
Effects of FeSO4 and Organic Additives on Soil Properties and Microbiota during Model Soybean Planting in Saline-Alkali Soil

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Site
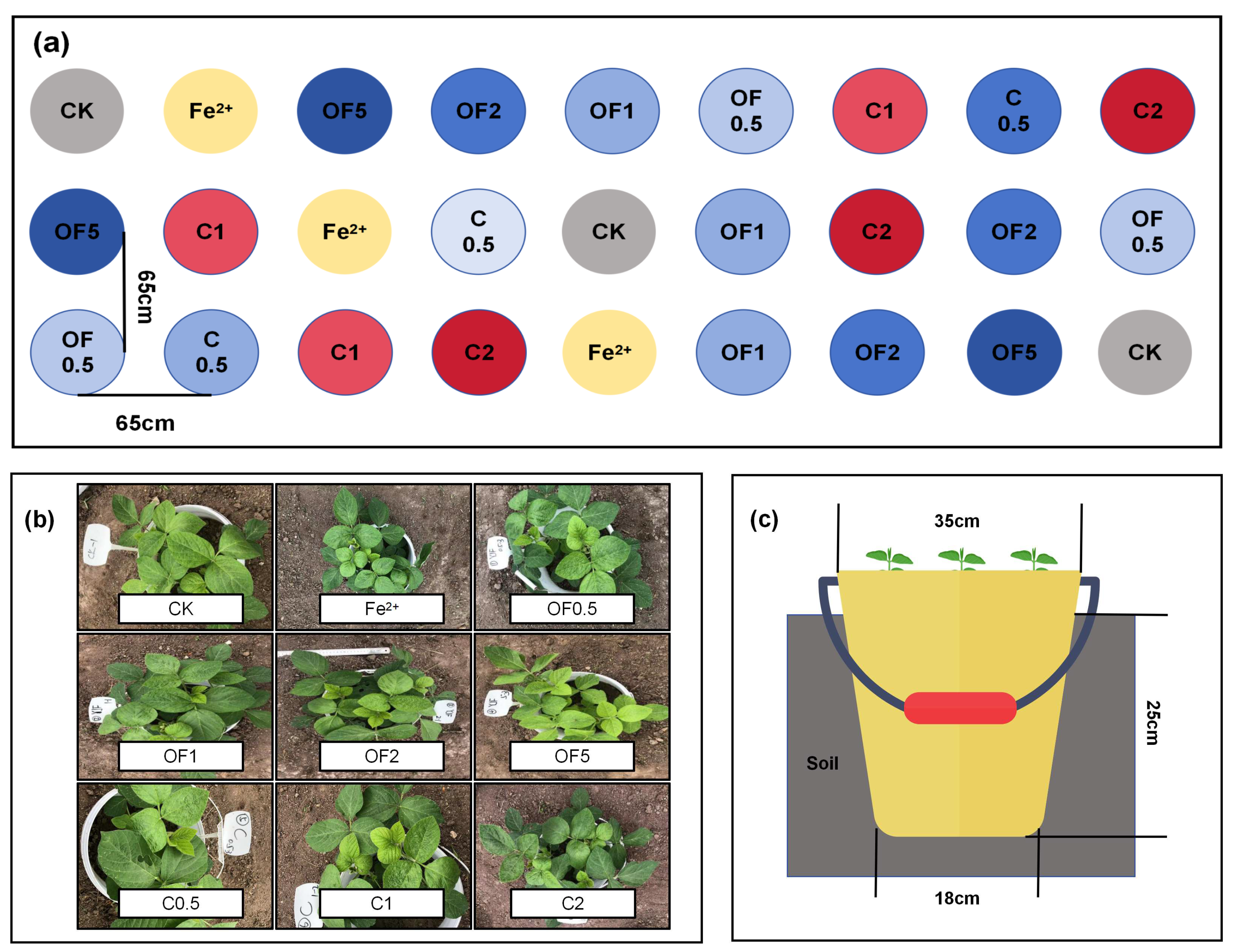
2.2. Experimental Design
2.3. Soil Sampling and Analysis
2.4. Soil Microbial Community Analysis
2.5. Statistical Analysis
3. Results
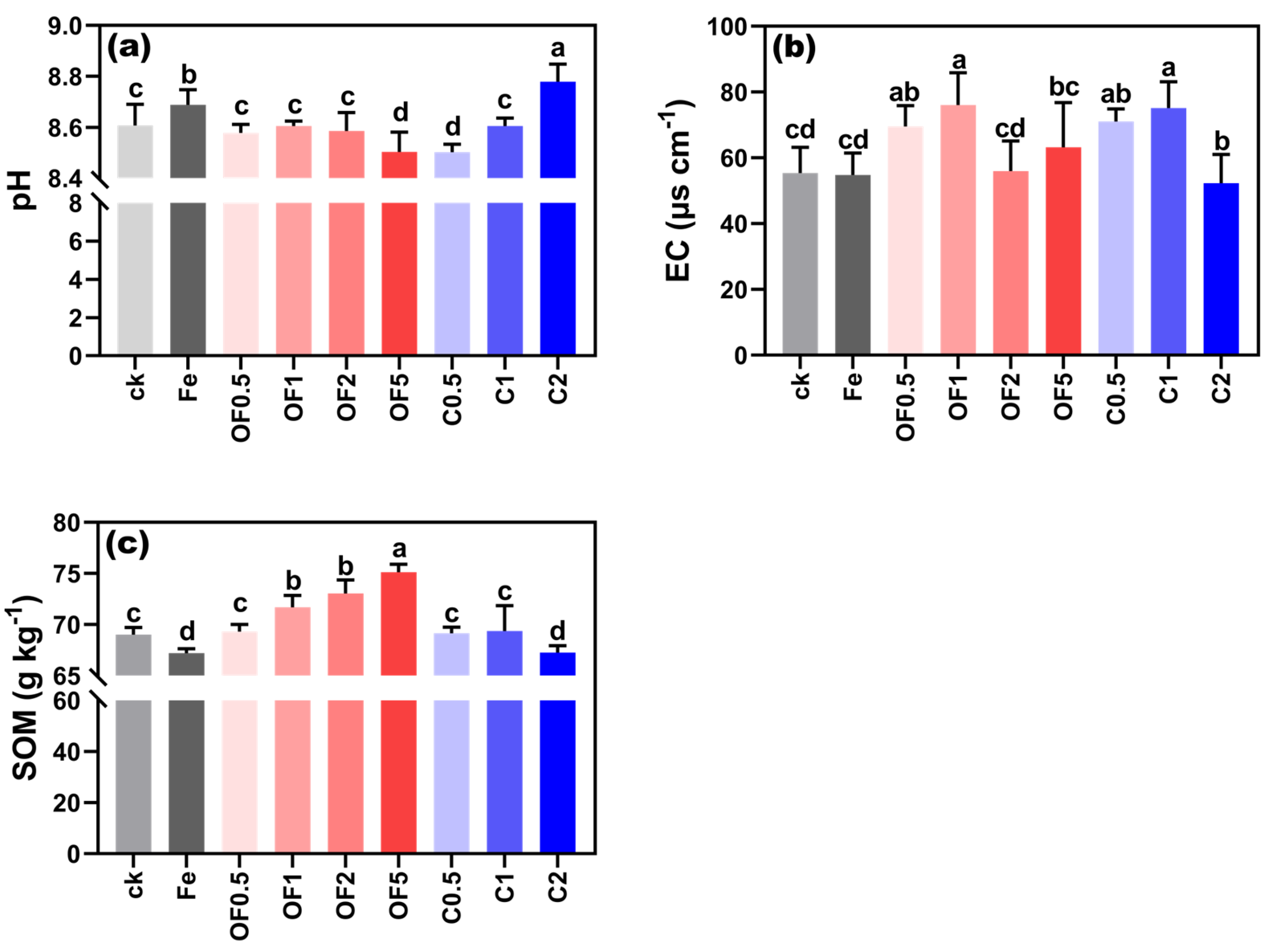
3.1. Soil pH, EC, and Organic Matter
3.2. Nutrients in Soil
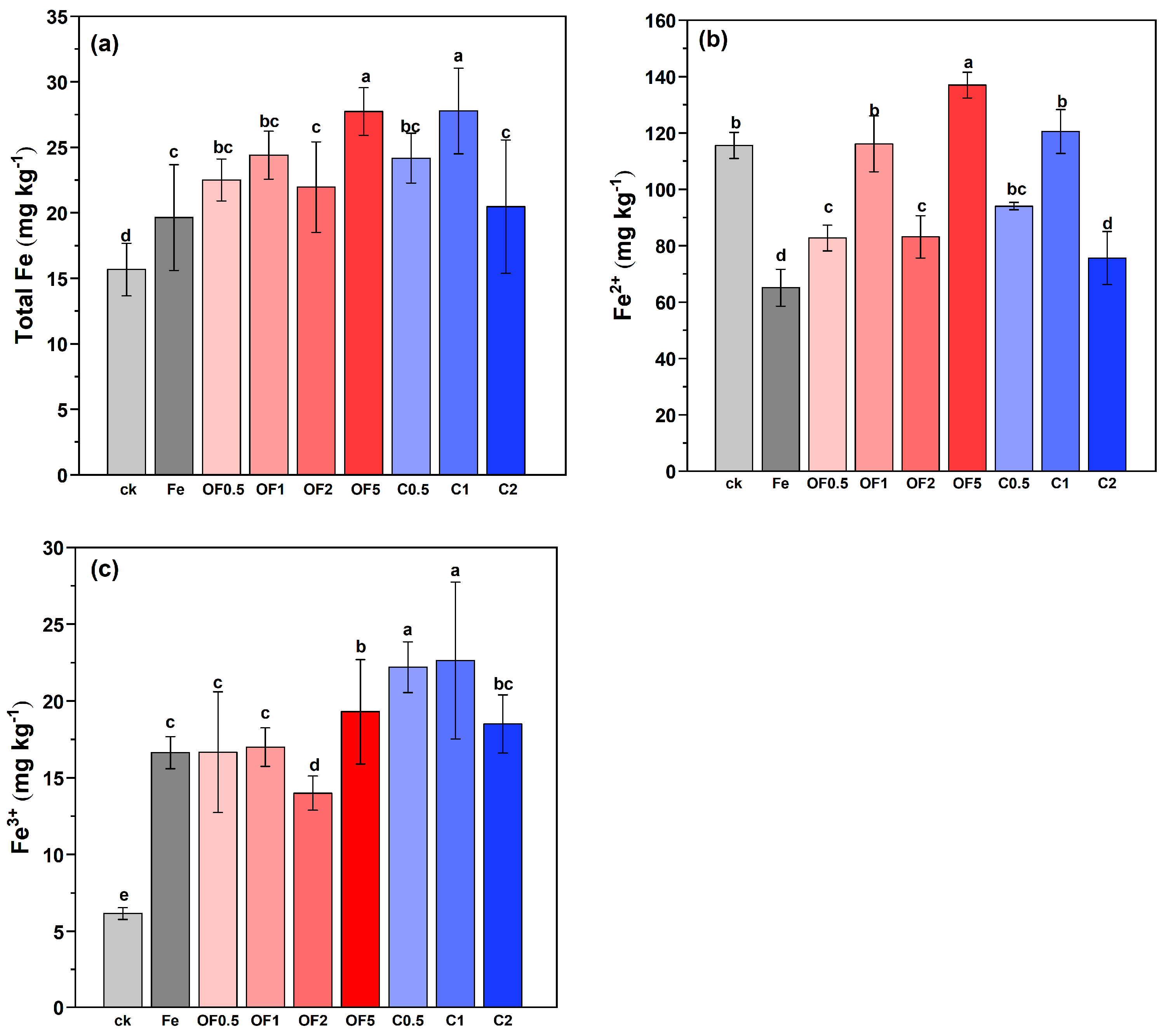
3.3. Changes in Iron Forms in Soil
3.4. Effect of Adding Organic Material on the Microbial Community of Saline-Alkali Soils
3.4.1. Alpha Diversity Analysis
3.4.2. PCA of Microbial Communities of Soil Subjected to Different Treatments
3.4.3. Changes in Microbial Community Diversity at the Phylum Level
3.4.4. Changes in Microbial Community Diversity at the Genus Level
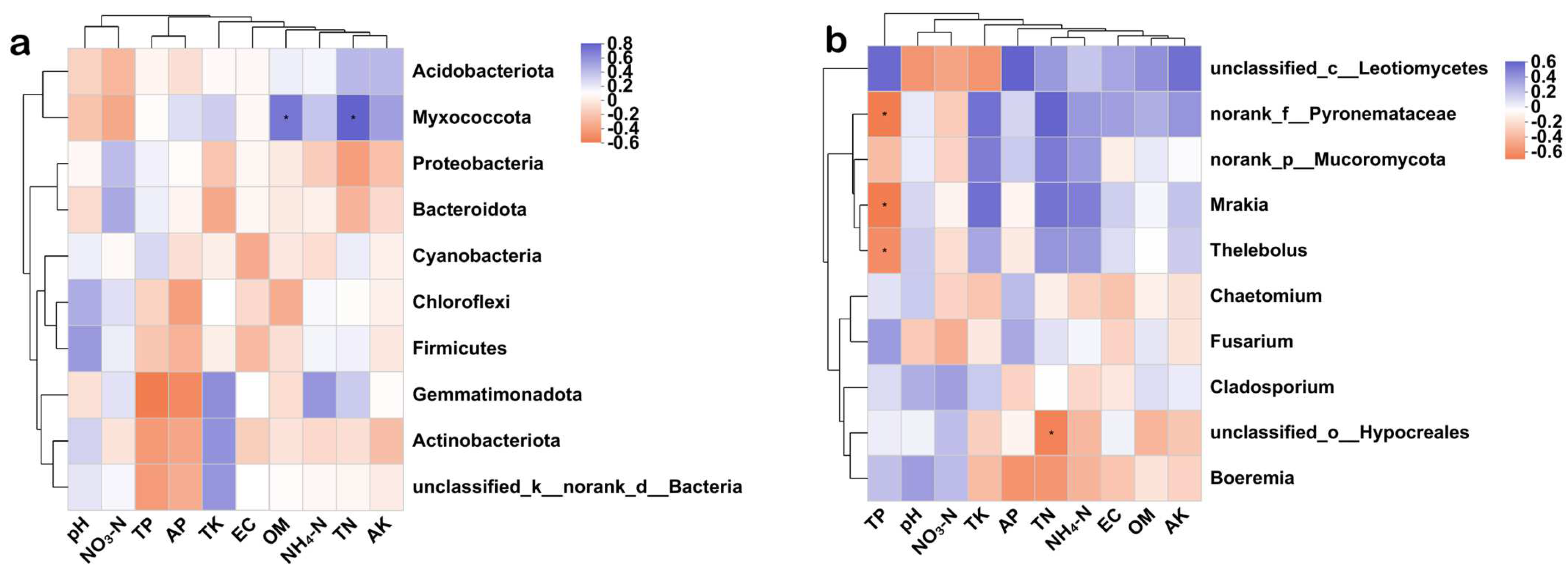
3.4.5. Correlation Analysis between Soil Microbial Community and Soil Environmental Factors
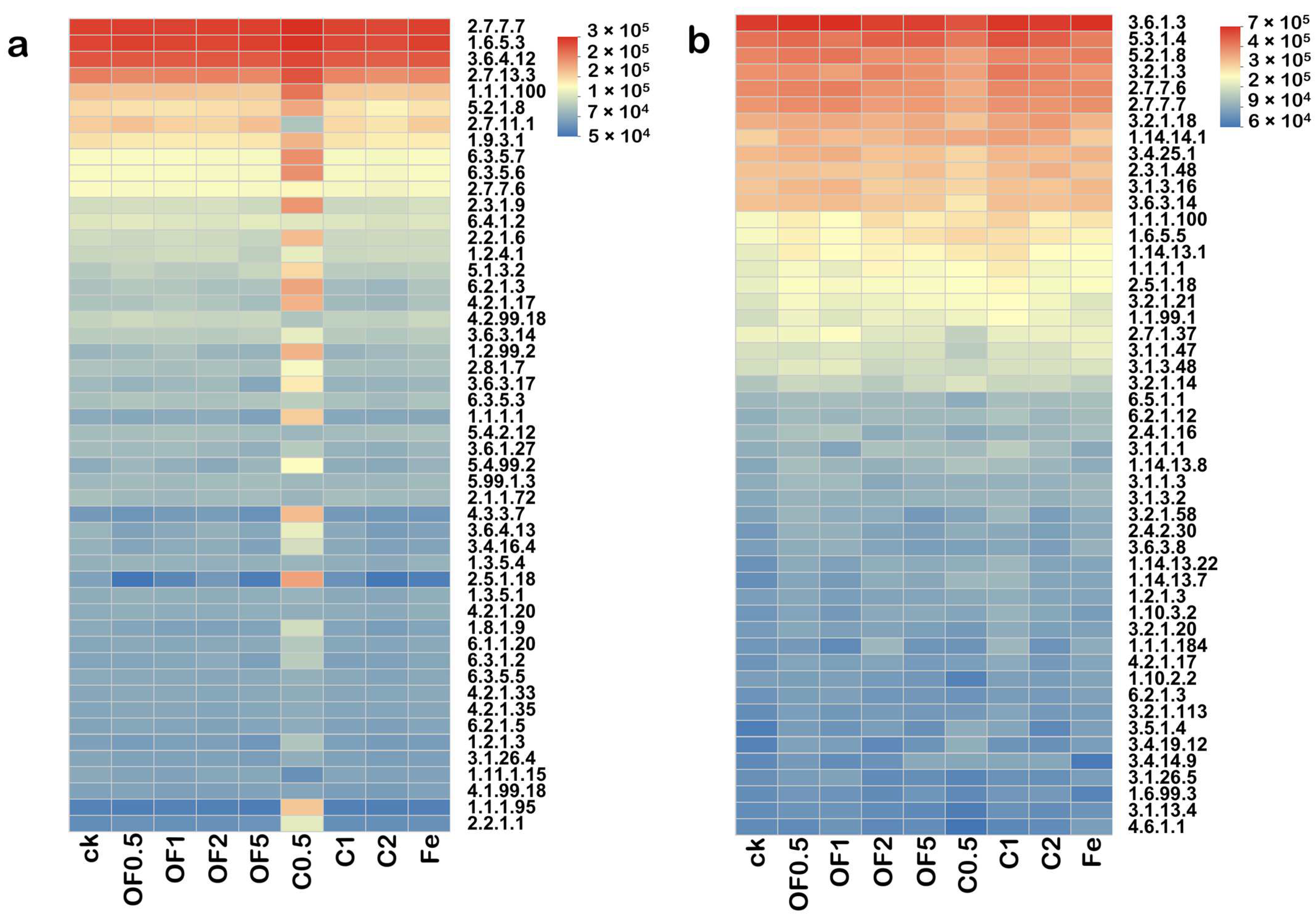
3.4.6. Functional Prediction Analysis of Soil Microbial Communities
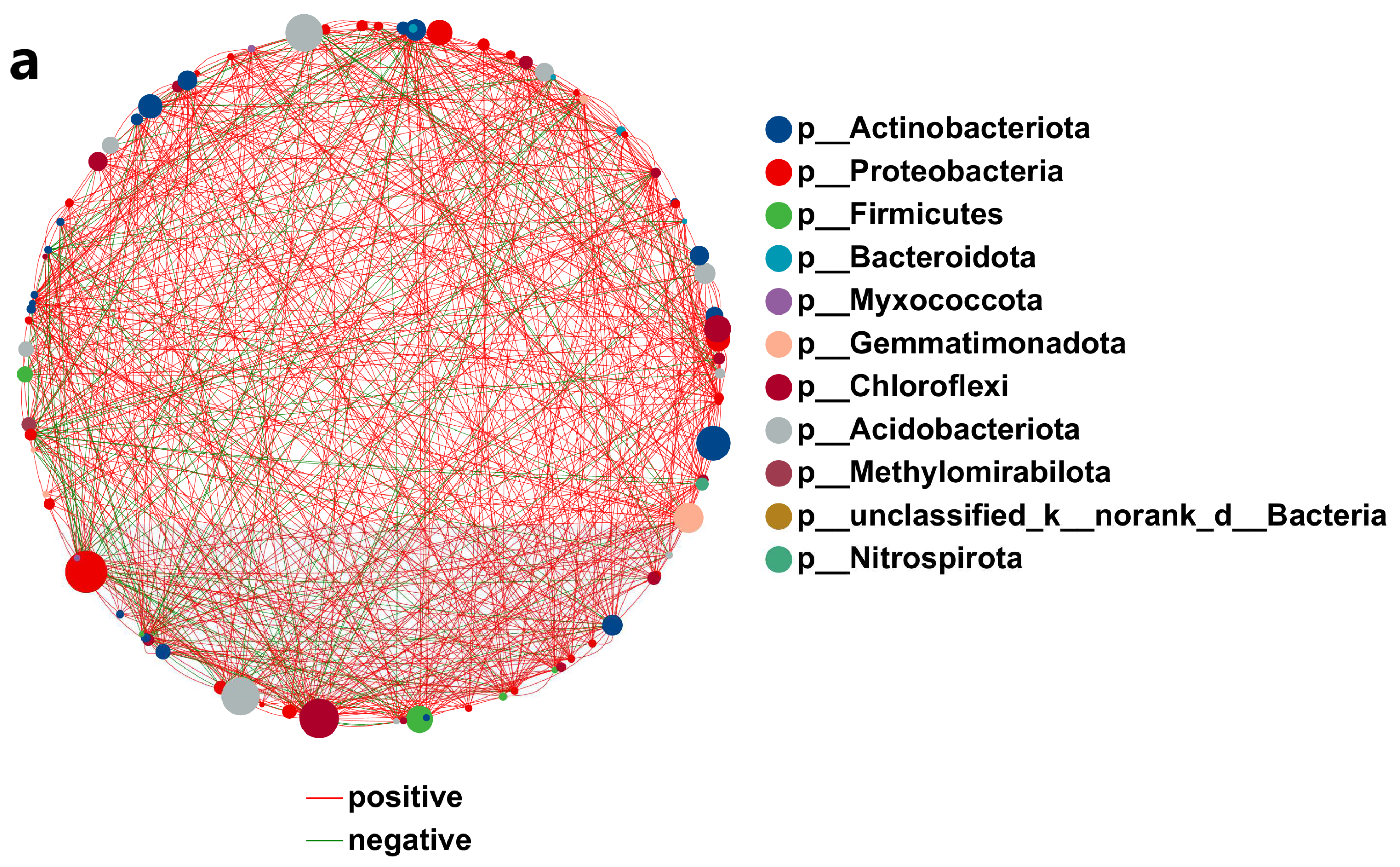
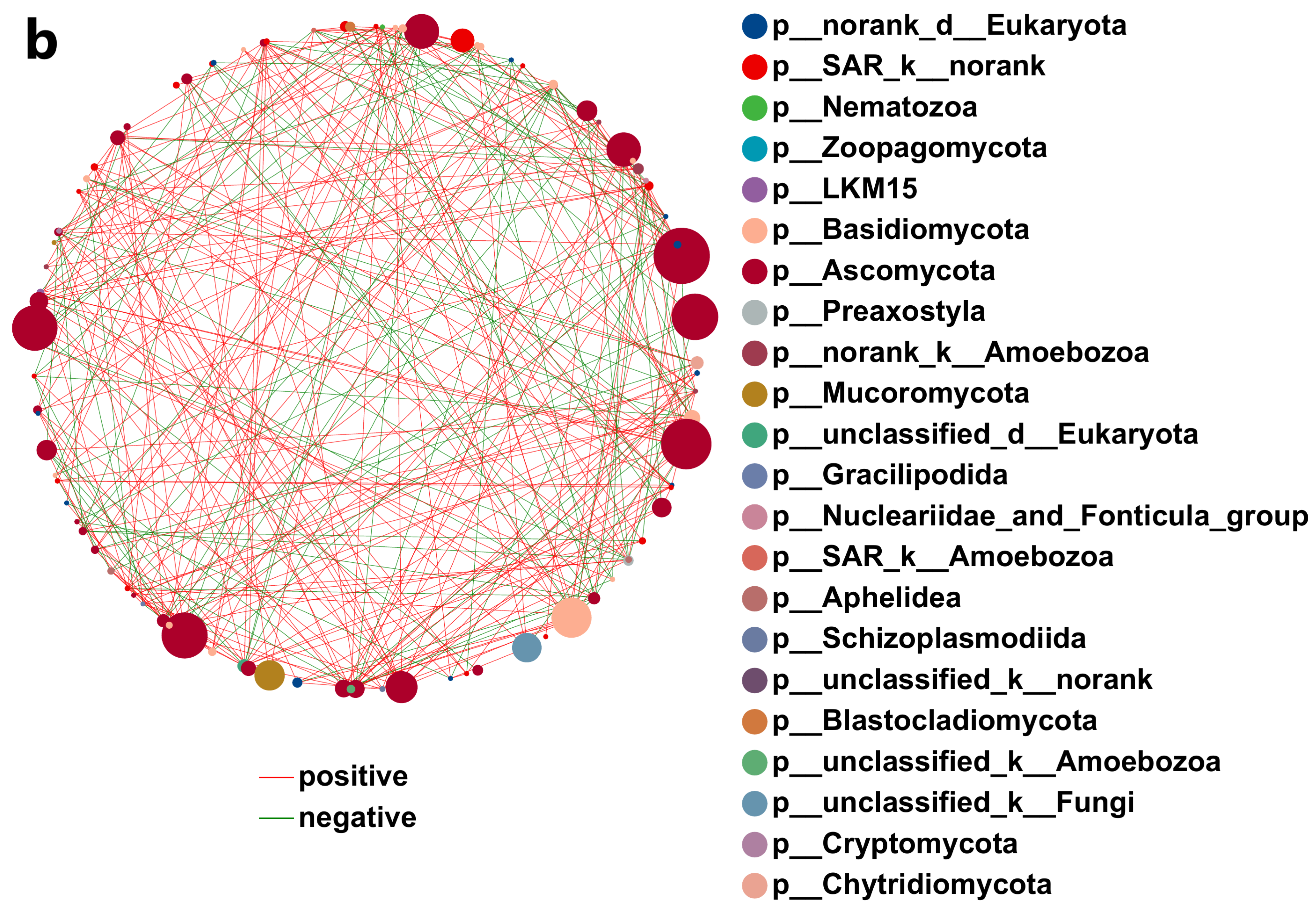
3.4.7. Network Analysis of Soil Microbial Correlations
4. Discussion
4.1. Effect of Adding Organic Material on Physiochemical Properties of Saline-Alkali Soils
4.2. Effect of Adding Iron Ions and Organic Material on Microbial Communities of Saline-Alkali Soils
4.3. Effects of Organic Material Addition on Soil Fungal Communities in Saline-Alkali Soils
4.4. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, X.D.; Yaa, O.K.; Wu, J.G. Effects of Different Organic Materials Application on Soil Physicochemical Properties in a Primary Saline-Alkali Soil. Eurasian Soil Sci. 2020, 53, 798–808. [Google Scholar] [CrossRef]
