves of eggplant at the seedling stage under Cd stress (Figure 3). The subcellular distribution of Cd in all parts of the eggplant was significantly decreased with the application of various types and levels of P fertilizers compared to the P0 level. The most significant reduction in the subcellular distribution of Cd in eggplant roots, stems, and leaves was observed with the application of DCP at the P3 level (p < 0.05), whereas the reduction was not significant with SSP. Relative to P0, the application of DCP at the P3 level led to reductions in the cell wall, organelle, and soluble fraction Cd contents of eggplant roots of 39.58%, 66.95%, and 36.98%, respectively; of eggplant stems of 46.28%, 69.02%, and 47.60%, respectively; and of eggplant leaves of 58.59%, 78.71%, and 57.55%, respectively. Applying P fertilizer changed the Cd distribution proportion at the subcellular level compared to P0. The trend observed for the distribution of Cd across all parts of the eggplant was as follows: cell walls > soluble fractions > organelles (Figure 4). The application of P fertilizer significantly increased the proportion of Cd in cell walls and soluble fractions of roots, stems, and leaves and decreased the proportion of Cd in organelles.
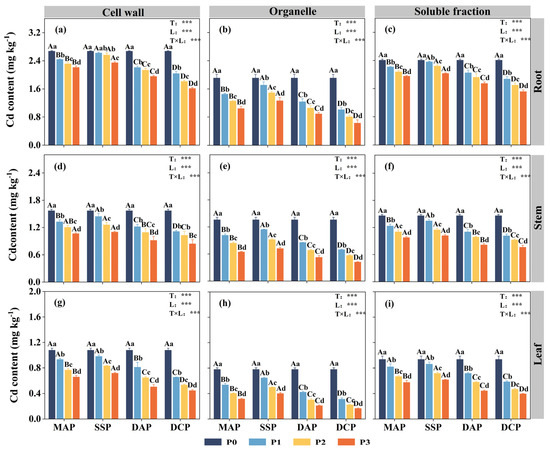
Figure 3.
Subcellular distribution of Cd in various parts of eggplant seedlings treated with different P fertilizers. (a) Root cell wall Cd content. (b) Root organelle Cd content. (c) Root soluble fraction Cd content. (d) Stem cell wall Cd content. (e) Stem organelle Cd content. (f) Stem soluble fraction Cd content. (g) Leaf cell wall Cd content. (h) Leaf organelle Cd content. (i) Leaf soluble fraction Cd content. A, B, C, and D represent significant differences between types at the same level of P fertilizer, while a, b, c, and d represent significant differences between levels of the same P fertilizer type. T and L indicate the significance levels of two-factor ANOVA for P fertilizer type and P fertilizer level, respectively. *** p < 0.05.
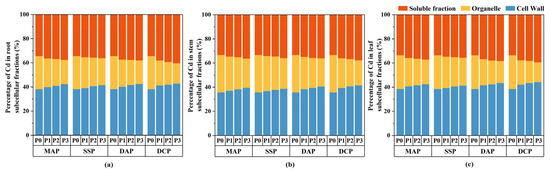
Figure 4.
