
Effects of Nitrogen Application and Planting Density Interaction on the Silique-Shattering Resistance and Yield of Direct-Seeding Rapeseed (Brassica napus L.) in Sichuan


Abstract
1. Introduction
2. Materials and Methods
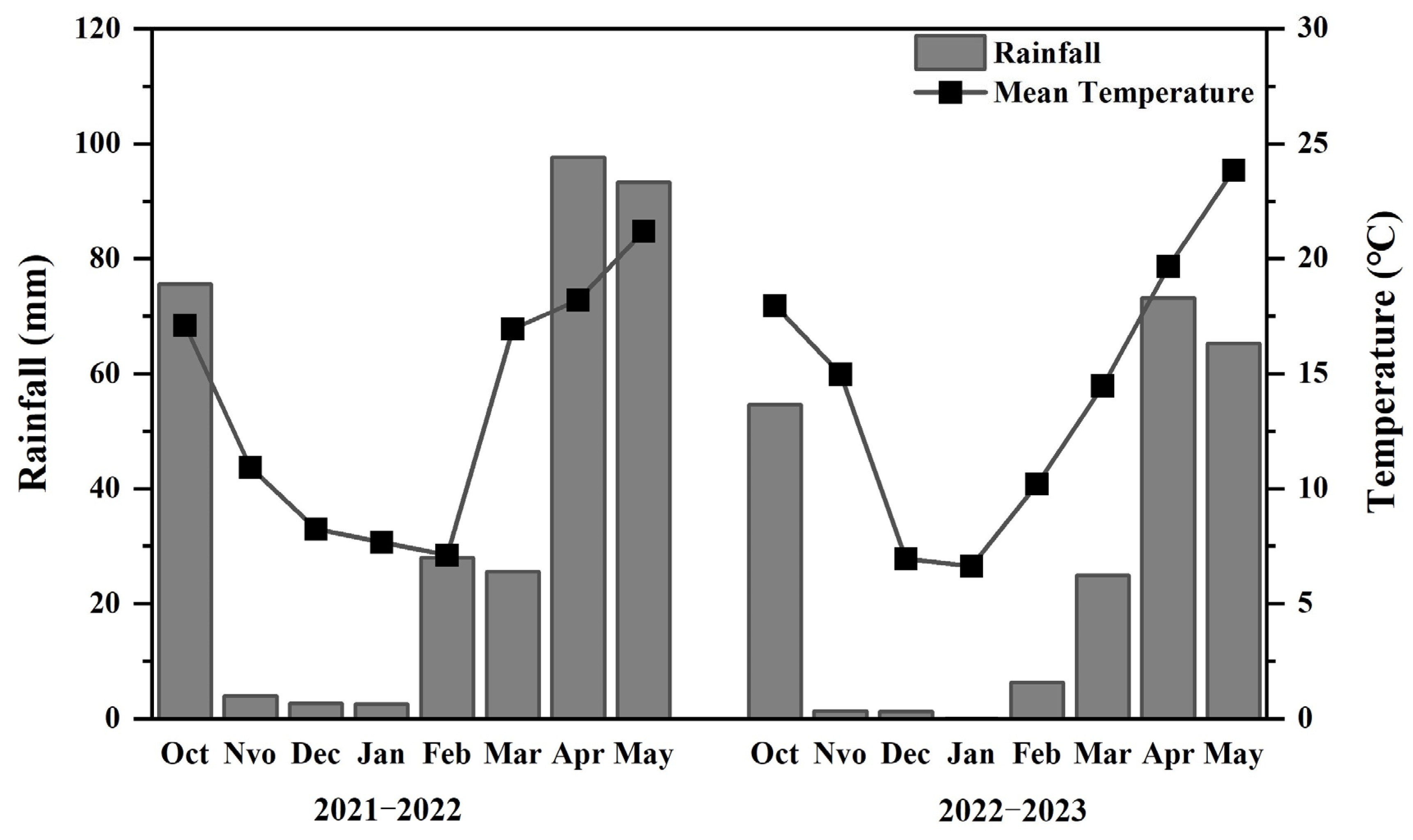
2.1. Experimental Site
2.2. Experimental Design and Field Management
2.3. Measurement Items and Methods
2.3.1. Determination of Morphological Indexes of Rapeseed Siliques
2.3.2. Moisture Content of the Siliques
2.3.3. Determination of Soluble Sugar Content in Silique Peel
2.3.4. Determination of Lignin and Cellulose Content in Silique Peel
2.3.5. Enzyme Activity Determination
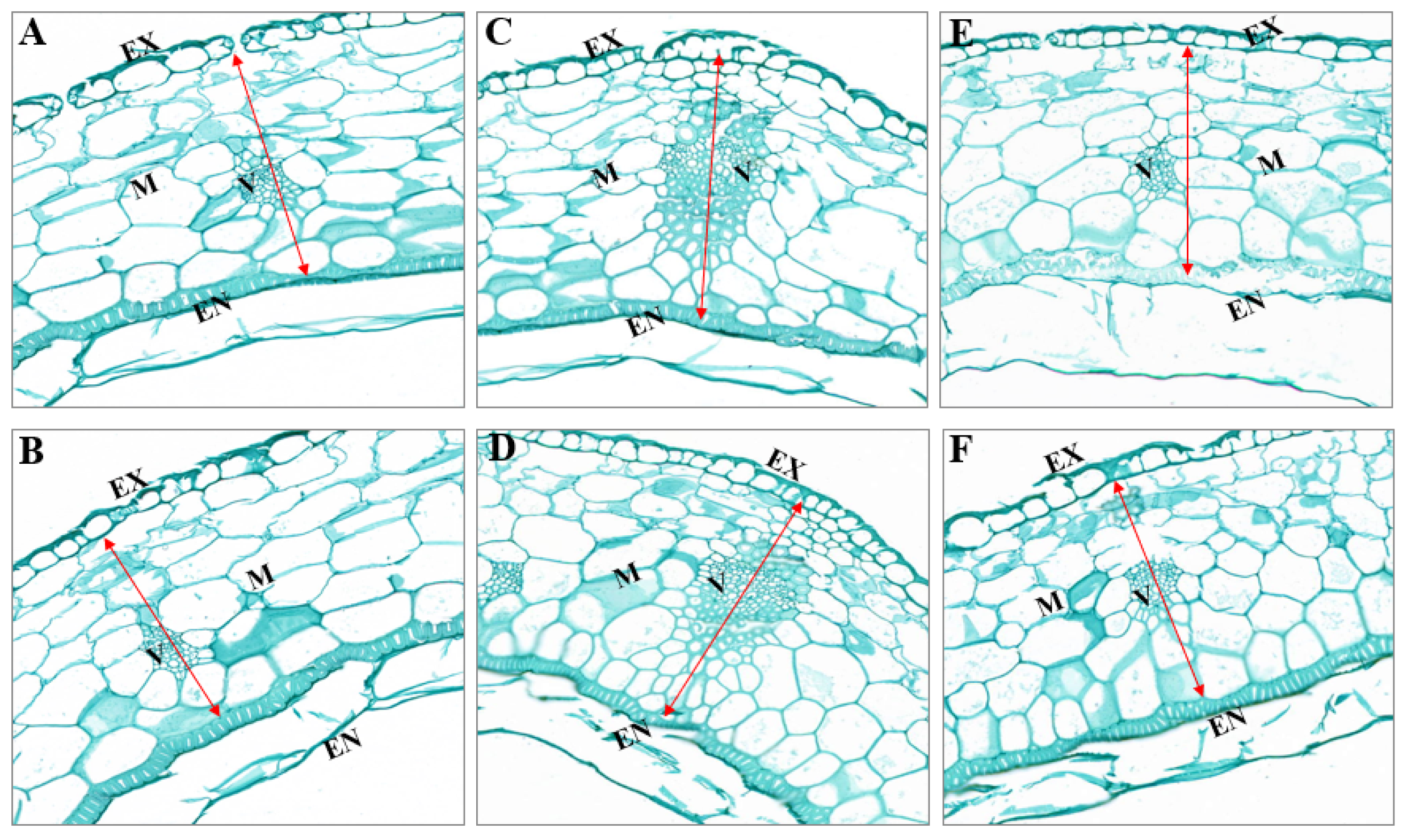
2.3.6. Cross-Section Structure of Silique Shell
2.3.7. Measurement of Yield and Its Component Factors
2.3.8. Determination of Silique Shatter Resistance Index
2.4. Statistical Analysis
3. Results
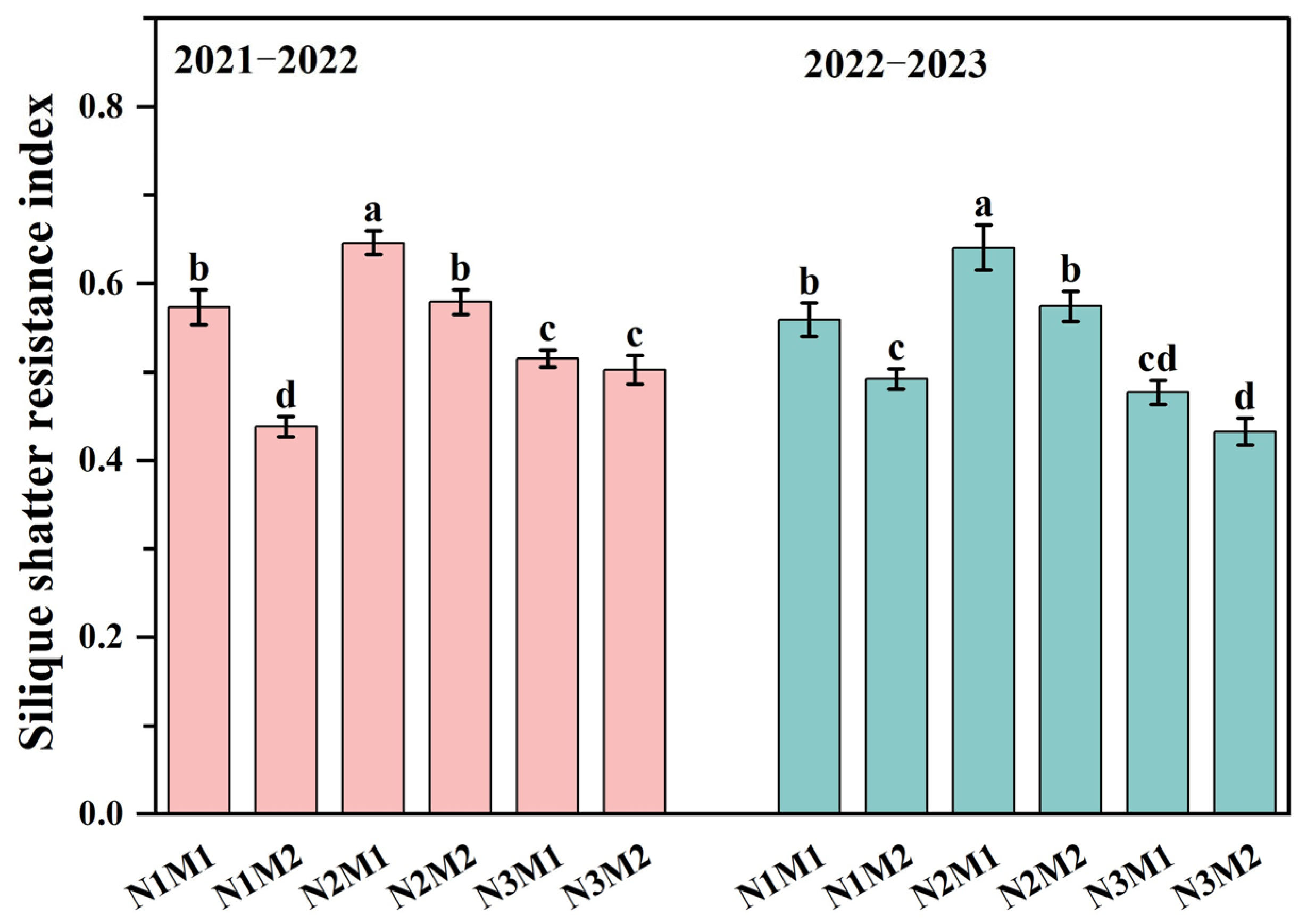
3.1. Effect of Nitrogen and Density Interaction on Silique Shatter Resistance Index
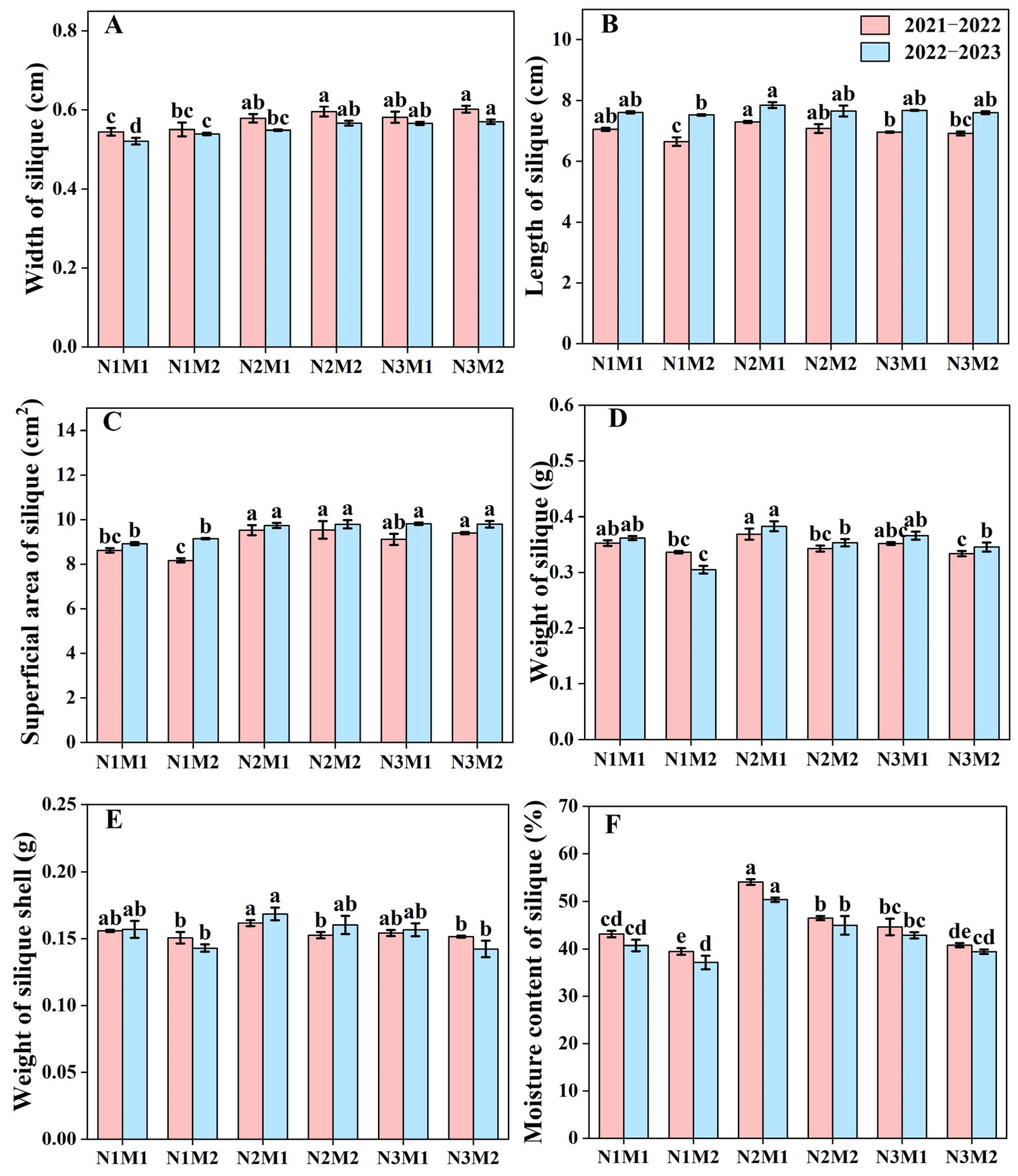
3.2. Effects of Nitrogen Density Interaction on Morphological and Physiological Indexes of Siliques
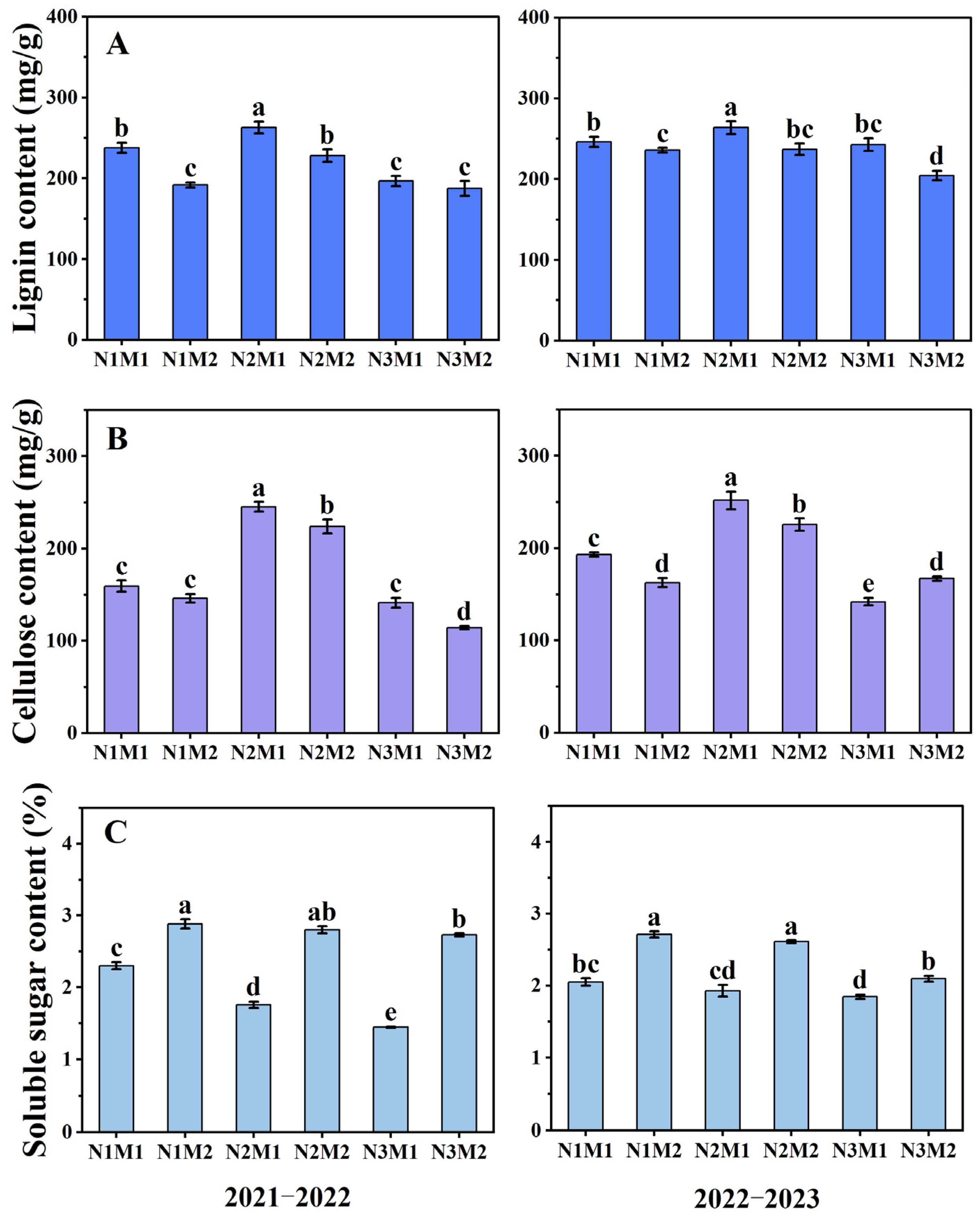
3.3. Effect of Nitrogen and Density Interaction on Carbohydrate in Silique Shell
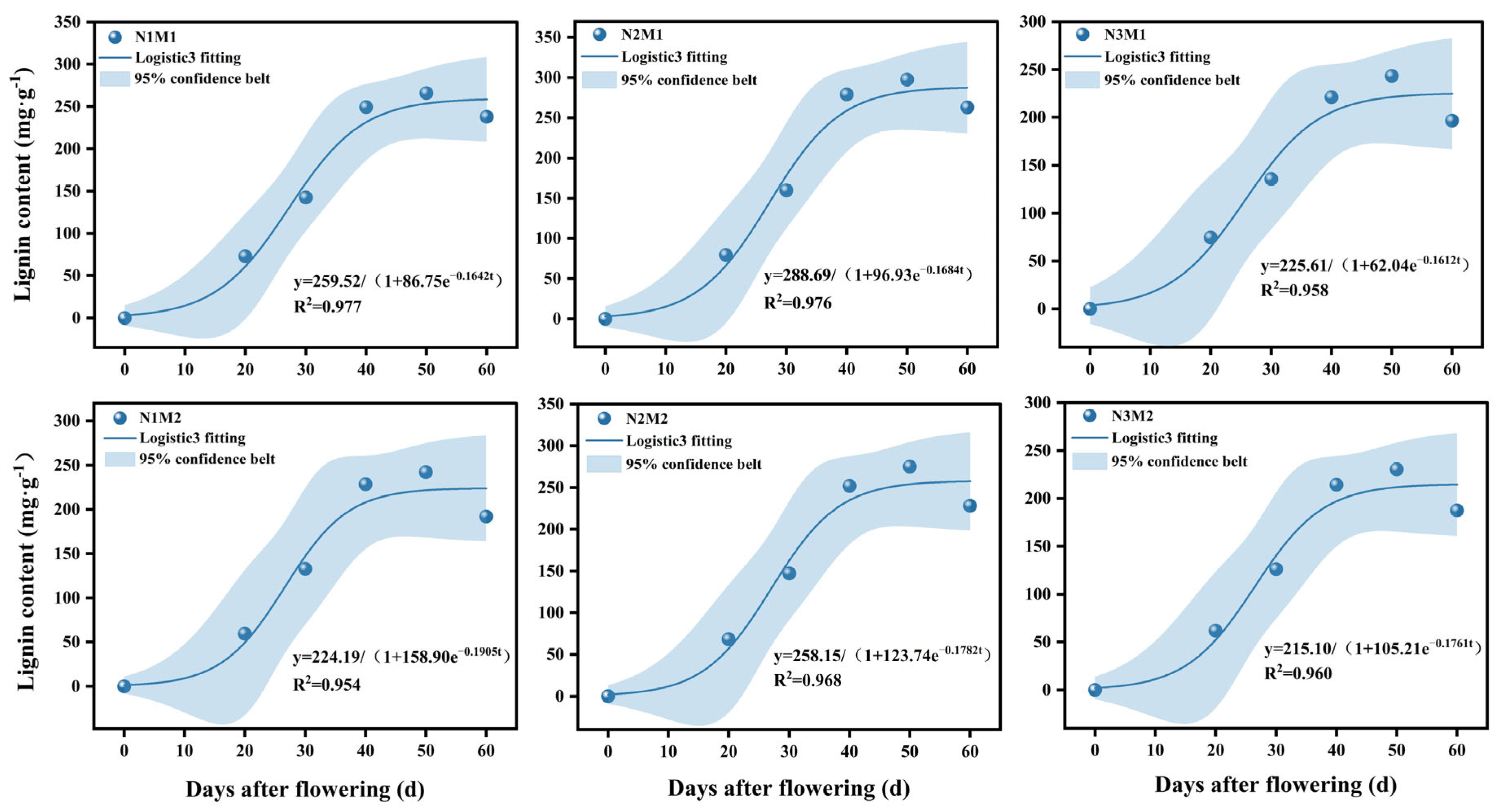
3.4. Effect of Nitrogen and Density Interaction on Parameter Characteristics of Lignin Accumulation Model in Silique Shell
3.5. Effect of Nitrogen and Density Interaction on Enzyme Activity in Silique Shell
3.6. Effects of Nitrogen Density Interaction on Yield and Yield Components
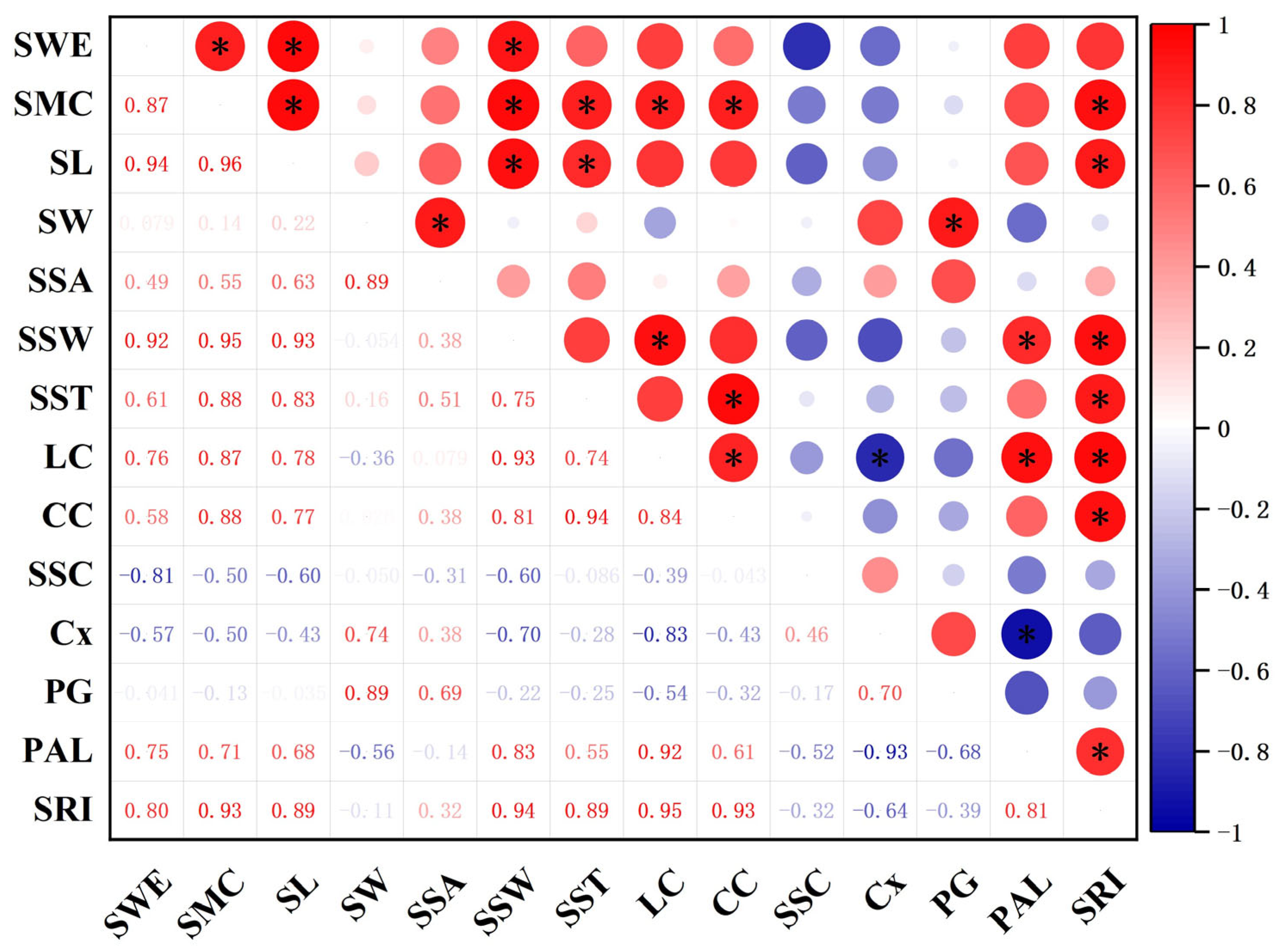
3.7. Correlation Analysis between Silique Characteristics and Shatter Resistance Index
4. Discussion
4.1. Effects of Nitrogen and Density Interaction on Silique Characteristics
4.2. Influence of Nitrogen and Density Interaction on Silique Shatter Resistance Index and Yield
4.3. Correlation between Silique Characteristics and Silique Shatter Resistance Index
