
Ruminant Grazing Lands in the Tropics: Silvopastoral Systems and Tithonia diversifolia as Tools with Potential to Promote Sustainability
 , ,
, ,
Abstract
:1. Introduction
2. Ruminant Production and Food Security—A Brief Overview
3. Silvopastoralism in Ruminant Production
4. Tithonia diversifolia and Sustainable Ruminant Production

{kind=link}
{kind=link}
{kind=link}
| Treatments | CH4 Production | Unit | NDF (%) | ADF (%) | A:P Ratio | Authors’ Discussion | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| TRT 1 | CON | TRT | CON | ||||||
| 10% TD with Lolium perenne | 29.3 | 30.5 | mL/gDM | 53.8 | 27.1 | 2.00 | 2.06 | No differences in CH4 and A:P ratio. | [103] |
| 20% TD with Lolium perenne | 25.9 | 52.1 | 28.9 | 1.95 | Decreased CH4 due to the presence of tannins. | ||||
| 33% TD with Pennisetum purpureum | 1.5 | 2.4 | mmol/g | 69.1 | 52.1 | 2.16 | 2.06 | No differences in CH4 and A:P ratio. | [104] |
| 75% TD with Pennisetum purpureum | 8.6 | 18.9 | mL/gDOM | 59.1 | 49.6 | 3.27 | 2.63 | Decreased CH4. Similar acetate and decreased propionate. | [105] |
| 75% TD with Cynodon dactylon | 4 | 7.3 | mL/gDOM | 61.3 | 48.3 | 3.41 | 3.04 | No differences in CH4. Similar propionate and increased acetate. | |
| TD extract with Commercial Concentrate (TMR) | 25.3 | 42.9 | mL/kgDOM | 30.1 | 21.4 | 1.39 | 1.99 | Decreased CH4 due to the presence of tannins. Similar propionate and decreased acetate. | [100] |
| TD extract with lucerne hay | 18.2 | 36.8 | 40.6 | 32.1 | 1.71 | 2.16 | Decreased CH4 due to the presence of tannins. Similar propionate and decreased acetate. | ||
| TD extract with Eragostis curvula | 5.8 | 47.7 | 78.4 | 49.2 | 1.63 | 2.48 | Decreased CH4 due to the presence of tannins. Increased propionate and decreased acetate. | ||
| 6.9% TD with sugarcane and concentrate | 0.7 | 0.5 | mL/gIDM | 29.4 | - | 0.90 | 0.71 | TD inclusion produced more CH4 due to increased A:P ratio. Increased acetate and decreased propionate. | [80] |
| 15.2% TD with sugarcane and concentrate | 1.2 | 30.7 | - | 1.09 | |||||
| 29.2% TD with sugarcane and concentrate | 3.3 | 34.5 | - | 1.55 | |||||
| 25% TD with Urochloa brizantha | ~22.9 | 26.2 | mg/gIDM | ~55.5 | ~38.3 | 2.37 | 3.56 | Decreased CH4 due to decreased A:P ratio. Decreased acetate and increased propionate. | [76] |
| 30% TD with Cynodon nlemfuensis | 0.9 | 6.5 | mL/100 mL | - | - | - | - | Decreased CH4 due to the presence of tannins. | [16] |
| 30% TD with Cynodon nlemfuensis | 9.2 | 65.2 | uL/gDM | 33.4 | 29.5 | - | - | Decreased CH4 due to the presence of tannins. | [106] |
| 30% de TD with Cynodon nlemfuensis | 47.2 | 35.3 | 30.4 | - | - | Decreased CH4. | |||
| 100% TD | 15.7 | 43.4 | mL/gDDM | 39 | 27.2 | - | - | Decreased CH4 due to the presence of tannins. | [99] |
| 5% TD with Cechrus clandestinum | 34.8 | 43.4 | - | - | 4.01 | 4.52 | Decreased CH4 due to the presence of tannins. Decreased acetate and increased propionate. | ||
| 3% TD with Cechrus clandestinum, concentrate and fat | 41 | 60.3 | - | - | 3.97 | 4.80 | Decreased CH4 due to the presence of tannins. Similar acetate and increased propionate. | ||
| Treatments | DMI 1 g/Day | NI g/Day | NF % NI | NU % NI | NR % NI | Authors’ Discussion | Reference |
|---|---|---|---|---|---|---|---|
| 0% TD extract + Cassava + concentrate | 378 | 6.98 | 39 | 39.8 | 21.2 | 80% TD resulted in decreased fecal N excretion and higher urinary N excretion. N retention was similar in all treatments except at 80%. | [111] |
| 20% TD extract + Cassava + concentrate | 374 | 6.92 | 31.4 | 46.7 | 22 | ||
| 40% TD extract + Cassava + concentrate | 371 | 6.87 | 29.7 | 46.4 | 23.9 | ||
| 80% TD extract + Cassava + concentrate | 318 | 5.88 | 25.9 | 56.3 | 17.9 | ||
| 0% TD + Dichanthium aristatum | 410 | 9.78 | 24.9 | 12.9 | 62.1 | TD inclusion resulted in decreased N excretion in feces and urine. N retention was higher with TD inclusion. | [114] |
| 25% TD + Dichanthium aristatum | 704 | 68.93 | 6.8 | 7.3 | 85.9 | ||
| 0% TD + Brachiaria decumbens | 652 | 63.23 | 48 | 35 | 16 | TD inclusion resulted in higher N excretion in feces and urine. N retention was higher with TD inclusion. | [88] |
| 35% TD + Brachiaria decumbens | 840 | 113.48 | 34 | 26 | 39 | ||
| 0% TD + Panicum maximum | 312 | 7.59 | 31.5 | 14.5 | 54 | No difference in fecal N excretion. Inclusions of 20% and 30% resulted in decreased urine N excretion. N retention was reduced by 20% and 30% TD diets. | [117] |
| 10% TD + Panicum maximum | 311 | 6.84 | 31.9 | 14.8 | 53.4 | ||
| 20% TD + Panicum maximum | 306 | 5.83 | 37 | 11.3 | 51.6 | ||
| 30% TD + Panicum maximum | 305 | 5.76 | 42 | 6.3 | 51.7 | ||
| TD 0% + sugarcane + concentrate | 1860 | 563 | 34.5 | 11 | 54.6 | No significant effects of TD inclusion on N excretion and nitrogen balance. | [11] |
| TD 6.5% + sugarcane + concentrate | 1890 | 564.1 | 35.6 | 10 | 54.4 | ||
| TD 15.4% + sugarcane + concentrate | 1870 | 557.2 | 35.5 | 11.4 | 53.1 | ||
| 0% TD + Pennisetum purpureum + sugarcane | 1050 | 13.05 | 50.73 | 39.2 | 9 | TD inclusion increased N excretion in feces and urine. Only at 20% inclusion was there positive N retention. Other inclusions were not significant. | [113] |
| 20% TD + Pennisetum purpureum + sugarcane | 1510 | 19.93 | 47.22 | 30 | 23.3 | ||
