
The TIR1/AFB Family in Solanum melongena: Genome-Wide Identification and Expression Profiling under Stresses and Picloram Treatment

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of the TIR1/AFB Gene Family
2.2. Comparative Phylogenetic Analysis and Classification of the S. melongena TIR1/AFB Genes
2.3. Identification of Duplicated TIR1/AFB Genes in S. melongena
2.4. Promoter cis-Elements of the S. melongena TIR1/AFB Genes
2.5. 3D Protein Structure Prediction
2.6. Gene Structures, Conserved Domains, and Motif Analysis
2.7. Prediction of miRNAs and Target Genes
2.8. RNA Sequencing and Data Analysis
2.8.1. Plant Growth and Sample Collection
2.8.2. Stress Treatment
2.8.3. RNA-Seq Library Construction and Sequencing
2.9. Validation of Selected Gene Expression by Real-Time Quantitative Reverse Transcription PCR (qRT–PCR)
3. Results
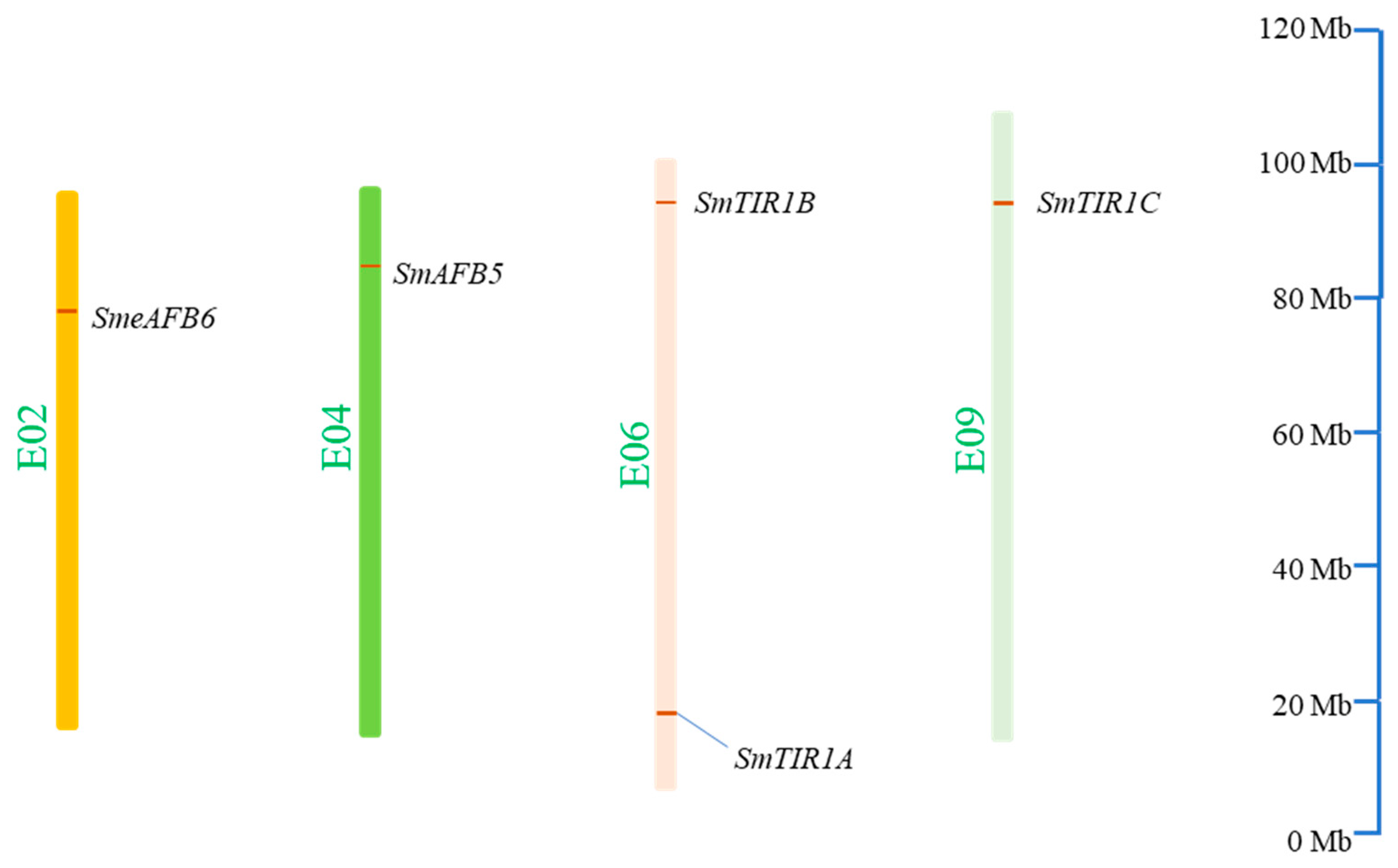
3.1. Identification of TIR1/AFB Genes in S. melongena
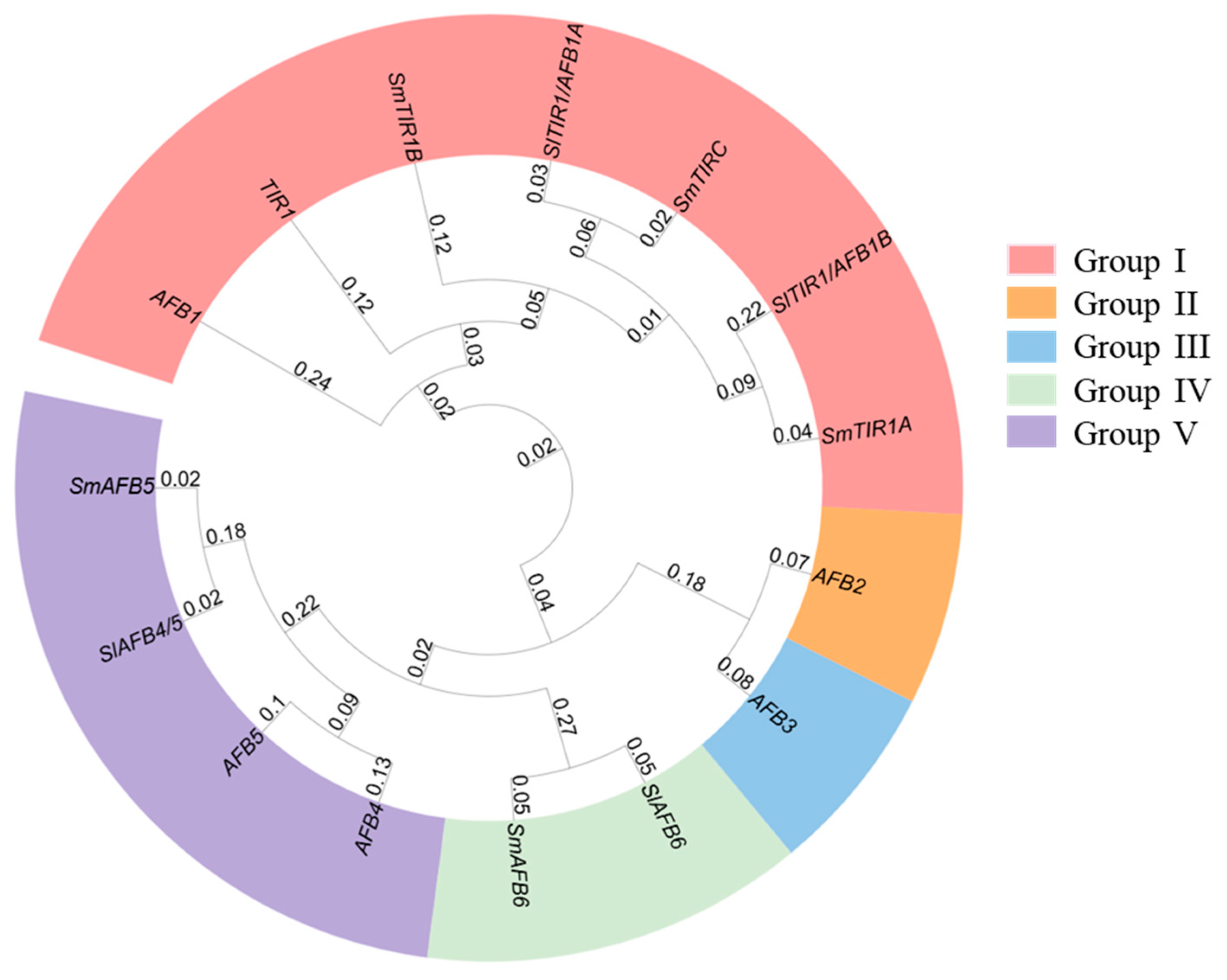
3.2. Phylogenetic Analysis and Classification of the S. melongena TIR1/AFB Genes
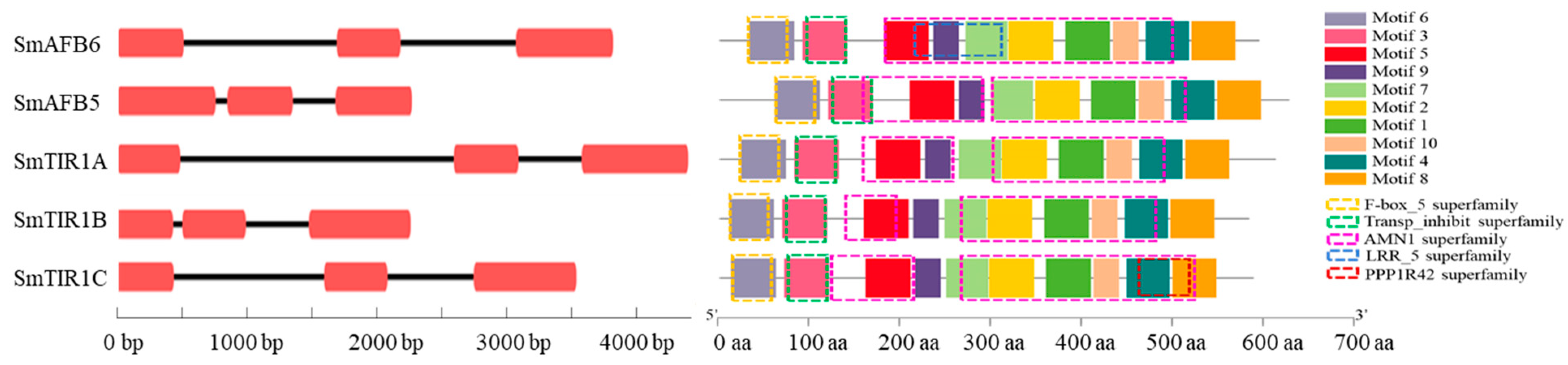
3.3. TIR1/AFB Gene Structure and Conserved Protein Motifs
3.4. Gene Duplicates among S. melongena TIR1/AFBs
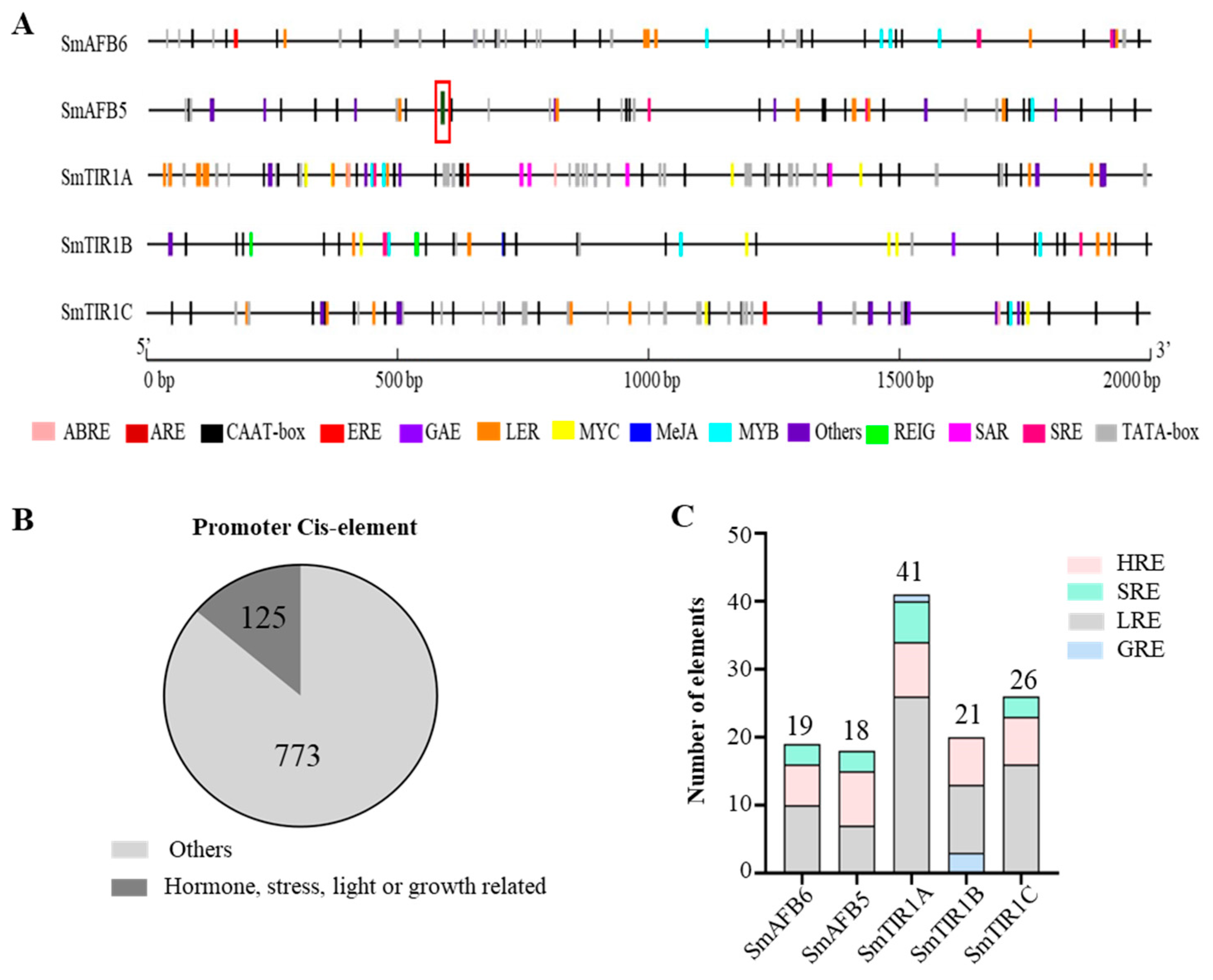
3.5. Promoter cis-Elements in the TIR1/AFB Genes
3.6. 3D Structure of the TIR1/AFB Proteins in S. melongena
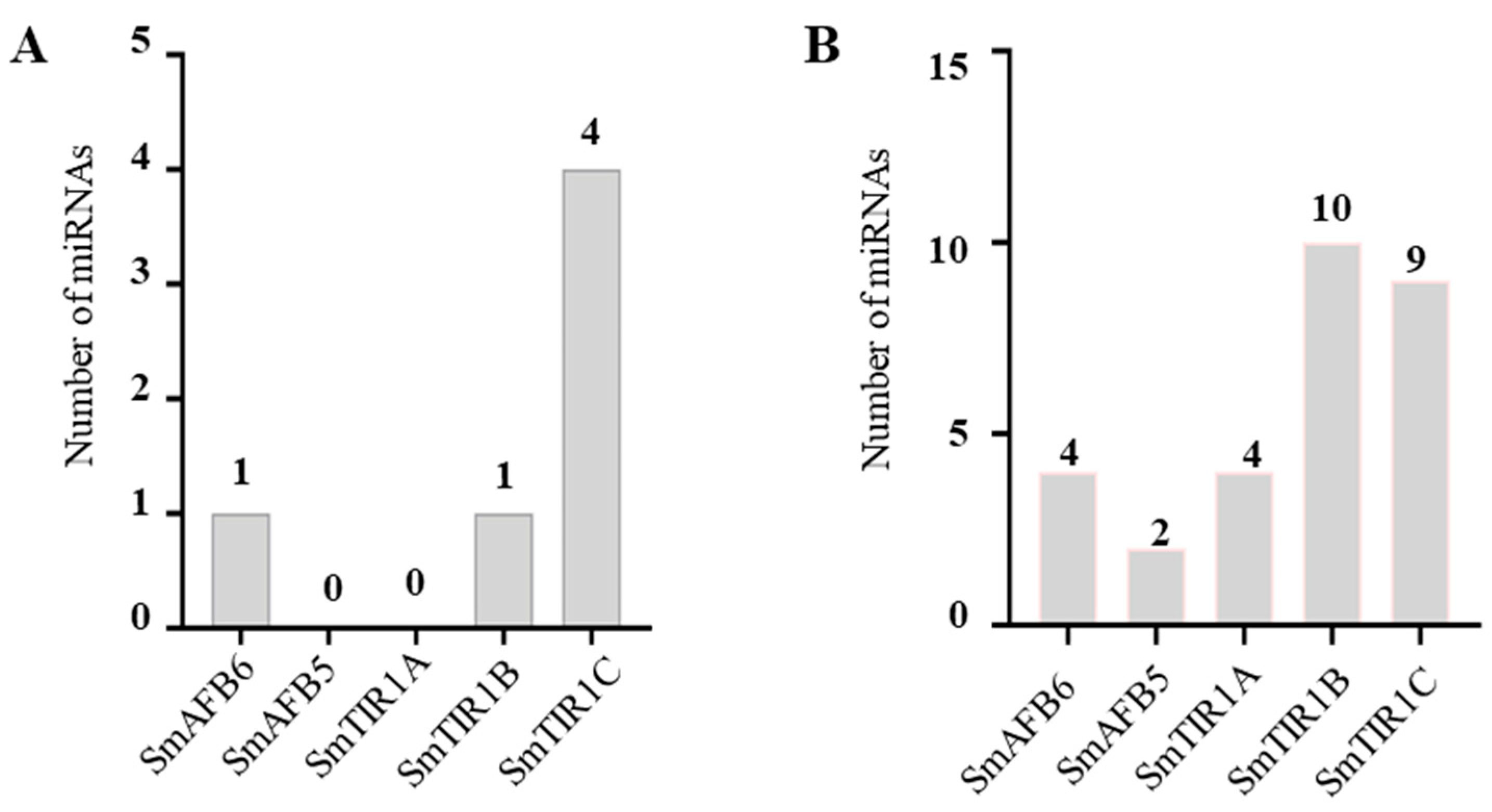
3.7. Identification of Potential miRNAs Targeting SmTIR1/AFB Genes
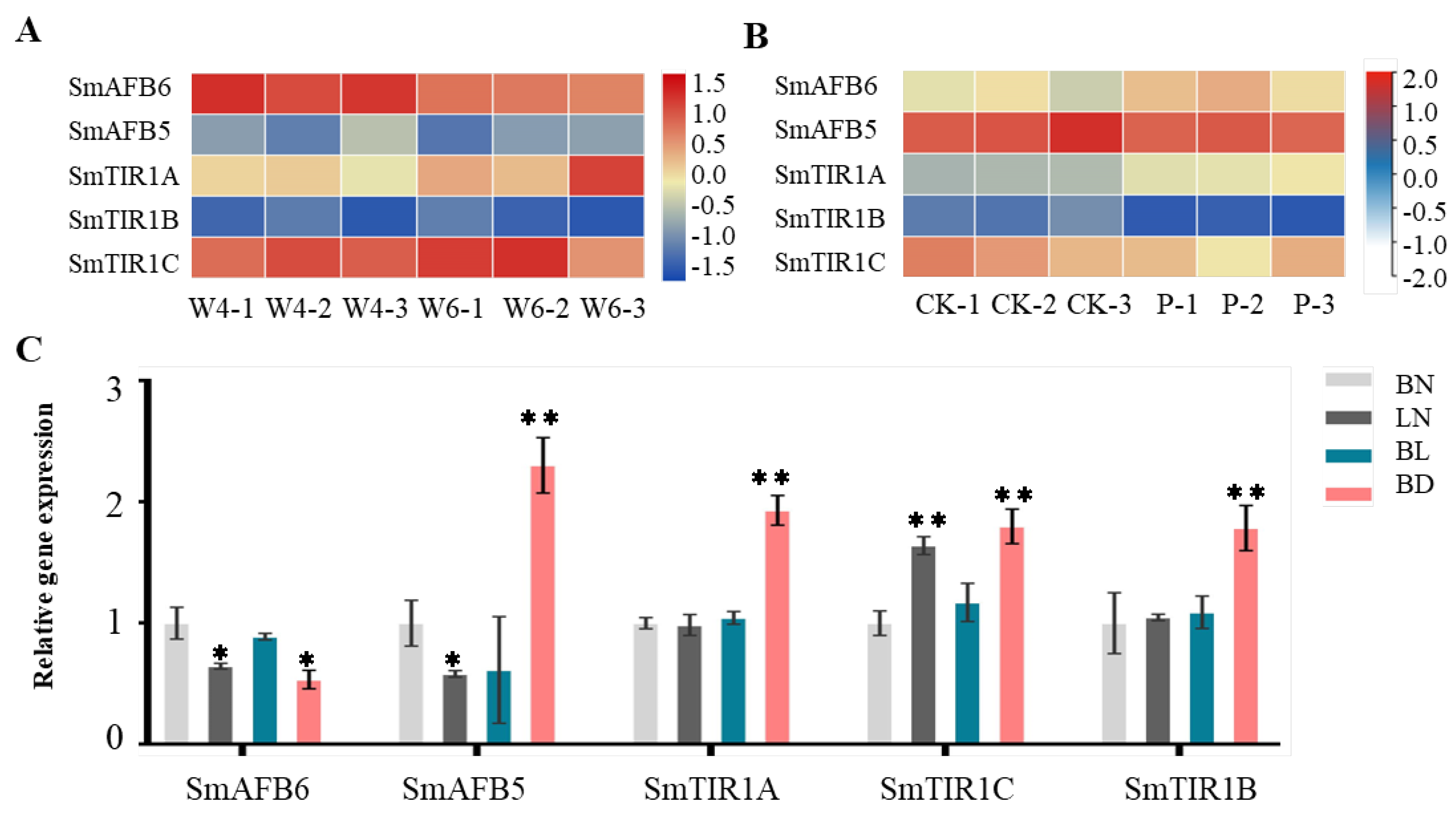
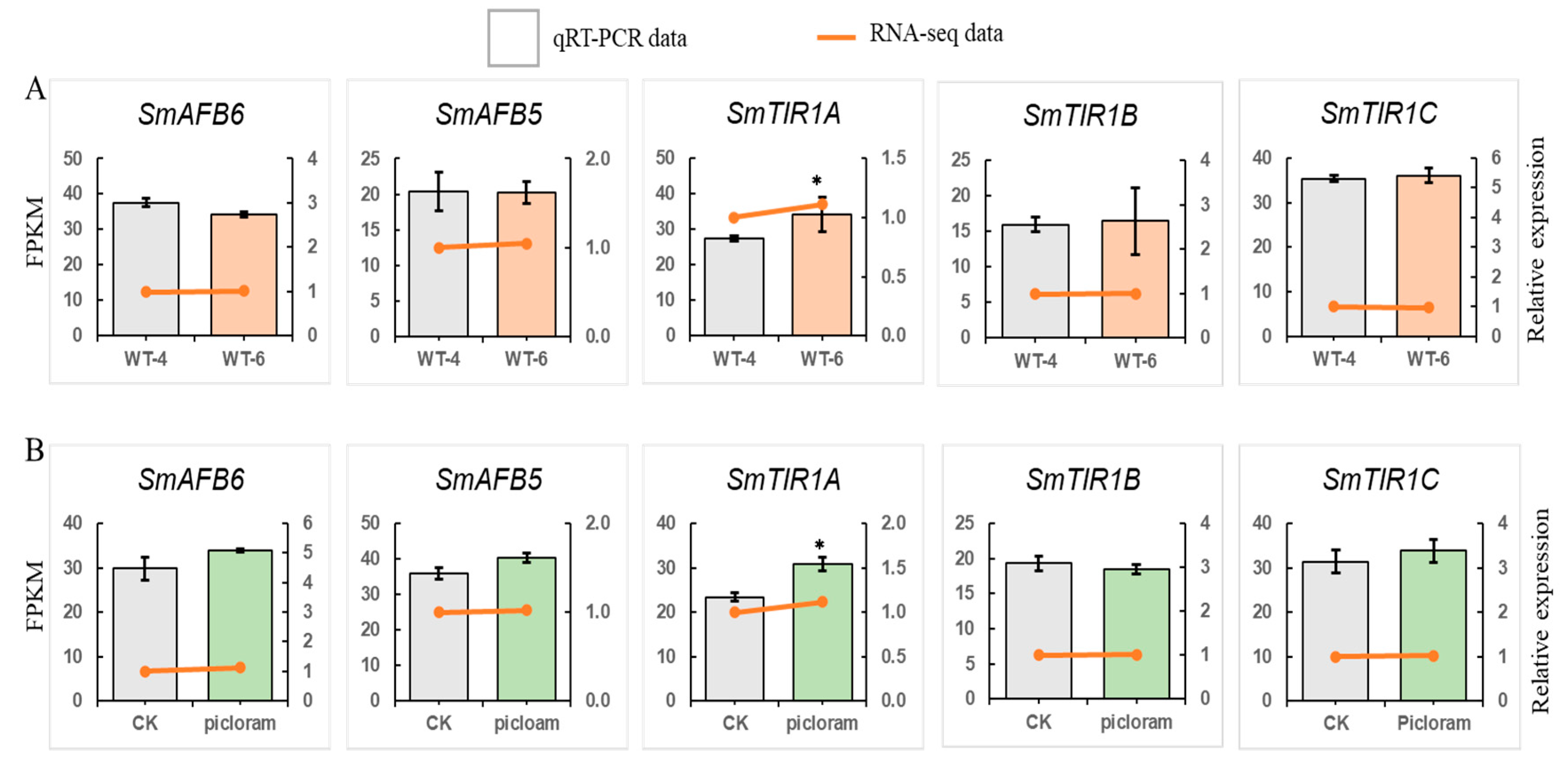
3.8. Transcriptome Analysis of SmTIR1/AFB Gene Expression
3.9. Knockdown of SmAFB5 Increases Resistance to Drought
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calderon Villalobos, L.I.A.; Lee, S.; De Oliveira, C.; Ivetac, A.; Brandt, W.; Armitage, L.; Sheard, L.B.; Tan, X.; Parry, G.; Mao, H.; et al. A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 2012, 8, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Kwiatkowski, M.; Chen, H.; Hoermayer, L.; Sinclair, S.; Zou, M.; del Genio, C.I.; Kubeš, M.F.; Napier, R.; Jaworski, K.; et al. Adenylate cyclase activity of TIR1/AFB auxin receptors in plants. Nature 2022, 611, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Parry, G.; Calderon-Villalobos, L.I.; Prigge, M.; Estelle, M. Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. USA 2009, 106, 22540–22545. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Platre, M.; Kadakia, N.; Zhang, Y.; Greenham, K.; Szutu, W.; Pandey, B.K.; Bhosale, R.A.; Bennett, M.J.; Bennett, M.J.; et al. Genetic analysis of the Arabidopsis TIR1/AFB auxin receptors reveals both overlapping and specialized functions. eLife 2020, 9, e54740. [Google Scholar] [CrossRef] [PubMed]
- Ozga, J.A.; Jayasinghege, C.P.A.; Kaur, H.; Gao, L.; Nadeau, C.D.; Reinecke, D.M. Auxin receptors as integrators of developmental and hormonal signals during reproductive development in pea. J. Exp. Bot. 2022, 73, 4094–4112. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Estelle, M. Diversity and specificity: Auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Huang, Y.; Qi, P.; Lian, G.; Hu, X.; Han, N.; Wang, J.; Zhu, M.; Qian, Q.; Bian, H. Functional analysis of auxin receptor OsTIR1/OsAFB family members in rice grain yield, tillering, plant height, root system, germination, and auxinic herbicide resistance. New Phytol. 2021, 229, 2676–2692. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Hormonal interactions underlying parthenocarpic fruit formation in horticultural crops. Hortic. Res. 2022, 9, uhab024. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, J.; Cui, L.; Zhang, T.; Wu, Z.; Zhu, P.-Y.; Meng, Y.-J.; Zhang, K.-J.; Yu, X.-Q.; Lou, Q.-F.; et al. New insights into the roles of cucumber TIR1 homologs and miR393 in regulating fruit/seed set development and leaf morphogenesis. BMC Plant Biol. 2017, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, I.; Sherif, S.M.; Jones, B.; Mila, I.; Kumar, P.P.; Bouzayen, M.; Jayasankar, S. TIR1-like auxin-receptors are involved in the regulation of plum fruit development. J. Exp. Bot. 2014, 65, 5205–5215. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghege, C.P.A.; Ozga, J.A.; Nadeau, C.D.; Kaur, H.; Reinecke, D.M. TIR1 auxin receptors are implicated in the differential response to 4-Cl-IAA and IAA in developing pea fruit. J. Exp. Bot. 2019, 70, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Lu, Y.; Li, Q.; Luo, S.; Shen, S.; Li, N.; Chen, X. TIR1/AFB proteins: Active players in abiotic and biotic stress signaling. Front. Plant Sci. 2022, 13, 1083409. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Vargas, F.; Godoy, T.; Tejos, R.; O’Brien, J.A. Overexpression of the auxin receptor AFB3 in Arabidopsis results in salt stress resistance and the modulation of NAC4 and SZF1. Int. J. Mol. Sci. 2020, 21, 9528. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Zhao, L.; Li, W.; Zhao, J.; Yan, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. Integrated metabolo-transcriptomics and functional characterization reveals that the wheat auxin receptor TIR1 negatively regulates defense against Fusarium graminearum. J. Integr. Plant Biol. 2021, 63, 340–352. [Google Scholar] [CrossRef] [PubMed]
- Fousia, S.; Tsafouros, A.; Roussos, P.A.; Tjamos, S.E. Increased resistance to Verticillium dahliae in Arabidopsis plants defective in auxin signalling. Plant Pathol. 2018, 67, 1749–1757. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Han, N.; Bian, H.; Liu, X.; Chan, Z. Hydrogen sulfide regulates abiotic stress tolerance and biotic stress resistance in Arabidopsis. J. Integr. Plant Biol. 2015, 57, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Vitor, S.C.; Duarte, G.T.; Saviani, E.E.; Vincentz, M.G.A.; Oliveira, H.C.; Salgado, I. Nitrate reductase is required for the transcriptional modulation and bactericidal activity of nitric oxide during the defense response of Arabidopsis thaliana against Pseudomonas syringae. Planta 2013, 238, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Benny, J.; Pisciotta, A.; Caruso, T.; Martinelli, F. Identification of key genes and its chromosome regions linked to drought responses in leaves across different crops through meta-analysis of RNA-Seq data. BMC Plant Biol. 2019, 19, 194. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Dalal, M.; Verma, R.K.; Kumar, S.V.; Yadav, S.K.; Pushkar, S.; Kushwaha, S.R.; Bhowmik, A.; Chinnusamy, V. Auxin protects spikelet fertility and grain yield under drought and heat stresses in rice. Environ. Exp. Bot. 2018, 150, 9–24. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Terrile, M.C.; Bartoli, C.G.; D’ippólito, S.; Casalongué, C.A. Auxin signaling participates in the adaptative response against oxidative stress and salinity by interacting with redox metabolism in Arabidopsis. Plant Mol. Biol. 2010, 74, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Muskett, P.R.; Chuang, H.-W.; Parker, J.E. Arabidopsis SGT1b is required for SCFTIR1-mediated auxin response. Plant Cell 2003, 15, 1310–1319. [Google Scholar] [PubMed]
- Liu, Y.; Li, D.; Zhang, S.; Zhang, L.; Gong, J.; Li, Y.; Chen, J.; Zhang, F.; Liao, X.; Chen, Z.; et al. Integrated Analysis of Microarray, Small RNA, and Degradome Datasets Uncovers the Role of MicroRNAs in Temperature-Sensitive Genic Male Sterility in Wheat. Int. J. Mol. Sci. 2022, 23, 8057. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, X.; Napier, R.; Dong, L.; Li, J. Mode of Action of a Novel Synthetic Auxin Herbicide Halauxifen-Methyl. Agronomy 2022, 12, 1659. [Google Scholar] [CrossRef]
- Luo, P.; Di, D.; Wu, L.; Yang, J.; Lu, Y.; Shi, W. MicroRNAs are involved in regulating plant development and stress response through fine-tuning of TIR1/AFB-dependent auxin signaling. Int. J. Mol. Sci. 2022, 23, 510. [Google Scholar] [CrossRef]
- Bian, H.; Xie, Y.; Guo, F.; Han, N.; Ma, S.; Zeng, Z.; Wang, J.; Yang, Y.; Zhu, M. Distinctive expression patterns and roles of the miRNA393/TIR1 homolog module in regulating flag leaf inclination and primary and crown root growth in rice (Oryza sativa). New Phytol. 2012, 196, 149–161. [Google Scholar] [PubMed]