- Gong, L.J.; Li, X.F.; Wu, S.; Jiang, L.Q. Prediction of potential distribution of soybean in the frigid region in China with MaxEnt modeling. Ecol. Inform. 2022, 72, 101834. [Google Scholar] [CrossRef]
- Colombo, C.; Palumbo, G.; He, J.Z.; Pinton, R.; Cesco, S. Review on Iron Availability in Soil: Interaction of Fe Minerals, Plants, and Microbes. J. Soils Sediments 2014, 14, 538–548. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops–What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Wang, S.; Chen, Q.; Li, Y.; Zhang, N.; Zhao, B.; Zhuo, Y.; Chen, C. Experimental Investigation on Distribution Profiles of Heavy Metals in the Desulfurization Gypsum Amended Saline and Sodic Soils. Ecol. Environ. Sci. 2013, 22, 851–856. [Google Scholar]
- Ye, X.X.; Wang, H.; Cao, X.L.; Jin, X.J.; Cui, F.Q.; Bu, Y.Y.; Liu, H.; Wu, W.W.; Takano, T.; Liu, S.K. Transcriptome profiling of Puccinellia tenuiflora during seed germination under a long-term saline-alkali stress. BMC Genom. 2019, 20, 589. [Google Scholar] [CrossRef]
- Shilev, S. Plant-growth-promoting bacteria mitigating soil salinity stress in plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Sun, Y.T.; Zhou, L.R.; Meng, Q.F.; Zhao, J.; Ma, X.F. Effects of long-term manure application on the distribution of soil water-stable aggregate, organic carbon, and nitrogen in meadow alkaline soils. Soil Fertil. Sci. China 2014, 1, 6–23. [Google Scholar]
