
Glomus mongioiense, a New Species of Arbuscular Mycorrhizal Fungi from Italian Alps and the Phylogeny-Spoiling Issue of Ribosomal Variants in the Glomus Genus

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Single Species Pot Cultures
2.2. Morphological Analysis
2.3. Molecular Analysis
2.4. Phylogenetic Analyses
3. Results
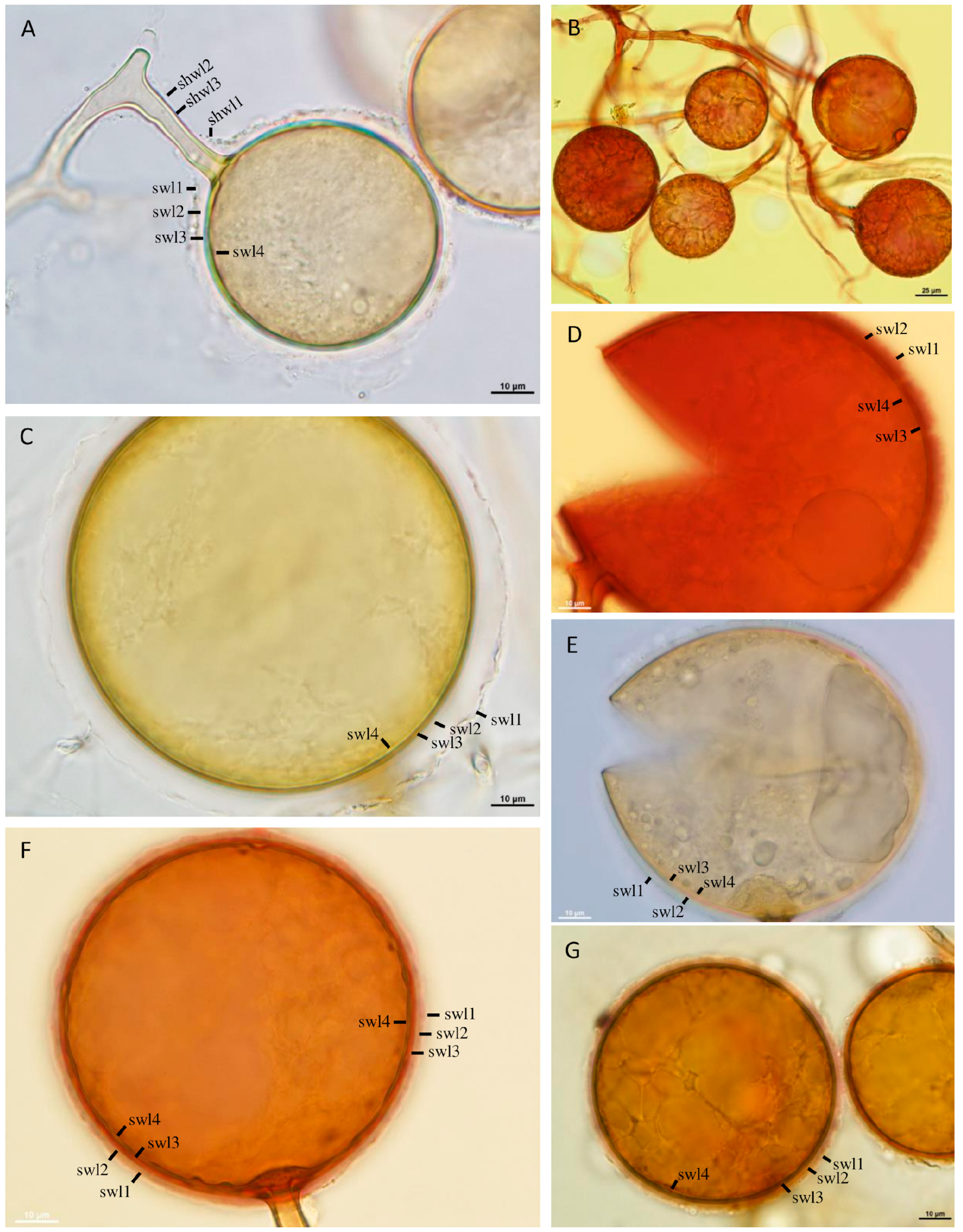
3.1. Taxonomy
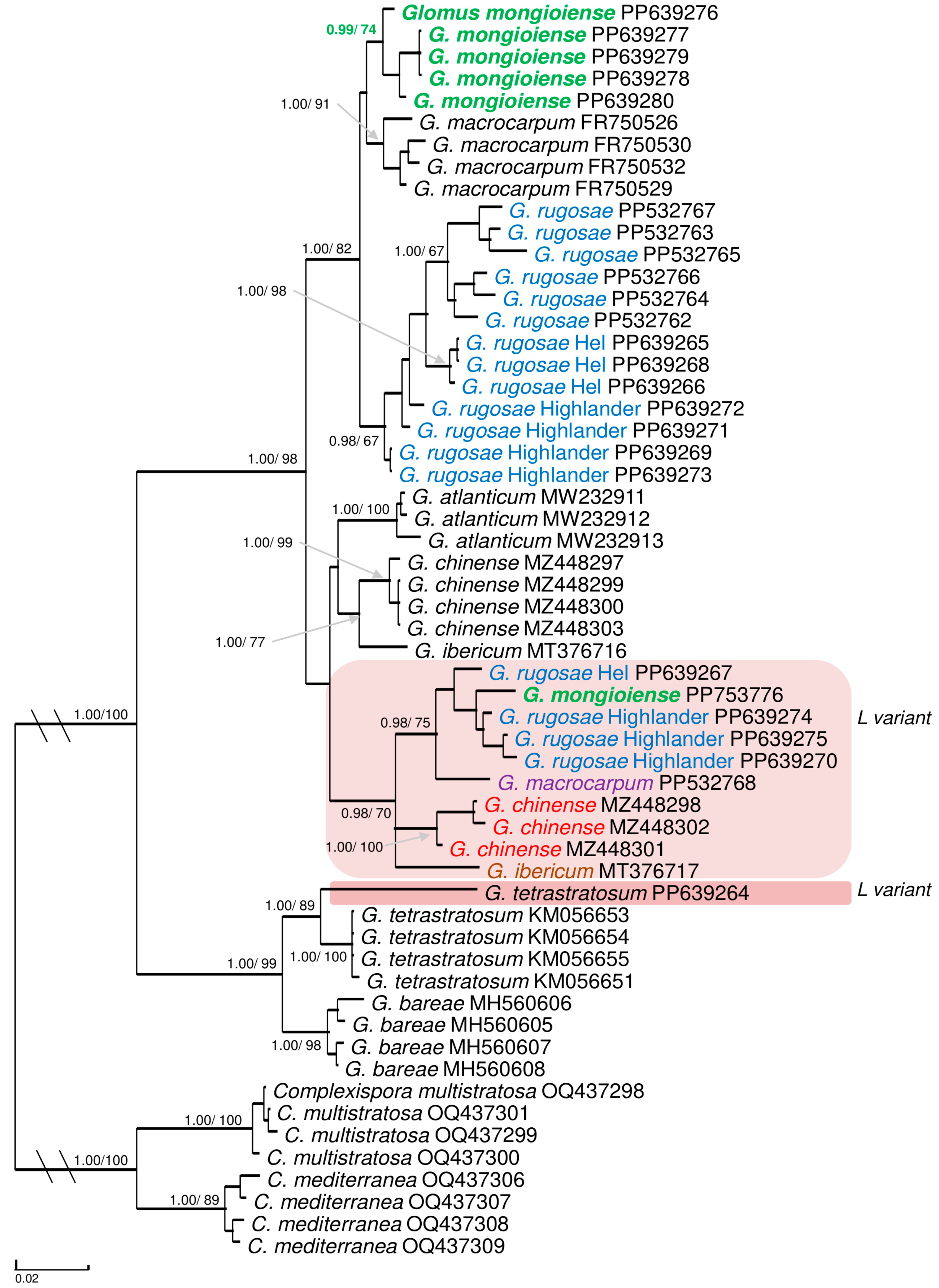
3.2. Molecular and Phylogenetic Analyses
4. Discussion
4.1. Comparison with Glomus Species
4.2. Ribosomal Variants: A “Forgotten” Issue
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilkes, T.I.; Warner, D.J.; Edmonds-Brown, V.; Davies, K.G.; Denholm, I. Zero Tillage Systems Conserve Arbuscular Mycorrhizal Fungi, Enhancing Soil Glomalin and Water Stable Aggregates with Implications for Soil Stability. Soil Syst. 2021, 5, 4. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of Arbuscular Mycorrhizal Fungi on Soil Fertility: Contribution in the Improvement of Physical, Chemical, and Biological Properties of the Soil. Front. Fungal Biol. 2022, 3, 723892. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xiao, Q.; Geng, X.; Lin, K.; Li, Z.; Li, Y.; Chen, J.; Li, X. Arbuscular mycorrhizal fungi alter rhizosphere bacterial diversity, network stability and function of lettuce in barren soil. Sci. Hortic. 2024, 323, 112533. [Google Scholar] [CrossRef]
- Pandit, A.; Kochar, M.; Srivastava, S.; Johny, L.; Adholeya, A. Diversity and Functionalities of Unknown Mycorrhizal Fungal Microbiota. Microbiol. Res. 2022, 256, 126940. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.J.; Demarmels, R.; Watts-Williams, S.J.; McLaughlin, M.J.; Kafle, A.; Ketelsen, C.; Soupir, A.; Bücking, H.; Cavagnaro, T.R.; van der Heijden, M.G.A. Global evaluation of commercial arbuscular mycorrhizal inoculants under greenhouse and field conditions. Appl. Soil Ecol. 2022, 169, 104225. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.; Dai, D.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Sánchez-García, M.; Niezgoda, P.; Zubek, S.; Fernández, F.; Vila, A.; Al-Yahya’ei, M.N.; Symanczik, S.; Milczarski, P.; Malinowski, R.; et al. A new order, Entrophosporales, and three new Entrophospora species in Glomeromycota. Front. Microbiol. 2022, 13, 962856. [Google Scholar] [CrossRef]
