
Transcriptome-Wide Genetic Variations in the Legume Genus Leucaena for Fingerprinting and Breeding

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA-seq and Variant Calling
2.3. Variant Filtration, Marker Development, and Phylogenetic Analysis
2.4. Genomic DNA Isolation
2.5. Polymerase Chain Reaction (PCR) and Scoring
2.6. Field Trials and F1 Phenotyping
3. Results and Discussion
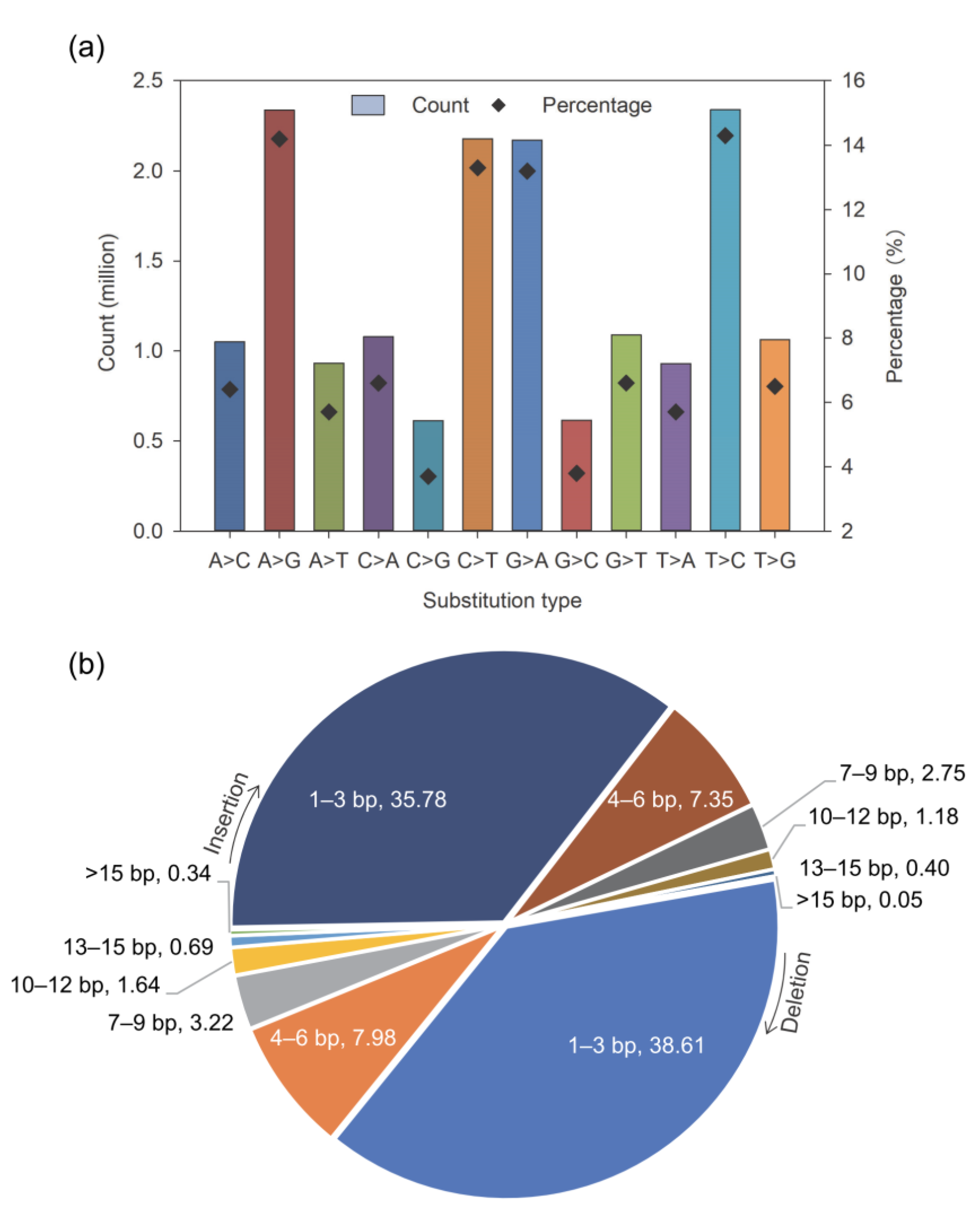
3.1. Transcriptome-Wide Genetic Variants in Leucaena Detected by RNA Sequencing
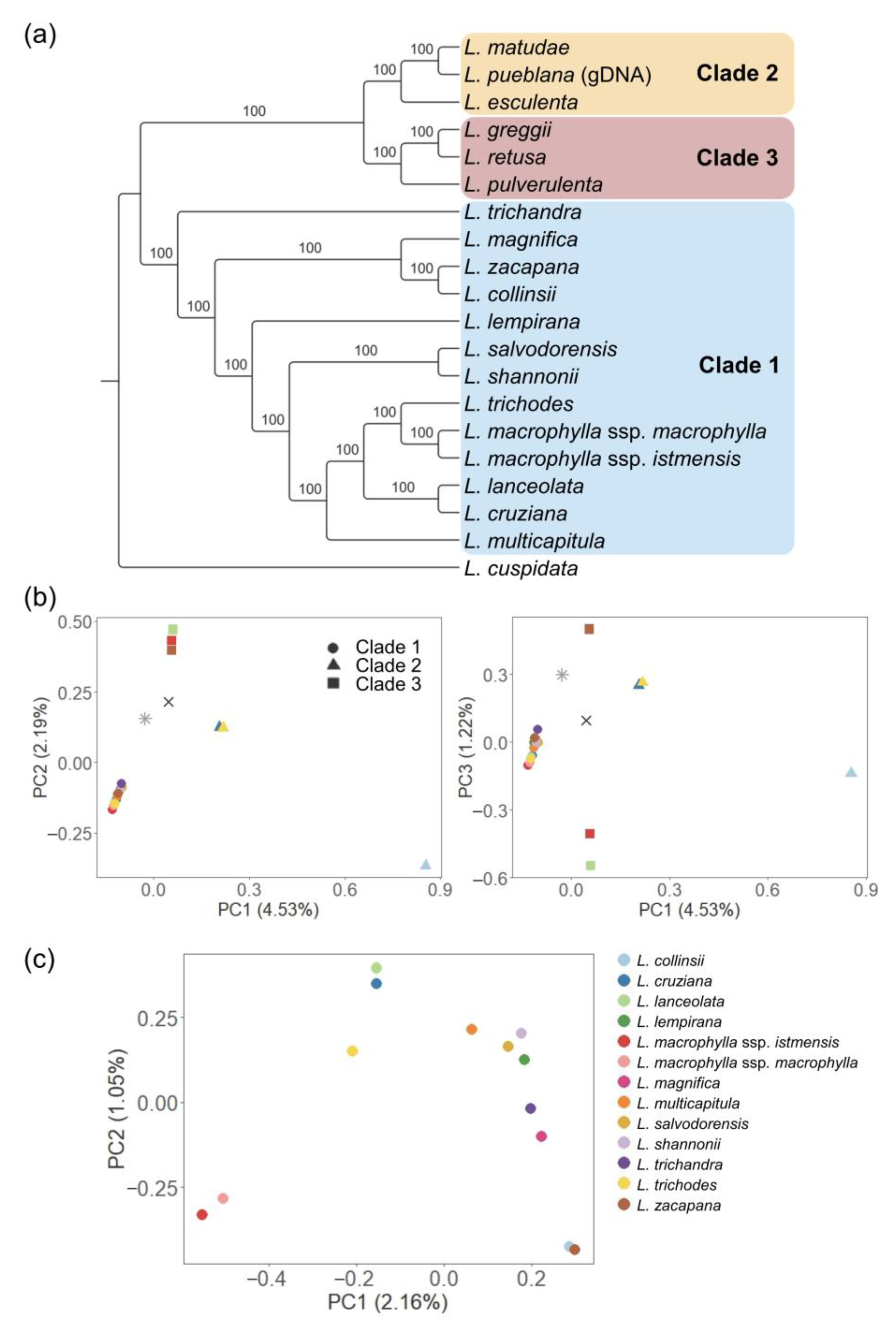
3.2. The Genus-Wide Sequence Variations Reflect the Phylogenetic History of Leucaena and Serve as a Genotyping Reference for Research and Breeding
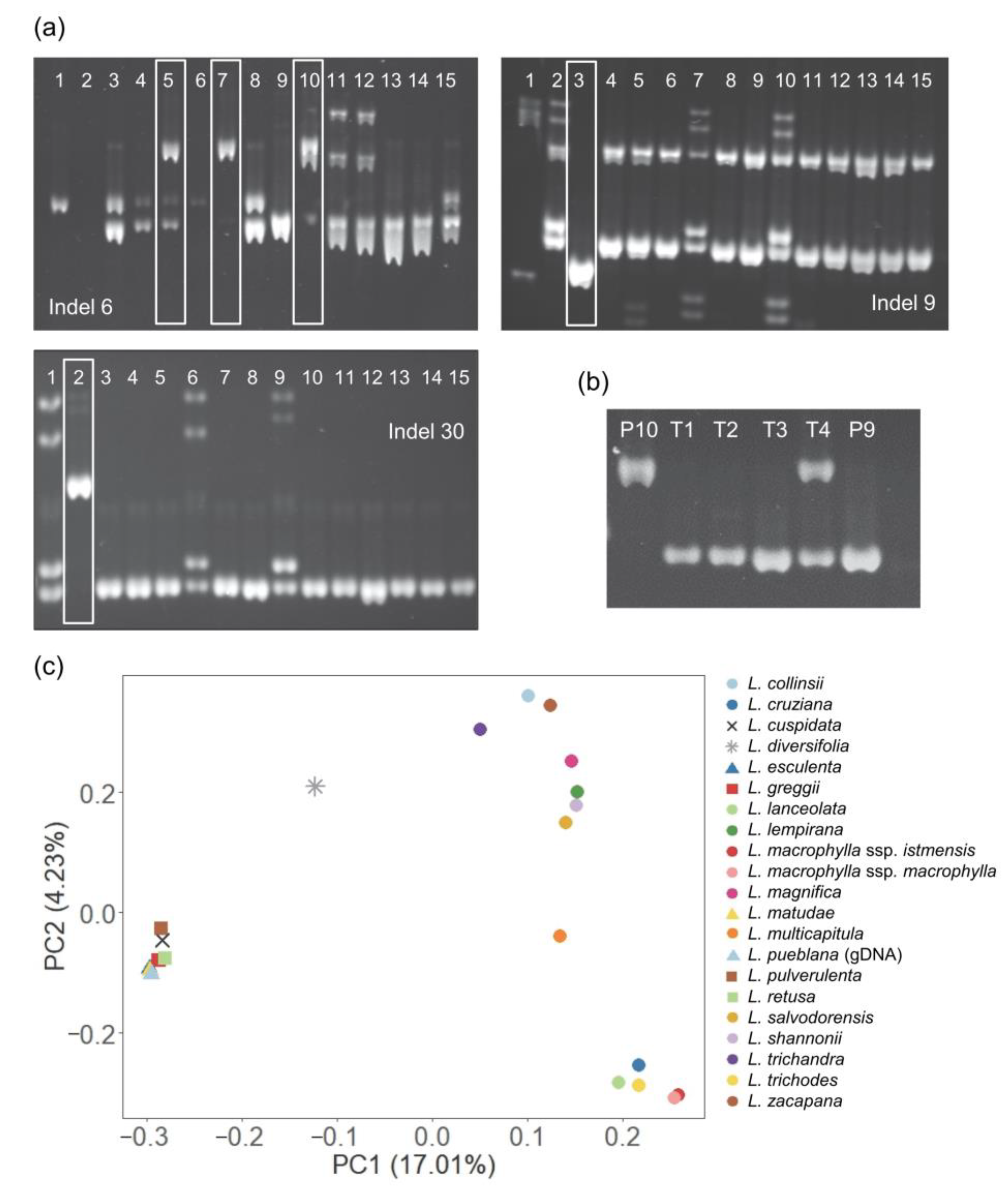
3.3. SNP Markers Outperformed Indels for Leucaena Genotyping
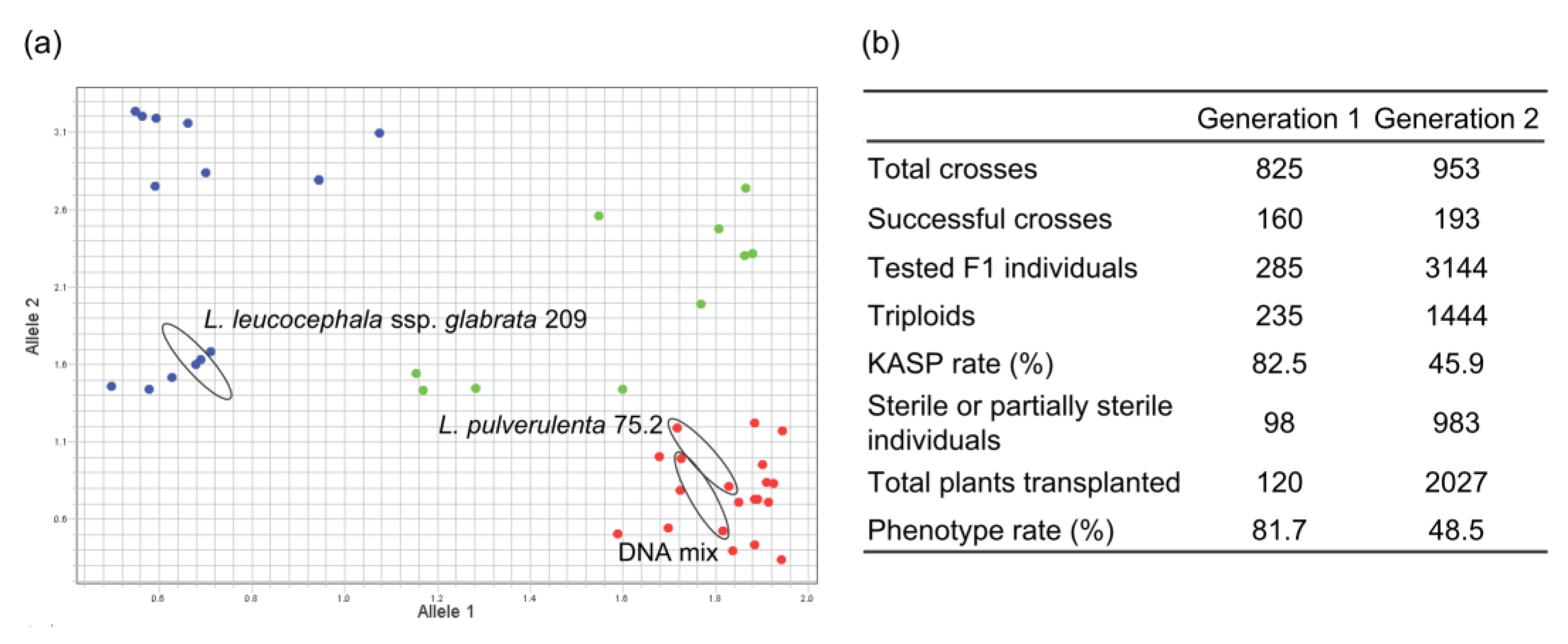
3.4. Reliability of Marker Fingerprinting Validated by Field Observations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, C.E. Leucaena: A Genetic Resources Handbook; Tropical Forestry Paper No 37; Oxford Forestry Institute: Oxford, UK, 1998. [Google Scholar]