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, H.; Hao, M.; Ding, B.; Mei, D.; Wang, W.; Wang, H.; Zhou, R.; Liu, J.; Li, C.; Hu, Q. Base editing with high efficiency in allotetraploid oilseed rape by A3A—PBE system. Plant Biotechnology J. 2021, 19, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Vera, C.L.; Downey, R.K.; Woods, S.M.; Raney, J.P.; McGregor, D.I.; Elliott, R.H.; Johnson, E.N. Yield and quality of canola seed as affected by stage of maturity at swathing. Can. J. Plant Sci. 2007, 87, 13–26. [Google Scholar] [CrossRef]
- Pari, L.; Assirelli, A.; Suardi, A.; Civitarese, V.; Del Giudice, A.; Santangelo, E. Seed losses during the harvesting of oilseed rape (Brassica napus L.) at on-farm scale. J. Agric. Eng. 2013, 44, e136. [Google Scholar] [CrossRef]
- Gulden, R.H.; Andrea, C.; Syrovy, L.D.; Shirtliffe, S.J. Pod drop in Brassica napus is linked to weight-ad justed pod-retention resistance. Field Crop Res. 2017, 205, 34–44. [Google Scholar] [CrossRef]
- Qing, Y.; Li, Y.; Xu, L.; Ma, Z.; Tan, X.; Wang, Z. Oilseed rape (Brassica napus L.) pod shatter resistance and its relationship with whole plant and pod characteristics. Ind. Crop Prod. 2021, 166, 113459. [Google Scholar] [CrossRef]
- Hu, Q.; Hua, W.; Yin, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed research and production in China. The Crop J. 2017, 5, 127–135. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, C.; Chen, H.; Wang, P.; Wang, P.; Song, C.; Zhang, X.; Wang, D. Multi-gene co-expression can improve compre hensive resistance to multiple abiotic stresses in Brassica napus L. Plant Sci. 2018, 274, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Cao, J.S.; Li, G.J.; Wu, X.H.; Wang, B.G.; Xu, P.; Hu, T.T.; Lu, Z.F.; Patrick, J.W.; Ruan, Y.L. Genotypic differences in pod wall and seed growth relate to invertase activities and assimilate transport pathways in asparagus bean. Ann. Bot. 2012, 109, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Kuai, J.; Sun, Y.; Liu, T.; Zhang, P.; Zhou, M.; Wu, J.; Zhou, G. Physiological mechanisms behind differences in pod shattering resistance in rapeseed (Brassica napus L.) varieties. PLoS ONE 2016, 11, e0157341. