Abstract
Rotation fallow is an effective way to overcome the obstacles associated with continuous cropping, being beneficial for the growth and development of crops. Soil micro-organisms are closely related to soil fertility, plant productivity, soil pathogenic bacteria, and crop health in agricultural ecosystems. To explore the effects of different rotation fallow modes on the diversity and functions of the soil bacterial community, a study was conducted in an arid area in the western foothills of the Greater Khingan Mountains. Using spring wheat variety Longmai 36 as the research material, this study systematically analyzed the changes and functional differences in soil physicochemical and biological characteristics, as well as microbial communities (endosphere, rhizosphere, and bulk soil) in spring wheat fields under five rotation fallow modes: Wheat2016–Wheat2017–Wheat2018(WWW), Wheat2016–Rape2017–Fallow2018(WRF), Wheat2016–Potato2017–Fallow2018(WPF), Wheat2016–Fallow2017–Rape2018(WFR), and Wheat2016–Fallow2017–Potato2018 (WFP). The results indicate that, compared to WWW, the soil urease activity, microbial biomass nitrogen content, and microbial biomass phosphorus content were significantly increased in the WFP mode (p < 0.05). In particular, the soil moisture content, organic matter, and total potassium content were increased by 6.88%, 3.34%, and 25.57%, respectively. The Shannon index and chao1 index of bulk soil (BS) and rhizosphere (RS) bacteria were significantly higher than those of endosphere (ER) bacteria (p < 0.05). Both ecological niche and rotation fallow modes affected the relative abundance of dominant bacteria, and the relative abundance of beneficial bacteria, such as Bacteroidetes, Firmicutes, and Verrucomimicrobia, significantly increased in the rotation fallow modes. The complexity and stability of bacterial networks, and abundance of nitrogen-cycling-related functional taxa were significantly improved, while the abundance of pathogen-related functional taxa was significantly decreased. The differences in soil bacterial community structure were closely related to soil physicochemical properties. Compared to ER, BS and RS bacterial communities, which are more susceptible to soil physicochemical properties, and soil pH are key driving forces for bacterial community distribution. In summary, compared with continuous cropping, the rotation fallow mode is beneficial for conserving soil moisture and nutrients, stabilizing soil pH, (i.e., making the soil tend to be neutral), increasing the abundance of beneficial bacteria in the soil, enhancing the complexity and stability of microbial ecological networks, and increasing the abundance of nitrogen-cycling-related functional taxa, thus improving crop growth and development.
1. Introduction
Soil micro-organisms play important roles in soil functions, ecosystem stability, and material cycling. They mainly participate in the formation of humus in soil, the decomposition and release of soil nutrients, and the conversion and cycling of energy [1]. In agricultural ecosystems, when functional microbacteria are limited by environmental changes, other microbial groups may exercise the same function to maintain the functional stability of the agricultural ecosystem [1,2]. Therefore, if community species are replaced in space or time due to environmental changes, the increase in soil microbial diversity can help to achieve functional complementarity between species, maintain soil functional diversity, and ensure ecosystem stability [3]. In addition, soil micro-organisms play important roles in the growth, development, and immunity of host plants [4,5]. When a host plant is under environmental pressure, soil micro-organisms can assist the plant in resisting stress and promoting its growth and development through synthesizing plant biomass, improving soil nutrient availability, and inhibiting the growth and reproduction of pathogenic micro-organisms [6,7]. Therefore, the improvement of the soil microbial diversity is of great significance for the sustainable development of agricultural ecology.
Soil micro-organisms, as an important component of agricultural ecosystems, are driven by the agricultural environment and host plant species and varieties [8]. Research has shown that different cultivation practices can lead to changes in the composition of soil microbial communities [9,10]. Continuous cropping is a common cultivation method that refers to the continuous cultivation of the same crop on the same cultivated land. Long-term continuous cropping can lead to an increase in the number of pathogenic micro-organisms and deterioration due to soil-borne pathogenic diseases, thereby affecting the stability of agricultural ecosystems [11]. In addition, long-term continuous cropping can lead to a decrease in soil carbon storage, a significant reduction in microbial availability of carbon sources, and a decrease in diversity [12]. To solve the problem of intensified soil-borne diseases and severe soil-quality degradation caused by long-term continuous cropping, crop rotation and fallow farming systems have been widely promoted and applied in many countries and regions, with significant ecological and socio-economic benefits [13].
Rotation fallow is an international practice that combines the planting system of rotation with the cultivation system of fallow, which solves problems, such as decreased soil fertility, deterioration of the ecological environment, and imbalanced grain production structure in farmland. Different crop types can induce the enrichment of specific micro-organisms through their nutrient preferences and root exudates [14,15]. Compared to continuous cropping, crop rotation can effectively regulate the dynamic balance of soil nutrients, improve the soil micro-environment in farmland, and, thus, affect soil microbial diversity. However, different crop rotations or fallow fields can result in differences in the composition and quantity of root exudates in the soil environment, leading to the recruitment of different soil microbial communities. The number of pathogenic bacteria, such as Phoma, Myrothecium, and Verticillium species, significantly decreased, and wheat–potato rotation with fallow soil occurred while crop yield and water use efficiency were significantly increased [16]. Corn–soybean rotation significantly increased the soil pH and nutrient content (e.g., N and P); increased the abundance of beneficial bacteria, such as Bradyrizobium sp., Gemmatimonas sp., and Mortierella sp.; improved soil fertility; and increased crop yield [17]. Therefore, in order to better understand the impact of rotation fallow modes on soil microbial community composition, this study aims to clarify key functional taxa and identify the main environmental variables driving microbial community changes, in order to obtain suitable rotation fallow modes for soil improvement, soil fertility cultivation, and crop yield increase in the arid area of the western foothills of the Greater Khingan Mountains. Spring wheat (Longmai 36) was taken as the research object, and five crop rotation fallow modes were considered. The changes in soil microbial diversity, microbial community composition, and abundance of beneficial micro-organisms under different rotation fallow modes were systematically analyzed, thus revealing the response mechanisms of soil micro-organisms to different rotation fallow modes.
2. Materials and Methods
2.1. Site Description and Experimental Design
This study was conducted as a long-term rotation fallow experiment at the Tenihe Soil Management and Ecological Restoration Scientific Observation and Experimental Station of the Inner Mongolia Autonomous Region Academy of Agricultural and Animal Husbandry Sciences (120°48′ E, 49°55′ N, altitude 650 m). This area is located in a middle temperate, semi-arid continental grassland climate zone, with an annual average temperature of 2.2 °C. The frost-free period is 90–105 days, and the annual average precipitation is 373–474 mm. The precipitation from June to September accounts for about 80% of the annual precipitation. The effective rainfall in the whole growth period of spring wheat in 2019 was 223.9 mm. The soil type of cultivated land is chernozem. The basic nutrient content of 0–20 cm of soil is shown in Table 1.

Table 1.
Basic physical and chemical properties of a 0~20 cm soil layer in the experimental land.
Long-term treatment was started in 2016, with the previous crop (2015) being rape. The experiment adopted a randomized block design (Figure 1), and a total of 5 treatment modes were set up: Wheat2016–Wheat2017– Wheat2018 (WWW); Wheat2016–Rape2017–Fallow2018 (WRF); Wheat2016–Potato2017–Fallow2018 (WPF); Wheat2016–Fallow2017–Rape2018 (WFR); and Wheat2016–Fallow2017–Potato2018 (WFP). The test material was spring wheat Longmai 36. Each processing method was set with 3 replicates, totaling 15 communities, with a community area of 9 square meters and a block spacing of 0.5 m. The experimental site was equipped with a 1.0 m bandwidth protection row. After land preparation (plowing, harrowing, and leveling), spring wheat sowing was carried out on 5 May 2019, with a sowing amount of 300 kg/hm−2 and a row spacing of 0.15 m. Urea: 60 kg/hm−2, diammonium phosphate: 180 kg/hm−2, potassium sulfate: 30 kg/hm−2 were applied as seed fertilizer before sowing, without topdressing in the later stage. Other management methods were the same as those in these fields.
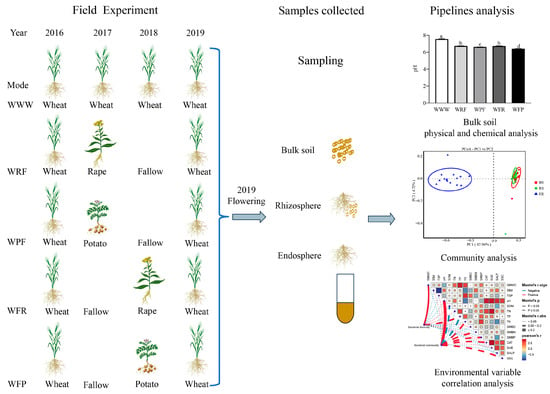
Figure 1.
Field experiments and experimental design. The lowercase letters on the bar chart indicate the significance of different rotation fallow modes at the p < 0.05 level.
2.2. Sample Collection
Bulk soil, rhizosphere, and endosphere samples were collected at the flowering stage of spring wheat. Referring to Pershina et al. [18], the specific approach was to use the “S” sampling method in the community, remove the topsoil at the sampling point, and collect soil samples from the 0–20 cm soil layer using a soil drill. Each treatment was repeated three times. After thoroughly and evenly sifting the soil, the sample was divided into two parts. One part was placed in a plastic bag and stored at 4 °C for the soil microbial biomass measurement; the other was screened with a 1 mm sterile mesh to remove impurities (spring wheat roots, leaves, etc.), then dried in the shade and stored in a sealed plastic bag containing desiccant for the soil physicochemical and biological properties’ determination. The specific determination method refers to Liu et al. [19]. We randomly selected 5 representative spring wheat plants in each community, shook the roots to mix the bulk soil and loose soil, and placed them in sterile tubes as bulk soil (BS) samples. We removed large pieces of soil from the roots and quickly transferred the wheat to the laboratory on ice. Then, we placed the wheat roots in sterile tubes, washed them with PBS buffer, and centrifuged them to obtain the rhizosphere (RS) sample. We washed the wheat roots clean with PBS buffer and sterile water, absorbed the surface moisture of the roots, cut them into small pieces with sterile scissors, mixed them well, and placed them in sterile tubes as endosphere (ER) samples (Figure 1). The samples of BS, RS, and ER were all frozen in liquid nitrogen and stored at −80 °C. A total of 45 samples was collected (treated with 5 rotation fallow modes) × 3 sample types (BS, RS, and ER) × 3 replicates and were used for bacterial diversity sequencing.
2.3. Determination of Soil Biological and Physicochemical Properties
The determination of the mass water content of soil (SMWC) involves the utilization of the drying and weighing method. Soil samples are collected and placed in an aluminum box with a known weight (g1) before being weighed (g2). Subsequently, they are transferred to an oven set at 105 °C and dried until a constant weight is achieved (g3). The soil moisture content is then calculated using the formula w = (g2 − g3)/(g3 − g1) × 100. Referring to Bai et al.‘s [20] analysis methods, the soil bulk density (SBd) and total soil porosity (TSP) were assessed using the ring knife method. Soil pH was measured utilizing the potentiometric method. The content of soil organic matter (SOM) was determined through the potassium dichromate dilution heat method, while the total nitrogen (TN) content was ascertained via the Kjeldahl digestion method. Total phosphorus (TP) content in the soil was determined by employing the HClO4-H2SO4 digestion method. Total potassium in the soil (TK) was quantified using an NaOH melt flame photometer (Jingke fp6410, Shanghai, China). Soil urease (SUE) activity was measured utilizing the indophenol blue colorimetric method. Soil alkaline phosphatase (SALP) detection employed sodium benzoate phosphate’s colorimetric approach, while the sucrase activity assessment utilized 3,5-dinitrosalicylic acid’s colorimetric technique. Catalase activity measurement involved potassium permanganate titration. The determination of soil microbial biomass carbon content (SMBC) utilized the chloroform fumigation extraction volumetric analysis methodology. Soil microbial biomass nitrogen (SMBN) was determined using chloroform fumigation extraction and the ninhydrin colorimetric method. Soil microbial biomass phosphorus (SMBP) was determined using the chloroform fumigation extraction total phosphorus determination method.
2.4. DNA Extraction, Amplification, and Construction of Illumina MiSeq Sequencing Library
DNA was extracted from soil samples using the Fast DNA® Spin Kit for Soil (Thomas Fisher Scientific, Waltham, MA, USA). The concentration and purity of DNA were detected by NanoDrop2000uv-Vis (Thermo Fisher Scientific, Waltham, MA, USA), and the quality of DNA was detected by 1% agarose gel electrophoresis. Place an appropriate amount of the sample into a centrifuge tube, dilute the sample to 1 ng with sterile water/µL, and store it at −80 °C for future use. Primers 515F(GTGCCAGCMGCCGCGGTAA)/806R (GGACTACHVGGGTWTCTAAT) were used to amplify the V4 variable region of the bacterial 16S rRNA gene [21]. All PCRs were carried out in 30 µL reactions with 15 µL of Phusion® High-Fidelity PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 0.2 µM of forward and reverse primers, and about 10 ng of template DNA. Thermal cycling consisted of initial denaturation at 98 °C for 1 min, followed by 35 cycles of denaturation at 98 °C for 10 s, annealing at 50 °C for 30 s, and elongation at 72 °C for 30 s. Finally, 72 °C for 5 min. The PCR product was extracted from 1.5% agarose gel and purified using the GeneJET Gel Extraction Kit (Thermo Fisher Scientific, Waltham, MA, USA), according to manufacturer’s instructions. The quality and content of DNA were tested using NanoDrop2000 (Thermo Fisher Scientific, Waltham, MA, USA). According to the standard protocols of Novo Genentech Co., LTD. (Beijing, China), sequencing libraries were generated using the Illumina TruSeq DNA PCR-Free Library Preparation Kit (Illumina, San Diego, CA, USA), following the manufacturer’s recommendations. Library quality was assessed on the Qubit@ 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA) and Agilent Bioanalyzer 2100 system (Agilent Technologies Inc., Santa Clara, CA, USA). the library was sequenced on an Illumina NovaSeq platform. Raw data were uploaded to the NCBI SRA database (accession number: PRJNA1104604).