- Sithole, N.; Tsvuura, Z.; Kirkman, K.; Magadlela, A. Nitrogen source preference and growth carbon costs of Leucaena leucocephala (Lam.) de Wit saplings in South African grassland soils. Plants 2021, 10, 2242. [Google Scholar] [CrossRef]
- Khanna, N.K.; Shukla, O.P.; Gogate, M.G.; Narkhede, S.L. Leucaena for paper industry in Gujarat, India: Case study. Trop. Grassl.-Forrajes. Trop. 2019, 7, 200–209. [Google Scholar] [CrossRef]
- Alemán-Ramirez, J.; Okoye, P.U.; Torres-Arellano, S.; Mejía-Lopez, M.; Sebastian, P. A review on bioenergetic applications of Leucaena leucocephala. Ind. Crop. Prod. 2022, 182, e114847. [Google Scholar] [CrossRef]
- Jube, S.; Borthakur, D. Development of an Agrobacterium-mediated transformation protocol for the tree-legume Leucaena leucocephala using immature zygotic embryos. Plant Cell Tiss. Org. 2009, 96, 325–333. [Google Scholar] [CrossRef]
- Buck, S.; Rolfe, J.; Lemin, C.; English, B. Adoption, profitability and future of Leucaena feeding systems in Australia. Trop. Grassl.-Forrajes. Trop. 2019, 7, 303–314. [Google Scholar] [CrossRef]