- Liang, C.J.; Bruell, C.J.; Marley, M.C.; Sperry, K.L. Persulfate oxidation for in situ remediation of TCE. II. Activated by chelated ferrous ion. Chemosphere 2004, 55, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wang, M.H.; Chen, L.Y.; Liang, X.J.; Wu, Z.G.; Lin, Z.H.; Zuo, J.; Feng, X.Y.; Zhao, J.; Liao, H.; et al. Soybean Fe-S cluster biosynthesis regulated by external iron or phosphate fluctuation. Plant Cell Rep. 2015, 34, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Zabaleta, R.; Fabani, M.P.; Rodriguez, R.; Mazza, G. Effects of the amendment with almond shell, bio-waste and almond shell-based biochar on the quality of saline-alkali soils. J. Environ. Manag. 2022, 318, 115604. [Google Scholar] [CrossRef]
- Singh, H.; Northup, B.K.; Rice, C.W.; Prasad, P.V.V. Biochar applications influence soil physical and chemical properties, microbial diversity, and crop productivity: A meta-analysis. Biochar 2022, 4, 8. [Google Scholar] [CrossRef]
- Liang, J.P.; Li, Y.; Si, B.C.; Wang, Y.Z.; Chen, X.G.; Wang, X.F.; Chen, H.R.; Wang, H.R.; Zhang, F.C.; Bai, Y.G.; et al. Optimizing biochar application to improve soil physical and hydraulic properties in saline-alkali soils. Sci. Total Environ. 2021, 771, 144802. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.S.; Liu, X.M.; Tang, J.C. Enhanced Degradation of Petroleum Hydrocarbons by Immobilizing Multiple Bacteria on Wheat Bran Biochar and Its Effect on Greenhouse Gas Emission in Saline-Alkali Soil. Chemosphere 2022, 286, 131663. [Google Scholar] [CrossRef]
- Luo, C.F.; Zhang, X.R.; Gong, Z.Q.; Jia, Y.J.; Zhao, X.; Liu, F.F.; Guo, W. Effects of a combination of activated iron tailings and magnesium-modified biochar on rice seedling growth and properties of saline-alkali soil. J. Agro-Environ. Sci. 2024, 43, 68. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Esmail, S.E.; El-Attar, A.B.; Othman, E.Z.; El-Bahbohy, R.M. Prospective Practice for Compound Stress Tolerance in Thyme Plants Using Nanoparticles and Biochar for Photosynthesis and Biochemical Ingredient Stability. Agronomy 2022, 12, 1069. [Google Scholar] [CrossRef]