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Yu, Y.K.; Zhu, K.M.; Ding, L.N.; Wang, Z.; Yang, Y.H.; Cao, J.; Xu, L.Z.; Li, Y.M.; Tan, X.L. Down-regulation of MAN NANASE7 gene in Brassica napus L. enhances silique dehiscence-resistance. Plant Cell Rep. 2021, 40, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.P.; Basarkar, P.W.; Salimath, P.M.; Patil, S.A. Role of cell wall-degrading enzymes in pod-shattering process of soybean, Glycine max (L.). Merrill. Curr. Sci. 2002, 82, 58–61. [Google Scholar]
- Wen, Y.C.; Dong, T.; Zhi, C.; Shen, J.X.; Zhang, S.F. Advances in studies of pod shattering resistance in rapeseed. J. Plant Genet. Resour. 2009, 1, 140–145. [Google Scholar]
- Tao, Z.; Huang, Y.; Zhang, L.; Wang, X.; Liu, G.; Wang, H. BnLATE, a Cys2/His2-type zinc-finger protein, enhances silique shat tering resistance by negatively regulating lignin accumulation in the silique walls of Brassica napus. PLoS ONE 2017, 12, e0168046. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Liu, J.; Cheng, H.; Li, C.; Fu, L.; Wang, W.; Wang, H.; Hao, M.; Mei, D.; Liu, K.; et al. A lignified-layer bridge controlled by a single recessive gene is associated with high pod-shatter resistance in Brassica napus L. Crop J. 2022, 10, 638–646. [Google Scholar] [CrossRef]
- Liu, X.; Tu, B.; Zhang, Q.; Herbert, S.J. Physiological and molecular aspects of pod shattering resistance in crops. Czech J. Genet. Plant Breed. 2019, 55, 87–92. [Google Scholar] [CrossRef]
- Li, Y.S.; Yu, C.B.; Zhu, S.; Hu, X.J.; Liao, X.; Liao, X.S.; Che, Z. High planting density benefits to mechanized harvest and nitrogen application rates of oilseed rape (Brassica napus L.). Soil Sci. Plant Nutr. 2014, 60, 384–392. [Google Scholar] [CrossRef]
- Li, Z.; Gao, G.; Xu, L.; Wang, Z.; Wang, C.; Yang, T.; Kuai, J.; Wang, B.; Xu, Z.; Zhao, J.; et al. Reducing nitrogen application at high planting density enhances secondary cell wall formation and decreases stem lodging in rapeseed. Eur. J. Agron. 2024, 156, 127162. [Google Scholar] [CrossRef]