- Li, K.; Wei, Y.-H.; Wang, R.-H.; Mao, J.-P.; Tian, H.-Y.; Chen, S.-Y.; Li, S.-H.; Tahir, M.-M.; Zhang, D. Mdm-miR393b-mediated adventitious root formation by targeted regulation of MdTIR1A expression and weakened sensitivity to auxin in apple rootstock. Plant Sci. 2021, 308, 110909. [Google Scholar] [PubMed]
- Luo, M.; Gao, J.; Peng, H.; Pan, G.; Zhang, Z. MiR393-targeted TIR1-like (F-box) gene in response to inoculation to R-Solani in Zea mays. Acta Physiol. Plant. 2014, 36, 1283–1291. [Google Scholar] [CrossRef]
- Shi, G.; Wang, S.; Wang, P.; Zhan, J.; Tang, Y.; Zhao, G.; Li, F.; Ge, X.; Wu, J. Cotton miR393-TIR1 module regulates plant defense against Verticillium dahliae via auxin perception and signaling. Front. Plant Sci. 2022, 13, 888703. [Google Scholar]
- Wojcik, A.M.; Gaj, M.D. miR393 contributes to the embryogenic transition induced in vitro in Arabidopsis via the modification of the tissue sensitivity to auxin treatment. Planta 2016, 244, 231–243. [Google Scholar] [PubMed]
- Du, W.; Lu, Y.; Luo, S.; Yu, P.; Shen, J.; Wang, X.; Xuan, S.; Wang, Y.; Zhao, J.; Li, N.; et al. Genome-wide transcriptome analysis reveals that upregulated expression of Aux/IAA genes is associated with defective leaf growth of the slf mutant in eggplant. Agronomy 2022, 12, 2647. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A high-quality chromosome-level genome assembly reveals genetics for important traits in eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
- Sarwar, R.; Jiang, T.; Ding, P.; Gao, Y.; Tan, X.; Zhu, K. Genome-wide analysis and functional characterization of the DELLA gene family associated with stress tolerance in B. napus. BMC Plant Biol. 2021, 21, 286. [Google Scholar]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Perspective—Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [PubMed]
- Yadav, S.; Kushwaha, H.R.; Kumar, K.; Verma, P.K. Comparative structural modeling of a monothiol GRX from chickpea: Insight in iron-sulfur cluster assembly. Int. J. Biol. Macromol. 2012, 51, 266–273. [Google Scholar] [PubMed]