Proportion of Cd distribution in subcellular fractions in eggplant seedlings under different P fertilizer application rates. (a) Percentage distribution of Cd in root subcellular fractions. (b) Percentage distribution of Cd in stem subcellular fractions. (c) Percentage distribution of Cd in leaf subcellular fractions.
3.5. Chemical Morphology of Cd in the Roots of Eggplant
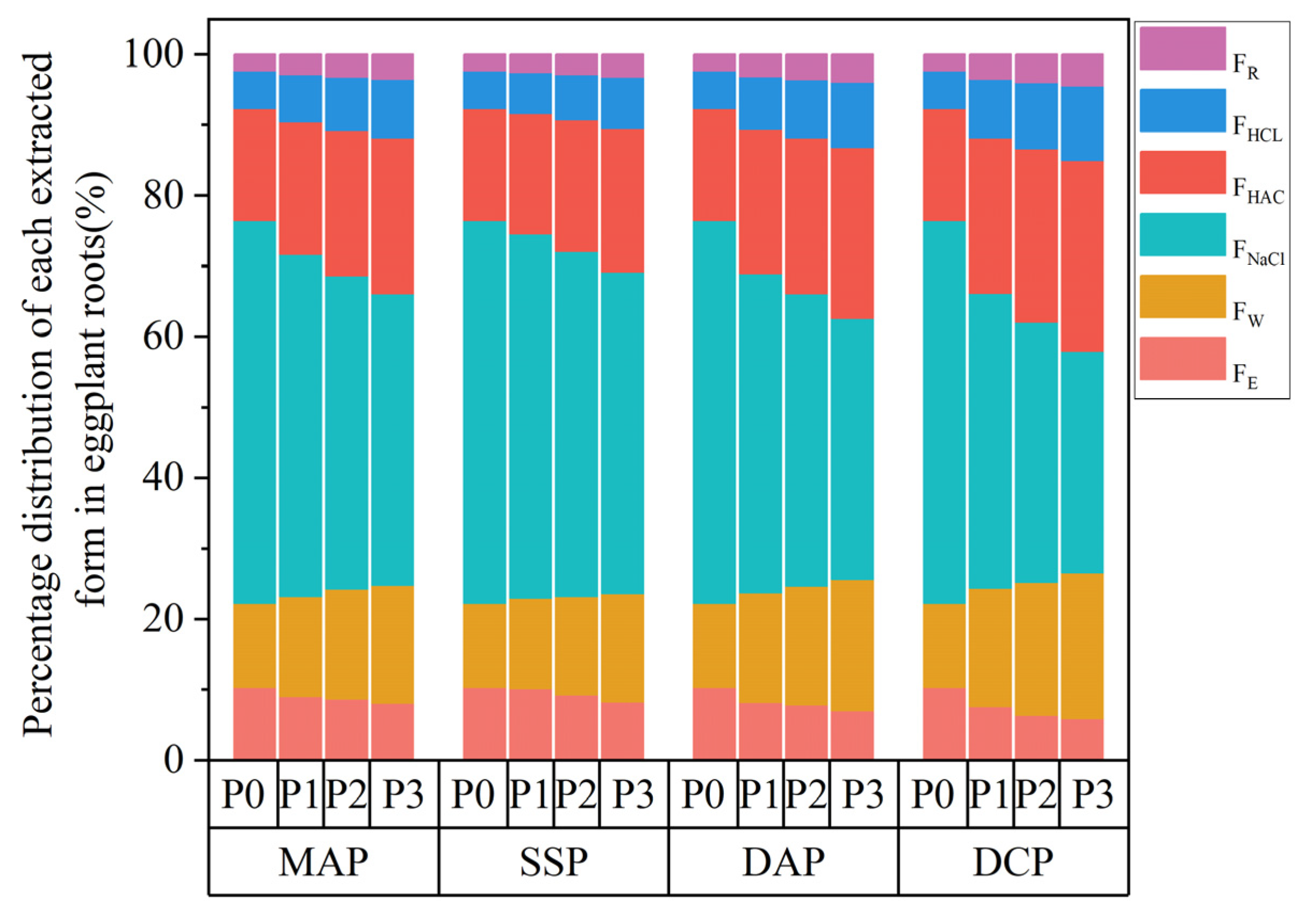
Two-way ANOVA revealed that the type and level of P fertilizer, along with their interaction, significantly (p < 0.01) influenced FE, FW, and FNaCl in seedling roots of eggplant under Cd stress (Table 2). Under the P0 condition, each extraction state in roots exhibited the order FNaCl > FHAC > FW > FHCL > FE > FR. The application of different types and levels of P fertilizers led to a decrease in FE and FNaCl and an increase in FW, FHAC, FHCL, and FR in eggplant roots. The application of P fertilizer led to changes in the proportions of each extractive state distribution in eggplant (Figure 5). The proportions of FNaCl and FE decreased gradually, while the proportions of FW, FHAC, FHCL, and FR increased. Notably, FNaCl still exhibited the highest proportion among the extractive states. Application of DCP at the P3 level decreased FE and FNaCl in eggplant roots by 65.45% and 64.65%, respectively, and increased FW, FHAC, FHCL, and FR in eggplant roots by 6.20%, 4.01%, 20.87%, and 17.85%, respectively, as compared to the P0 level. Regarding the assigned percentage for each chemical morphology, the FE percentage decreased by 43.43%, and the FHCL percentage significantly increased by 97.93% at the P3 level compared to the P0 level with the application of DCP. These were the two extraction states with the most pronounced increase and decrease in the percentage of chemical morphologies in eggplant roots.
Table 2.
Cd chemical morphologies in eggplant seedling roots under various P fertilizers (mg kg−1).
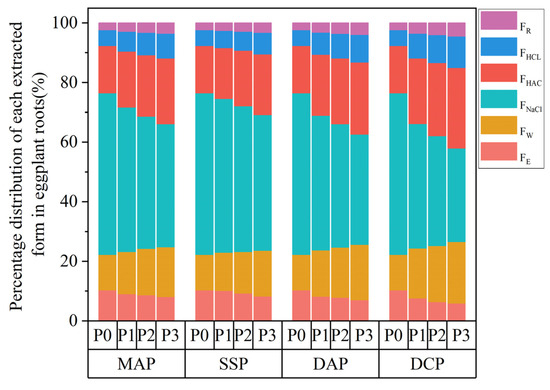
Figure 5.
Percentage of cadmium in the chemical morphologies of eggplant seedling roots with the application of different phosphorus fertilizers.