| 35% TD + Pennisetum purpureum + sugarcane | 1550 | 25.51 | 47.75 | 34.1 | 18.7 | ||
| 50% TD + Pennisetum purpureum + sugarcane | 1520 | 30.11 | 47.23 | 37.6 | 15.6 | ||
| 0% TD + Brachiaria + concentrate | 1471 | 66.4 | 25.1 | 55.4 | 19.5 | No effect of TD inclusion on fecal N excretion. Urinary N excretion was Decreased with 12% TD inclusion. | [112] |
| 6% TD + Brachiaria + concentrate | 1432 | 66.4 | 24.9 | 54.4 | 20.7 | ||
| 12% TD + Brachiaria + concentrate | 1452 | 66 | 27.1 | 43.7 | 29.2 | ||
| 0% TD + Panicum maximum | 1580 | 30.5 | - | - | 76 | TD inclusions led to increased N retention. | [115] |
| 25% TD + Panicum maximum | 1970 | 56.6 | - | - | 71.9 | ||
| 50% TD + Panicum maximum | 2070 | 64.4 | - | - | 75 | ||
| 75% TD + Panicum maximum | 2130 | 72.8 | - | - | 76.7 |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Boval, M.; Dixon, R.M. The Importance of Grasslands for Animal Production and Other Functions: A Review on Management and Methodological Progress in the Tropics. Animal 2012, 6, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Flachowsky, G.; Meyer, U.; Südekum, K.-H. Land Use for Edible Protein of Animal Origin—A Review. Animals 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock—A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations–FAO: Rome, Italy, 2013. [Google Scholar]
- Benchaar, C.; Greathead, H. Essential Oils and Opportunities to Mitigate Enteric Methane Emissions from Ruminants. Anim. Feed Sci. Technol. 2011, 166–167, 338–355. [Google Scholar] [CrossRef]
- Belanche, A.; Hristov, A.N.; van Lingen, H.J.; Denman, S.E.; Kebreab, E.; Schwarm, A.; Kreuzer, M.; Niu, M.; Eugène, M.; Niderkorn, V.; et al. Prediction of Enteric Methane Emissions by Sheep Using an Intercontinental Database. J. Clean. Prod. 2023, 384, 135523. [Google Scholar] [CrossRef]
- Eisler, M.C.; Lee, M.R.F.; Tarlton, J.F.; Martin, G.B.; Beddington, J.; Dungait, J.A.J.; Greathead, H.; Liu, J.; Mathew, S.; Miller, H.; et al. Agriculture: Steps to Sustainable Livestock. Nature 2014, 507, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E.; Dias de Oliveira, E.A.; Contin, D.R.; Delvecchio, G.; Viciedo, D.O.; de Moraes, M.A.; de Mello Prado, R.; de Pinho Costa, K.A.; Braga, M.R.; Martinez, C.A. Warming and Water Deficit Impact Leaf Photosynthesis and Decrease Forage Quality and Digestibility of a C4 Tropical Grass. Physiol. Plant. 2019, 165, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Pirhofer-Walzl, K.; Rasmussen, J.; Høgh-Jensen, H.; Eriksen, J.; Søegaard, K.; Rasmussen, J. Nitrogen Transfer from Forage Legumes to Nine Neighbouring Plants in a Multi-Species Grassland. Plant Soil 2012, 350, 71–84. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An International Terminology for Grazing Lands and Grazing Animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Jose, S.; Dollinger, J. Silvopasture: A Sustainable Livestock Production System. Agrofor. Syst. 2019, 93, 1–9. [Google Scholar] [CrossRef]
- Ribeiro, R.S.; Terry, S.A.; Sacramento, J.P.; Silveira, S.R.E.; Bento, C.B.P.; da Silva, E.F.; Mantovani, H.C.; Gama, M.A.S.D.; Pereira, L.G.R.; Tomich, T.R.; et al. Tithonia Diversifolia as a Supplementary Feed for Dairy Cows. PLoS ONE 2016, 11, e0165751. [Google Scholar] [CrossRef]
- Krüger, A.; Márquez, S.; Bizzuti, B.; Ovani, V.; Takahashi, L.; Santos, A.; Lima, P.; Maurício, R.; Abdalla, A. Tithonia Diversifolia and Tropical Grasses Intercropping as a Sustainable Alternative for Ruminants. In Proceedings of the International Symposium on Sustainable Animal Production and Health, Viena, Austria, 28 June–2 July 2021. [Google Scholar]
- Nziguheba, G.; Merckx, R.; Palm, C.A. Mutuo Combining Tithonia Diversifolia and Fertilizers for Maize Production in a Phosphorus Deficient Soil in Kenya. Agrofor. Syst. 2002, 55, 165–174. [Google Scholar] [CrossRef]
- Calsavara, L.H.F.; Ribeiro, R.S.; Silveira, S.R.; Delarota, G.; Freitas, D.S.; Sacramento, J.P.; Paciullo, D.S.C.; Maurício, R.M. Potential of Tithonia Diversifolia as Source of Forage for Ruminants. Livest. Res. Rural Dev. 2016, 28, 1–9. [Google Scholar]
- Mabou Tagne, A.; Marino, F.; Cosentino, M. Tithonia diversifolia (Hemsl.) A. Gray as a Medicinal Plant: A Comprehensive Review of Its Ethnopharmacology, Phytochemistry, Pharmacotoxicology and Clinical Relevance. J. Ethnopharmacol. 2018, 220, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Delgado, D.C.; Galindo, J.; González, R.; González, N.; Scull, I.; Dihigo, L.; Cairo, J.; Aldama, A.I.; Moreira, O. Feeding of Tropical Trees and Shrub Foliages as a Strategy to Reduce Ruminal Methanogenesis: Studies Conducted in Cuba. Trop. Anim. Health Prod. 2012, 44, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Rock, C.L.; Flatt, S.W.; Pakiz, B.; Taylor, K.S.; Leone, A.F.; Brelje, K.; Heath, D.D.; Quintana, E.L.; Sherwood, N.E. Weight Loss, Glycemic Control, and Cardiovascular Disease Risk Factors in Response to Differential Diet Composition in a Weight Loss Program in Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Care 2014, 37, 1573–1580. [Google Scholar] [CrossRef]