- Campbell, S.; Vogler, W.; Brazier, D.; Vitelli, J.; Brooks, S. Weed Leucaena and its significance, implications and control. Trop. Grassl.-Forrajes. Trop. 2019, 7, 280–289. [Google Scholar] [CrossRef]
- Real, D.; Han, Y.; Bailey, C.D.; Vasan, S.; Li, C.; Castello, M.; Broughton, S.; Abair, A.; Crouch, S.; Revell, C. Strategies to breed sterile Leucaena for Western Australia. Trop. Grassl.-Forrajes. Trop. 2019, 7, 80–86. [Google Scholar] [CrossRef]
- Brewbaker, J.L. Breeding Leucaena: Tropical multipurpose leguminous tree. In Plant Breeding Reviews; Janick, J., Ed.; Wiley-Blackwell Inc.: Hoboken, NJ, USA, 2016; Volume 40, pp. 43–123. [Google Scholar] [CrossRef]
- Real, D.; Revell, C.; Han, Y.; Li, C.; Castello, M.; Bailey, C.D. Successful creation of seedless (sterile) Leucaena germplasm developed from interspecific hybridisation for use as forage. Crop Pasture Sci. 2022, 74, 783–796. [Google Scholar] [CrossRef]
- Lema, M. Marker-assisted selection in comparison to conventional plant breeding. Agric. Res. Technol. 2018, 14, e555914. [Google Scholar] [CrossRef]