3.6. Correlation Analysis of Subcellular and Chemical Morphologies in Eggplant Seedlings
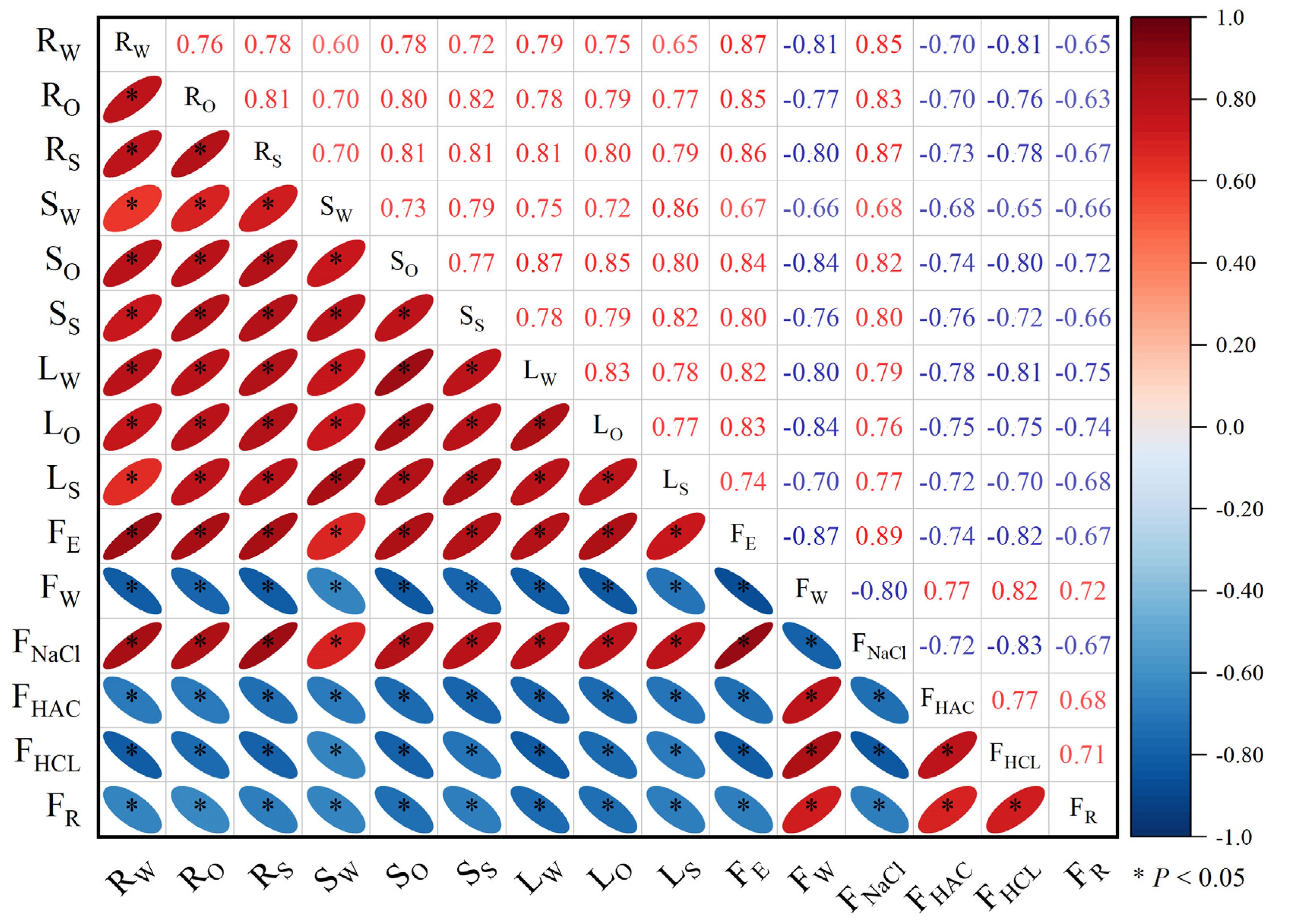
Correlation analysis revealed a significant correlation (p < 0.05) between the subcellular distribution of Cd in various parts of eggplant seedlings and the chemical morphology of Cd in roots (Figure 6). FE and FNaCl were significantly positively correlated with the subcellular distribution of Cd in roots, stems, and leaves of eggplants in the seedling stage, while FW, FHAC, FHCL, and FR were significantly negatively correlated with the subcellular distribution of Cd in roots, stems, and leaves of eggplants in the seedling stage. Following the application of P fertilizer, the subcellular distribution of Cd in various parts of eggplant seedlings decreased, causing a reduction in cell wall and protoplasmic membrane permeability and an increase in retention capacity. This led to a decrease in Cd bound to proteins, pectin, and other substances on the cell wall (FNaCl and FE) and an increase in the weakly migratory activity of FW, FHAC, FHCL, and FR.
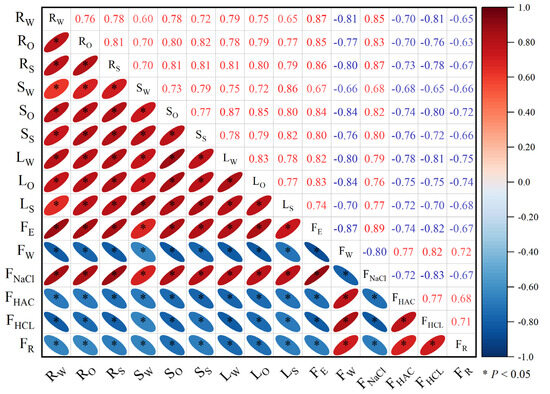
Figure 6.
Correlation between subcellular distribution and chemical morphology of Cd in roots at the seedling stage of eggplant treated with different P fertilizers. RW: root cell wall; RO: root organelle; RS: root soluble fraction; SW: stem cell wall; SO: stem organelle; SS: stem soluble fraction; LW: leaf cell wall; LO: leaf organelle; LS: leaf soluble fraction.
4. Discussion
4.1. Effect of Phosphorus Application on Eggplant Biomass and Cadmium Content under Cadmium Stress
P fertilizer was effective in mitigating the stress effect of Cd on eggplant biomass and promoting plant growth [37]. Both aboveground and belowground biomass of eggplant at the seedling stage increased gradually with the addition of P fertilizer (Figure 1), consistent with the findings of Li [38]. Eggplant biomass significantly increased with higher levels of P fertilizer. This is attributed to P being an essential nutrient for plant growth, participating in the synthesis and metabolism of plant carbohydrates and proteins, promoting plant growth, and ultimately increasing eggplant biomass. Cd contents in eggplant roots, stems, and leaves were higher under the no P fertilizer treatment in this experiment, representing the plant’s greatest exposure to the toxic effects of Cd (Figure 2). Application of P fertilizer, especially at the P3 level with DCP, significantly reduced Cd content in all parts of the eggplant seedling, indicating that P application had a notable effect on alleviating Cd toxicity. This aligns with the findings of previous research by some scholars [11,12,13,14,26]. Previous studies have highlighted that P exhibits tolerance and physiological responses to Cd in mangrove forests under Cd stress, leading to the accumulation of most Cd in mangrove roots. The addition of P induces the immobilization of Cd in mangrove roots [39]. Moreover, studies suggest that P addition can mitigate the mechanistic effects of Cd toxicity on ryegrass roots and reduce the leaching of Cd from contaminated soil [40]. The bioconcentration factors of Cd without P application and translocation factors were higher, while the bioconcentration and translocation factors of eggplant parts gradually decreased after the application of P fertilizer, which could enrich more Cd in the roots and reduce the migration of Cd to the aboveground parts (Table 1). This is because the application of P fertilizer promotes the formation of precipitation between P and Cd, and the Cd content absorbed and accumulated in eggplant roots and aboveground is reduced, and insoluble P precipitation occurs in the root cell wall, vesicles, etc., which prevents upward transport of Cd, and further reduces the possibility of enrichment and transport of Cd in eggplant tissues to aboveground parts [15].