- Silva, G.A.; Corazon-Guivin, M.A.; de Assis, D.M.A.; Oehl, F. Blaszkowskia, a new genus in Glomeraceae. Mycol. Prog. 2023, 22, 74. [Google Scholar] [CrossRef]
- Tulasne, L.R.; Tulasne, C. Fungi nonnulli hipogaei, novi v. minus cognito act. Giorn. Bot. Ital. 1845, 2, 55–63. [Google Scholar]
- Tulasne, L.R.; Tulasne, C. Fungi Hypogaei. Histoire et Monographie des Champignons Hypogds; Legare Street Press: Paris, France, 1851. [Google Scholar]
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae of the Pacific Northwest. Micol. Mem. 1974, 5, 1–76. [Google Scholar]
- Schüßler, A.; Walker, C. The Glomeromycota: A Species List with New Families and New Genera; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Oehl, F.; Silva, G.A.; Goto, B.T.; Maia, L.C.; Sieverding, E. Glomeromycota: Two new classes and a new order. Mycotaxon 2011, 116, 365–379. [Google Scholar] [CrossRef]
- Oehl, F.; Silva, G.A.; Goto, B.T.; Sieverding, E. Glomeromycota: Three new genera and glomoid species reorganized. Mycotaxon 2011, 116, 75–120. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Chwat, G.; Góralska, A.; Ryszka, P.; Kovács, G.M. Two new genera, Dominikia and Kamienskia, and D. disticha sp. nov. in Glomeromycota. Nova Hedwig. 2015, 100, 225–238. [Google Scholar] [CrossRef]
- Sieverding, E.; Silva, G.A.; Reinhard, B.; Oehl, F. Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon 2015, 129, 373–386. [Google Scholar] [CrossRef]
- Berch, S.M.; Fortin, J.A. Lectotypification of Glomus macrocarpum and proposal of new combinations: Glomus australe, Glomus versiforme, and Glomus tenebrosum (Endogonaceae). Can. J. Bot. 1983, 61, 2608–2617. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; Goto, B.T.; Kozłowska, A. Halonatospora gen. nov. with H. pansihalos comb. nov. and Glomus bareae sp. nov. (Glomeromycota; Glomeraceae). Botany 2018, 96, 737–748. [Google Scholar] [CrossRef]
- Guillén, A.; Serrano-Tamay, F.J.; Peris, J.B.; Arrillaga, I. Glomus ibericum, Septoglomus mediterraneum, and Funneliformis pilosus, three new species of arbuscular mycorrhizal fungi. Mycologia 2020, 112, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Błaszkowski, J.; Niezgoda, P.; Zubek, S.; Meller, E.; Milczarski, P.; Malicka, M.; Goto, B.T.; Woźniak, G.; Moreira, H.; Magurno, F. Dominikia bonfanteae and Glomus atlanticum, two new species in the Glomeraceae (phylum Glomeromycota) with molecular phylogenies reconstructed from two unlinked loci. Mycol. Prog. 2021, 20, 131–148. [Google Scholar] [CrossRef]