- Xiong, X.; Chang, L.; Khalid, M.; Zhang, J.; Huang, D. Alleviation of drought stress by nitrogen application in Brassica campestris ssp. Chinensis L. Agron. J. 2018, 8, 66. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Q.; Dong, X.; Liu, Y.; Lu, J. Herbicide and nitrogen rate effects on weed suppression, N uptake, use efficiency and yield in winter oilseed rape (Brassica napus L.). Glob. Ecol. Conserv. 2019, 17, e00529. [Google Scholar] [CrossRef]
- Zuo, Q.; Liu, J.; Wang, L.; Yang, G.; Leng, S. Yield, dry matter and N characteristics in canola as affected by fertilizer N rate and split-application ratio under high soil fertility condition. J. Plant Nutr. 2020, 43, 655–666. [Google Scholar] [CrossRef]
- Wen, G.; Ma, B.L.; Luce, M.S.; Liu, K.; Mooleki, P.S.; Crittenden, S.; Gulden, R.; Semach, G.; Tiege, P.; Lokuruge, P. Optimizing nitrogen fertilization for hybrid canola (Brassica napus L.) production across Canada. Field Crop Res. 2023, 302, 109048. [Google Scholar] [CrossRef]
- Zhan, N.; Xu, K.; Ji, G.; Yan, G.; Chen, B.; Wu, X.; Cai, G. Research progress in high-efficiency utilization of nitrogen in rape seed. Int. J. Mol. Sci. 2023, 24, 7752. [Google Scholar] [CrossRef] [PubMed]
- Laidig, F.; Feike, T.; Lichthardt, C.; Schierholt, A.; Piepho, H.P. Breeding progress of nitrogen use efficiency of cereal crops, winter oilseed rape and peas in long-term variety trials. Theor. Appl. Genet. 2024, 137, 45. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Konvalina, P.; Neugschwandtner, R.W.; Kopecký, M.; Amirahmadi, E.; Bucur, D.; Walkiewicz, A. Interaction of biochar with chemical, green and biological nitrogen fertilizers on nitrogen use efficiency indices. Agron. J. 2022, 12, 2106. [Google Scholar] [CrossRef]
- Gu, X.; Cheng, Z.; Du, Y.; Cai, H.; Li, Y.; Li, Y.; Fang, H.; Sun, S. Optimizing planting density to improve growth, yield and resources use efficiencies of winter oilseed rape under ridge-furrow film mulching. J. Integr. Agric. 2024. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Kadkol, G.P.; Pang, E.C.K.; Taylor, P.W.J. Identification of RAPD markers linked to recessive genes confer ring siliqua shatter resistance in Brassica rapa. Plant Breed. 2003, 122, 479–484. [Google Scholar] [CrossRef]