4.2. Effect of P Application under Cd Stress on the Subcellular Distribution and Chemical Morphology of Cd in Eggplants
Selective regional distribution of heavy metal contaminants at the subcellular level may partially alleviate their toxic effects on organisms [41,42]. The plant cell wall serves as the primary barrier preventing Cd2+ transport from the external environment into the plant cell, and it is also the initial storage site for Cd [43]. The negatively charged surface of the cell wall interacts with positively charged Cd2+, facilitating the complexation of Cd on the cell wall and diminishing Cd’s entry into the cell [44,45]. The soluble portion of the cell primarily comprises cytoplasm and vesicles, which serve as the primary organs for Cd accumulation [46]. The vesicles contain a significant amount of sulfur-rich states and organic acids capable of chelating and sequestering Cd to prevent damage caused by the heavy metal to the organelles [47]. Higher levels of free Cd2+ in the cell walls and soluble fraction of the plants and the cell walls and the soluble fractions of eggplant roots, stems, and leaves were more influenced by Cd toxicity in the absence of P treatments (Figure 3). Previous studies have shown that barley complexes a significant amount of Cd in the soluble fraction or stores it in the cell wall when exposed to Cd stress, thereby reducing Cd damage to organelles [48,49]. The present study aligns with these findings. In this study, it was observed that the Cd content of the organelles was lower than that of the cell wall and the soluble fraction of the cells under no P treatment. The application of P fertilizer resulted in a reduction in Cd content in subcellular fractions across all parts of eggplant. Following the entry of Cd into the plant, P demonstrated the ability to restrict Cd mobility by promoting the formation of insoluble Cd-P complexes in the vesicles along with pectate/protein integrated Cd in the cell wall [50]. This process inhibited Cd uptake by subcellular fractions in all parts of the eggplant [51]. Studies have demonstrated that two types of vegetable hearts in Cd-contaminated environments can mitigate the subcellular distribution of Cd and reduce the risk of Cd pollution in the food chain through the application of P fertilizer [18]. The application of P fertilizer altered the distribution proportion of Cd in subcellular fractions (Figure 4). The proportion of Cd increased in the eggplant cell wall and soluble fraction, while decreasing in organelles. Other researchers demonstrated that applying P fertilizer in a high-Cd pollution environment increased the proportion of Cd in the cell walls of rice roots and shoots, indicating a detoxifying effect of P on rice organelles [52]. The allocation proportion of Cd in eggplant root and shoot organelles was significantly lower than that in roots, rendering Cd less toxic to stems and leaves than to roots. P and Cd formed metal chelates in the plant’s cell wall, reducing the long-distance transport of Cd2+ in the xylem and limiting its mobility to the aboveground parts of the plant [53]. Reduction in the Cd content of subcellular fractions in various parts of eggplant with the most significant change in the distribution proportion was observed when DCP was applied at the P3 level in this study.
Cd in eggplant exists in various chemical morphologies in the roots, and these different morphologies can influence the activities, toxicity, migratory capacity, and ease of substrate separation of cadmium [54]. Generally, Cd in plants is predominantly found in the form of FNaCl. FNaCl is the form in which Cd binds to proteins, and Cd exhibits a strong affinity for sulfur groups in proteins or other organic compounds. This interaction can interfere with enzyme activity and impact normal crop growth and development [55]. FE exhibited higher migratory and toxic properties. Organic acid salts of FW and M(H2PO4)2 could form chelates with Cd at the binding sites on the surface of the protoplast membrane, thereby limiting the migratory properties of Cd in eggplant. FHAC represents the form in which Cd binds to MHPO4, and the adsorption of Cd by P species reduces the amount of Cd in the active state, decreasing Cd mobility within the plant. Cd exhibits the weakest migratory activity and toxicity in FHCl and FR [56]. In this study, under no P treatment, the extraction state of Cd in the roots showed FNaCl > FHAC > FW > FHCl > FE > FR (Table 2), and the application of P fertilizer changed the chemical morphology of Cd in the roots of eggplants, in which FE and FNaCl, which have high migratory activity, were decreasing, and their percentage was also decreasing (Figure 5). Previous studies have shown that the application of P fertilizer enhances the ability of phosphate to bind to Cd in plants, leading to a gradual increase in FW and an increase in the weaker migratory activities of FHAC, FHCl, and FR. These findings align with the results of the present study [57].