- Mottet, A.; de Haan, C.; Falcucci, A.; Tempio, G.; Opio, C.; Gerber, P. Livestock: On Our Plates or Eating at Our Table? A New Analysis of the Feed/Food Debate. Glob. Food Sec. 2017, 14, 1–8. [Google Scholar] [CrossRef]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The Rumen Microbiome: A Crucial Consideration When Optimising Milk and Meat Production and Nitrogen Utilisation Efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- McLaren, S.; Berardy, A.; Henderson, A.; Holden, N.; Huppertz, T.; Jolliet, O.; De Camillis, C.; Renouf, M.; Rugani, B.; Saarinen, M.; et al. Integration of Environment and Nutrition in Life Cycle Assessment of Food Items: Opportunities and Challenges; Food and Agriculture Organization of the United Nations–FAO: Rome, Italy, 2021; ISBN 978-92-5-135532-9. [Google Scholar]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed]
- Wyness, L. The Role of Red Meat in the Diet: Nutrition and Health Benefits. Proc. Nutr. Soc. 2016, 75, 227–232. [Google Scholar] [CrossRef]
- Givens, D.I. Review: Dairy Foods, Red Meat and Processed Meat in the Diet: Implications for Health at Key Life Stages. Animal 2018, 12, 1709–1721. [Google Scholar] [CrossRef]
- Cerri, C.C.; Moreira, C.S.; Alves, P.A.; Raucci, G.S.; de Almeida Castigioni, B.; Mello, F.F.C.; Cerri, D.G.P.; Cerri, C.E.P. Assessing the Carbon Footprint of Beef Cattle in Brazil: A Case Study with 22 Farms in the State of Mato Grosso. J. Clean. Prod. 2016, 112, 2593–2600. [Google Scholar] [CrossRef]
- McAuliffe, G.A.; Takahashi, T.; Orr, R.J.; Harris, P.; Lee, M.R.F. Distributions of Emissions Intensity for Individual Beef Cattle Reared on Pasture-Based Production Systems. J. Clean. Prod. 2018, 171, 1672–1680. [Google Scholar] [CrossRef]
- Sakita, G.Z.; Lima, P.M.T.; Abdalla Filho, A.L.; Bompadre, T.F.V.; Ovani, V.S.; Bizzuti, B.E.; da Costa, W.D.S.; do Prado Paim, T.; Campioni, T.S.; de Oliva Neto, P.; et al. Treating Tropical Grass with Fibrolytic Enzymes from the Fungus Trichoderma Reesei: Effects on Animal Performance, Digestibility and Enteric Methane Emissions of Growing Lambs. Anim. Feed Sci. Technol. 2022, 286, 115253. [Google Scholar] [CrossRef]
- Loeb, J. COP26: Is Change Afoot in Livestock Farming? Vet. Rec. 2021, 189, 348–351. [Google Scholar] [CrossRef]
- Eisen, M.B.; Brown, P.O. Rapid Global Phaseout of Animal Agriculture Has the Potential to Stabilize Greenhouse Gas Levels for 30 Years and Offset 68 Percent of CO2 Emissions This Century. PLoS Clim. 2022, 1, e0000010. [Google Scholar] [CrossRef]
- de Oliveira Silva, R.; Barioni, L.G.; Hall, J.A.J.; Folegatti Matsuura, M.; Zanett Albertini, T.; Fernandes, F.A.; Moran, D. Increasing Beef Production Could Lower Greenhouse Gas Emissions in Brazil If Decoupled from Deforestation. Nat. Clim. Chang. 2016, 6, 493–497. [Google Scholar] [CrossRef]
- Teague, W.R.; Apfelbaum, S.; Lal, R.; Kreuter, U.P.; Rowntree, J.; Davies, C.A.; Conser, R.; Rasmussen, M.; Hatfield, J.; Wang, T.; et al. The Role of Ruminants in Reducing Agriculture’s Carbon Footprint in North America. J. Soil Water Conserv. 2016, 71, 156–164. [Google Scholar] [CrossRef]
- Boval, M.; Angeon, V.; Rudel, T. Tropical Grasslands: A Pivotal Place for a More Multi-Functional Agriculture. Ambio 2017, 46, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; Parr, C.L. Beyond the Forest Edge: Ecology, Diversity and Conservation of the Grassy Biomes. Biol. Conserv. 2010, 143, 2395–2404. [Google Scholar] [CrossRef]
- Bezabih, M.; Pellikaan, W.F.; Tolera, A.; Khan, N.A.; Hendriks, W.H. Chemical Composition and in Vitro Total Gas and Methane Production of Forage Species from the M Id R Ift V Alley Grasslands of Ethiopia. Grass Forage Sci. 2014, 69, 635–643. [Google Scholar] [CrossRef]
- Lima, P.M.T.; Moreira, G.D.; Sakita, G.Z.; Natel, A.S.; Mattos, W.T.; Gimenes, F.M.A.; Gerdes, L.; McManus, C.; Abdalla, A.L.; Louvandini, H. Nutritional Evaluation of the Legume Macrotyloma Axillare Using In Vitro and in Vivo Bioassays in Sheep. J. Anim. Physiol. Anim. Nutr. 2018, 102, e669–e676. [Google Scholar] [CrossRef] [PubMed]
- Rao, I.M.; Peters, M.; Castro, A.; Schultze-Kraft, R.; White, D.; Fisher, M.; Miles, J.W.; Lascano Aguilar, C.E.; Blümmel, M.; Bungenstab, D.J.; et al. LivestockPlus—The Sustainable Intensification of Forage-Based Agricultural Systems to Improve Livelihoods and Ecosystem Services in the Tropics. Trop. Grassl. Forrajes Trop. 2015, 3, 59. [Google Scholar] [CrossRef]
- Teutscherová, N.; Vázquez, E.; Sotelo, M.; Villegas, D.; Velásquez, N.; Baquero, D.; Pulleman, M.; Arango, J. Intensive Short-Duration Rotational Grazing Is Associated with Improved Soil Quality within One Year after Establishment in Colombia. Appl. Soil Ecol. 2021, 159, 103835. [Google Scholar] [CrossRef]
- Rao, I.; Miles, J.; Wenzl, J.; Lowl-Gaume, A.; Cardoso, J.; Polania, J.; Rincón, J.; Hoyos, V.; Frossard, E.; Wagatsuma, T.; et al. Mechanisms of Adaptation of Brachiaria Grasses to Abiotic Stress Factors in the Tropics. In Proceedings of the III International Symposium on Forage Breeding, Bonito, Brazil, 7–11 November 2011. [Google Scholar]
- Navas Panadero, A.; Montaña, V. Comportamiento de Tithonia Diversifolia, Bajo Condiciones de Bosque Húmedo Tropical. Rev. Investig. Vet. del Perú 2019, 30, 721–732. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-Tool for Comprehensive Science Mapping Analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 February 2024).
- FAOSTAT. Land Use and Agricultural Area. Available online: https://www.fao.org/faostat/en/#country (accessed on 14 February 2024).