- Song, X.; Liu, G.; Duan, W.; Liu, T.; Huang, Z.; Ren, J.; Li, Y.; Hou, X. Genome-wide identification, classification and expression analysis of the heat shock transcription factor family in Chinese cabbage. Mol. Genet. Genom. 2014, 289, 541–551. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [PubMed]
- Zhang, M.; Zhang, H.; Tan, J.; Huang, S.; Chen, X.; Jiang, D.; Xiao, X. Transcriptome analysis of eggplant root in response to root-knot nematode infection. Pathogens 2021, 10, 470. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Perrot-Rechenmann, C. Cellular responses to auxin: Division versus Expansion. Cold Spring Harb. Perspect. Biol. 2010, 2, a001446. [Google Scholar]
- Gidhi, A.; Mohapatra, A.; Fatima, M.; Jha, S.K.; Kumar, M.; Mukhopadhyay, K. Insights of auxin signaling F-box genes in wheat (Triticum aestivum L.) and their dynamic expression during the leaf rust infection. Protoplasma 2023, 260, 723–739. [Google Scholar] [PubMed]
- Mazur, E.; Kulik, I.; Hajny, J.; Friml, J. Auxin canalization and vascular tissue formation by TIR1/AFB-mediated auxin signaling in Arabidopsis. New Phytol. 2020, 226, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Li, H.; Wang, X.; Ji, X.; You, C. Genome-wide identification of apple auxin receptor family genes and functional characterization of MdAFB1. Hortic. Plant J. 2023, 9, 645–658. [Google Scholar]
- Cai, Z.; Zeng, D.-E.; Liao, J.; Cheng, C.; Sahito, Z.A.; Xiang, M.; Fu, M.; Chen, Y.; Wang, D. Genome-wide analysis of auxin receptor family genes in Brassica juncea var. tumida. Genes 2019, 10, 165. [Google Scholar] [CrossRef]
- Shu, W.; Liu, Y.; Guo, Y.; Zhou, H.; Zhang, J.; Zhao, S.; Lu, M. A Populus TIR1 gene family survey reveals differential expression patterns and responses to 1-naphthaleneacetic acid and stress treatments. Front. Plant Sci. 2015, 6, 719. [Google Scholar]
- Nagy, G.; Nagy, L. Motif grammar: The basis of the language of gene expression. Comput. Struct. Biotechnol. J. 2020, 18, 2026–2032. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, K.; Agarwal, N.; Pile, L.; Ansari, A. Gene architecture facilitates intron-mediated enhancement of transcription. Front. Mol. Biosci. 2021, 8, 669004. [Google Scholar] [CrossRef] [PubMed]