4.3. Mechanisms of Applying Different P Fertilizers to Mitigate Cd Pollution
The application of different P fertilizers can alter the soil pH (Figure S1) and diminish the effectiveness of soil Cd. Previous studies have explored the immobilization of Cd-contaminated soil through the application of SSP and DAP. The use of SSP and DAP can lower the pH value of the soil, subsequently decreasing the migration and dissolution of Cd in the soil [58]. However, this study was conducted in calcareous soil, which is alkaline. The NH4+ and H2PO42− present in MAP and DAP form complexes with metal cations like Mn2+, Mg2+, and Ca2+ in calcareous soil, for example, CaHPO4 or Ca3(PO4)2 precipitates. This complexation reduces the adsorption sites of effective ions in MAP and DAP for Cd2+, thereby weakening the efficacy of Cd pollution treatment through the application of MAP and DAP in this study. The aqueous solution of SSP is acidic. When applied, it neutralized the calcareous soil in this study, diminishing the effectiveness of SSP in limiting Cd. Numerous studies have demonstrated that PO42− is readily immobilized in calcareous soil, consequently diminishing fertilizer effectiveness. However, Ca2+ also has a significant effect on the fixation of cadmium [59]. The DCP aqueous solution, with its weak alkalinity, can increase soil pH (Figure S1), enhancing the negative charge on the surface of clay minerals, hydrated oxides, and organic matter, consequently improving their adsorption capacity for heavy metals. Research indicates a significant negative correlation between pH value and cadmium availability. With increasing pH, cadmium availability in the soil decreases, resulting in reduced cadmium uptake by plants [60,61]. Cadmium exhibits the lowest effectiveness in soil due to its high Ca2+ content and pH. Thus, in this study, the application of DCP had the most pronounced effect on P3 levels.
5. Conclusions
Application of different types and levels of P fertilizers to calcareous Cd-contaminated soils increased above- and belowground biomass and decreased Cd contents in all parts of eggplant during the seedling stage. P fertilizer application influenced the subcellular distribution of Cd, leading to a gradual increase in the proportion of Cd in the cell wall and cell soluble fractions, along with a gradual decrease in the proportion of Cd in the organelles. Additionally, it further altered the chemical morphology of Cd in the roots of eggplant seedlings. In this study, the most significant effect of DCP application was its reducing the damage of Cd pollution to all parts of eggplant at the seedling stage. Moreover, the reduction in Cd toxicity was more obvious with higher P fertilizer application. Therefore, the application of 0.35 g kg−1 of DCP in calcareous soil had the greatest effect on the subcellular distribution and chemical morphology of Cd in eggplant, which could effectively reduce the toxic effects of Cd pollution on plants.
6. Prospects
This study constituted preliminary research and had several shortcomings such that further exploration is needed:
- (1)
- Only the chemical morphology of cadmium in roots was investigated; the chemical morphology in stems and leaves during the seedling stage remained unstudied.
- (2)
- The study focused solely on the seedling stage of eggplant and lacked a systematic examination of the mature stage, thus presenting limitations in the exploration.
- (3)
- Further investigation of the mechanism by which phosphorus fertilizer mitigates cadmium toxicity is warranted.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14050932/s1, Figure S1: Soil pH of different P fertilizers applied. A, B, C, and D represent significant differences between types at the same level of P fertilizer, while a, b, c, and d represent significant differences between levels of the same P fertilizer type. T and L indicate the significance levels of two-factor ANOVA for P fertilizer type and P fertilizer level, respectively. *** p < 0.05.
Author Contributions
Methodology and original draft preparation were conducted by Q.M.; methodology, project administration, review and editing of the manuscript, funding acquisition, and supervision were performed by W.F.; review and editing of the manuscript were carried out by F.L. and G.W.; formal analysis was conducted by X.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study received support from the National Natural Science Foundation of China (NSFC) Joint Fund (grant no. U1710255–4) and the Academic Recovery Research Program of Shanxi Agricultural University (grant no. 2020xshf20).
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cao, C.; Zhang, N.; Tao, R.; Zheng, J.; Hu, H.; Li, J.; Ma, Y.; Liao, X. Screening for Low-Cadmium Accumulation in Maize Varieties Based on Species Sensitivity Distribution and Research on Soil Environmental Thresholds. Agronomy 2023, 13, 1960. [Google Scholar] [CrossRef]