- Hu, Y.; Javed, H.H.; Du, Y.L.; Liao, Q.W.; Ye, W.; Zhou, J.; Peng, X.; Arslan, M.; Raza, A.; Wu, Y.C. Improving lignin metabolism, lodging resistance, and yield of rapeseed (Brassica napus L.) by applying straw-fermented fertilizer. J. Soil Sci. Plant Nut. 2023, 23, 2832–2848. [Google Scholar] [CrossRef]
- Khan, A.A.; McNeilly, T.; Collins, J.C. Accumulation of amino acids, proline, and carbohydrates in response to aluminum and manganese stress in maize. J. Plant Nut. 2000, 23, 1303–1314. [Google Scholar] [CrossRef]
- Morgan, C.L.; Bruce, D.M.; Child, R.; Ladbrooke, Z.L.; Arthur, A.E. Genetic variation for pod shatter resistance among lines of oilseed rape developed from synthetic B. napus. Field Crop Res. 1998, 58, 153–165. [Google Scholar] [CrossRef]
- Noda, H.M.; Muraoka, H.; Nasahara, K.N.; Saigusa, N.; Murayama, S.; Koizumi, H. Phenology of leaf morphological, photo synthetic, and nitrogen use characteristics of canopy trees in a cool-temperate deciduous broadleaf forest at Takayama, central Japan. Ecol. Res. 2015, 30, 247–266. [Google Scholar] [CrossRef]
- Song, Z.; Xu, G.; Feng, Y.; Li, J.; Luo, J.; Wang, X.; Gao, Y.; You, X.; Ren, H. Effects of Annual Straw Incorporation Combined with Application of Nitrogen Fertilizer in Rice Season on Dry Matter and Nutrient Accumulation Characteristics of Subsequent Rapeseed. Agron. J. 2023, 13, 1514. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, L.; Ding, Y.; Yao, X.; Wu, X.; Weng, F.; Li, G.; Liu, Z.; Tang, S.; Ding, C.; et al. Nitrogen fertilizer application affects lodging resistance by altering secondary cell wall synthesis in japonica rice (Oryza sativa). J. Plant Res. 2017, 130, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Yang, Y.; Liu, L.; Li, Z.; Zhu, C.; Hu, Q.; Liu, J. Decryption of superior allele and candidate genes for total lignin contents of rapeseed. Ind. Crop Prod. 2024, 214, 118483. [Google Scholar] [CrossRef]
- Kuai, J.; Sun, Y.; Zhou, M.; Zhang, P.; Zuo, Q.; Wu, J.; Zhou, G. The effect of nitrogen application and planting density on the radiation use efficiency and the stem lignin metabolism in rapeseed (Brassica napus L.). Field Crop Res. 2016, 199, 89–98. [Google Scholar] [CrossRef]