2.5. Data Processing Analysis
Fastp software (https://github.com/OpenGene/fastp, version 0.19.6) (accessed on 12 September 2020) was used to perform quality control on the raw sequencing data [22]. Paired-end reads from the original DNA fragments were merged by using FLASH software (https://www.cbcb.umd.edu/software/flash, version 1.2.11) (accessed on 18 September 2020) [23], and reads were filtered by QIIME (Quantitative Insights Into Microbial Ecology) (https://qiime2.org/) (accessed on 25 September 2020) quality filters. Sequences with ≥97% similarity were assigned to the same OTUs. Standardize the OTU matrix using the trimmed mean of M values method in R package edge R [24]. The RDP classifier (http://rdp.cme.msu.edu/, version 2.11) (accessed on 15 December 2020) was used to compare the SILVA (https://www.arb-silva.de/) (accessed on 20 December 2020) ribosomal RNA database for the taxonomic annotation of OTUs, with a confidence threshold of 70%, and to statistically analyze the community composition of each sample at different community classification levels [25]. On the NovoMagic Cloud platform (https://magic.novogene.com/) (accessed on 10 December 2022), we conducted bacterial diversity analysis. Chao1 and Shannon indices were calculated by Qiime. The Wilcox rank sum test and one-way analysis of variance were used to analyze the alpha diversity between treatment groups. R (Version 2.15.3) ade4 and ggplot2 were used for the principal co-ordinates analysis (PCoA) of community differences at different spatial locations. NMDS (non-metric multidimensional scaling) analysis and the Scheirer–Ray–Hare test were conducted on the differences in bacterial composition among different rotation fallow modes using R (Version 2.15.3) vegan [26]. LEfSe software (https://github.com/SegataLab/lefse, Version 1.1.1) (accessed on 12 February 2023) was used (LDA > 3.4, p < 0.05) to analyze significant differences in bacterial levels among different rotation fallow modes. Based on the Spearman correlation |r| > 0.6 p < 0.05, a collinearity network analysis was conducted on the top 100 bacterial genera, and visualized using graphviz-2.38.0 software to calculate network diameter, modularity, clustering coefficient, graph density, and average path length. FAPROTAX (http://www.loucalab.com/archive/FAPROTAX/, Version 1.2.0) (accessed on 20 February 2023) [27] was used to predict bacterial function.
The soil physical and chemical properties were statistically analyzed using Excel 2016 (Microsoft, Redmond, WA, USA), and a one-way analysis of variance (ANOVA) was performed on each variable using IBM SPSS Statistics 22.0 (IBMCorp., Amonk, NY, USA). The significant differences among the treatments were compared (p < 0.05), and the Spearman correlation between the variables was analyzed. Graphpad Prism 8 (Graphpad, La Jolla, CA, USA) was used for visualization. Analyze the correlation between environmental factors and dominant bacterial genera through redundancy analysis (RDA). We screened environmental factors at the operational taxonomic unit (OTU) level using the Mantel test and variance inflation factor (VIF). We calculated the ecosystem multifunctionality index (MFI) using the mean method based on key environmental variables [28]. Using the OmicStudio tool (https://www.omicstudio.cn/tool) (accessed on 20 March 2023) Perform RDA, network related heatmap analysis, and linear regression analysis. Soil factors are represented by key environmental variables, bacterial alpha diversity is represented by the Shannon index, bacterial community differences are represented by the Bray–Curtis beta diversity index, nitrogen-related functional taxa (NRFT) are represented by the average predicted nitrogen-cycling-related functional taxa, and soil microbial network complexity is represented by the clustering coefficient (CC).
3. Results
3.1. Analysis of Soil Physicochemical Properties and Biological Properties
Soil physical and chemical properties are important indicators reflecting soil health, which are closely related to soil material circulation and energy flow. Through the analysis of soil physicochemical and biological properties under different rotation fallow modes, it was found that different rotation fallow modes had significant effects on the properties (ANOVA, p < 0.05). Compared to the continuous cropping mode of spring wheat (WWW), the spring wheat–fallow–potato mode (WFP), spring wheat–fallow–rape mode (WFR), and spring wheat–rape–fallow mode (WRF) increased soil mass water content (SMWC) by 6.88%, 4.31%, and 2.96%, respectively. Furthermore, the total potassium (TK) content in the soil increased by 25.57%, 13.36%, and 3.44%, and that under the spring wheat–potato–fallow mode (WPF) increased by 8.65%; except for WRF, the organic matter content (SOM) in the soil increased significantly under the other three modes, with WPF, WFP, and WFR leading to increases of 3.68%, 3.34%, and 3.34%, respectively. The soil pH value of the WWW spring wheat field was 7.54, significantly higher than the other four crop rotation fallow modes (Figure 2 and Figure S1). The activities of catalase (CAT) and alkaline phosphatase (SALP) in WWW soil were significantly increased, while the urease (SUE) activity, soil microbial biomass nitrogen (SMBN), and soil microbial biomass phosphorus (SMBP) content in WFP soil were significantly higher than those in the other four modes (ANOVA, p < 0.05).
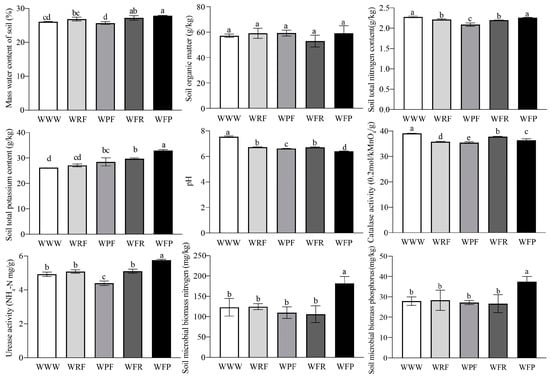
Figure 2.
Analysis of soil physical and chemical properties under different crop rotation fallow modes. The error bars on the bar chart are represented by standard deviation (SD), and the lowercase letters on the bar chart indicate the significance of different rotation fallow modes at the p < 0.05 level.
3.2. Analysis of Soil Bacterial Diversity
Soil micro-organisms are an important component of the soil microenvironment. Rotation fallow changes the physical and chemical properties of soil, to some extent, and may also have a significant impact on soil microbial diversity. Therefore, we performed bacterial 16 S rRNA amplicon sequencing on 45 samples collected from three different locations (soil bulk, rhizosphere, and endosphere) under five crop rotation fallow modes. It was found that the rotation fallow mode had no significant effect on bacterial alpha diversity (Duncan, p = 0.148), while the ecological niche significantly affected bacterial alpha diversity (Duncan, p < 0.001). Moreover, under the same rotation fallow mode, Shannon and chao1 were found to be ordered as follows: Bulk soil > Rhizosphere > Endosphere (Figure 3A,B; Tables S1 and S2). The bacterial Shannon indices for soil bulk, rhizosphere, and endosphere were significantly increased under the WFR mode, compared with WWW (Duncan, p = 0.001).
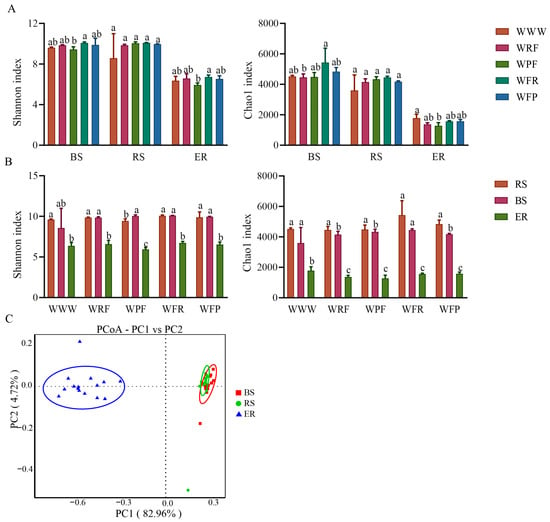
Figure 3.
Diversity of bacterial communities in different root space positions under different crop rotation fallow modes. (A) Analysis of differences in bacterial Shannon index and Chao1 index between different rotation fallow modes in the same spatial position. The error bars on the bar chart are represented by standard deviation (SD), and the lowercase letters on the bar chart indicate the significance of different rotation fallow modes at the p < 0.05 level. (B) Analysis of differences in Shannon index and Chao1 index of bacteria at different spatial positions under the same rotation fallow modes. (C) Principal coordinate analysis (PCoA) of bacteria in different ecological niches. BS: bulk soil; RS: rhizosphere; ER: endosphere.
Using principal coordinate analysis (PCoA), non-metric multidimensional scaling (NMDS), and the rank-based Scheirer–Ray–Hare test, the bacterial community structure under different rotation fallow modes was analyzed. It was found that the bacterial community was significantly distinguished by the ecological niche (PC1 = 82.96%, H niche > H mode, p < 0.001; Table S2) and, compared with endosphere, bulk soil, and rhizosphere, the sample points were clustered and their community structure was similar (Figure 3C). In the same ecological niche, the bacterial communities under the five rotation fallow modes were significantly distinguished, with WWW located in a single quadrant, WPF and WRF located in the same quadrant, and WFR and WFP located in the same quadrant (Figure S2). These results indicate that a niche is the main factor driving bacterial community changes, while crop rotation fallow modes are secondary factors (Figure 3C). The bacterial communities under WPF and WRF rotations were similar, and the bacterial communities under WFR and WFP rotations were also similar. Therefore, crop rotation sequence may also be the reason for the differences in bacterial communities between samples.
3.3. Analysis of Differences in Soil Bacterial Community Composition
The species optimization annotation of bacterial operational taxonomic units (OTUs) was conducted at three different positions (BS, RS, and ER) under five rotation fallow modes at the phylum level. The top three bacterial phyla, in terms of relative abundance rankings, were found to be Proteobacteria (20.42–51.57%), Actinobacteria (7.57–27.94%), and Bacteroidetes (3.50–16.06%). The relative abundance of Proteobacteria showed BS > RS > ER in the WWW, WRF, and WPF modes, and RS > BS > ER in WFR and WFP modes (Figure 4A). In BS and ER samples, compared with WWW, the relative abundance of Firmicutes was increased by over 93.75% in the rotation fallow mode, while the relative abundance of Acidobacteria significantly decreased, with a decrease of no less than 25.89% (Table S3). Under the rotation fallow mode, the relative abundances of Bacteroidetes and Verrucomicrobia in BS and RS samples significantly increased, while ER significantly decreased. The results show that both the crop rotation fallow mode and ecological niche can affect the relative abundance of dominant bacteria. The relative abundances of Bacteroidetes, Firmicutes, and Verrucomicrobia were significantly increased under crop rotation fallow modes, while Actinobacteria tended to accumulate in wheat roots.
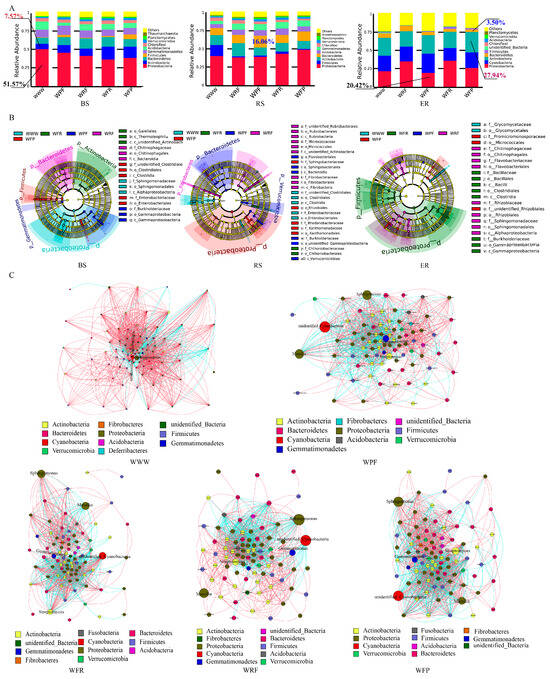
Figure 4.
Differences in bacterial community composition among different crop rotation fallow modes. (A) Composition of bacterial communities in different crop rotation fallow modes. (B) LEfSe analyzes the evolutionary branching diagram, from the inner circle to the outer circle; the species at the phylum, class, order, family, and genus levels are in order, and the species names represented by the English letters in the figure are shown in the legend on the right. The red, green, blue, purple, and light-blue nodes in the branches are the microbial groups that play an important role in WFP, WFR, WPF, WRF, and WWW rotations, respectively, and the yellow nodes represent the microbial groups that do not play an important role in the five treatments. (C) Horizontal correlation network of bacterial genera in different crop rotation fallow modes. The circular nodes in the figure represent different genera, the node size represents abundance, the node color represents different bacterial phyla, the red line represents a positive correlation, and the green line represents a negative correlation.
Linear discriminant analysis effect size (LEfSe) was used to analyze species with significant differences in three spatial locations under the five crop rotation fallow modes. The bacteria found to have significant differences were included in 5 phyla, 9 classes, 14 orders, 19 families, and 17 genera (Figure 4B). Under the WWW, WPF, WFR, and WFP modes, the BS biomarkers were Sphingomonas, Massilia, Segetibacter, and Clostridia, respectively. Under the WWW, WRF, and WFR modes, the RS biomarkers were Ruminococcus, Rubrobacteria, and Chthoniobacteraceae, respectively. In the WPF mode, the RS biomarkers were Massilia and Segetibacter, while, in the WFP mode, the RS biomarkers were Rhodanobacter and Pedobacter. There were 11 biomarkers in ER, of which only 1 was in the WWW mode, which was Glycomyces. WRF, WPF, and WFR all had two biomarkers each: Sphingomonas and Flavobacterium; Amycolatopsis and Niastella; and Massilia and Bacillus, respectively. There were four biomarkers in the WFP mode: Promicromonospora, Devosia, Phyllobacterium, and Neorhizobium (Figure S3). The results show that there are few species and a simple species composition in the WWW mode. In contrast, under the WFP mode, there were many species of bacteria with a rich species composition, with most bacteria having the ability to inhibit plant pathogenic micro-organisms, which may be more conducive to farmland ecological restoration and crop growth.