- Yu, F.; Goto, B.T.; Magurno, F.; Błaszkowski, J.; Wang, J.; Ma, W.; Feng, H.; Liu, Y. Glomus chinense and Dominikia gansuensis, two new Glomeraceae species of arbuscular mycorrhizal fungi from high altitude in the Tibetan Plateau. Mycol. Prog. 2022, 21, 32. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Zubek, S.; Milczarski, P.; Malinowski, R.; Goto, B.T.; Niezgoda, P. Glomus rugosae, a new arbuscular mycorrhizal species in Glomeraceae (phylum Glomeromycota) from maritime sand dunes of Poland and an ash pond of Czech Republic. Phytotaxa 2024, 644, 271–280. [Google Scholar] [CrossRef]
- Fratianni, S.; Acquaotta, F. The climate of Italy. In Landscapes and Landforms of Italy; Soldati, M., Marchetti, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 29–38. [Google Scholar]
- Firth, C.; Fleet, L. Information Sheet on Ramsar Wetlands (RIS). 2006. Available online: http://www.ramsar.org/ris/key_ris_index.htm (accessed on 1 April 2024).
- Gunn, G.F. Measuring nocturnal near-surface urban heat island intensity in the small, mid-latitude city of Inverness, Scotland. Scott. Geogr. J. 2023, 140, 192–209. [Google Scholar] [CrossRef]
- Polderman, P.J.; Polderman-Hall, R.A. Algal communities in Scottish saltmarshes. Br. J. Psychol. 1980, 15, 59–71. [Google Scholar] [CrossRef]
- Woch, M.W.; Kapusta, P.; Stanek, M.; Możdżeń, K.; Grześ, I.M.; Rożej-Pabijan, E.; Stefanowicz, A.M. Effects of invasive Rosa rugosa on Baltic coastal dune communities depend on dune age. NeoBiota 2023, 82, 163–187. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen Publishing: London, UK, 1983. [Google Scholar]
- Furrazola, E.; Torres-Arias, Y.; Ferrer, R.L.; Herrera, R.A.; Berbara, R.; Goto, B.T. Glomus crenatum (Glomeromycetes), a new ornamented species from Cuba. Mycotaxon 2011, 116, 132–143. [Google Scholar] [CrossRef]
- Goto, B.T.; Jardim, J.G.; da Silva, G.A.; Furrazola, E.; Torres-Arias, Y.; Oehl, F. Glomus trufemii (Glomeromycetes), a new sporocarpic species from Brazilian sand dunes. Mycotaxon 2012, 120, 1–9. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Jobim, K.; Niezgoda, P.; Meller, E.; Malinowski, R.; Milczarski, P.; Zubek, S.; Magurno, F.; Casieri, L.; Bierza, W.; et al. New Glomeromycotan Taxa, Dominikia glomerocarpica sp. nov. and Epigeocarpum crypticum gen. nov. et sp. nov. From Brazil, and Silvaspora gen. nov. From New Caledonia. Front. Microbiol. 2021, 12, 655910. [Google Scholar] [CrossRef] [PubMed]