- Wang, C.-C.; Zhang, Q.-C.; Yan, C.-A.; Tang, G.-Y.; Zhang, M.-Y.; Ma, L.Q.; Gu, R.-H.; Xiang, P. Heavy metal(loid)s in agriculture soils, rice, and wheat across China: Status assessment and spatiotemporal analysis. Sci. Total Environ. 2023, 882, 163361. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Huang, X.; Sun, L.; Li, S.; Chen, Y.; Cao, X.; Wang, W.; Dai, J.; Rinnan, R. Screening stably low cadmium and moderately high micronutrients wheat cultivars under three different agricultural environments of China. Chemosphere 2020, 241, 125065. [Google Scholar] [CrossRef]
- Chen, R.; de Sherbinin, A.; Ye, C.; Shi, G. China’s Soil Pollution: Farms on the Frontline. Science 2014, 344, 691. [Google Scholar] [CrossRef]
- Chen, H.; Teng, Y.; Lu, S.; Wang, Y.; Wang, J. Contamination features and health risk of soil heavy metals in China. Sci. Total Environ. 2015, 512–513, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, H.; Kopittke, P.M.; Zhao, F.-J. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, Y.; Hou, J.; Wu, L.; Christie, P.; Liu, W. Microbial community assembly of the hyperaccumulator plant Sedum plumbizincicola in two contrasting soil types with three levels of cadmium contamination. Sci. Total Environ. 2023, 863, 160917. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Yang, W.; Chen, Y.; Yang, L.; Zhou, H.; Yang, Y.; Zhao, Z.; Wu, P.; Zia-ur-Rehman, M. Exploring the mechanism of Cd uptake and translocation in rice: Future perspectives of rice safety. Sci. Total Environ. 2023, 897, 165369. [Google Scholar] [CrossRef]
- Wang, X.; Ai, S.; Liao, H. Deciphering Interactions between Phosphorus Status and Toxic Metal Exposure in Plants and Rhizospheres to Improve Crops Reared on Acid Soil. Cells 2023, 12, 441. [Google Scholar] [CrossRef] [PubMed]
- Barrow, N.J. How understanding soil chemistry can lead to better phosphate fertilizer practice: A 68 year journey (so far). Plant Soil 2022, 476, 117–131. [Google Scholar] [CrossRef]
- Wielgusz, K.; Praczyk, M.; Irzykowska, L.; Świerk, D. Fertilization and soil pH affect seed and biomass yield, plant morphology, and cadmium uptake in hemp (Cannabis sativa L.). Ind. Crops Prod. 2022, 175, 114245. [Google Scholar] [CrossRef]
- Dai, M.; Liu, W.; Hong, H.; Lu, H.; Liu, J.; Jia, H.; Yan, C. Exogenous phosphorus enhances cadmium tolerance by affecting cell wall polysaccharides in two mangrove seedlings Avicennia marina (Forsk.) Vierh and Kandelia obovata (S., L.) Yong differing in cadmium accumulation. Mar. Pollut. Bull. 2018, 126, 86–92. [Google Scholar] [CrossRef]
- Wu, C.; Yan, S.; Zhang, H.; Luo, Y. Chemical forms of cadmium in a calcareous soil treated with different levels of phosphorus-containing acidifying agents. Soil Res. 2015, 53, 105. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Rehman, M.Z.U.; Ali, S.; Rizwan, M.; Naeem, A.; Maqsood, M.A.; Khalid, H.; Rinklebe, J.; Ok, Y.S. Residual effects of monoammonium phosphate, gypsum and elemental sulfur on cadmium phytoavailability and translocation from soil to wheat in an effluent irrigated field. Chemosphere 2017, 174, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Zhao, B.; Peng, Y.; El-Badri, A.M.; Batool, M.; Wang, C.; Wang, Z.; Huang, W.; Wang, T.; Li, Z.; et al. Auxin plays a key role in nitrogen and plant density-modulated root growth and yield in different plant types of rapeseed. Field Crops Res. 2023, 302, 109066. [Google Scholar] [CrossRef]
- Han, X.-q.; Xiao, X.-y.; Guo, Z.-h.; Xie, Y.-h.; Zhu, H.-w.; Peng, C.; Liang, Y.-q. Release of cadmium in contaminated paddy soil amended with NPK fertilizer and lime under water management. Ecotoxicol. Environ. Saf. 2018, 159, 38–45. [Google Scholar] [CrossRef]
- You, Y.; Ju, C.; Wang, L.; Wang, X.; Ma, F.; Wang, G.; Wang, Y. The mechanism of arbuscular mycorrhizal enhancing cadmium uptake in Phragmites australis depends on the phosphorus concentration. J. Hazard. Mater. 2022, 440, 129800. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Wang, Y.; Yang, Z.; Yuan, J. Effects of phosphorus supplied in soil on subcellular distribution and chemical forms of cadmium in two Chinese flowering cabbage (Brassica parachinensis L.) cultivars differing in cadmium accumulation. Food Chem. Toxicol. 2011, 49, 2260–2267. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Wang, H.; Zhang, B.; He, Y.; Wang, H.; Zhu, Y.; Holm, P.E.; Shi, Y. Comparing cadmium uptake kinetics, xylem translocation, chemical forms, and subcellular distribution of two tobacco (Nicotiana tabacum L.) cultivars. Ecotoxicol. Environ. Saf. 2023, 254, 114738. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, J.; Zhang, Y.; Wang, H.; Wei, S.; Zhang, X.; Zhang, D.; Ma, H.; Ding, Q.; Ma, L. Morphophysiological, proteomic and metabolomic analyses reveal cadmium tolerance mechanism in common wheat (Triticum aestivum L.). J. Hazard. Mater. 2023, 445, 130499. [Google Scholar] [CrossRef]