To further clarify the differences in bacterial community composition among the different crop rotation and fallow modes, we selected the top 100 species with total abundance at the genus classification level for microbial network construction (Spearman correlation, p < 0.01). There were significant differences in bacteria-related networks among the five rotation fallow modes (Figure 4C). Compared with the WWW mode, the rotation fallow modes exhibited a more complex and compact network structure, with a higher average degree, clustering coefficient (CC), and lower degree of modularity. In addition, the number and connectivity of negatively correlated bacteria in the WWW network were significantly lower than those under the crop rotation fallow modes (Figure 4C and Figure S4, Table S4). These results indicate that, compared to the WWW mode, bacterial interactions under rotation fallow modes are more aggregated and complex, while the long-term continuous cropping of spring wheat leads to decreases in negatively correlated bacteria and network stability.
3.4. Analysis of Functional Differences between Soil Dominant Bacteria
To clarify the metabolic and ecological functions of dominant bacteria enriched under the five modes, a functional annotation was performed on OTUs enriched in three ecological niches under the modes based on FAPROTAX. It was found that bacterial functional taxa were related to the ecological cycling of soil carbon, nitrogen, sulfur, and other elements, and were significantly influenced by the modes and bacterial ecological niches (Figure 5 and Figure S5). The main dominant functional taxa under the five modes and three niches were chemoheterotrophic and aerobic heterotrophic (Figure 5A). Compared with the WWW mode, the rotation fallow mode had a higher abundance of functional taxa related to cycling nitrogen, sulfur, arsenic, manganese, and other elements (Figure S5A), and nitrogen-related functional taxa (NRFT) were significantly enriched (Figure 5B). Meanwhile, the abundance of functional taxa of plant pathogens was significantly decreased. Among them, the functional classification of WFP-mode bacteria is rich and uniform, with significant advantages in the cycling of soil elements, such as carbon, nitrogen, sulfur, arsenic, and manganese. Compared with the WWW mode, the functional taxa under the rotation fallow modes in BS were relatively evenly distributed, especially in the WFP mode; in the RS and WWW modes, plant pathogens were significantly higher than in the rotation fallow modes (ANOVA, p < 0.05, Figure S5B). In ER, the main functional taxa were similar in composition under the five modes. The results show that different rotation fallow modes have a significant impact on the bacterial functions in BS and RS; meanwhile, the main functional taxa in ER were the result of host plant selection. Compared with the WWW mode, bacterial community functions under the crop rotation fallow modes were relatively complex, the bacterial community of plant pathogens was relatively low, and the abundance of nitrogen-cycling-related functional taxa was significantly increased.
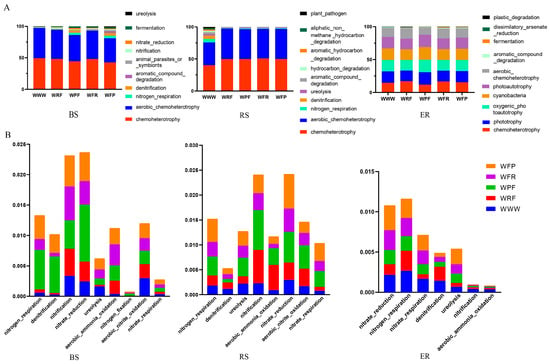
Figure 5.
Prediction of bacterial function in the root system of spring wheat under different crop rotation fallow modes. (A) The top 10 functional groups in terms of the abundance ranking for different crop rotation fallow modes. The horizontal axis represents rotation fallow modes, the vertical axis represents relative abundance, and colors represent functional taxa. (B) Nitrogen-cycling-related functional taxa in the top 100 functional taxa ranked by the abundance of different crop rotation fallow modes. The horizontal axis represents nitrogen-related functional taxa, the vertical axis represents relative abundance, and the color represents rotation fallow modes. BS: bulk soil; RS: rhizosphere; ER: endosphere.
3.5. Analysis of the Correlation between Dominant Bacteria and Environmental Factors
The differences in soil bacterial composition caused by the rotation fallow mode are closely related to soil properties (Figure 6 and Figure S6, Table 2). Through the Spearman correlation analysis of dominant bacterial phyla in three ecological niches (bulk soil, rhizosphere, and endosphere) under the five modes, it was found that Proteobacteria were significantly correlated with SMWC, TK, and total phosphorus content (TP) (p < 0.05). Actinobacteria were significantly positively correlated with TK and SMBN, while Bacteroides and Firmicutes were significantly negatively correlated with pH, SUE, and SALP. Acidobacteria were significantly positively correlated with pH, TN, CAT, SUE, and soil sucrase (SSC) (p < 0.05, Figure S6A). To further determine the key environmental factors affecting the composition of bacterial communities, we conducted RDA to analyze the correlations between environmental factors and dominant bacterial genera, and screened the environmental factors affecting the OTU levels of bacteria through the Mantel test and VIF (Table 2 and Table S5, Figure S6B). RDA analysis revealed a significant correlation between environmental factors and dominant bacterial genera (p < 0.05). The biomarker species (Figure 6) under different rotation fallow modes, such as Sphingomonas, Massilia, Segetibacter, Flavobacterium, Amycolatopsis, Bacillus, and Devosia, were significantly correlated with pH, SUE, CAT, SALP, TN, SMWC, TK, TP, SMWC, and SMBN (p < 0.05). In addition, the Mantel test revealed that BS and RS bacterial communities were more affected by soil properties, compared to ER (Table 2), and pH (BS: r = 0.445, p = 0.005; RS: r = 0.4765, p = 0.001), TN (BS: r = 0.4994, p = 0.001; RS: r = 0.3111, p = 0.041), TP (BS: r = 0.3625, p = 0.013), TK (BS: r = 0.2906, p = 0.018), SUE (BS: r = 0.4302, p = 0.002; RS: r = 0.4717, p = 0.002) CAT (BS: r = 0.2724, p = 0.025; RS: r = 0.5389, p = 0.001), and SSC (BS: r = 0.3715, p = 0.02; RS: r = 0.2992, p = 0.044) were significantly correlated with bacterial community structure. Meanwhile, the VIF results also demonstrate that pH, TK, SUE, and CAT, among other factors, are key influencing factors for the differences in sample and community distribution. In summary, our results demonstrate that the differences in soil bacterial composition caused by rotation fallow modes are closely related to soil characteristics. In particular, compared to ER, BS and RS bacterial communities are more susceptible to soil properties, and pH, TK, TN, SUE, and CAT are key environmental variables driving bacterial community distribution.
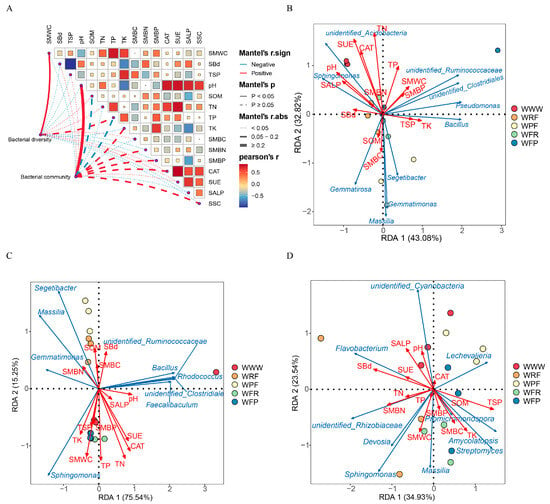
Figure 6.
Correlation analysis between environmental factors and root space (soil bulk, rhizosphere, and endosphere) bacteria in spring wheat. (A) Correlation analysis between environmental factors and bacterial alpha diversity and differences in bacterial communities. Bacterial diversity is represented by the Shannon index of bacteria, while differences in bacterial communities are represented by the beta diversity index based on Bray–Curtis index. The red line represents a positive correlation, while the blue line represents a negative correlation. The thickness of the line indicates the magnitude of the correlation coefficient, while the solid line indicates a significant correlation. SBd: soil bulk density, TSP: total soil porosity, and SMBC: soil microbial biomass carbon. (B) RDA analysis of the correlation between BS dominant bacteria and environmental variables; (C) RDA analysis of the correlation between RS dominant bacteria and environmental variables; (D) RDA analysis of the correlation between ER dominant bacteria and environmental variables. The red line represents environmental factors, and the blue line represents dominant bacteria.
Table 2.
Correlation between environmental factors and microbial communities analyzed by the mantel test.
3.6. The Impact of Key Environmental Variables on the Complexity of Microbial Networks and Nitrogen-Related Functional Taxa
The complexity of bacterial networks and changes in nitrogen-cycling-related functional taxa under different rotation fallow modes were found to be significantly correlated with key environmental variables, which mainly affect the composition of bacterial communities (Figure 7). We analyzed the impact of key environmental variables on bacterial network complexity and nitrogen-cycling-related functional taxa through univariate linear regression. The results indicate that bacterial network complexity is significantly positively correlated with TN (R2 = 0.131, p < 0.001), TK (R2 = 0.156, p < 0.001), and MFI (R2 = 0.036, p < 0.001), and negatively correlated with pH (R2 = 0144, p = 0.011); meanwhile, NRFT is significantly positively correlated with TK (R2 = 0.116, p < 0.001) and negatively correlated with pH (R2 = 0.163, p = 0.006), TN (R2 = 0.199, p = 0.002), CAT (R2 = 0.184, p = 0.003), SUE (R2 = 0.199, p = 0.002), and MFI (R2 = 0.208, p = 0.002) (see Figure 7).
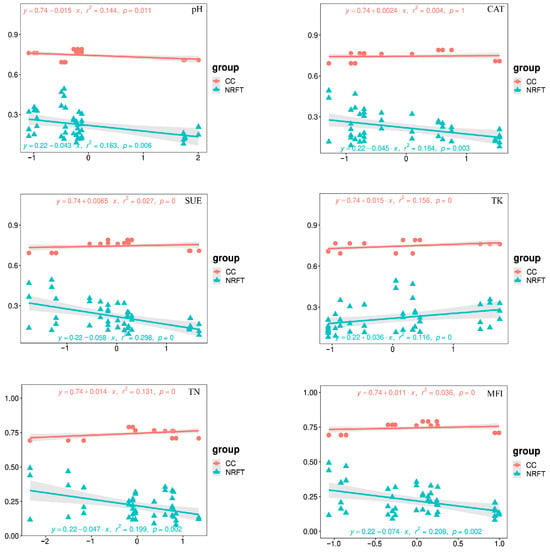
Figure 7.
Linear regression analysis of key environmental variables with the CC and NRFT. Clustering coefficient (CC) represents the complexity of the soil bacterial network; nitrogen-related functional taxa (NRFT) represents the predicted bacterial population with a nitrogen-cycling function.
4. Discussion
Rotation fallow is an important agronomic practice known to improve productivity, reduce crop diseases, enhance soil fertility, and promote yield improvement [29,30,31]. During rotation, diverse crops are introduced into the cultivation area, and their unique growth characteristics and differentiated management methods often lead to changes in the soil microenvironment, which may alter the structure of soil microbial communities and cause differential responses in particular groups. The results of this study support this viewpoint, to some extent. Compared to the continuous cropping of spring wheat, the rotation of spring wheat with potato or rape was found to present changes in the soil environment and microbial community. However, the spring wheat–fallow–potato mode seems to have a more positive impact on soil nutrient maintenance and element cycling, especially nitrogen (Figure 5). Therefore, our results not only reveal the key mechanism of improving farmland quality through crop rotation and fallow, but also provide a suitable crop rotation and fallow model for studying the region.
4.1. Reasonable Rotation Fallow Has a Positive Impact on Soil Nutrient Maintenance and Soil Function
Soil health refers to the ability of soil, as an important living system, to function within its ecosystem and land-use boundaries [32]. Soil physicochemical and biological properties are the core and comprehensive manifestations of soil health. This study indicated that, compared with the WWW rotation, the rotation fallow modes significantly increased the contents of SMWC and TK, while the difference in SOM content was not significant. The changes in SMWC and TK contents are similar to the results of Zhang et al. [29] and Liu et al. [17], while the SOM content result is similar to that in the study of Bates et al. [33]. Rotation significantly changes soil biological characteristics and basal respiration, but TC is less sensitive to different cropping systems. Except for WFP, the TN and TP contents in the WWW rotation were significantly higher than those under other rotation fallow modes, indicating that different crop rotations and crop sequences have different effects on soil physical and chemical properties, and continuous cropping can also increase the content of certain nutrients in the soil. Liu et al. [17] found that long-term soybean rotation significantly increased soil TN and TP. Similarly, Chen et al. [34] investigated soil nutrients after 20 years of the continuous cropping of Achyranthes bidentata, and found that soil nutrients—including TC, TN, and TP—were significantly increased. We also found that the soil pH of WWW was significantly higher than that under the rotation fallow modes. This is consistent with Zhong et al. [35], who reported that the long-term continuous cultivation of bananas can lead to a significant increase in soil pH. However, Liu et al. [17] found that, compared with soybean continuous cropping, the soybean–corn rotation significantly increased soil pH, which differs from our results. It is speculated that this is due to the studied crop types, as well as different regions and planting years.