- Walker, C. Taxonomic concepts in the Endogonaceae: Spore wall characteristics in species descriptions. Mycotaxon 1983, 18, 443–455. [Google Scholar]
- Błaszkowski, J. Glomeromycota; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2012. [Google Scholar]
- Goto, B.T.; Maia, L.C. Glomerospores: A new denomination for the spore of Glomeromycota, a group molecularly distinct from the zygomycota. Mycotaxon 2006, 96, 129–132. [Google Scholar]
- Jobim, K.; Błaszkowski, J.; Niezgoda, P.; Kozłowska, A.; Zubek, S.; Mleczko, P.; Chachuła, P.; Ishikawa, N.K.; Goto, B.T. New sporocarpic taxa in the phylum Glomeromycota: Sclerocarpum amazonicum gen. et sp. nov. in the family Glomeraceae (glomerales) and Diversispora sporocarpia sp. nov. in the Diversisporaceae (Diversisporales). Mycol. Prog. 2019, 18, 369–384. [Google Scholar] [CrossRef]
- Krüger, M.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based species level detection of Glomeromycota: One PCR primer set for all arbuscular mycorrhizal fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef]
- Błaszkowski, J.; Yamato, M.; Niezgoda, P.; Zubek, S.; Milczarski, P.; Malinowski, R.; Meller, E.; Malicka, M.; Goto, B.T.; Uszok, S.; et al. A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycol. Prog. 2023, 22, 34. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Abadi, S.; Azouri, D.; Pupko, T.; Mayrose, I. Model selection may not be a mandatory step for phylogeny reconstruction. Nat. Commun. 2019, 10, 934. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.A.; Krompass, D.; Stamatakis, A. Performance, accuracy, and web server for evolutionary placement of short sequence reads under maximum likelihood. Syst. Biol. 2011, 60, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Barbera, P.; Stamatakis, A. Genesis and Gappa: Processing, analyzing and visualizing phylogenetic (placement) data. Bioinformatics 2020, 36, 3263–3265. [Google Scholar] [CrossRef] [PubMed]
- Malicka, M.; Magurno, F.; Piotrowska-Seget, Z. Phenol and Polyaromatic Hydrocarbons are Stronger Drivers Than Host Plant Species in Shaping the Arbuscular Mycorrhizal Fungal Component of the Mycorrhizosphere. Int. J. Mol. Sci. 2022, 23, 12585. [Google Scholar] [CrossRef] [PubMed]