- Aria, M.; Alterisio, A.; Scandurra, A.; Pinelli, C.; D’Aniello, B. The Scholar’s Best Friend: Research Trends in Dog Cognitive and Behavioral Studies. Anim. Cogn. 2021, 24, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.F.; Maurício, R.M.; Moreira, G.R.; Gonçalves, L.C.; Borges, I.; Pereira, L.G.R. Nutritional Evaluation of “Braquiarão” Grass in Association with “Aroeira” Trees in a Silvopastoral System. Agrofor. Syst. 2010, 79, 189–199. [Google Scholar] [CrossRef]
- de Castro Santos, D.; Júnior, R.G.; Vilela, L.; Pulrolnik, K.; Bufon, V.B.; de Souza França, A.F. Forage Dry Mass Accumulation and Structural Characteristics of Piatã Grass in Silvopastoral Systems in the Brazilian Savannah. Agric. Ecosyst. Environ. 2016, 233, 16–24. [Google Scholar] [CrossRef]
- Gomes, F.J.; Cavalli, J.; Pedreira, B.C.; Pedreira, C.G.S.; Holschuch, S.G.; Pereira, D.H. Forage Nutritive Value of Marandu Palisade Grass under Clipping in a Silvopastoral System. Agrofor. Syst. 2022, 96, 79–88. [Google Scholar] [CrossRef]
- Paciullo, D.S.C.; De Carvalho, C.A.B.; Aroeira, L.J.M.; Morenz, M.J.F.; Lopes, F.C.F.; Rossiello, R.O.P. Morphophysiology and Nutritive Value of Signalgrass under Natural Shading and Full Sunlight. Pesqui. Agropecuária Bras. 2007, 42, 573–579. [Google Scholar] [CrossRef]
- Ainsworth, J.A.W.; Moe, S.R.; Skarpe, C. Pasture Shade and Farm Management Effects on Cow Productivity in the Tropics. Agric. Ecosyst. Environ. 2012, 155, 105–110. [Google Scholar] [CrossRef]
- Casanova-Lugo, F.; Villanueva-López, G.; Alcudia-Aguilar, A.; Nahed-Toral, J.; Medrano-Pérez, O.R.; Jiménez-Ferrer, G.; Alayón-Gamboa, J.A.; Aryal, D.R. Effect of Tree Shade on the Yield of Brachiaria Brizantha Grass in Tropical Livestock Production Systems in Mexico. Rangel. Ecol. Manag. 2022, 80, 31–38. [Google Scholar] [CrossRef]
- Paciullo, D.S.C.; Campos, N.R.; Gomide, C.A.M.; Castro, C.R.T.D.; Tavela, R.C.; Rossiello, R.O.P. Crescimento de Capim-Braquiária Influenciado Pelo Grau de Sombreamento e Pela Estação Do Ano. Pesqui. Agropecuária Bras. 2008, 43, 917–923. [Google Scholar] [CrossRef]
- Abraham, E.M.; Kyriazopoulos, A.P.; Parissi, Z.M.; Kostopoulou, P.; Karatassiou, M.; Anjalanidou, K.; Katsouta, C. Growth, Dry Matter Production, Phenotypic Plasticity, and Nutritive Value of Three Natural Populations of Dactylis Glomerata L. under Various Shading Treatments. Agrofor. Syst. 2014, 88, 287–299. [Google Scholar] [CrossRef]
- Gomes, F.J.; Pedreira, B.C.; Santos, P.M.; Bosi, C.; Lulu, J.; Pedreira, C.G.S. Microclimate Effects on Canopy Characteristics of Shaded Palisadegrass Pastures in a Silvopastoral System in the Amazon Biome of Central Brazil. Eur. J. Agron. 2020, 115, 126029. [Google Scholar] [CrossRef]
- Resende, L.D.O.; Müller, M.D.; Kohmann, M.M.; Pinto, L.F.G.; Cullen Junior, L.; de Zen, S.; Rego, L.F.G. Silvopastoral Management of Beef Cattle Production for Neutralizing the Environmental Impact of Enteric Methane Emission. Agrofor. Syst. 2020, 94, 893–903. [Google Scholar] [CrossRef]
- Filho, J.F.L.; de Oliveira, H.M.R.; de Souza Barros, V.M.; dos Santos, A.C.; de Oliveira, T.S. From Forest to Pastures and Silvopastoral Systems: Soil Carbon and Nitrogen Stocks Changes in Northeast Amazônia. Sci. Total Environ. 2024, 908, 168251. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.C.; Segura, M.; Andrade, H.J. Agroforestry Systems Affect Soil Organic Carbon Stocks and Fractions in Deforested Landscapes of Amazonia. Agrofor. Syst. 2024, 98, 1139–1151. [Google Scholar] [CrossRef]
- Archimède, H.; Eugène, M.; Marie Magdeleine, C.; Boval, M.; Martin, C.; Morgavi, D.P.; Lecomte, P.; Doreau, M. Comparison of Methane Production between C3 and C4 Grasses and Legumes. Anim. Feed Sci. Technol. 2011, 166–167, 59–64. [Google Scholar] [CrossRef]
- Melesse, A.; Steingass, H.; Schollenberger, M.; Holstein, J.; Rodehutscord, M. Nutrient Compositions and in Vitro Methane Production Profiles of Leaves and Whole Pods of Twelve Tropical Multipurpose Tree Species Cultivated in Ethiopia. Agrofor. Syst. 2019, 93, 135–147. [Google Scholar] [CrossRef]
- Bahamonde, H.A.; Peri, P.L.; Alvarez, R.; Barneix, A.; Moretto, A.; Martínez Pastur, G. Litter Decomposition and Nutrients Dynamics in Nothofagus Antarctica Forests under Silvopastoral Use in Southern Patagonia. Agrofor. Syst. 2012, 84, 345–360. [Google Scholar] [CrossRef]