Soil enzyme activity and microbial biomass are closely related to soil function, as important biological traits. Agricultural techniques, such as crop rotation and continuous cropping, as well as crop types, can alter the soil environment and affect soil enzyme activity [36]. In this study, it was found that WFP significantly increased soil urease activity, soil microbial biomass nitrogen, and soil microbial biomass phosphorus content. Gonnety et al. [37] found that soil enzyme activity decreased significantly when corn was planted continuously for a long period of time, compared with that when Chromolaena odorata was fallow for 2 years, and it was believed that the difference was caused by different land-use modes. Borase et al. [38] found that crop rotation significantly affects soil enzyme activity and microbial biomass. Compared with the maize–wheat rotation, pigeonpea–wheat significantly increased SMBC and SMBN contents, as well as alkaline phosphatase activity. In this study, WWW significantly increased the activities of soil catalase and alkaline phosphatase (Figure 1). This differs from the research results of Liu et al. [39], who found that the continuous cultivation of American ginseng caused a decrease in soil alkaline phosphatase activity; the reason for this change is that changes in soil pH and salinity cause changes in the microbial community structure. In addition, we observed a highly significant correlation between soil pH and alkaline phosphatase and catalase (Figure 6A). Compared with the rotation fallow mode, soil pH significantly increased under the WWW rotation. Further research is needed to determine whether this is the reason for the increase in catalase and alkaline phosphatase under the WWW rotation. Malobane et al. [40] argued that crop rotation only affects acid phosphatase and, compared to sorghum—fallow—sorghum, sorghum—vetch—sorghum enhanced acid phosphatase activity. Yusuf et al. [41] found that the crop rotation sequence has a significant impact on soil microbial biomass carbon and nitrogen. Soybean crop rotation can significantly increase the soil nitrogen content and increase crop yield, mainly attributed to the growth characteristics of nitrogen-fixing bacteria, which are enriched in legume crops. In addition, the introduction of soil secretions or residues by plants themselves, as well as their preference for nutrients, can directly alter the soil environment and affect microbial content [42].
In summary, different rotation fallow modes can affect soil physicochemical and biological properties. The imbalance of soil nutrients caused by continuous cropping can be improved through crop rotation and fallow, while improper crop combinations or rotation sequences may be detrimental to the maintenance of soil nutrients. This is mainly attributed to the diversity of crop rotation and management methods, which together regulate the multifunctional aspects of soil [43]. Therefore, selecting a reasonable crop rotation and fallow system can have a positive impact on both soil nutrient maintenance and soil function.
4.2. Rotation Fallow Mode Can Affect the Diversity of Bacterial Communities, but Its Effect Is Weaker Than That of Ecological Niches
The soil microbial community is influenced by the type of host plant and its ecological niche [8]. The results of this study indicate that the rotation fallow mode does not significantly affect bacterial alpha diversity (Figure 2), while niche significantly affects bacterial alpha diversity, with the Shannon index and chao1 index both being in the order of Bulk soil = Rhizosphere > Endosphere. These research results are similar to those of previous studies [8,44,45,46]. The plant microbiome is largely formed by host selection, affected by factors such as the genotype and compartment niche. Host plants directly recruit the rhizosphere microbiome through plant signals and root exudates, and tend to select micro-organisms that can penetrate the root tissue to form endophytes. The diversity of the endosphere is usually lower than that of the rhizosphere and bulk soil. Similarly, based on the NMDS of the bacterial Bray–Curtis distance PCoA in different ecological niches and bacterial Bray–Curtis distance between rotation and fallow modes, we observed that ecological niches are the main driving factor of bacterial community structure, while rotation and fallow modes are secondary factors. In addition, in the analysis of dominant bacterial phyla in different spatial positions under the crop rotation and fallow modes, it was found that, compared to the dominant bacterial phyla in the rhizosphere and bulk soil, the distribution of endosphere bacterial phyla under different crop rotation and fallow modes was more uniform. This result can also be explained by the strong driving force of the niche in community construction, that is, the selection effect of the host plant (i.e., spring wheat) allows host-preferred bacteria and immune-resistant bacteria to parasitize within its roots. In summary, these results indicate that the contribution of ecological niches shaped by host plants to bacterial community construction is greater than that of crop rotation and fallow management strategies.
4.3. Reasonable Rotation Fallow Cultivation Is Beneficial for the Colonization and Growth of Beneficial Micro-organisms
The decomposition and release of soil nutrients, energy conversion, and material cycling are driven by soil micro-organisms and, thus, are closely related to community composition and metabolic functions [47]. The results of this study indicate that Proteobacteria, Actinobacteria, and Acidobacteria are dominant bacterial phyla under rotation fallow modes, similar to previous studies on the wheat microbiome [46,48,49]. This may be attributed to these bacterial phyla being the most common bacterial phyla in soil, which are widely present in various agricultural systems and soil types [50,51]. In particular, Bacteroidetes, Firmicutes, and Verrucomicrobia are eutrophic bacteria [52], while Acidobacteria are oligotrophic bacteria [53]. Compared with the continuous cropping mode, the relative abundance of Bacteroidetes, Firmicutes, and Verrucomicrobia was increased under the rotation fallow modes, while the relative abundance of Acidobacteria decreased (Figure 3), suggesting that the rotation fallow mode leads to a higher soil nutrient content, further confirming our results for soil nutrients under different modes. In addition, LEfSe analysis revealed that, compared with continuous cropping, the rotation fallow modes, especially WFP, significantly increased the types and numbers of main biomarkers (Figure 4), playing an important role in the cycling of soil carbon, nitrogen, phosphorus, sulfur, and other elements; ecological restoration; pathogen antagonism; and promotion of plant growth. For example, Massilia [54] can synthesize various secondary metabolites and enzymes, with functions such as phosphorus solubilization, phenanthrene degradation, and heavy metal tolerance. Clostridia [55] participates in the cycling of soil carbon-source substances, producing organic acids to inhibit the growth of soil pathogens; Phylobacterium [56] participates in soil phosphorus cycling, assisting crops in resisting abiotic stress; and Pedobacter [57] regulates wheat growth and development through synthesizing indole derivatives and thiobacilli to resist the wheat stripe rust pathogen. Flavobacteria [58] can be used as an effective biocontrol bacterial group for the prevention and control of tomato wilt disease. This supports the view that crop rotation fallow is an effective measure to overcome the obstacles associated with continuous cropping. However, although the rotation fallow mode has a rich variety of beneficial biomarker species, there are differences in the types and numbers of biomarker species between different modes, (e.g., WFP and WFR). The reason for this is that the residual effects (plant tissue or root exudates) of different crops in the rotation system can alter the soil microenvironment and stimulate the growth of specific micro-organisms; the effects of this stimulation may also affect the assembly of subsequent soil microbial communities [59,60]. Similarly, differences in crop cultivation can also lead to similar results. In addition to the legacy effects of crop cultivation, previous crop management strategies (e.g., cultivation, fertilization, and pesticides) may also have legacy effects that affect the microbial community composition of subsequent crops. As Kan et al. [61] found in the wheat–corn rotation system, compared to the tillage and stubble treatments, the no-tillage and stubble treatments of wheat straw led to higher organic carbon content input into the soil. The changes in carbon source types and contents are closely related to soil microbial composition [62]. It is worth noting that the contribution of legacy effects of previous crop management strategies to the microbial assembly of subsequent crops is not yet clear, although recent research reports have indicated that crop rotation has a greater impact on the soil microbial assembly than fertilization [63]. Overall, rotation fallow can affect microbial community structure through the legacy effects of crop and management practices, and, thus, selecting appropriate crop rotation and planting practices is beneficial for the growth of beneficial bacterial colonies.
4.4. Rotation Fallow Enhances the Complexity, Stability, and Relative Abundance of Nitrogen-Cycling Functional Bacterial Taxa in Bacterial Communities
As is well-known, soil microbial communities can regulate various ecosystem functions, and ecosystem processes often occur through complex interactions between microbial communities [64,65]. This study indicates that, compared with the WWW rotation, the rotation fallow modes exhibited a more complex and compact microbial network structure (Figure 5). Consistent with previous research findings, diversified crop rotation can enhance the complexity of underground microbial networks [43]. This is mainly attributed to the introduction of diverse crops that provide diverse litter and root exudates, thus directly or indirectly affecting the association of soil microbial communities [43,66]. Different host crops, such as potatoes and rape, recruit specific microbial groups through root exudates and block other groups. Diversified microbial environments can affect microbial community structure and alter microbial interactions [67,68]. In addition, diverse crop rotation systems can generate legacy effects and provide favorable niches for specific microbial communities through root action, leading to greater niche sharing and interactions among microbial communities [69].
The stability of microbial networks can characterize the ability of micro-organisms to resist external interference [70]. The results of this study reveal that, compared with the continuous cropping mode, crop rotation fallow is characterized by a lower degree of modularity, tending to form a large network module and an increase in negative correlation co-occurrence (Figure S4). Some studies have shown that environmental pressure makes the microbial community network unstable. In habitats with higher environmental pressure, microbial diversity is reduced, and positive correlation co-occurrence is predominant [71]. Our results indicate that the long-term continuous cropping of spring wheat may cause certain environmental stresses or nutrient limitations for micro-organisms. Micro-organisms can only survive by changing their survival strategies through niche differentiation, thus forming multiple smaller, positively correlated modules.
Changes in network complexity and stability affect the functional structure of microbial communities and the functional processes of ecosystems. Ecological network changes caused by crop rotation and fallow may affect the ability of ecosystems to provide stable ecological services. Through the FAPROTAX functional prediction, we found that the bacterial taxa related to soil nitrogen-cycling functions were significantly enriched during rotation fallow treatment compared to the WWW rotation (Figure 5), which is consistent with previous research results. Hao et al. [72] found that crop diversification (rotation or intercropping) significantly increased the abundance of soil nitrogen-cycling functional bacteria. The main reason for this phenomenon may also be attributed to the legacy effects of crops. Crop diversification increases both aboveground and underground biomass, and abundant organic matter provides resources for microbial growth. Surprisingly, soil TN content is not always positively correlated with soil microbial nitrogen-cycling-related taxa (Figure 6), and, compared with the WWW mode, other rotation fallow modes, except for WFP, presented significantly decreased levels of such groups. It can be speculated that the nitrogen-cycling taxa increased by rotation fallow may promote processes, such as nitrogen ammonification and nitrification in the input soil, which are beneficial for plant absorption and utilization. In addition, different microbial communities that mediate the same nitrogen-cycling process have different activities and environmental adaptability [73], and are influenced by variables such as crop type, planting methods, duration of diversification, and soil texture [72]. Further research is needed to investigate the impact of increased nitrogen-cycling functional taxa on soil nitrogen content.
4.5. Key Environmental Variables Affecting Microbial Community Assembly
Microbial community assembly is influenced by two ecological processes: deterministic and stochastic [74]. The results of this study indicate that the endosphere in spring wheat roots is mainly influenced by the selectivity of the host plant, while RS and BS are more susceptible to environmental factors, especially BS (Figure S6). This may imply that deterministic processes are the main ecological processes that affect the inter-niche microbial community structure in spring wheat soil areas, rather than random processes. In addition, as a unique ecological niche, the rhizosphere is influenced by both environmental and biological variables, which, to some extent, can weaken the effect of spatial heterogeneity on bacteria. In this study, rhizosphere bacteria were mainly influenced by pH and biological traits (CAT and SUE), while non-rhizosphere soil was also influenced by soil nutrients (e.g., TN, TP, and TK), in addition to pH and soil biological traits. This is similar to the results reported in previous research [75]. These results indicate that pH, TK, TN, SUE, and CAT play key roles in driving the distribution of bacterial communities. Studies have also indicated that SOC is another important factor affecting bacterial community assembly, which is significantly correlated with the soil aggregation structure and can provide nutritional resources and a favorable growth environment for micro-organisms [76]. In our study, soil organic matter content did not significantly affect bacterial community structure. This may be because the five rotation fallow modes did not cause significant differences in the soil organic matter content, which may be influenced by soil fertility conditions and planting cycles. Further research is needed to determine the specific reasons for these observations.
The main ecological functions of soil are nutrient cycling and a resistance to environmental stresses. We observed a positive correlation between bacterial network complexity and the soil multifunctionality index, which is consistent with previous studies [77,78]. The main reason for this is that complex communities have advantages in terms of resource utilization and information transmission, and bacteria with similar environments and resource preferences have strong interactions, making information transmission more efficient. In our study, TN, TP, and TK were significantly positively correlated with bacterial complexity (Figure 7), indicating that rich nutrition may help to enhance interactions within the microbiome. However, there was a negative correlation between NRFT and the soil ecological function index (MFI). Does this mean that a significant increase in certain specific microbial groups may affect the complexity of the microbial network and hinder soil ecological multifunctionality? We need to further explore this question in future research. In summary, compared to continuous cropping, rotation fallow tillage complicates the soil microbial network and increases the abundance of nitrogen-cycling-related microbial communities, which may enhance nitrogen availability, maintain nutrient cycling, and improve agricultural productivity.
In summary, we propose a model diagram of the impact mechanism of crop rotation and fallow mode on soil bacteria (Figure 8). The crop rotation fallow mode improved soil moisture and soil aeration, to a certain extent, through the “legacy effect” due to the rotation of different crops, thus promoting a neutral and stable soil pH, which, in turn, maintains enzyme activity in a benign state, thereby increasing soil microbial biomass, carbon, nitrogen, phosphorus, total nitrogen, phosphorus, potassium, and other nutrients, as well as promoting the efficient utilization of soil nutrients, which is conducive to crop growth. In addition, the beneficial bacteria left behind can also colonize in the soil, increase microbial diversity, promote the cycling of elements, such as C and N in the soil, and assist crops in resisting pathogenic hazards. And the well-growing crops can provide a sufficient environment for micro-organisms through root secretions and residues, enriching special micro-organisms, improving the abundance of beneficial micro-organisms, activating soil nutrients, and then realizing the combined effect of land enrichment.