- Hughes, C.E.; Bailey, C.D.; Harris, S.A. Divergent and reticulate species relationships in Leucaena (Fabaceae) inferred from multiple data sources: Insights into polyploid origins and nrDNA polymorphism. Am. J. Bot. 2002, 89, 1057–1073. [Google Scholar] [CrossRef]
- Govindarajulu, R.; Hughes, C.E.; Bailey, C.D. Phylogenetic and population genetic analyses of diploid Leucaena (Leguminosae; Mimosoideae) reveal cryptic species diversity and patterns of divergent allopatric speciation. Am. J. Bot. 2011, 98, 2049–2063. [Google Scholar] [CrossRef]
- Rajarajan, K.; Uthappa, A.R.; Handa, A.K.; Chavan, S.B.; Vishnu, R.; Shrivastava, A.; Handa, A.; Rana, M.; Sahu, S.; Humar, N.; et al. Genetic diversity and population structure of Leucaena leucocephala (Lam.) de Wit genotypes using molecular and morphological attributes. Genet. Resour. Crop. Evol. 2022, 69, 71–83. [Google Scholar] [CrossRef]
- Walton, C.S. Leucaena (Leucaena leucocephala) in Queensland; Queensland Department of Natural Resources and Mines: Brisbane, Australia, 2003. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0009/57294/IPA-Leucaena-PSA.pdf (accessed on 13 November 2023).