- Dalpé, Y.; Plenchette, C.; Frenot, Y.; Gloaguen, J.C.; Strullu, D.G. Glomus kerguelense, a new Glomales species from sub-Antarctic. Mycotaxon 2002, 84, 51–60. [Google Scholar]
- Rodriguez, A.; Clapp, J.P.; Robinson, L.; Dodd, J.C. Studies on the diversity of the distinct phylogenetic lineage encompassing Glomus claroideum and Glomus etunicatum. Mycorrhiza 2005, 15, 33–46. [Google Scholar] [CrossRef]
- VanKuren, N.W.; den Bakker, H.C.; Morton, J.B.; Pawlowska, T.E. Ribosomal RNA gene diversity, effective population size, and evolutionary longevity in asexual Glomeromycota. Evolution 2013, 67, 207–224. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Yildirir, G.; Rizzi, Y.S.; Malar, M.C.; Nicol, A.M.; Sorwar, E.; Villeneuve-Laroche, M.; Chen, E.C.H.; Iwasaki, W.; Brauer, E.K.; et al. Arbuscular mycorrhizal fungi heterokaryons have two nuclear populations with distinct roles in host–plant interactions. Nat. Microbiol. 2023, 8, 2142–2153. [Google Scholar] [CrossRef] [PubMed]
- Yildirir, G.; Sperschneider, J.; Malar, M.C.; Chen, E.C.H.; Iwasaki, W.; Cornell, C.; Corradi, N. Long reads and Hi-C sequencing illuminate the two-compartment genome of the model arbuscular mycorrhizal symbiont Rhizophagus irregularis. New Phytol. 2022, 233, 1097–1107. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magurno, F.; Uszok, S.; Bierza, K.; Bakr, J.; Kende, Z.; Bessa de Queiroz, M.; Casieri, L. Glomus mongioiense, a New Species of Arbuscular Mycorrhizal Fungi from Italian Alps and the Phylogeny-Spoiling Issue of Ribosomal Variants in the Glomus Genus. Agronomy 2024, 14, 1350. https://doi.org/10.3390/agronomy14071350
Magurno F, Uszok S, Bierza K, Bakr J, Kende Z, Bessa de Queiroz M, Casieri L. Glomus mongioiense, a New Species of Arbuscular Mycorrhizal Fungi from Italian Alps and the Phylogeny-Spoiling Issue of Ribosomal Variants in the Glomus Genus. Agronomy. 2024; 14(7):1350. https://doi.org/10.3390/agronomy14071350
Chicago/Turabian StyleMagurno, Franco, Sylwia Uszok, Karolina Bierza, Jawdat Bakr, Zoltan Kende, Mariana Bessa de Queiroz, and Leonardo Casieri. 2024. "Glomus mongioiense, a New Species of Arbuscular Mycorrhizal Fungi from Italian Alps and the Phylogeny-Spoiling Issue of Ribosomal Variants in the Glomus Genus" Agronomy 14, no. 7: 1350. https://doi.org/10.3390/agronomy14071350