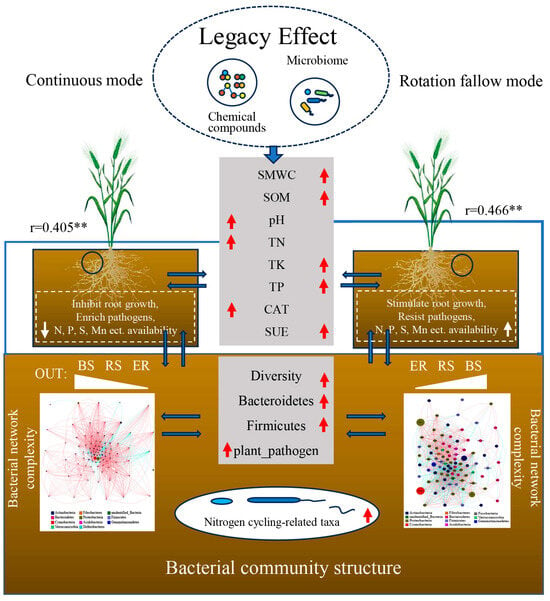
Figure 8.
Model diagram of the impact mechanism of the crop rotation fallow mode on soil bacteria. The gray modules in the figure represent the soil properties and bacterial community structure indicators detected in this study. The dashed module represents the inferred results based on this study. The red and white arrows indicate the up or down regulation indicators in the crop rotation fallow mode or continuous cropping mode. The left side refers to the continuous cropping mode, and the right side refers to the crop rotation fallow mode. The arrow pointing upward indicates the upregulation, and the arrow pointing downward indicates the downregulation. The blue arrow indicates the relationship between the soil, bacteria, and plants. The blue line represents the correlation between key environmental variables (pH and TN) and bacterial community structure based on the Mantel test analysis. The thickness of the line represents the magnitude of the correlation, and the correlation coefficient r value is expressed as the mean correlation coefficient between environmental variables and bulk soil and rhizosphere bacterial communities, **: p < 0.05. BS: bulk soil; RS: rhizosphere; ER: endosphere.
5. Conclusions
Compared with the WWW mode, reasonable crop rotation fallow modes were found to be beneficial for conserving soil moisture, maintaining soil nutrient balance, and ensuring that the soil pH tends to be neutral. The ecological niche and rotation fallow mode both affected the relative abundance of dominant bacteria. The relative abundance of beneficial bacterial phyla, such as Bacteroidetes, Firmicutes, and Verrucomicrobia, was significantly increased under the rotation fallow mode, and the complexity and stability of bacterial networks and abundance of nitrogen-cycling related functional taxa were significantly improved. The differences in soil bacterial composition due to the rotation fallow mode were closely related to soil characteristics. Compared to ER, BS and RS bacterial communities were more susceptible to soil properties, and soil pH was a key environmental variable driving the distribution of bacterial communities. In summary, compared to continuous cropping, the rotation fallow mode complexity of the soil microbial network and abundance of nitrogen-cycling-related microbial communities increase, which may enhance nitrogen availability, maintain nutrient cycling, and improve agricultural productivity. In particular, the spring wheat–fallow–potato mode can be used as a suitable rotation fallow model for maintaining soil fertility in the western foothills of the Greater Khingan Mountains.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14071456/s1, Figure S1: Analysis of soil physicochemical and biological properties under different rotation fallow modes; Figure S2: Non-metric multidimensional scaling (NMDS) analysis of sample differences under different rotation fallow modes with the same root spatial position; Figure S3: Analysis of significant differences in biomarkers under different rotation fallow modes with the same root spatial position; Figure S4: Frequency analysis of positive and negative correlations bacteria in the top 100 bacterial genera in abundance rankings under different rotation fallow modes; Figure S5: Prediction and analysis of the main dominant bacterial functions under different rotation fallow modes with the same root spatial position; Figure S6: Environmental factor correlation analysis; Table S1: Diversity index of bulk soil, rhizospheres and endosphere bacteria in spring wheat under different rotation fallow modes; Table S2: Scheirer–Ray–Hare test analysis of variance table; Table S3: Mean relative abundance of major bacteria under different crop rotation fallow modes; Table S4: Topological properties of bacterial networks under different rotation fallow modes; Table S5: Correlation between environmental factors and microbial communities analyzed by mantel test; Figure S6: Environmental factor correlation analysis.
Author Contributions
Conceptualization, Z.L. and X.Z. (Xiaoqing Zhao); methodology, X.Z. (Xiaoqing Zhao) and G.S.; software, G.S. and J.F.; validation, G.S., J.F., S.W., X.Z. (Xiangqian Zhang) and Z.L.; formal analysis, G.S., Y.C. and S.S.; investigation, G.S., J.F., S.W., X.Z. (Xiaoqing Zhao) and Z.L.; resources, G.S., J.F., J.W. (Jianguo Wang), L.Z., S.S. and J.W. (Jianhui Wu); data curation, G.S., J.F. and F.Z.; writing—original draft preparation, G.S.; writing—review and editing, G.S. and J.F.; visualization, G.S. and X.Z. (Xiaoqing Zhao); supervision, G.S.; project administration, X.Z. (Xiaoqing Zhao); funding acquisition, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2022YFD1500901-5), the National Natural Science Foundation of China (U23A20195, 31860356, 32260457), the Inner Mongolia Autonomous Region Natural Science Foundation Project Key Projects (2022ZD13), the Inner Mongolia Agriculture and Animal Husbandry Innovation Fund Project (2022CXJJN08 and 2022CXJJM04), and the Scientific and Technological Projects of Grassland Talents in Inner Mongolia Autonomous Region and the Leading Talent Project of “Science and Technology Leading Talent Team Project of Inner Mongolia Autonomous Region” (2022LJRC0010).
Data Availability Statement