- Russell, J.R.; Hedley, P.E.; Cardle, L.; Dancey, S.; Morris, J.; Booth, A.; Odee, D.; Mwaura, L.; Omondi, W.; Angaine, P.; et al. tropiTree: An NGS-based EST-SSR resource for 24 tropical tree species. PLoS ONE 2014, 9, e102502. [Google Scholar] [CrossRef]
- Tan, J.; Guo, J.J.; Yin, M.Y.; Wang, H.; Dong, W.P.; Zeng, J.; Zhou, S.L. Next generation sequencing-based molecular marker development: A case study in Betula alnoides. Molecules 2018, 23, 2963. [Google Scholar] [CrossRef]
- Mammadov, J.; Aggarwal, R.; Buyyarapu, R.; Kumpatla, S. SNP markers and their impact on plant breeding. Int. J. Plant. Genomics 2012, 2012, e728398. [Google Scholar] [CrossRef]
- Nagy, I.; Barth, S.; Mehenni-Ciz, J.; Abberton, M.T.; Milbourne, D. A hybrid next generation transcript sequencing-based approach to identify allelic and homeolog-specific single nucleotide polymorphisms in allotetraploid white clover. BMC Genomics 2013, 14, e100. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. 2016. Available online: https://ggplot2.tidyverse.org/ (accessed on 11 March 2024).