- Sousa, L.F.; Maurício, R.M.; Paciullo, D.S.C.; Silveira, S.R.; Ribeiro, R.S.; Calsavara, L.H.; Moreira, G.R. Forage Intake, Feeding Behavior and Bio-Climatological Indices of Pasture Grass, under the Influence of Trees, in a Silvopastoral System. Trop. Grassl. Forrajes Trop. 2015, 3, 129. [Google Scholar] [CrossRef]
- Zambrano, E.R.A.; Edvan, R.L.; Oliveira, M.E.; Da Costa Araujo, D.L.; Costa, J.V.; Da Silva, I.R.; Barros, D.M.A.; De Sousa Reis, G.; Dias-Silva, T.P. Characterization of Pasture of Andropogon Grass and Behavior of Grazing Goats in a Silvopastoral System. Agrofor. Syst. 2021, 95, 1155–1165. [Google Scholar] [CrossRef]
- Ovani, V.S.; Pérez-Márquez, S.; da Silva, B.T.N.; Louvandini, H.; Abdalla, A.L.; de Azevedo Olival, A. Potential of Chloroleucon Acacioides Trees as an Alternative Feed Supplement for Grazing Ruminants in a Tropical Silvopastoral System. J. Agric. Food Res. 2023, 11, 100524. [Google Scholar] [CrossRef]
- Kilgour, R.J. In Pursuit of “Normal”: A Review of the Behaviour of Cattle at Pasture. Appl. Anim. Behav. Sci. 2012, 138, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, L.; Ominski, K.H.; He, M.; Xu, Z.; Krause, D.O.; Acharya, S.N.; Wittenberg, K.M.; Liu, X.L.; Stanford, K.; et al. Screening of Condensed Tannins from Canadian Prairie Forages for Anti–Escherichia Coli O157:H7 with an Emphasis on Purple Prairie Clover (Dalea purpurea Vent). J. Food Prot. 2013, 76, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Hoste, H.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Mueller-Harvey, I.; Sotiraki, S.; Louvandini, H.; Thamsborg, S.M.; Terrill, T.H. Tannin Containing Legumes as a Model for Nutraceuticals against Digestive Parasites in Livestock. Vet. Parasitol. 2015, 212, 5–17. [Google Scholar] [CrossRef] [PubMed]
- MacAdam, J.; Villalba, J. Beneficial Effects of Temperate Forage Legumes That Contain Condensed Tannins. Agriculture 2015, 5, 475–491. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional Aspects of Ecologically Relevant Phytochemicals in Ruminant Production. Front. Vet. Sci. 2021, 8, 155. [Google Scholar] [CrossRef]
- Albores-Moreno, S.; Alayón-Gamboa, J.A.; Miranda-Romero, L.A.; Alarcón-Zúñiga, B.; Jiménez-Ferrer, G.; Ku-Vera, J.C.; Piñeiro-Vázquez, A.T. Effect of Tree Foliage Supplementation of Tropical Grass Diet on in Vitro Digestibility and Fermentation, Microbial Biomass Synthesis and Enteric Methane Production in Ruminants. Trop. Anim. Health Prod. 2019, 51, 893–904. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Morsy, A.S.; Sallam, S.M.A.; Lucas, R.C.; Louvandini, H.; Kreuzer, M.; Abdalla, A.L. Contribution of Condensed Tannins and Mimosine to the Methane Mitigation Caused by Feeding Leucaena leucocephala. Arch. Anim. Nutr. 2013, 67, 169–184. [Google Scholar] [CrossRef]
- Pachas, N.A.; Radrizzani, A.; Murgueitio, E.; Uribe, F.; Zapata Cadavid, Á.; Chará, J.; Ruiz, T.E.; Escalante, E.; Mauricio, R.M.; Ramírez-Avilés, L. Establishment and Management of Leucaena in Latin America. Trop. Grassl. Forrajes Trop. 2019, 7, 127–132. [Google Scholar] [CrossRef]
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the Abatement of Methane and Nitrous Oxide from Ruminant Production: A Review. Livest. Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Mueller-Harvey, I.; Bee, G.; Dohme-Meier, F.; Hoste, H.; Karonen, M.; Kölliker, R.; Lüscher, A.; Niderkorn, V.; Pellikaan, W.F.; Salminen, J.-P.; et al. Benefits of Condensed Tannins in Forage Legumes Fed to Ruminants: Importance of Structure, Concentration, and Diet Composition. Crop Sci. 2019, 59, 861–885. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen Fixation in Annual and Perennial Legume-Grass Mixtures across a Fertility Gradient. Plant Soil 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Delgado, J.A.; Groffman, P.M.; Nearing, M.A.; Goddard, T.; Reicosky, D.; Lal, R.; Kitchen, N.R.; Rice, C.W.; Towery, D.; Salon, P. Conservation Practices to Mitigate and Adapt to Climate Change. J. Soil Water Conserv. 2011, 66, 118A–129A. [Google Scholar] [CrossRef]
- Mauricio, R.; Ribeiro, R.; Paciullo, D.; Cangussú, M.; Murgueitio, E.; Chará, J.; Estrada, M. Silvopastoral Systems in Latin America for Biodiversity Environmental and Socioeconomic Improvements; Elsevier: Amsterdam, Netherlands, 2019; ISBN 9780128110508. [Google Scholar]