- Grüter, R.; Costerousse, B.; Mayer, J.; Mäder, P.; Thonar, C.; Frossard, E.; Schulin, R.; Tandy, S. Long-term organic matter application reduces cadmium but not zinc concentrations in wheat. Sci. Total Environ. 2019, 669, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Zhu, Y.; Shao, K.; Zhang, Q.; Ye, G.; Shen, J. Metals source apportionment in farmland soil and the prediction of metal transfer in the soil-rice-human chain. J. Environ. Manag. 2020, 260, 110092. [Google Scholar] [CrossRef]
- Nishigaki, T.; Tsujimoto, Y.; Rinasoa, S.; Rakotoson, T.; Andriamananjara, A.; Razafimbelo, T. Phosphorus uptake of rice plants is affected by phosphorus forms and physicochemical properties of tropical weathered soils. Plant Soil 2019, 435, 27–38. [Google Scholar] [CrossRef]
- Chevallier, T.; Cournac, L.; Hamdi, S.; Gallali, T.; Bernoux, M. Temperature dependence of CO2 emissions rates and isotopic signature from a calcareous soil. J. Arid Environ. 2016, 135, 132–139. [Google Scholar] [CrossRef]
- Ma, S.; Nan, Z.; Hu, Y.; Chen, S.; Yang, X.; Su, J. Phosphorus supply level is more important than wheat variety in safe utilization of cadmium-contaminated calcareous soil. J. Hazard. Mater. 2022, 424, 127224. [Google Scholar] [CrossRef] [PubMed]
- Azzi, V.; Kanso, A.; Kazpard, V.; Kobeissi, A.; Lartiges, B.; El Samrani, A. Lactuca sativa growth in compacted and non-compacted semi-arid alkaline soil under phosphate fertilizer treatment and cadmium contamination. Soil Tillage Res. 2017, 165, 1–10. [Google Scholar] [CrossRef]
- Barchi, L.; Aprea, G.; Rabanus-Wallace, M.T.; Toppino, L.; Alonso, D.; Portis, E.; Lanteri, S.; Gaccione, L.; Omondi, E.; van Zonneveld, M.; et al. Analysis of >3400 worldwide eggplant accessions reveals two independent domestication events and multiple migration-diversification routes. Plant J. 2023, 116, 1667–1680. [Google Scholar] [CrossRef]
- Yuan, H.; Sun, L.; Tai, P.; Liu, W.; Li, X.; Hao, L. Effects of grafting on root-to-shoot cadmium translocation in plants of eggplant (Solanum melongena) and tomato (Solanum lycopersicum). Sci. Total Environ. 2019, 652, 989–995. [Google Scholar] [CrossRef]
- Cao, F.; Cai, Y.; Liu, L.; Zhang, M.; He, X.; Zhang, G.; Wu, F. Differences in photosynthesis, yield and grain cadmium accumulation as affected by exogenous cadmium and glutathione in the two rice genotypes. Plant Growth Regul. 2015, 75, 715–723. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Liang, L.; Lin, Z.; Su, X.; Zhang, W. Immobilization of cadmium in contaminated soils using sulfidated nanoscale zero-valent iron: Effectiveness and remediation mechanism. J. Hazard. Mater. 2021, 420, 126605. [Google Scholar] [CrossRef]
- Yu, S.; Sheng, L.; Mao, H.; Huang, X.; Luo, L.; Li, Y. Physiological response of Conyza Canadensis to cadmium stress monitored by Fourier transform infrared spectroscopy and cadmium accumulation. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 229, 118007. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, H.; He, D.; He, X.; Yan, Y.; Wu, K.; Wei, H. Effects of Exogenous Organic Acids on Cd Tolerance Mechanism of Salix variegata Franch. Under Cd Stress. Front. Plant Sci. 2020, 11, 594352. [Google Scholar] [CrossRef]
- Weigel, H.J.; Jäger, H.J. Subcellular Distribution and Chemical Form of Cadmium in Bean Plants. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef]
- Wu, F.-B.; Dong, J.; Qian, Q.Q.; Zhang, G.-P. Subcellular distribution and chemical form of Cd and Cd–Zn interaction in different barley genotypes. Chemosphere 2005, 60, 1437–1446. [Google Scholar] [CrossRef]
- Marchiol, L.; Assolari, S.; Sacco, P.; Zerbi, G. Phytoextraction of heavy metals by canola (Brassica napus) and radish (Raphanus sativus) grown on multicontaminated soil. Environ. Pollut. 2004, 132, 21–27. [Google Scholar] [CrossRef]
- Meng, Q.; Fan, W.; Liu, F.; Wang, G.; Di, X. Effect of Phosphorus Application on Eggplant Cadmium Accumulation and Soil Cadmium Morphology. Sustainability 2023, 15, 16236. [Google Scholar] [CrossRef]
- Santoro, V.; Schiavon, M.; Celi, L. Role of soil abiotic processes on phosphorus availability and plant responses with a focus on strigolactones in tomato plants. Plant Soil 2023, 194, 1–49. [Google Scholar] [CrossRef]
- Li, Z.; Guo, F.; Cornelis, J.-T.; Song, Z.; Wang, X.; Delvaux, B. Combined Silicon-Phosphorus Fertilization Affects the Biomass and Phytolith Stock of Rice Plants. Front. Plant Sci. 2020, 11, 67. [Google Scholar] [CrossRef]
- Dai, M.; Lu, H.; Liu, W.; Jia, H.; Hong, H.; Liu, J.; Yan, C. Phosphorus mediation of cadmium stress in two mangrove seedlings Avicennia marina and Kandelia obovata differing in cadmium accumulation. Ecotoxicol. Environ. Saf. 2017, 139, 272–279. [Google Scholar] [CrossRef]
- Jia, H.; Wu, Y.; Zhang, M.; Ye, J.; Du, D.; Wang, H. Role of phosphorus on the biogeochemical behavior of cadmium in the contaminated soil under leaching and pot experiments. J. Environ. Sci. 2024, 137, 488–499. [Google Scholar] [CrossRef]
- Wang, K.; Qiao, Y.; Zhang, H.; Yue, S.; Li, H.; Ji, X.; Liu, L. Influence of metal-contamination on distribution in subcellular fractions of the earthworm (Metaphire californica) from Hunan Province, China. J. Environ. Sci. 2018, 73, 127–137. [Google Scholar] [CrossRef]