- Ortiz, E.M. vcf2phylip v2.0: Convert a VCF Matrix into Several Matrix Formats for Phylogenetic Analysis. Available online: https://zenodo.org/records/2540861 (accessed on 9 September 2019).
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szcześniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef]
- Pearce, S.; Vazquez-Gross, H.; Herin, S.Y.; Hane, D.; Wang, Y.; Gu, Y.Q.; Dubcovsky, J. WheatExp: An RNA-seq expression database for polyploid wheat. BMC Plant Biol. 2015, 15, 299. [Google Scholar] [CrossRef]
- Wang, S.; Li, Z.; Guo, X.; Fang, Y.; Xiang, J.; Jin, W. Comparative analysis of microsatellite, SNP, and InDel markers in four Rhododendron species based on RNA-seq. Breeding Sci. 2018, 68, 536–544. [Google Scholar] [CrossRef]
- Bachlava, E.; Taylor, C.A.; Tang, S.; Bowers, J.E.; Mandel, J.R.; Burke, J.M.; Knapp, S.J. SNP discovery and development of a high-density genotyping array for sunflower. PLoS ONE 2012, 7, e29814. [Google Scholar] [CrossRef]
- Lv, Y.; Liu, Y.; Zhao, H. mInDel: A high-throughput and efficient pipeline for genome-wide InDel marker development. BMC Genomics 2016, 17, e290. [Google Scholar] [CrossRef]
- Sehn, J.K. Insertions and deletions (Indels). In Clinical Genomics; Kulkarni, S., Pfiefer, J., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 129–150. [Google Scholar] [CrossRef]
- Karunarathne, S.; Walker, E.; Sharma, D.; Li, C.; Han, Y. Genetic resources and precise gene editing for targeted improvement of barley abiotic stress tolerance. J. Zhejiang Univ. Sci. B 2023, 24, 1069–1092. [Google Scholar] [CrossRef]
- Govindarajulu, R.; Hughes, C.E.; Alexander, P.J.; Bailey, C.D. The complex evolutionary dynamics of ancient and recent polyploidy in Leucaena (Leguminosae; Mimosoideae). Am. J. Bot. 2011, 98, 2064–2076. [Google Scholar] [CrossRef]
- Abair, A.; Hughes, C.E.; Bailey, C.D. The evolutionary history of Leucaena: Recent research, new genomic resources and future directions. Trop. Grassl.-Forrajes. Trop. 2019, 7, 65–73. [Google Scholar] [CrossRef]
- von Maydell, D.; Beleites, C.; Stache, A.; Riewe, D.; Krähmer, A.; Marthe, F. Genetic variation of annual and biennial caraway (Carum carvi) germplasm offers diverse opportunities for breeding. Ind. Crop. Prod. 2024, 208, e117798. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; De Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Primers 2021, 1, 59. [Google Scholar] [CrossRef]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Nagy, L.G.; Kocsubé, S.; Csanádi, Z.; Kovács, G.M.; Petkovits, T.; Vágvölgyi, C.; Papp, T. Re-mind the gap! Insertion—Deletion data reveal neglected phylogenetic potential of the nuclear ribosomal internal transcribed spacer (ITS) of fungi. PLoS ONE 2012, 7, e49794. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Cohen, O.; Pupko, T.; Huchon, D. Indel reliability in indel-based phylogenetic inference. Genome Biol. Evol. 2014, 6, 3199–3209. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analysing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Liu, S.; An, Y.; Tong, W.; Qin, X.; Samarina, L.; Guo, R.; Xia, X.; Wei, C. Characterization of genome-wide genetic variations between two varieties of tea plant (Camellia sinensis) and development of InDel markers for genetic research. BMC Genomics 2019, 20, 935. [Google Scholar] [CrossRef]