- Gao, Y.; Shao, G.C.; Yang, Z.; Zhang, K.; Lu, J.; Wang, Z.Y.; Wu, S.Q.; Xu, D. Influences of Soil and Biochar Properties and Amount of Biochar and Fertilizer on the Performance of Biochar in Improving Plant Photosynthetic Rate: A Meta-Analysis. Eur. J. Agron. 2021, 130, 126345. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Li, G.; Andersen, M.N.; Liu, F. Biochar Enhances Yield and Quality of Tomato Under Reduced Irrigation. Agric. Water Manag. 2014, 138, 37–44. [Google Scholar] [CrossRef]
- Singh, K. Microbial and Enzyme Activities of Saline and Sodic Soils. Land Degrad. Dev. 2016, 27, 706–718. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Wu, L.N.; Jiang, Y.; Zhao, F.Y.; He, X.F.; Liu, H.F.; Yu, K. Increased organic Fertilizer application and reduced chemical Fertilizer application affect the soil properties and bacterial communities of grape rhizosphere soil. Sci. Rep. 2020, 10, 9568. [Google Scholar] [CrossRef]
- Merl, T.; Rasmussen, M.R.; Koch, L.R.; Sondergaard, J.V.; Bust, F.F.; Koren, K. Measuring soil pH at in situ like conditions using optical pH sensors (pH-optodes). Soil Biol. Biochem. 2022, 175, 108862. [Google Scholar] [CrossRef]
- Li, X.J.; Wang, X.; Zhao, Q.; Zhang, Y.Y.; Zhou, Q.X. In situ representation of soil/sediment conductivity using electrochemical impedance spectroscopy. Sensors 2016, 16, 625. [Google Scholar] [CrossRef]
- Hui, R.; Zhao, R.M.; Liu, L.C.; Li, X.R. Effect of snow cover on water content, carbon and nutrient availability, and microbial biomass in complexes of biological soil crusts and subcrust soil in the desert. Geoderma 2022, 406, 115505. [Google Scholar] [CrossRef]