- Grabski, D.F.; Broseus, L.; Kumari, B.; Rekosh, D.; Hammarskjold, M.; Ritchie, W. Intron retention and its impact on gene expression and protein diversity: A review and a practical guide. Wiley Interdiscip. Rev.-RNA 2021, 12, e1631. [Google Scholar] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [PubMed]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Ma, H.; DePamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2007, 50, 873–885. [Google Scholar] [PubMed]
- Grant, D.; Cregan, P.; Shoemaker, R.C. Genome organization in dicots: Genome duplication in Arabidopsis and synteny between soybean and Arabidopsis. Proc. Natl. Acad. Sci. USA 2000, 97, 4168–4173. [Google Scholar] [PubMed]
- Dugas, D.V.; Bartel, B. MicroRNA regulation of gene expression in plants. Curr. Opin. Plant Biol. 2004, 7, 512–520. [Google Scholar] [PubMed]
- Martin, A.; Adam, H.; Díaz-Mendoza, M.; Żurczak, M.; González-Schain, N.D.; Suárez-López, P. Graft-transmissible induction of potato tuberization by the microRNA miR172. Development 2009, 136, 2873–2881. [Google Scholar] [PubMed]
- Naqvi, A.R.; Haq, Q.M.R.; Mukherjee, S.K. MicroRNA profiling of tomato leaf curl new delhi virus (tolcndv) infected tomato leaves indicates that deregulation of miR159/319 and miR172 might be linked with leaf curl disease. Virol. J. 2010, 7, 281. [Google Scholar] [PubMed]
- Candar-Cakir, B.; Arican, E.; Zhang, B. Small RNA and degradome deep sequencing reveals drought-and tissue-specific micrornas and their important roles in drought-sensitive and drought-tolerant tomato genotypes. Plant Biotechnol. J. 2016, 14, 1727–1746. [Google Scholar] [PubMed]
- Omidvar, V.; Mohorianu, I.; Dalmay, T.; Fellner, M. Identification of miRNAs with potential roles in regulation of anther development and male-sterility in 7B-1 male-sterile tomato mutant. BMC Genom. 2015, 16, 878. [Google Scholar]