- Piskol, R.; Ramaswami, G.; Li, J.B. Reliable identification of genomic variants from RNA-seq data. Am. J. Hum. Genet. 2013, 93, 641–651. [Google Scholar] [CrossRef]
- Huang, Z.; Peng, G.; Gossen, B.D.; Yu, F. Fine mapping of a clubroot resistance gene from turnip using SNP markers identified from bulked segregant RNA-Seq. Mol. Breed. 2019, 39, 131. [Google Scholar] [CrossRef]
- Muñoz-Espinoza, C.; Di Genova, A.; Sánchez, A.; Correa, J.; Espinoza, A.; Meneses, C.; Maass, A.; Orellana, A.; Hinrichsen, P. Identification of SNPs and InDels associated with berry size in table grapes integrating genetic and transcriptomic approaches. BMC Plant Biol. 2020, 20, e365. [Google Scholar] [CrossRef]
- Chen, Z.; Tang, D.; Ni, J.; Li, P.; Wang, L.; Zhou, W.; Li, C.; Lan, H.; Li, L.; Liu, J. Development of genic KASP SNP markers from RNA-Seq data for map-based cloning and marker-assisted selection in maize. BMC Plant. Biol. 2021, 21, 157. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Chen, S.; Chen, L.; Sun, K.; Huang, C.; Zhou, D.; Huang, Y.; Wang, J.; Liu, Y.; Wang, H.; et al. Development of a core SNP arrays based on the KASP method for molecular breeding of rice. Rice 2019, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Mavi, G.S.; Gill, M.S.; Saini, D.K. Utilization of KASP technology for wheat improvement. Cereal Res. Commun. 2020, 48, 409–421. [Google Scholar] [CrossRef]
- Tao, D.; Kalendar, R.; Paterson, A.H. Editorial: Interspecific hybridisation in plant biology. Front. Plant Sci. 2022, 13, e1026492. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | nRefHom | nNonRefHom | nHets | nTransitions | nTransversions | nIndels | NCBI Accession Number |
|---|---|---|---|---|---|---|---|
| Leucaena collinsii Britton & Rose (2×) | 8428728 | 5,228,368 | 2,806,318 | 4,440,773 | 3,593,913 | 450,406 | SRX2719653 |
| Leucaena cruziana Britton & Rose (2×) | 8,284,936 | 5,330,027 | 2,839,134 | 4,515,216 | 3,653,945 | 459,723 | SRX2719651 |
| Leucaena cuspidata Standl. (2×) | 7,554,107 | 5,839,938 | 3,033,407 | 4,891,943 | 3,981,402 | 486,368 | SRX2719650 |
| Leucaena diversifolia (Schltdl.) Benth. (4×) | 7,686,304 | 4,977,243 | 3,791,795 | 4,830,917 | 3,938,121 | 458,478 | SRX2719649 |
| Leucaena esculenta (DC.) Benth. (2×) | 7,024,119 | 6,160,890 | 3,218,074 | 5,159,900 | 4,219,064 | 510,737 | SRX2719648 |
| Leucaena greggii S. Watson (2×) | 7,282,246 | 6,090,881 | 3,048,606 | 5,037,017 | 4,102,470 | 492,087 | SRX2719647 |
| Leucaena lanceolata S. Watson (2×) | 8,312,577 | 5,360,102 | 2,782,799 | 4,505,372 | 3,637,529 | 458,342 | SRX2719645 |
| Leucaena lempirana C.E. Hughes (2×) | 8,429,399 | 5,192,417 | 2,842,191 | 4,444,316 | 3,590,292 | 449,813 | SRX2719644 |
| Leucaena macrophylla subsp. istmensis C.E. Hughes (2×) | 8,244,333 | 5,462,404 | 2,743,421 | 4,536,361 | 3,669,464 | 463,662 | SRX2719641 |