- Hua, S.; Chen, Z.H.; Zhang, Y.; Yu, H.; Lin, B.; Zhang, D. Chlorophyll and carbohydrate metabolism in developing silique and seed are prerequisite to seed oil content of Brassica napus L. Bot. Stud. 2014, 55, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, B.; Kuai, J.; Li, Z.; Bai, R.; Zhou, G. Planting density and variety intercropping improve organ biomass distribution of rapeseed to alleviate the trade--off between yield and lodging resistance. Crop Sci. 2021, 61, 2696–2712. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Kuai, J.; Ullah, S.; Fahad, S.; Zhou, G. Optimization of nitrogen rate and planting density for improving yield, nitrogen use efficiency, and lodging resistance in oilseed rape. Front. Plant Sci. 2017, 8, 246598. [Google Scholar] [CrossRef] [PubMed]
- Kuai, J.; Yang, Y.; Sun, Y.; Zhou, G.; Zuo, Q.; Wu, J.; Ling, X. Paclobutrazol increases canola seed yield by enhancing lodging and pod shatter resistance in Brassica napus L. Field Crop Res. 2015, 180, 10–20. [Google Scholar] [CrossRef]
- Zhang, M.; Li, G.; Yang, Y.; Jin, M.; Wang, G. Test trials and analysis of pod-shattering characteristics of harvested rapeseed silique. Appl. Sci. 2023, 13, 9369. [Google Scholar] [CrossRef]
- Braatz, J.; Harloff, H.J.; Emrani, N.; Elisha, C.; Heepe, L.; Gorb, S.N.; Jung, C. The effect of INDEHISCENT point mutations on silique shatter resistance in oilseed rape (Brassica napus). Theor. Appl. Genet. 2018, 131, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Malhi, S.S.; Brandt, S.A.; McDonald, C.L. Assessment of seed shattering resistance and yield loss in five oilseed crops. Can. J. Plant Sci. 2008, 88, 267–270. [Google Scholar] [CrossRef]
- Holzenkamp, K.; Link, W.; Möllers, C. Effect of a low seed lignin content on seed yield and quality in winter oilseed rape (Bras- sica napus). Plant Breed. 2022, 141, 801–806. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Jiang, Y.; Sun, H.; Duan, R.; Qu, J.; Yao, D.; Liu, S.; Guan, S. Formation mechanism and occurrence law of pod shattering in soybean: A Review. Phyton 2022, 91, 1327–1340. [Google Scholar] [CrossRef]