- Olival, A.D.A.; Souza, S.E.X.F.D.; Leles, G.M. Effects of Native Tree Species on Forage Quality and Availability in the Mato-Grossense Amazon. Rev. Ibero-Americana Ciências Ambient. 2021, 12, 146–156. [Google Scholar] [CrossRef]
- Rivera, J.; Chará, J.; Arango, J.; Barahona, R. Effect of Different Genotypes of Tithonia diversifolia on Fermentation of Feed Mixtures with Urochloa Brizantha Cv. Marandú. Crop Pasture Sci. 2021, 72, 850–859. [Google Scholar] [CrossRef]
- Van Sao, N.; Mui, N.T.; Van Binh, D. Biomass Production of Tithonia diversifolia (Wild Sunflower), Soil Improvement on Sloping Land and Use as High Protein Foliage for Feeding Goats. Livest. Res. Rural Dev. 2010, 22, 1–7. [Google Scholar]
- García, I.R. Potencialidades de Tithonia diversifolia (Hemsl.) Gray En La Alimentación Animal. Livest. Res. Rural Dev. 2017, 29, 1–11. [Google Scholar]
- Mahecha, L.; Rosales, M. Valor Nutricional Del Follaje de Botón de Oro (Tithonia diversifolia [Hemsl] Gray) En La Producción Animal En El Trópico. Livest. Res. Rural Dev. 2005, 17, 1–9. [Google Scholar]
- Terry, S.A.; Ribeiro, R.S.; Freitas, D.S.; Delarota, G.D.; Pereira, L.G.R.; Tomich, T.R.; Maurício, R.M.; Chaves, A.V. Effects of Tithonia diversifolia on in Vitro Methane Production and Ruminal Fermentation Characteristics. Anim. Prod. Sci. 2016, 56, 437. [Google Scholar] [CrossRef]
- Pérez-Márquez, S.; Ovani, V.S.; Lima, P.D.M.T.; Lana, Â.M.Q.; Louvandini, H.; Abdalla, A.L.; Maurício, R.M. Tithonia diversifolia Improves In Vitro Rumen Microbial Synthesis of Sheep Diets without Changes in Total Gas and Methane Production. Agronomy 2023, 13, 2768. [Google Scholar] [CrossRef]
- Partey, S.T. Effect of Pruning Frequency and Pruning Height on the Biomass Production of Tithonia diversifolia (Hemsl) A. Gray. Agrofor. Syst. 2011, 83, 181–187. [Google Scholar] [CrossRef]
- Chagas-Paula, D.A.; Oliveira, R.B.; Rocha, B.A.; Da Costa, F.B. Ethnobotany, Chemistry, and Biological Activities of the Genus Tithonia (Asteraceae). Chem. Biodivers. 2012, 9, 210–235. [Google Scholar] [CrossRef]
- Mauricio, R.M. Feeding Ruminants Using Tithonia diversifolia as Forage. J. Dairy, Vet. Anim. Res. 2017, 5, 00146. [Google Scholar] [CrossRef]
- Ovani, V.; Pérez-Márquez, S.; Evangelista, V.; Louvandini, H.; Abdalla, A. Root and Biomass Production of Tithonia diversifolia in Acidic Soil Levels. In Proceedings of the 57o Reunião da Sociedade Brasileira de Zootecnia—Tropical Animal Science and Practice to Feed the Planet, Campinas, Brazil, 25–29 July 2022. [Google Scholar]
- Argüello-Rangel, J.; Mahecha-Ledesma, L.; Angulo-Arizala, J. Perfil Nutricional y Productivo de Especies Arbustivas En Trópico Bajo, Antioquia (Colombia). Cienc. Tecnol. Agropecu. 2020, 21, 1–20. [Google Scholar] [CrossRef]
- Chin, N.; Hue, K. Supplementing Tithonia diversifolia with Guinea Grass or Tree Foliages: Effects on Feed Intake and Live Weight Gain of Growing Goats. Livest. Res. Rural Dev. 2012, 24, 188. [Google Scholar]
- Durango, S.G.; Barahona, R.; Bolívar, D.; Chirinda, N.; Arango, J. Feeding Strategies to Increase Nitrogen Retention and Improve Rumen Fermentation and Rumen Microbial Population in Beef Steers Fed with Tropical Forages. Sustainability 2021, 13, 10312. [Google Scholar] [CrossRef]
- Guatusmal-Gelpud, C.; Escobar-Pachajoa, L.D.; Meneses-Buitrago, D.H.; Cardona-Iglesias, J.L.; Castro-Rincón, E. Producción y Calidad de Tithonia diversifolia y Sambucus Nigra En Trópico Altoandino Colombiano. Agron. Mesoam. 2020, 31, 193–208. [Google Scholar] [CrossRef]
- Lezcano, Y.; Soca, M.; Ojeda, F.; Roque, E.; Fontes, D.; Montejo, I.; Santana, H.; Martínez, J.; Cubillas, N. Caracterización Bromatológica de Tithonia diversifolia (Hemsl) A Gray En Dosetapas de Dos Etapas de Su Ciclo Fisiológico. Pastos y Forrajes 2012, 35, 275–282. [Google Scholar]
- Londoño, J.M.B.; Carabalí, A.G.; Londoño, M.A.B. Yield, Agronomic Parameters and Nutritional Quality of Tithonia diversifolia in Response to Different Fertilization Levels. Rev. Mex. Ciencias Pecu. 2019, 10, 789–800. [Google Scholar] [CrossRef]
- Mahecha, L.; Londoño, J.D.; Angulo, J. Agronomic and Nutritional Assessment of an Intensive Silvopastoral System: Tithonia diversifolia, Sambucus nigra, Cynodon nlemfuensis, and Urochloa plantaginea. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2022, 92, 37–47. [Google Scholar] [CrossRef]