| Leucaena macrophylla subsp. macrophylla Benth. (2×) | 8,208,257 | 5,382,254 | 2,857,991 | 4,554,355 | 3,685,890 | 465,318 | SRX2719640 |
| Leucaena magnifica (C.E. Hughes) C.E. Hughes (2×) | 8,465,544 | 5,150,898 | 2,848,108 | 4,415,835 | 3,583,171 | 449,270 | SRX2719639 |
| Leucaena matudae (Zarate) C.E. Hughes (2×) | 6,982,398 | 6,187,703 | 3,231,807 | 5,181,555 | 4,237,955 | 511,912 | SRX2719638 |
| Leucaena multicapitula Schery (2×) | 8,372,304 | 5,299,403 | 2,788,487 | 4,474,205 | 3,613,685 | 453,626 | SRX2719637 |
| Leucaena pueblana Britton & Rose (2×) | 5,793,382 | 8,948,739 | 1,578,880 | 5,790,542 | 4,737,077 | 592,819 | SRX2719610 |
| Leucaena pulverulenta (Schltdl.) Benth. (2×) | 7,251,662 | 6,145,143 | 3,024,462 | 5,046,850 | 4,122,755 | 492,553 | SRX2719635 |
| Leucaena retusa Benth. (2×) | 7,248,430 | 6,151,778 | 3,021,196 | 5,057,547 | 4,115,427 | 492,416 | SRX2719634 |
| Leucaena salvadorensis Standl. ex Britton & Rose (2×) | 8,350,557 | 5,157,831 | 2,950,991 | 4,481,168 | 3,627,654 | 454,441 | SRX2719633 |
| Leucaena shannonii Donn.Sm. (2×) | 8,391,792 | 5,170,550 | 2,899,176 | 4,459,365 | 3,610,361 | 452,302 | SRX2719632 |
| Leucaena trichandra (Zucc.) Urb. (2×) | 8,773,909 | 4,785,556 | 2,925,521 | 4,257,895 | 3,453,182 | 428,834 | SRX2719631 |
| Leucaena trichodes (Jacq.) Benth. (2×) | 8,285,059 | 5,390,552 | 2,777,911 | 4,516,372 | 3,652,091 | 460,298 | SRX2719630 |
| Leucaena zacapana (C.E. Hughes) R. Govind. & C.E. Hughes (2×) | 8,419,838 | 5,192,514 | 2,851,655 | 4,448,285 | 3,595,884 | 449,813 | SRX2719629 |
| Total number of unique variants | SNPs 16,396,328 | 9,024,214 | 7,372,114 | 816,282 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Abair, A.; van der Zanden, J.; Nageswara-Rao, M.; Vasan, S.P.; Bhoite, R.; Castello, M.; Bailey, D.; Revell, C.; Li, C.; et al. Transcriptome-Wide Genetic Variations in the Legume Genus Leucaena for Fingerprinting and Breeding. Agronomy 2024, 14, 1519. https://doi.org/10.3390/agronomy14071519
Han Y, Abair A, van der Zanden J, Nageswara-Rao M, Vasan SP, Bhoite R, Castello M, Bailey D, Revell C, Li C, et al. Transcriptome-Wide Genetic Variations in the Legume Genus Leucaena for Fingerprinting and Breeding. Agronomy. 2024; 14(7):1519. https://doi.org/10.3390/agronomy14071519
Chicago/Turabian StyleHan, Yong, Alexander Abair, Julian van der Zanden, Madhugiri Nageswara-Rao, Saipriyaa Purushotham Vasan, Roopali Bhoite, Marieclaire Castello, Donovan Bailey, Clinton Revell, Chengdao Li, and et al. 2024. "Transcriptome-Wide Genetic Variations in the Legume Genus Leucaena for Fingerprinting and Breeding" Agronomy 14, no. 7: 1519. https://doi.org/10.3390/agronomy14071519
APA StyleHan, Y., Abair, A., van der Zanden, J., Nageswara-Rao, M., Vasan, S. P., Bhoite, R., Castello, M., Bailey, D., Revell, C., Li, C., & Real, D. (2024). Transcriptome-Wide Genetic Variations in the Legume Genus Leucaena for Fingerprinting and Breeding. Agronomy, 14(7), 1519. https://doi.org/10.3390/agronomy14071519