- Wu, B.; Cui, Z.; Ma, L.; Li, X.; Wang, H.; Wang, Y.; Yan, B.; Dong, H.; Gao, Y. Effects of planting density—Potassium interaction on the coordination among the lignin synthesis, stem lodging resistance, and grain yield in oil flax. Agron. J. 2023, 13, 2556. [Google Scholar] [CrossRef]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. The role of the pod in seed development: Strategies for manipulating yield. New Phytol. 2011, 190, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, Y.Z. Seed shattering: From models to crops. Front. Plant Sci. 2015, 6, 146140. [Google Scholar] [CrossRef] [PubMed]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | pH | Total N (g·kg−1) | Available K (mg·kg−1) | Available P (mg·kg−1) | Organic C (g·kg−1) |
|---|---|---|---|---|---|
| 2021–2022 | 5.94 | 2.13 | 131.97 | 9.79 | 34.40 |
| 2022–2023 | 5.96 | 1.93 | 128.65 | 9.65 | 33.93 |
| Treatment | Lignin Accumulation Model Equation | R2 | t0 | t1 | t2 | Δt |
|---|---|---|---|---|---|---|
| N1M1 | y = 259.52/(1 + 86.75e−0.1642t) | 0.977 | 27 | 19 | 35 | 16 |
| N1M2 | y = 224.19/(1 + 158.90e−0.1905t) | 0.954 | 27 | 20 | 34 | 14 |
| N2M1 | y = 288.69/(1 + 96.93e−0.1684t) | 0.976 | 27 | 19 | 35 | 16 |
| N2M2 | y = 258.15/(1 + 123.74e−0.1782t) | 0.968 | 27 | 20 | 34 | 14 |
| N3M1 | y = 225.61/(1 + 62.04e−0.1612t) | 0.958 | 26 | 17 | 34 | 17 |
| N3M2 | y = 215.10/(1 + 105.21e−0.1761t) | 0.960 | 26 | 19 | 34 | 15 |
| Years | Nitrogen Dose (N) | Sowing Density (M) | Effective Siliques Per Plant | Seeds Per Pod | 1000-Seed Weight (g) | Seed Yield (kg ha−1) | Oil Yield (kg ha−1) | Oil Content (%) |
|---|---|---|---|---|---|---|---|---|
| 2021–2022 | N1 | M1 | 135.97 d | 18.27 ab | 4.15 b | 1974.33 d | 985.33 d | 49.91 a |
| M2 | 108.94 e | 17.30 c | 4.06 c | 2124.67 c | 1043.45 c | 49.11 b | ||
| N2 | M1 | 169.17 a | 18.57 a | 4.19 a | 3651.33 a | 1684.73 a | 46.14 d | |
| M2 | 144.03 c | 17.66 bc | 4.15 b | 3711.83 a | 1703.81 a | 45.90 d | ||
| N3 | M1 | 161.20 b | 18.47 a | 4.18 a | 3141.67 b | 1519.92 b | 48.37 bc | |
| M2 | 131.06 d | 17.56 bc | 4.14 b | 3205.17 b | 1531.71 b | 47.79 c | ||