- Fang, H.Y. Effect of soil conservation measures and slope on runoff, soil, TN, and TP losses from cultivated lands in northern China. Ecol. Indic. 2021, 126, 107677. [Google Scholar] [CrossRef]
- Huang, T.T.; Yang, N.; Lu, C.; Qin, X.L.; Siddique, K.H.M. Soil organic carbon, total nitrogen, available nutrients, and yield under different straw returning methods. Soil Tillage Res. 2021, 214, 105171. [Google Scholar] [CrossRef]
- Bao, S.D. Analysis on Soil and Agricultural Chemistry; China Agricultural Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Hou, M.H.; Fan, F.; Zhang, Q.G.; Tai, J.C.; Su, Y.L.Q.Q.G.; Yan, J. Analysis of soil alkalization characteristics of different vegetation types in saline-alkali soil of Xiliao river province. J. Heilongjiang Bayi Agric. 2022, 34, 39–46. [Google Scholar] [CrossRef]
- Dey, G.; Banerjee, P.; Sharma, R.K.; Maity, J.P.; Etesami, H.; Shaw, A.K.; Huang, Y.-H.; Huang, H.-B.; Chen, C.-Y. Management of phosphorus in salinity-stressed agriculture for sustainable crop production by salt-tolerant phosphate-solubilizing bacteria—A review. Agronomy 2021, 11, 1552. [Google Scholar] [CrossRef]
- Bartsev, S.I.; Pochekutov, A.A. The vertical distribution of soil organic matter predicted by a simple continuous model of soil organic matter transformations. Ecol. Model. 2016, 328, 95–98. [Google Scholar] [CrossRef]
- Li, Q.W.; Zhang, L.W.; Ji, W.W.; Chang, Z.X.; Chong, R.F.; Li, D.L.; Li, J.Y. Feasibility of using ammonium iron (II) sulphate to passivate hexavalent chromium in polluted soil. Environ. Technol. 2020, 43, 1522–1531. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ben-Laouane, R.; Lahlali, R.; Anli, M.; Ikan, C.; Boutasknit, A.; Slimani, A.; Oufdou, K.; Baslam, M.; Ait Barka, E.; et al. Application of indigenous rhizospheric microorganisms and local compost as enhancers of lettuce growth, development, and salt stress tolerance. Microorganisms 2022, 10, 1625. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, S.S.; Naresh, R.K.; Mandal, A.; Singh, R.; Dhaliwal, M.K. Dynamics and transformations of micro-nutrients in agricultural soils as influenced by organic matter build-up: A review. Environ. Sustain. Indic. 2019, 1, 100007. [Google Scholar] [CrossRef]
- Vejan, P.; Khadiran, T.; Abdullah, R.; Ahmad, N. Controlled Release Fertilizer: A Review on Developments, Applications, and Potential in Agriculture. J. Control. Release 2021, 339, 321–334. [Google Scholar] [CrossRef]
- Wang, J.; Liao, L.R.; Wang, G.L.; Liu, H.F.; Wu, Y.; Liu, G.B.; Zhang, C. N-Induced Root Exudates Mediate the Rhizosphere Fungal Assembly and Affect Species Coexistence. Sci. Total Environ. 2022, 804, 150148. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Fahad, S.; Wu, L.; Zhou, W.; Xu, P.; Sun, Z.; Salam, A.; Imran, M.; Jiang, M.D.; Kuzyakov, Y.; et al. Labile Organic Matter Intensifies Phosphorus Mobilization in Paddy Soils by Microbial Iron(III) Reduction. Geoderma 2019, 352, 185–196. [Google Scholar] [CrossRef]
- Siedliska, A.; Baranowski, P.; Pastuszka-Woźniak, J.; Zubik, M.; Krzyszczak, J. Identification of plant leaf phosphorus content at different growth stages based on hyperspectral reflectance. BMC Plant Biol. 2021, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Yang, L.; Guo, J.; Yang, J.F.; Li, N.; Dai, J.; Feng, H.; Liu, N.; Han, X. Contrasting Effects of Nitrogen and Organic Fertilizers on Iron Dynamics in Soil after 38–Year Fertilization Practice. Agronomy 2023, 13, 371. [Google Scholar] [CrossRef]