The bacterial raw DNA sequences used in this study were deposited in the Sequence Read Achieve (SRA) of the NCBI database under the accession number PRJNA1104604 for open access.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Luo, G.; Wang, T.; Li, K.; Li, L.; Zhang, J.; Guo, S.; Ling, N.; Shen, Q. Historical Nitrogen Deposition and Straw Addition Facilitate the Resistance of Soil Multifunctionality to Drying-Wetting Cycles. Appl. Environ. Microbiol. 2019, 85, e02251-18. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.S.; Isbell, F.; Fujii, S.; Makoto, K.; Matsuoka, S.; Osono, T. Low Multifunctional Redundancy of Soil Fungal Diversity at Multiple Scales. Ecol. Lett. 2016, 19, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; de Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root Microbiota Drive Direct Integration of Phosphate Stress and Immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and Ecological Function of the Root Microbiome across Angiosperm Plant Species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Wang, Y.; Huang, Y.; Xu, J.; Zhang, P.; Wang, N.; Liu, X.; Chu, H.; Liu, G.; Jiang, H.; et al. Taxonomic Structure and Functional Association of Foxtail Millet Root Microbiome. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Garbeva, P.; van Elsas, J.D.; van Veen, J.A. Rhizosphere Microbial Community and Its Response to Plant Species and Soil History|Plant and Soil. Available online: https://link.springer.com/article/10.1007/s11104-007-9432-0 (accessed on 14 April 2024).
- Schlemper, T.R.; Leite, M.F.A.; Lucheta, A.R.; Shimels, M.; Bouwmeester, H.J.; van Veen, J.A.; Kuramae, E.E. Rhizobacterial Community Structure Differences among Sorghum Cultivars in Different Growth Stages and Soils. FEMS Microbiol. Ecol. 2017, 93, fix096. [Google Scholar] [CrossRef]
- Liu, H.; Pan, F.; Han, X.; Song, F.; Zhang, Z.; Yan, J.; Xu, Y. A Comprehensive Analysis of the Response of the Fungal Community Structure to Long-Term Continuous Cropping in Three Typical Upland Crops. J. Integr. Agric. 2020, 19, 866–880. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Arafat, Y.; Lin, W. Studies on Fungal Communities and Functional Guilds Shift in Tea Continuous Cropping Soils by High-Throughput Sequencing. Ann. Microbiol. 2020, 70, 7. [Google Scholar] [CrossRef]
- Bansal, S.; Yin, X.; Schneider, L.; Sykes, V.; Jagadamma, S.; Lee, J. Carbon Footprint and Net Carbon Gain of Major Long-Term Cropping Systems under No-Tillage. J. Environ. Manag. 2022, 307, 114505. [Google Scholar] [CrossRef] [PubMed]
- Haichar, F.E.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Heulin, T.; Achouak, W. Plant Host Habitat and Root Exudates Shape Soil Bacterial Community Structure. ISME J. 2008, 2, 1221–1230. [Google Scholar] [CrossRef]
- Zuppinger-Dingley, D.; Schmid, B.; Petermann, J.S.; Yadav, V.; De Deyn, G.B.; Flynn, D.F.B. Selection for Niche Differentiation in Plant Communities Increases Biodiversity Effects. Nature 2014, 515, 108–111. [Google Scholar] [CrossRef]
- Venter, Z.S.; Jacobs, K.; Hawkins, H.-J. The Impact of Crop Rotation on Soil Microbial Diversity: A Meta-Analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, J.; Yu, Z.; Yao, Q.; Li, Y.; Liang, A.; Zhang, W.; Mi, G.; Jin, J.; Liu, X.; et al. Long-Term Continuous Cropping of Soybean Is Comparable to Crop Rotation in Mediating Microbial Abundance, Diversity and Community Composition. Soil Tillage Res. 2020, 197, 104503. [Google Scholar] [CrossRef]
- Pershina, E.V.; Ivanova, E.A.; Korvigo, I.O.; Chirak, E.L.; Sergaliev, N.H.; Abakumov, E.V.; Provorov, N.A.; Andronov, E.E. Investigation of the Core Microbiome in Main Soil Types from the East European Plain. Sci. Total Environ. 2018, 631–632, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Yu, Z.; Shi, Y.; Chu, H.; Jin, J.; Liu, X.; Wang, G. High Throughput Sequencing Analysis of Biogeographical Distribution of Bacterial Communities in the Black Soils of Northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Bai, L.; Wang, Y.; Li, Y.; Zhang, X.; Lu, Z.; Zhang, D.; Sun, F.; Zhao, X. Changes in the Microbial Community in Maize (Zea mays L.) Root Spatial Structure Following Short-Term Nitrogen Application. ACS Omega 2023, 8, 208–218. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S rRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, G.; Wang, Y.; Fei, J.; Xiangmin, R.; Peng, J.; Tian, C.; Zhang, Y. Crop Rotation-Driven Change in Physicochemical Properties Regulates Microbial Diversity, Dominant Components, and Community Complexity in Paddy Soils. Agric. Ecosyst. Environ. 2023, 343, 108278. [Google Scholar] [CrossRef]
- Sun, L.; Wang, S.; Narsing Rao, M.P.; Shi, Y.; Lian, Z.-H.; Jin, P.-J.; Wang, W.; Li, Y.-M.; Wang, K.-K.; Banerjee, A.; et al. The Shift of Soil Microbial Community Induced by Cropping Sequence Affect Soil Properties and Crop Yield. Front. Microbiol. 2023, 14, 1095688. [Google Scholar] [CrossRef]
- Meier, M.A.; Lopez-Guerrero, M.G.; Guo, M.; Schmer, M.R.; Herr, J.R.; Schnable, J.C.; Alfano, J.R.; Yang, J. Rhizosphere Microbiomes in a Historical Maize-Soybean Rotation System Respond to Host Species and Nitrogen Fertilization at the Genus and Subgenus Levels. Appl. Environ. Microbiol. 2021, 87, e0313220. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil Health and Sustainability: Managing the Biotic Component of Soil Quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the Global Distribution of Dominant Archaeal Populations in Soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, J.; Wu, L.; Lin, S.; Wang, J.; Li, Z.; Zhang, Z.; Lin, W. Effects of Continuous Monoculture of Achyranthes bidentata on Microbial Community Structure and Functional Diversity in Soil. Allelopath. J. 2015, 36, 197–211. [Google Scholar]
- Zhong, S.; Mo, Y.; Guo, G.; Zeng, H.; Jin, Z. Effect of Continuous Cropping on Soil Chemical Properties and Crop Yield in Banana Plantation. J. Agric. Sci. Technol. 2014, 16, 239–250. [Google Scholar]