- Naranjo, J.; Cuartas, C. Nutritional Characterization and Ruminal Degradation Kinetics of Some Forages with Potential for Ruminants Supplementation in the Highland Tropics of Colombia. CES Med. Zootec 2005, 6, 9–19. [Google Scholar]
- Verdecia, D.; Ramirez, J.; Leonard, I.; Alvarez, Y.; Bazán, Y.; Bodas, R.; Andrés, S.; Alvarez, J.; Giraldez, F.; Lopez, S. Nutritive Value of the Tithonia diversifolia in a Location of Valle Del Cauto. Rev. electrón. Vet 2011, 12, 1–13. [Google Scholar]
- Andries, J.I.; Buysse, F.X.; De Brabander, D.L.; Cottyn, B.G. Isoacids in Ruminant Nutrition: Their Role in Ruminal and Intermediary Metabolism and Possible Influences on Performances—A Review. Anim. Feed Sci. Technol. 1987, 18, 169–180. [Google Scholar] [CrossRef]
- Zebeli, Q.; Terrill, S.J.; Mazzolari, A.; Dunn, S.M.; Yang, W.Z.; Ametaj, B.N. Intraruminal Administration of Megasphaera elsdenii Modulated Rumen Fermentation Profile in Mid-Lactation Dairy Cows. J. Dairy Res. 2012, 79, 16–25. [Google Scholar] [CrossRef]
- Sampaio, C.B.; Detmann, E.; Paulino, M.F.; Valadares Filho, S.C.; de Souza, M.A.; Lazzarini, I.; Rodrigues Paulino, P.V.; de Queiroz, A.C. Intake and Digestibility in Cattle Fed Low-Quality Tropical Forage and Supplemented with Nitrogenous Compounds. Trop. Anim. Health Prod. 2010, 42, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Tambara, A.A.C.; Härter, C.J.; Rabelo, C.H.S.; Kozloski, G.V. Effects of Supplementation on Production of Beef Cattle Grazing Tropical Pastures in Brazil during the Wet and Dry Seasons: A Meta-Analysis. Rev. Bras. Zootec. 2021, 50, 1–22. [Google Scholar] [CrossRef]
- Cardona Iglesias, J.L.; Mahecha Ledesma, L.; Angulo Arizala, J. Efecto Sobre La Fermentación in Vitro de Mezclas de Tithonia Diversifolia, Cenchrus Clandestinum y Grasas Poliinsaturadas. Agron. Mesoam. 2017, 28, 405. [Google Scholar] [CrossRef]
- Akanmu, A.M.; Hassen, A.; Adejoro, F.A. Gas Production, Digestibility and Efficacy of Stored or Fresh Plant Extracts to Reduce Methane Production on Different Substrates. Animals 2020, 10, 146. [Google Scholar] [CrossRef]
- Folley, S.J.; French, T.H. Acetate as a Possible Precursor of Ruminant Milk Fat, Particularly the Short-Chain Fatty Acids. Nature 1949, 163, 174–175. [Google Scholar] [CrossRef]
- Lin, M.; Jiang, M.; Yang, T.; Tan, D.; Hu, G.; Zhao, G.; Zhan, K. Acetate-Induced Milk Fat Synthesis Is Associated with Activation of the MTOR Signaling Pathway in Bovine Mammary Epithelial Cells. Animals 2022, 12, 2616. [Google Scholar] [CrossRef]
- Rivera, J.E.; Villegas, G.; Chará, J.; Durango, S.G.; Romera, M.A.; Verchot, L. Effect of Tithonia diversifolia (Hemsl.) A. Gray intake on in vivo methane (CH4) emission and milk production in dual purpose cows in the Colombian Amazonian piedmont. Transl. Anim. Sci. 2022, 6, 1–12. [Google Scholar] [CrossRef]
- Herrera, J.P.N.; Preston, T.R.; Edmundo, J.; Guerrero, A.; Riascos, R. Producción de Metano En Una Incubación in Vitro de Cenchrus Clandestinus y Lolium Hybridum Suplementado Con Tithonia diversifolia En El Trópico Alto Del Departamento Del Putumayo, Colombia. Livest. Res. Rural Dev. 2020, 32, 1–6. [Google Scholar]
- Holguín, V.A.; Cuchillo-Hilario, M.; Mazabel, J.; Quintero, S.; Mora-Delgado, J. Efecto de La Mezcla Ensilada de Penisetum Purpureum y Tithonia Diversifolia Sobre La Fermentación Ruminal in Vitro y Su Emisión de Metano en el Sistema RUSITEC. Rev. Mex. Ciencias Pecu. 2020, 11, 19–37. [Google Scholar] [CrossRef]
- Pérez-Márquez, S.; Ribeiro, R.S.; Abdalla, A.L.; Lana, A.M.Q.; Maurício, R.M. Does Tithonia diversifolia Influences in Vitro Gas Production Parameters and Nutritive Value of Graminous Cynodon Spp and Penisetum purpurem When Associated? In Proceedings of the X Congreso Internacional Sobre Sistemas Silvopastoriles: Por una Producción Sostenible; CIPAV Editorial Asunción: Asunción, Paraguay, 2019; pp. 493–495. [Google Scholar]
- Olayinka, B.; Raiyemo, D.; Etejere, E. Phytochemical and Proximate Composition of Tithonia diversifolia (Hemsl) A Gray. Ann. Food Sci. Technol. 2015, 16, 195–200. [Google Scholar]