- Jin, Z.; Ottaviani, M.; Sikand, M. Modeling Sea Ice Albedo and Transmittance Measurements with a Fully-Coupled Radiative TransFer Model. Opt. Express 2023, 31, 21128–21152. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Medha, I.; Bhattacharya, J. Potassium-iron rice straw biochar composite for sorption of nitrate, phosphate, and ammonium ions in soil for timely and controlled release. Sci. Total Environ. 2020, 712, 136337. [Google Scholar] [CrossRef]
- Makete, N.; Rizzu, M.; Seddaiu, G.; Gohole, L.; Otinga, A. Fluoride toxicity in cropping systems: Mitigation, adaptation strategies and related mechanisms. A review. Sci. Total Environ. 2022, 833, 155129. [Google Scholar] [CrossRef]
- Kutovaya, O.V.; Vasilenko, E.S.; Lebedeva, M.P. Microbiological and micromorphological characteristics of extremely arid desert soils in the Ili Depression (Kazakhstan). Eurasian Soil Sci. 2012, 45, 1147–1158. [Google Scholar] [CrossRef]
- Lurthy, T.; Pivato, B.; Lemanceau, P.; Mazurier, S. Importance of the Rhizosphere Microbiota in Iron Biofortification of Plants. Front. Plant Sci. 2021, 12, 744445. [Google Scholar] [CrossRef] [PubMed]
- Rivkina, E.M.; Fedorov-Davydov, D.G.; Zakharyuk, A.G.; Shcherbakova, V.A.; Vishnivetskaya, T.A. Free Iron and Iron-Reducing Microorganisms in Permafrost and Permafrost-Affected Soils of Northeastern Siberia. Eurasian Soil Sci. 2020, 53, 1455–1468. [Google Scholar] [CrossRef]
- Rehman, A.U.; Nazir, S.; Irshad, R.; Tahir, K.; Rehman, K.; Islam, R.U.; Wahab, Z. Toxicity of Heavy Metals in Plants and Animals and Their Uptake by Magnetic Iron Oxide Nanoparticles. J. Mol. Liq. 2021, 321, 114455. [Google Scholar] [CrossRef]