- Wang, J.; Su, L.; Yang, J.; Yuan, J.; Yin, A.; Qiu, Q.; Zhang, K.; Yang, Z. Comparisons of cadmium subcellular distribution and chemical forms between low-Cd and high-Cd accumulation genotypes of watercress (Nasturtium officinale L. R. Br.). Plant Soil 2015, 396, 325–337. [Google Scholar] [CrossRef]
- Veach, A.M.; Yip, D.; Engle, N.L.; Yang, Z.K.; Bible, A.; Morrell-Falvey, J.; Tschaplinski, T.J.; Kalluri, U.C.; Schadt, C.W. Modification of plant cell wall chemistry impacts metabolome and microbiome composition in Populus PdKOR1 RNAi plants. Plant Soil 2018, 429, 349–361. [Google Scholar] [CrossRef]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal Interactions between Cadmium-Induced Cell Wall Responses and Oxidative Stress in Plants. Front. Plant Sci. 2017, 8, 286473. [Google Scholar] [CrossRef]
- Yu, H.; Guo, J.; Li, Q.; Zhang, X.; Huang, H.; Huang, F.; Yang, A.; Li, T. Characteristics of cadmium immobilization in the cell wall of root in a cadmium-safe rice line (Oryza sativa L.). Chemosphere 2020, 241, 125095. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Tenuzzo, B.A.; Carata, E.; Panzarini, E.; Luvisi, A.; De Bellis, L.; Vergine, M. Effects of Cadmium on Root Morpho-Physiology of Durum Wheat. Front. Plant Sci. 2022, 13, 936020. [Google Scholar] [CrossRef]
- Yang, L.P.; Zhu, J.; Wang, P.; Zeng, J.; Tan, R.; Yang, Y.Z.; Liu, Z.M. Effect of Cd on growth, physiological response, Cd subcellular distribution and chemical forms of Koelreuteria paniculata. Ecotoxicol. Environ. Saf. 2018, 160, 10–18. [Google Scholar] [CrossRef]
- Guo, J.; Ye, D.; Zhang, X.; Huang, H.; Wang, Y.; Zheng, Z.; Li, T.; Yu, H. Characterization of cadmium accumulation in the cell walls of leaves in a low-cadmium rice line and strengthening by foliar silicon application. Chemosphere 2022, 287, 132374. [Google Scholar] [CrossRef]
- Ma, P.; Zang, J.; Shao, T.; Jiang, Q.; Li, Y.; Zhang, W.; Liu, M. Cadmium distribution and transformation in leaf cells involved in detoxification and tolerance in barley. Ecotoxicol. Environ. Saf. 2023, 249, 114391. [Google Scholar] [CrossRef]
- Przybylska, A.; Wieczorek, P.; Obrępalska-Stęplowska, A. Meloidogyne arenaria candidate effector MaMsp4 interacts with maize (Zea mays L.) proteins involved in host defense response and cell wall modifications. Plant Soil 2023, 491, 501–523. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, C.; Wang, C.; Huang, Y.; Liu, Z. Increasing phosphate inhibits cadmium uptake in plants and promotes synthesis of amino acids in grains of rice. Environ. Pollut. 2020, 257, 113496. [Google Scholar] [CrossRef] [PubMed]
- Siebers, N.; Siangliw, M.; Tongcumpou, C. Cadmium uptake and subcellular distribution in rice plants as affected by phosphorus: Soil and hydroponic experiments. J. Soil Sci. Plant Nutr. 2013, 13, 833–844. [Google Scholar] [CrossRef]
- Sterckeman, T.; Thomine, S. Mechanisms of Cadmium Accumulation in Plants. Crit. Rev. Plant Sci. 2020, 39, 322–359. [Google Scholar] [CrossRef]
- Xu, Y.; Deng, G.; Guo, H.; Yang, M.; Yang, Q. Accumulation and sub cellular distribution of lead (Pb) in industrial hemp grown in Pb contaminated soil. Ind. Crops Prod. 2021, 161, 113220. [Google Scholar] [CrossRef]
- Gu, T.; Lu, Y.; Li, F.; Zeng, W.; Shen, L.; Yu, R.; Li, J. Microbial extracellular polymeric substances alleviate cadmium toxicity in rice (Oryza sativa L.) by regulating cadmium uptake, subcellular distribution and triggering the expression of stress-related genes. Ecotoxicol. Environ. Saf. 2023, 257, 114958. [Google Scholar] [CrossRef]
- Zheng, M.-M.; Feng, D.; Liu, H.-J.; Yang, G.-L. Subcellular distribution, chemical forms of cadmium and rhizosphere microbial community in the process of cadmium hyperaccumulation in duckweed. Sci. Total Environ. 2023, 859, 160389. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Zhou, Y.; Yang, Z.; Lin, B.; Yuan, J.; Wu, S. Comparisons in subcellular and biochemical behaviors of cadmium between low-Cd and high-Cd accumulation cultivars of pakchoi (Brassica chinensis L.). Front. Environ. Sci. Eng. 2014, 8, 226–238. [Google Scholar] [CrossRef]
- Thawornchaisit, U.; Polprasert, C. Evaluation of phosphate fertilizers for the stabilization of cadmium in highly contaminated soils. J. Hazard. Mater. 2009, 165, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, L.; Wang, Y.; Cao, Y.; Zhang, S. Synergistic enhanced passivation of phosphorus and cadmium in sediment by Ca/Al co-modified biochar. Chem. Eng. J. 2023, 474, 145539. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Liu, T.; Liu, B.; Huang, D.; Zhu, Q.; Xu, C. The influence of liming on cadmium accumulation in rice grains via iron-reducing bacteria. Sci. Total Environ. 2018, 645, 109–118. [Google Scholar] [CrossRef]
- He, L.-L.; Huang, D.-Y.; Zhang, Q.; Zhu, H.-H.; Xu, C.; Li, B.; Zhu, Q.-H. Meta-analysis of the effects of liming on soil pH and cadmium accumulation in crops. Ecotoxicol. Environ. Saf. 2021, 223, 112621. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).