| Variance analysis | N | ** | ns | ** | ** | ** | ** | |
| M | ** | ** | ** | ** | * | * | ||
| N × M | Ns | ns | ** | ns | ns | ns | ||
| 2022–2023 | N1 | M1 | 122.86 d | 21.93 bc | 3.77 d | 1821.39 e | 901.25 d | 49.47 a |
| M2 | 101.28 e | 21.19 c | 3.68 e | 2110.94 d | 1036.61 c | 49.10 ab | ||
| N2 | M1 | 175.97 a | 24.47 a | 4.02 a | 3350.08 b | 1523.99 b | 45.44 d | |
| M2 | 155.42 b | 22.83 bc | 3.91 b | 3627.14 a | 1649.16 a | 45.50 d | ||
| N3 | M1 | 143.14 c | 23.42 ab | 3.96 b | 3104.31 c | 1490.16 b | 47.99 bc | |
| M2 | 123.11 d | 21.42 c | 3.86 c | 3232.71 bc | 1541.42 b | 47.67 c | ||
| Variance analysis | N | ** | * | ** | ** | ** | ** | |
| M | ** | ** | ** | ** | ** | ns | ||
| N × M | Ns | ns | ns | ns | ns | ns | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Jiang, H.; Hu, Y.; Liu, L.; Ghaffor, K.; Javed, H.H.; Peng, X.; Guo, X.; Wu, Y. Effects of Nitrogen Application and Planting Density Interaction on the Silique-Shattering Resistance and Yield of Direct-Seeding Rapeseed (Brassica napus L.) in Sichuan. Agronomy 2024, 14, 1437. https://doi.org/10.3390/agronomy14071437
Luo Y, Jiang H, Hu Y, Liu L, Ghaffor K, Javed HH, Peng X, Guo X, Wu Y. Effects of Nitrogen Application and Planting Density Interaction on the Silique-Shattering Resistance and Yield of Direct-Seeding Rapeseed (Brassica napus L.) in Sichuan. Agronomy. 2024; 14(7):1437. https://doi.org/10.3390/agronomy14071437
Chicago/Turabian StyleLuo, Yuqin, Huayu Jiang, Yue Hu, Li Liu, Kamran Ghaffor, Hafiz Hassan Javed, Xiao Peng, Xiang Guo, and Yongcheng Wu. 2024. "Effects of Nitrogen Application and Planting Density Interaction on the Silique-Shattering Resistance and Yield of Direct-Seeding Rapeseed (Brassica napus L.) in Sichuan" Agronomy 14, no. 7: 1437. https://doi.org/10.3390/agronomy14071437
APA StyleLuo, Y., Jiang, H., Hu, Y., Liu, L., Ghaffor, K., Javed, H. H., Peng, X., Guo, X., & Wu, Y. (2024). Effects of Nitrogen Application and Planting Density Interaction on the Silique-Shattering Resistance and Yield of Direct-Seeding Rapeseed (Brassica napus L.) in Sichuan. Agronomy, 14(7), 1437. https://doi.org/10.3390/agronomy14071437