- Qin, L.; Freeman, C.; Jia, X.; Zhang, Z.; Liu, B.; Zhang, S.; Jiang, M. Microbial Enzyme Activity and Stoichiometry Signal the Effects of Agricultural Intervention on Nutrient Cycling in Peatlands. Ecol. Indic. 2021, 122, 107242. [Google Scholar] [CrossRef]
- Gonnety, J.T.; Assémien, E.F.L.; Guéi, A.M.; N’Dri, A.A.; Djina, Y.; Koné, A.W.; Tondoh, J.E. Effect of Land-Use Types on Soil Enzymatic Activities and Chemical Properties in Semi-Deciduous Forest Areas of Central-West Côte d’Ivoire. Biotechnol. Agron. Société Environ. 2012, 16, 478–485. [Google Scholar]
- Borase, D.N.; Nath, C.P.; Hazra, K.K.; Senthilkumar, M.; Singh, S.S.; Praharaj, C.S.; Singh, U.; Kumar, N. Long-Term Impact of Diversified Crop Rotations and Nutrient Management Practices on Soil Microbial Functions and Soil Enzymes Activity. Ecol. Indic. 2020, 114, 106322. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Z.; Niu, J.; Dang, K.; Zhang, S.; Wang, S.; Wang, Z. Changes in Physicochemical Properties, Enzymatic Activities, and the Microbial Community of Soil Significantly Influence the Continuous Cropping of Panax quinquefolius L. (American Ginseng). Plant Soil 2021, 463, 427–446. [Google Scholar] [CrossRef]
- Malobane, M.E.; Nciizah, A.D.; Nyambo, P.; Mudau, F.N.; Wakindiki, I.I.C. Microbial Biomass Carbon and Enzyme Activities as Influenced by Tillage, Crop Rotation and Residue Management in a Sweet Sorghum Cropping System in Marginal Soils of South Africa. Heliyon 2020, 6, e05513. [Google Scholar] [CrossRef]
- Yusuf, A.A.; Abaidoo, R.C.; Iwuafor, E.N.O.; Olufajo, O.O.; Sanginga, N. Rotation Effects of Grain Legumes and Fallow on Maize Yield, Microbial Biomass and Chemical Properties of an Alfisol in the Nigerian Savanna. Agric. Ecosyst. Environ. 2009, 129, 325–331. [Google Scholar] [CrossRef]
- Chinnadurai, C.; Gopalaswamy, G.; Balachandar, D. Long Term Effects of Nutrient Management Regimes on Abundance of Bacterial Genes and Soil Biochemical Processes for Fertility Sustainability in a Semi-Arid Tropical Alfisol. Geoderma 2014, 232–234, 563–572. [Google Scholar] [CrossRef]
- Yang, X.; Hu, H.-W.; Yang, G.-W.; Cui, Z.-L.; Chen, Y.-L. Crop Rotational Diversity Enhances Soil Microbiome Network Complexity and Multifunctionality. Geoderma 2023, 436, 116562. [Google Scholar] [CrossRef]
- Ji, N.; Liang, D.; Clark, L.V.; Sacks, E.J.; Kent, A.D. Host Genetic Variation Drives the Differentiation in the Ecological Role of the Native Miscanthus Root-Associated Microbiome. Microbiome 2023, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, Y.S.; Zhan, Y.; Zhang, Z.; Liu, Y.; Wei, Y.; Xu, T.; Li, J. Tomato Microbiome under Long-Term Organic and Conventional Farming. iMeta 2022, 1, e48. [Google Scholar] [CrossRef] [PubMed]
- Gdanetz, K.; Trail, F. The Wheat Microbiome Under Four Management Strategies, and Potential for Endophytes in Disease Protection. Phytobiomes J. 2017, 1, 158–168. [Google Scholar] [CrossRef]
- Shi, Y.; Delgado-Baquerizo, M.; Li, Y.; Yang, Y.; Zhu, Y.-G.; Peñuelas, J.; Chu, H. Abundance of Kinless Hubs within Soil Microbial Networks Are Associated with High Functional Potential in Agricultural Ecosystems. Environ. Int. 2020, 142, 105869. [Google Scholar] [CrossRef] [PubMed]
- Donn, S.; Kirkegaard, J.A.; Perera, G.; Richardson, A.E.; Watt, M. Evolution of Bacterial Communities in the Wheat Crop Rhizosphere. Environ. Microbiol. 2015, 17, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct Soil Microbial Diversity under Long-Term Organic and Conventional Farming. ISME J. 2014, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-Based Assessment of Bacterial Community Structure along Different Management Types in German Forest and Grassland Soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef]
- Shen, C.; Liang, W.; Shi, Y.; Lin, X.; Zhang, H.; Wu, X.; Xie, G.; Chain, P.; Grogan, P.; Chu, H. Contrasting Elevational Diversity Patterns between Eukaryotic Soil Microbes and Plants. Ecology 2014, 95, 3190–3202. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Semenov, M.; Yao, F.; Ye, J.; Bu, R.; Ma, R.; Lin, J.; Kurganova, I.; Wang, X.; et al. Temperature Sensitivity of SOM Decomposition Is Linked with a K-selected Microbial Community. Glob. Chang. Biol. 2021, 27, 2763–2779. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative Metagenomic, Phylogenetic and Physiological Analyses of Soil Microbial Communities across Nitrogen Gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; He, X.; Baer, M.; Beirinckx, S.; Tian, T.; Moya, Y.A.T.; Zhang, X.; Deichmann, M.; Frey, F.P.; Bresgen, V.; et al. Plant Flavones Enrich Rhizosphere Oxalobacteraceae to Improve Maize Performance under Nitrogen Deprivation. Nat. Plants 2021, 7, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lu, Y. Decomposition of Soil Polymeric Organic Matter by Bacteroidetes and Clostridia:Progress and Perspectives. Microbiol. China 2022, 49, 1147–1157. [Google Scholar] [CrossRef]
- Breitkreuz, C.; Buscot, F.; Tarkka, M.; Reitz, T. Shifts Between and Among Populations of Wheat Rhizosphere Pseudomonas, Streptomyces and Phyllobacterium Suggest Consistent Phosphate Mobilization at Different Wheat Growth Stages Under Abiotic Stress. Front. Microbiol. 2019, 10, 3109. [Google Scholar] [CrossRef]