- Tyurin, A.A.; Suhorukova, A.V.; Kabardaeva, K.V.; Goldenkova-Pavlova, I.V. Transient gene expression is an effective experimental tool for the research into the fine mechanisms of plant gene function: Advantages, limitations, and solutions. Plants 2020, 9, 1187. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Terrile, M.C.; Casalongue, C.A. Auxin and salicylic acid signalings counteract the regulation of adaptive responses to stress. Plant Signal. Behav. 2011, 6, 452–454. [Google Scholar] [PubMed]
- Chen, Z.; Hu, L.; Han, N.; Hu, J.; Yang, Y.; Xiang, T.; Zhang, X.; Wang, L. Overexpression of a miR393-resistant form of transport inhibitor response protein 1 (mTIR1) enhances salt tolerance by increased osmoregulation and Na+ exclusion in Arabidopsis thaliana. Plant Cell Physiol. 2015, 56, 73–83. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Instability Index | Amino Acids | GRAVY Score | Introns, Exons | Predicted Subcellular Location |
|---|---|---|---|---|---|
| SmAFB6 | 87.87 | 587 | −0.156 | 2, 3 | Nucleus, cytoplasm |
| SmAFB5 | 38.35 | 621 | −0.008 | 2, 3 | Nucleus, cytoplasm |
| SmTIR1A | 45.35 | 605 | 0.029 | 2, 3 | Chloroplast, cytoplasm |
| SmTIR1B | 52.81 | 576 | −0.024 | 2, 3 | Nucleus, cytoplasm |
| SmTIR1C | 55.93 | 581 | −0.06 | 2, 3 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, W.; Karamat, U.; Cao, L.; Li, Y.; Li, H.; Li, H.; Wei, L.; Yang, D.; Xia, M.; Li, Q.; et al. The TIR1/AFB Family in Solanum melongena: Genome-Wide Identification and Expression Profiling under Stresses and Picloram Treatment. Agronomy 2024, 14, 1413. https://doi.org/10.3390/agronomy14071413
Du W, Karamat U, Cao L, Li Y, Li H, Li H, Wei L, Yang D, Xia M, Li Q, et al. The TIR1/AFB Family in Solanum melongena: Genome-Wide Identification and Expression Profiling under Stresses and Picloram Treatment. Agronomy. 2024; 14(7):1413. https://doi.org/10.3390/agronomy14071413
Chicago/Turabian StyleDu, Wenchao, Umer Karamat, Liuqing Cao, Yunpeng Li, Haili Li, Haoxin Li, Lai Wei, Dongchen Yang, Meng Xia, Qiang Li, and et al. 2024. "The TIR1/AFB Family in Solanum melongena: Genome-Wide Identification and Expression Profiling under Stresses and Picloram Treatment" Agronomy 14, no. 7: 1413. https://doi.org/10.3390/agronomy14071413