- Moreira, G.D.; Lima, P.D.M.T.; Borges, B.O.; Primavesi, O.; Longo, C.; McManus, C.; Abdalla, A.; Louvandini, H. Tropical Tanniniferous Legumes Used as an Option to Mitigate Sheep Enteric Methane Emission. Trop. Anim. Health Prod. 2013, 45, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Makkar, H.P.S. Methane Mitigation from Ruminants Using Tannins and Saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.D.M.T.; Filho, A.L.A.; Issakowicz, J.; Ieda, E.H.; Corrêa, P.S.; de Mattos, W.T.; Gerdes, L.; McManus, C.; Abdalla, A.L.; Louvandini, H. Methane Emission, Ruminal Fermentation Parameters and Fatty Acid Profile of Meat in Santa Inês Lambs Fed the Legume Macrotiloma. Anim. Prod. Sci. 2020, 60, 665. [Google Scholar] [CrossRef]
- Yousuf, B.M.; Adefunmilayo Adeloye, A.; kehinde Okukpe, M.; Dauda Adeyemi, K.; Julius Ogundun, N. Influence of Dietary Sunflower (Tithonia diversifolia) Leaf Extracts on Performance Characteristics of Goats Fed Cassava Peeling Wastes-Based Diet. J. Agric. Technol. 2014, 10, 59–65. [Google Scholar]
- Chacón Góngora, P.A. Uso de Tithonia Diversifolia Como Forraje Alternativo Para La Reducción de Emisiones de Óxido Nitroso En Excretas de Vacas. CaTIE: Turrialba, Costa Rica, 2018. [Google Scholar]
- Ramírez-Rivera, U.; Sanginés-García, J.R.; Escobedo-Mex, J.G.; Cen-Chuc, F.; Rivera-Lorca, J.A.; Lara-Lara, P.E. Effect of Diet Inclusion of Tithonia diversifolia on Feed Intake, Digestibility and Nitrogen Balance in Tropical Sheep. Agrofor. Syst. 2010, 80, 295–302. [Google Scholar] [CrossRef]
- Castañeda Serrano, R.D.; Piñeros Varón, R.; Vélez Giraldo, A. Foliage of Tropical Arboreal Species in Feeding Ovines (Ovis aries): Intake, Digestibility and Balance Nitrogen. Boletín Científico Cent. Museos Mus. Hist. Nat. 2018, 22, 58–68. [Google Scholar] [CrossRef]
- Fajemisin, A.N.; Salihu, T.; Fadiyimu, A.; Alokan, A. Dietary Effect of Substituting Panicum Maximum with Tithonia Diversifolia Forage on Performance of Yankasa Sheep. In Proceedings of the XXII International Grassland Congress, Sidney, Australia, 15–19 September 2019; pp. 563–564. [Google Scholar]
- Rivera, J.E.; Villegas, G.; Chará, J.; Durango, S.; Romero, M.; Verchot, L. Silvopastoral Systems with Tithonia diversifolia (Hemsl.) A. Gray Reduce N2O–N and CH4 Emissions from Cattle Manure Deposited on Grasslands in the Amazon Piedmont. Agrofor. Syst. 2023, 98, 1091–1104. [Google Scholar] [CrossRef]
- Odedire, J.; Oloidi, F. Feeding Wild Sunflower (Tithonia diversifolia Hemsl., A. Gray) to West African Dwarf Goats as a Dry Season Forage Supplement. World J. Agric. Res. 2014, 2, 280–284. [Google Scholar] [CrossRef]
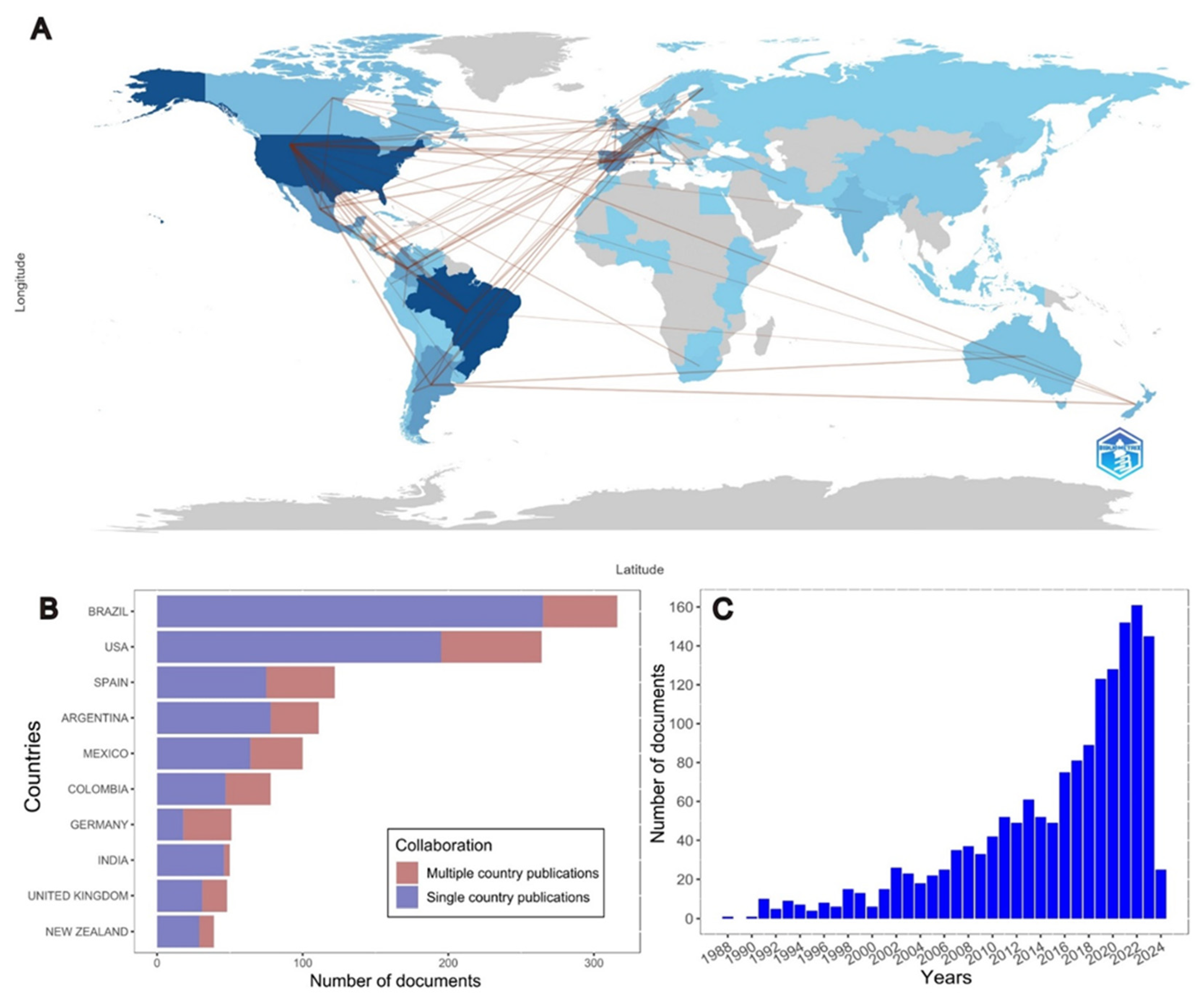

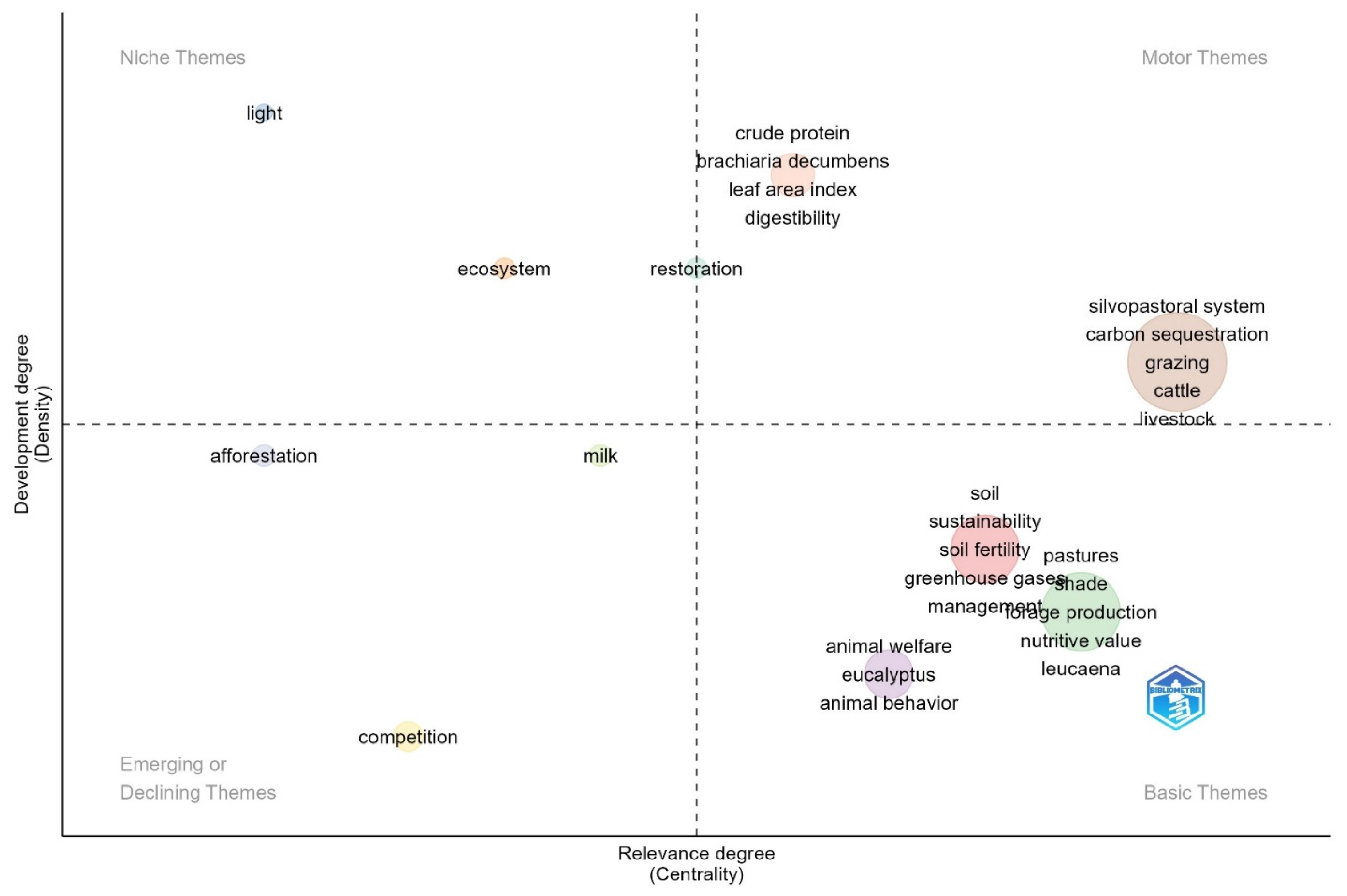
| References | DM | CP 1 | NDF | ADF | Obs. |
|---|---|---|---|---|---|
| Argüello-Rangel et al. [86] | 190 | 252 | 337 | 145 | |
| Calsavara et al. [14] | 200 | 165 | 476 | 333 | Whole plant |
| Calsavara et al. [14] | 195 | 225 | 410 | 261 | Leaves |
| Chin and Hue [87] | 146 | 239 | 384 | n/a | |
| Durango et al. [88] | 212 | 185 | 462 | 343 | |
| Guatusmal-Gelpud et al. [89] | n/a | 267 | 331 | 150 | |
| Lezcano et al. [90] | 101 | 219 | n/a | n/a | Rainy season |
| Lezcano et al. [90] | 127 | 190 | n/a | n/a | Dry season |
| Londoño et al. [91] | 185 | 273 | 268 | 169 | No fertilization |
| Mahecha and Rosales [79] | 172 | 242 | 253 | 304 | |
| Mahecha et al. [92] | n/a | 223 | 359 | 181 | |
| Naranjo and Cuartas [93] | 191 | 241 | 386 | 345 | |
| Van Sao et al. [77] | 146 | 239 | 384 | n/a | |
| Verdecia et al. [94] | 198 | 289 | 436 | 276 | Rainy season |
| Verdecia et al. [94] | 182 | 275 | 404 | 241 | Dry season |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krüger, A.M.; Lima, P.d.M.T.; Ovani, V.; Pérez-Marquéz, S.; Louvandini, H.; Abdalla, A.L. Ruminant Grazing Lands in the Tropics: Silvopastoral Systems and Tithonia diversifolia as Tools with Potential to Promote Sustainability. Agronomy 2024, 14, 1386. https://doi.org/10.3390/agronomy14071386
Krüger AM, Lima PdMT, Ovani V, Pérez-Marquéz S, Louvandini H, Abdalla AL. Ruminant Grazing Lands in the Tropics: Silvopastoral Systems and Tithonia diversifolia as Tools with Potential to Promote Sustainability. Agronomy. 2024; 14(7):1386. https://doi.org/10.3390/agronomy14071386
Chicago/Turabian StyleKrüger, Ana Maria, Paulo de Mello Tavares Lima, Vagner Ovani, Simón Pérez-Marquéz, Helder Louvandini, and Adibe Luiz Abdalla. 2024. "Ruminant Grazing Lands in the Tropics: Silvopastoral Systems and Tithonia diversifolia as Tools with Potential to Promote Sustainability" Agronomy 14, no. 7: 1386. https://doi.org/10.3390/agronomy14071386