- Zhang, L.-L.; Gan, L.-Z.; Xu, Z.-B.; Yang, F.; Li, Y.; Fan, X.-L.; Liu, X.-F.; Tian, Y.-Q.; Dai, Y.-M. Pedobacter chitinilyticus Sp. Nov., a Chitin-Degrading Bacterium Isolated from Wheat Leaf Tissue. Int. J. Syst. Evol. Microbiol. 2018, 68, 3713–3719. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.-J.; Kong, H.G.; Choi, K.; Kwon, S.-K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere Microbiome Structure Alters to Enable Wilt Resistance in Tomato. Nat. Biotechnol. 2018, 36, 1100–1109. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Zhang, Y.; Chen, G.; Li, Z.; Zhang, M. Straw Return Drives Soil Microbial Community Assemblage to Change Metabolic Processes for Soil Quality Amendment in a Rice-Wheat Rotation System. Soil Biol. Biochem. 2023, 185, 109131. [Google Scholar] [CrossRef]
- Hong, S.; Yuan, X.; Yang, J.; Yang, Y.; Jv, H.; Li, R.; Jia, Z.; Ruan, Y. Selection of Rhizosphere Communities of Diverse Rotation Crops Reveals Unique Core Microbiome Associated with Reduced Banana Fusarium Wilt Disease. New Phytol. 2023, 238, 2194–2209. [Google Scholar] [CrossRef]
- Kan, Z.-R.; Chen, Z.; Wei, Y.-X.; Virk, A.L.; Bohoussou, Y.N.; Lal, R.; Zhao, X.; Zhang, H.-L. Contribution of Wheat and Maize to Soil Organic Carbon in a Wheat-Maize Cropping System: A Field and Laboratory Study. J. Appl. Ecol. 2022, 59, 2716–2729. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Zhang, F. Abundant and Rare Bacteria Possess Different Diversity and Function in Crop Monoculture and Rotation Systems across Regional Farmland. Soil Biol. Biochem. 2022, 171, 108742. [Google Scholar] [CrossRef]
- Xie, Y.; Ouyang, Y.; Han, S.; Se, J.; Tang, S.; Yang, Y.; Ma, Q.; Wu, L. Crop Rotation Stage Has a Greater Effect than Fertilisation on Soil Microbiome Assembly and Enzymatic Stoichiometry. Sci. Total Environ. 2022, 815, 152956. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate Warming Enhances Microbial Network Complexity and Stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Melillo, J.M.; Frey, S.D.; DeAngelis, K.M.; Werner, W.J.; Bernard, M.J.; Bowles, F.P.; Pold, G.; Knorr, M.A.; Grandy, A.S. Long-Term Pattern and Magnitude of Soil Carbon Feedback to the Climate System in a Warming World. Science 2017, 358, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root Exudate Cocktails: The Link between Plant Diversity and Soil Microorganisms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, M.D.; Grandy, A.S.; Tiemann, L.K.; Weintraub, M.N. Crop Rotation Complexity Regulates the Decomposition of High and Low Quality Residues. Soil Biol. Biochem. 2014, 78, 243–254. [Google Scholar] [CrossRef]
- Xiong, W.; Jousset, A.; Li, R.; Delgado-Baquerizo, M.; Bahram, M.; Logares, R.; Wilden, B.; de Groot, G.A.; Amacker, N.; Kowalchuk, G.A.; et al. A Global Overview of the Trophic Structure within Microbiomes across Ecosystems. Environ. Int. 2021, 151, 106438. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Cong, W.-F.; Bezemer, T.M. Legacies at Work: Plant-Soil-Microbiome Interactions Underpinning Agricultural Sustainability. Trends Plant Sci. 2022, 27, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jin, L.; Zhang, C.; Li, S.; Zhou, T.; Hua, Z.; Wang, L.; Ji, S.; Wang, Y.; Gan, Y.; et al. Destabilized Microbial Networks with Distinct Performances of Abundant and Rare Biospheres in Maintaining Networks under Increasing Salinity Stress. iMeta 2023, 2, e79. [Google Scholar] [CrossRef]
- Hernandez, D.J.; David, A.S.; Menges, E.S.; Searcy, C.A.; Afkhami, M.E. Environmental Stress Destabilizes Microbial Networks. ISME J. 2021, 15, 1722–1734. [Google Scholar] [CrossRef]
- Hao, J.; Feng, Y.; Wang, X.; Yu, Q.; Zhang, F.; Yang, G.; Ren, G.; Han, X.; Wang, X.; Ren, C. Soil Microbial Nitrogen-Cycling Gene Abundances in Response to Crop Diversification: A Meta-Analysis. Sci. Total Environ. 2022, 838, 156621. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Wang, F.; Hu, C.; Liu, B. Metagenomics Reveals Taxon-Specific Responses of the Nitrogen-Cycling Microbial Community to Long-Term Nitrogen Fertilization. Soil Biol. Biochem. 2021, 156, 108214. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling Mechanisms That Mediate the Balance between Stochastic and Deterministic Processes in Microbial Succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Li, J.; Luan, X.; Yang, L.; Chen, W.; Ma, X.; Yang, Z.; Shangguan, Z. Biogeographical Patterns of Rhizosphere Microbial Communities in Robinia pseudoacacia Forests along a North–South Transect in the Loess Plateau. Geoderma 2023, 435, 116516. [Google Scholar] [CrossRef]
- Zhang, W.; Munkholm, L.J.; Liu, X.; An, T.; Xu, Y.; Ge, Z.; Xie, N.; Li, A.; Dong, Y.; Peng, C.; et al. Soil Aggregate Microstructure and Microbial Community Structure Mediate Soil Organic Carbon Accumulation: Evidence from One-Year Field Experiment. Geoderma 2023, 430, 116324. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, Q.; Zhu, H.; Reich, P.B.; Banerjee, S.; van der Heijden, M.G.A.; Sadowsky, M.J.; Ishii, S.; Jia, X.; Shao, M.; et al. Erosion Reduces Soil Microbial Diversity, Network Complexity and Multifunctionality. ISME J. 2021, 15, 2474–2489. [Google Scholar] [CrossRef]
- Bender, S.F.; Schulz, S.; Martínez-Cuesta, R.; Laughlin, R.J.; Kublik, S.; Pfeiffer-Zakharova, K.; Vestergaard, G.; Hartman, K.; Parladé, E.; Römbke, J.; et al. Simplification of Soil Biota Communities Impairs Nutrient Recycling and Enhances Above- and Belowground Nitrogen Losses. New Phytol. 2023, 240, 2020–2034. [Google Scholar] [CrossRef]
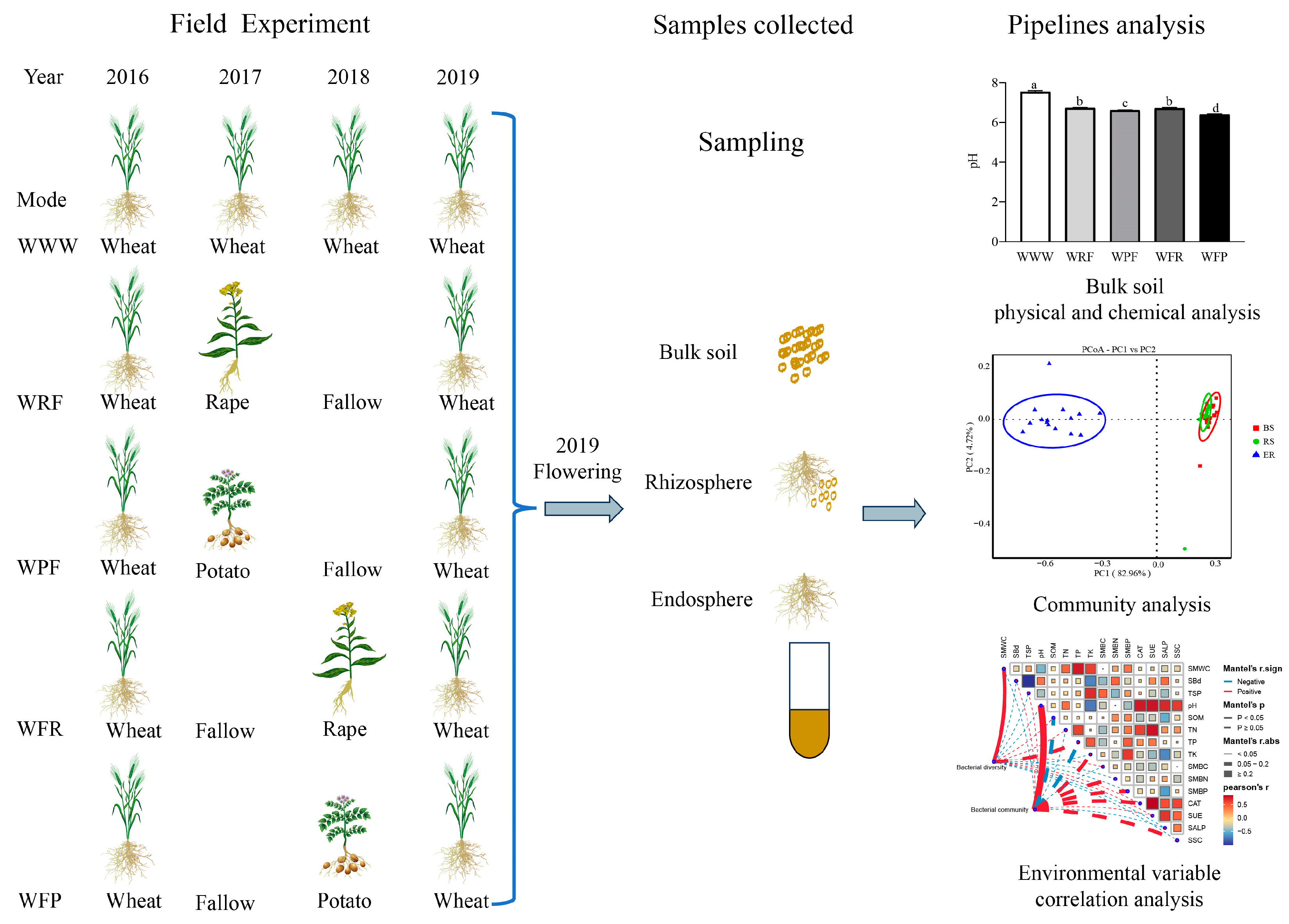
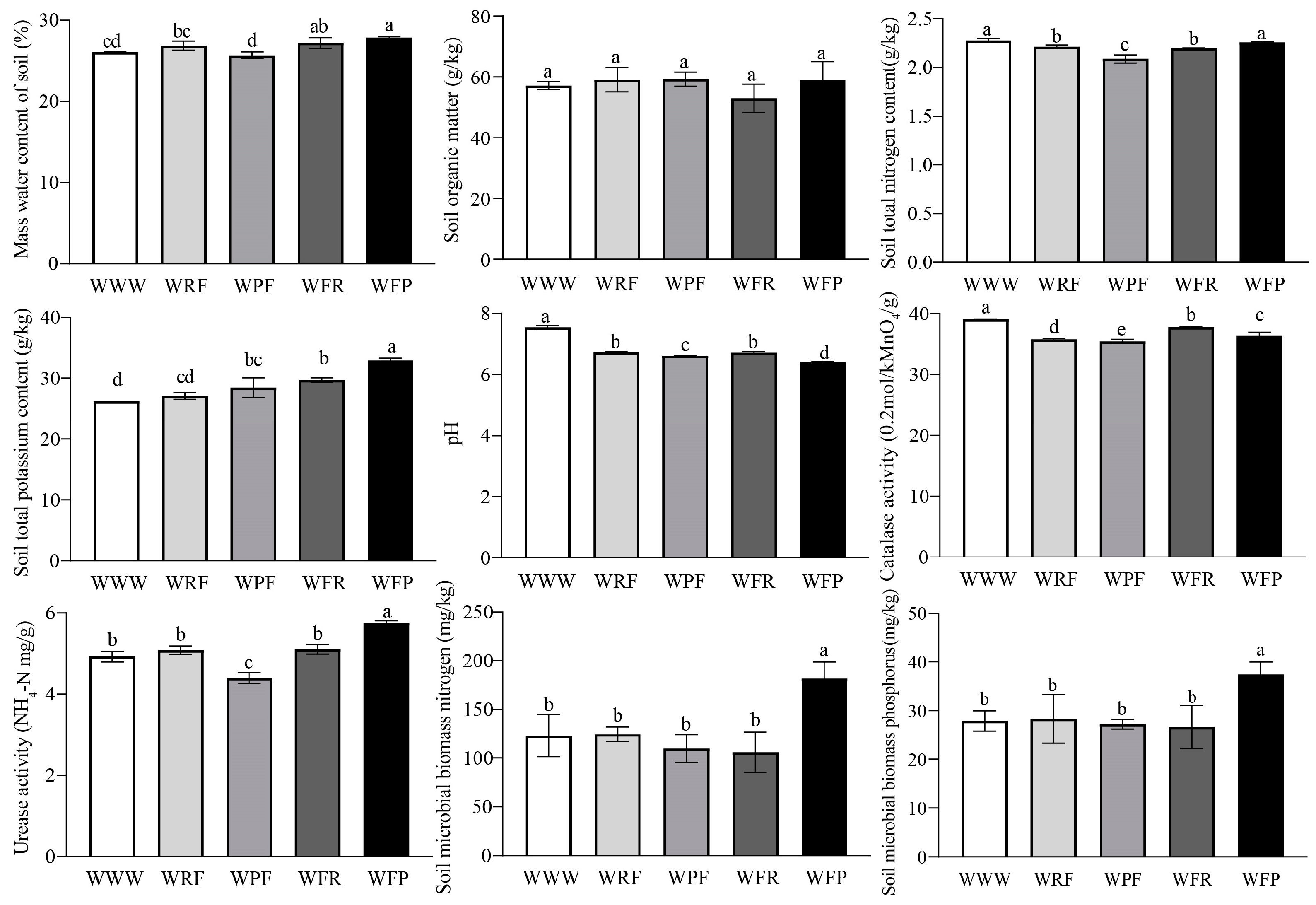
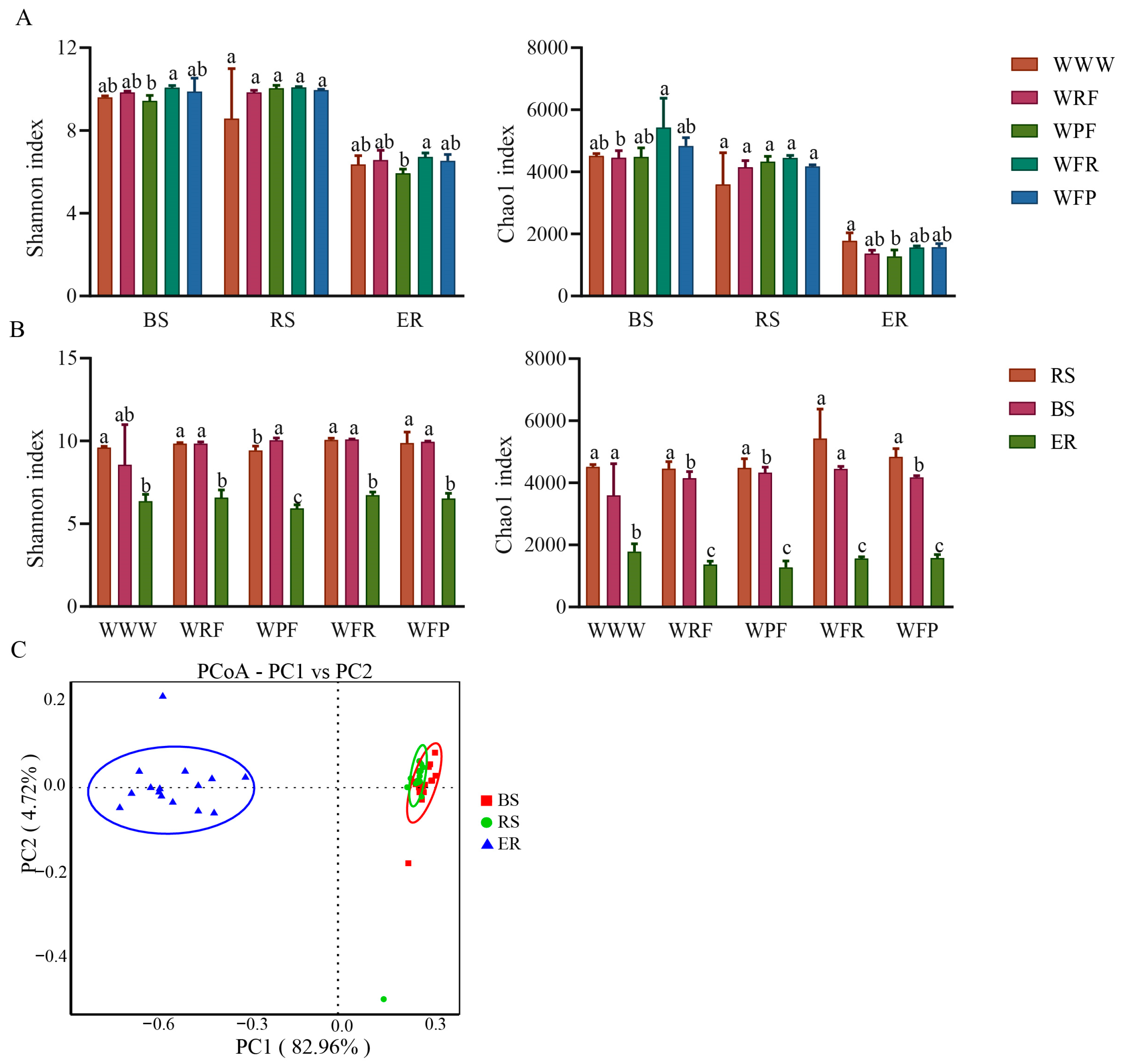
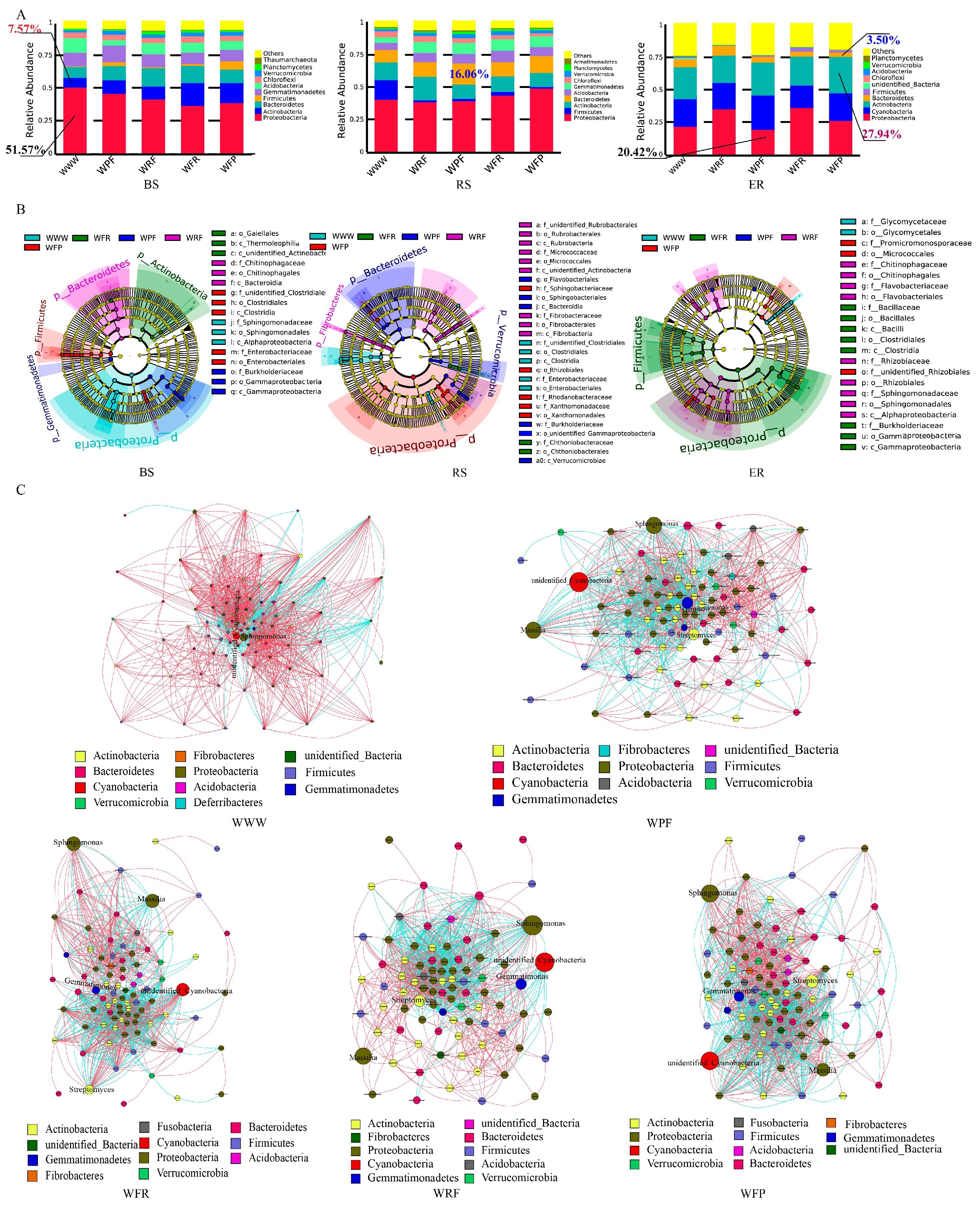
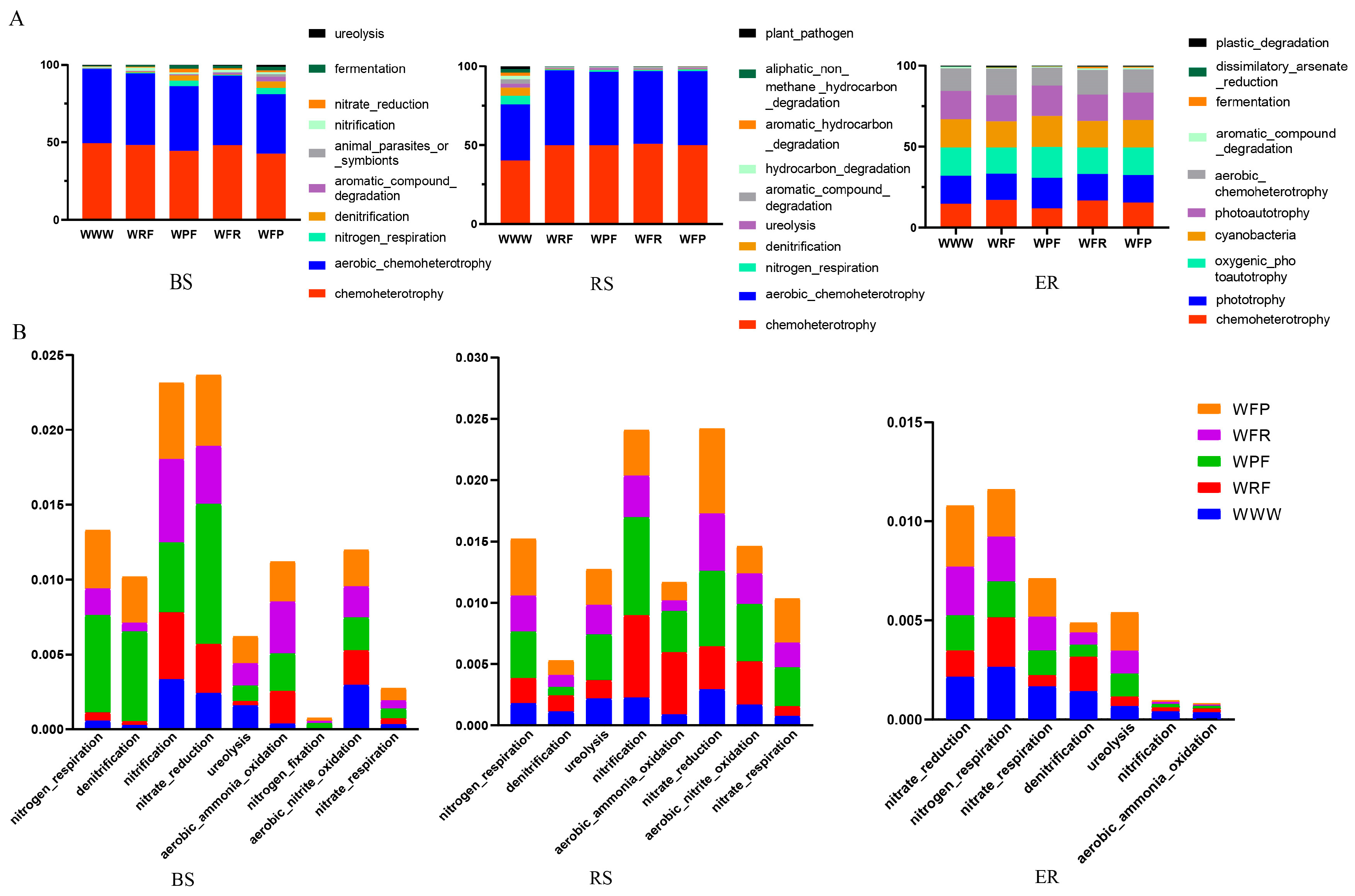
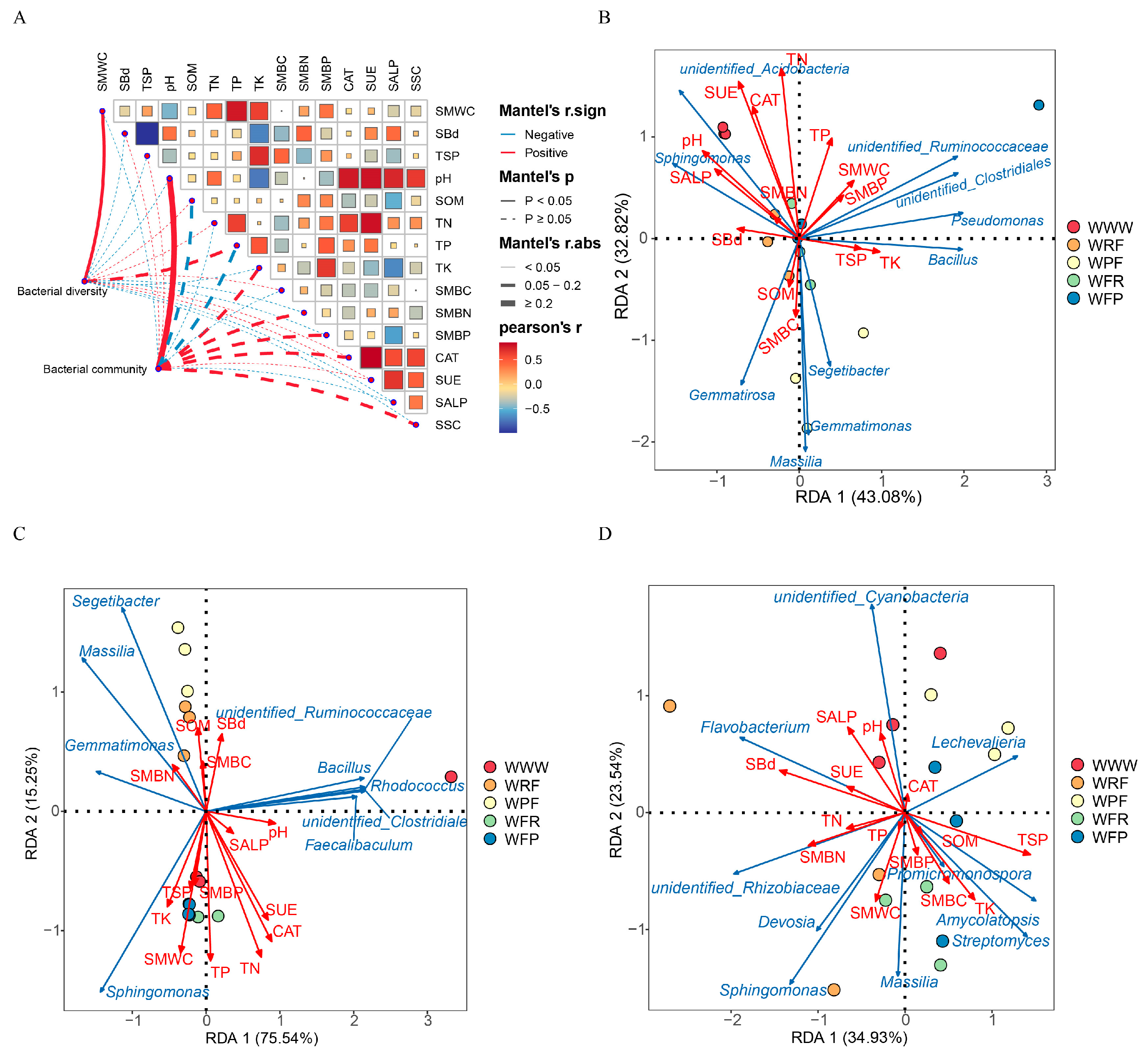
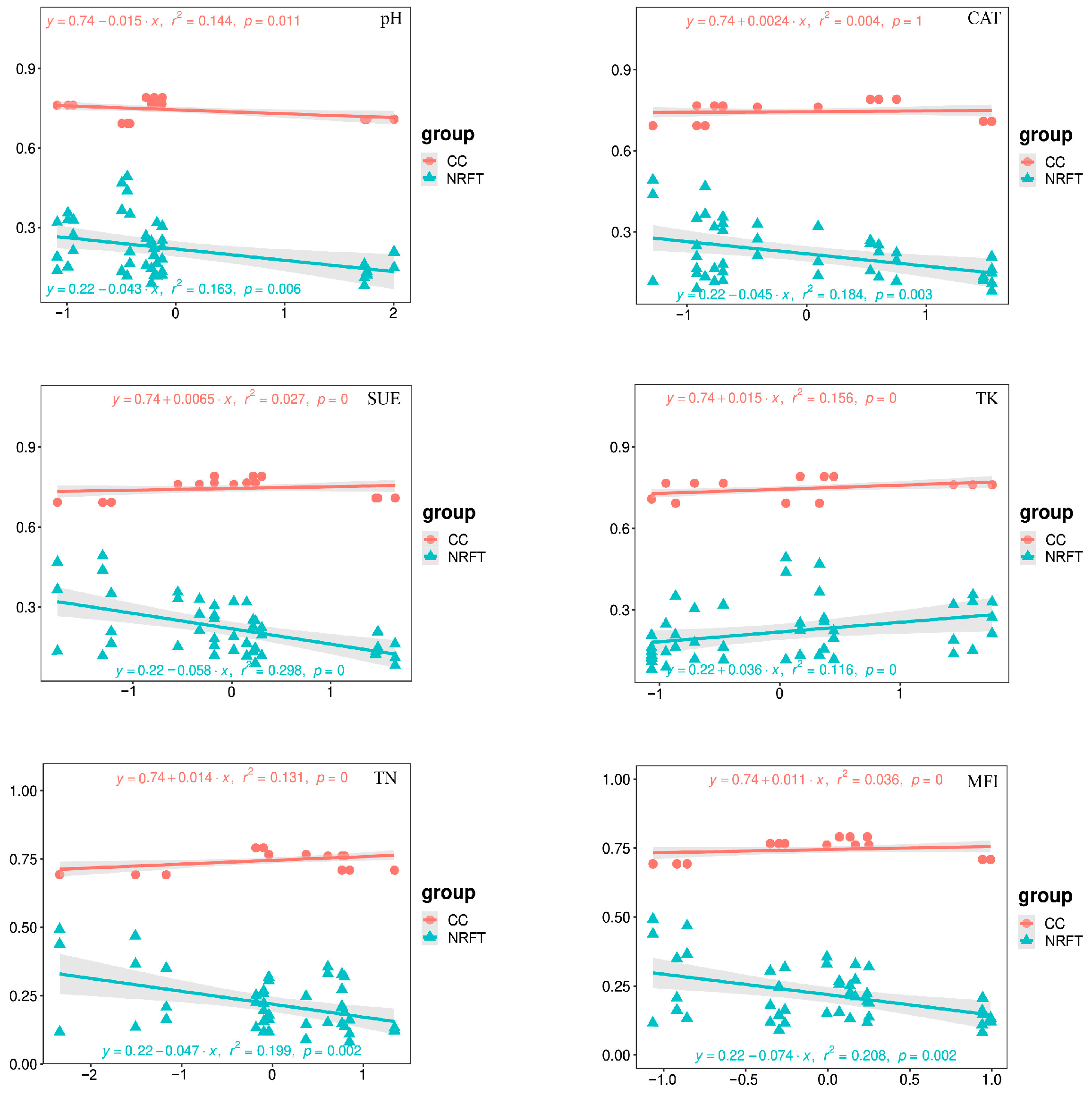
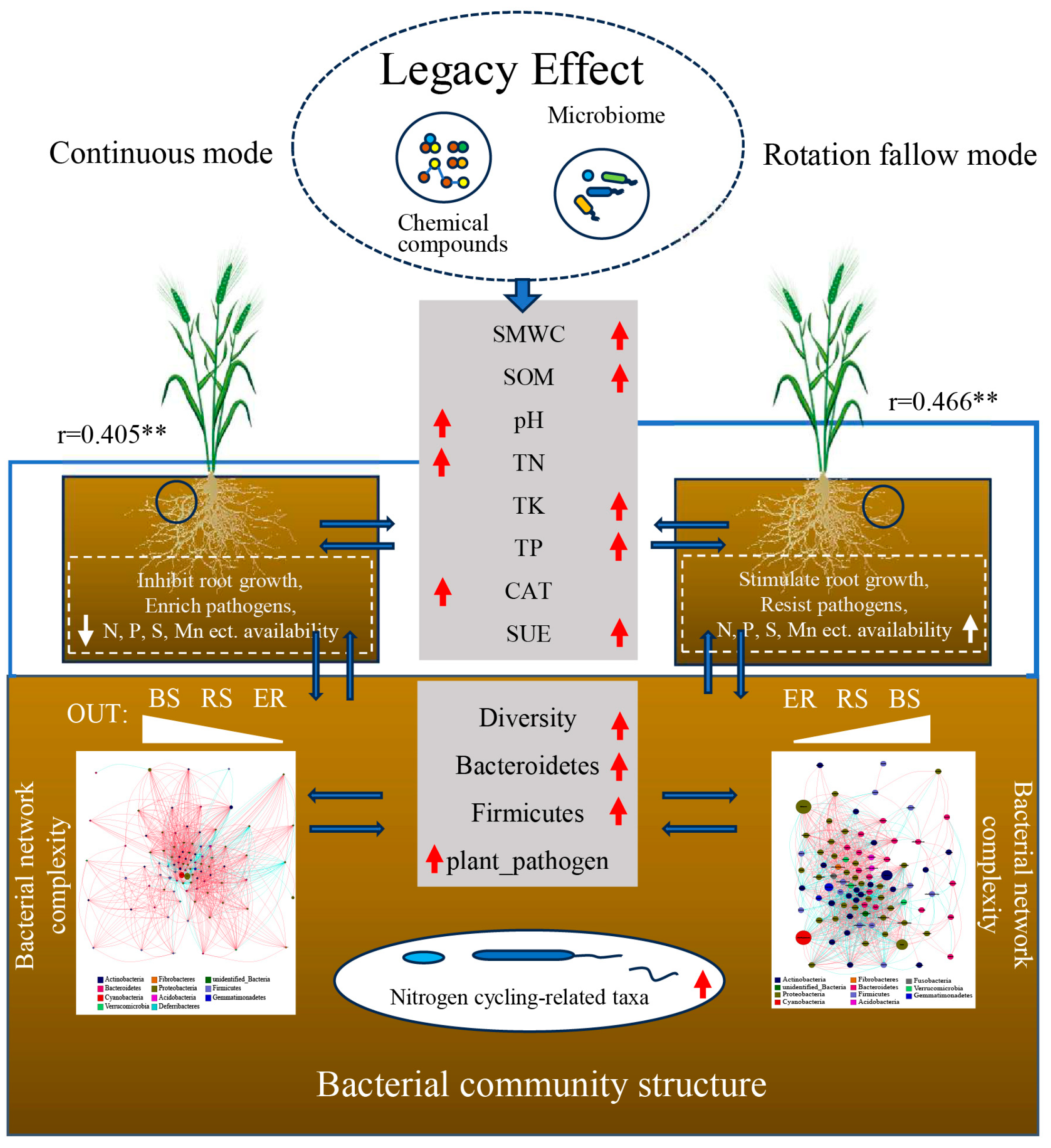
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).