- Hussain, I.; Afzal, S.; Ashraf, M.A.; Rasheed, R.; Saleem, M.H.; Alatawi, A.; Fahad, S. Effect of Metals or Trace Elements on Wheat Growth and Its Remediation in Contaminated Soil. J. Plant Growth Regul. 2023, 42, 2258–2282. [Google Scholar] [CrossRef]
- Bhat, B.A.; Islam, S.T.; Ali, A.; Sheikh, B.A.; Tariq, L.; Islam, S.U.; Hassan Dar, T.U. Role of Micronutrients in Secondary Metabolism of Plants. In Plant Micronutrients: Deficiency and Toxicity Management; Springer: Berlin/Heidelberg, Germany, 2020; pp. 311–329. [Google Scholar]
- Pattnaik, S.; Mohapatra, B.; Gupta, A. Plant Growth-Promoting Microbe Mediated Uptake of Essential Nutrients (Fe, P, K) for Crop Stress Management: Microbe–Soil–Plant Continuum. Front. Agron. 2021, 3, 689972. [Google Scholar] [CrossRef]
- Adegbeye, M.J.; Reddy, P.R.K.; Obaisi, A.I.; Elghandour, M.M.M.Y.; Oyebamiji, K.J.; Salem, A.Z.M.; Camacho-Díaz, L.M. Sustainable Agriculture Options for Production, Greenhouse Gases and Pollution Alleviation, and Nutrient Recycling in Emerging and Transitional Nations—An Overview. J. Clean. Prod. 2020, 242, 118319. [Google Scholar] [CrossRef]
- Su, C.; Zhang, M.L.; Lin, L.Y.; Yu, G.W.; Zhong, H.T.; Chong, Y.X. Reduction of iron oxides and microbial community composition in iron-rich soils with different organic carbon as electron donors. Int. Biodeterior. Biodegrad. 2020, 148, 104881. [Google Scholar] [CrossRef]
- Kim, H.; Jeon, J.; Lee, K.K.; Lee, Y.H. Compositional Shift of Bacterial, Archaeal, and Fungal Communities Is Dependent on Trophic Lifestyles in Rice Paddy Soil. Front. Microbiol. 2021, 12, 719486. [Google Scholar] [CrossRef]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef]
- Jing, J.Y.; Cong, W.F.; Bezemer, T.M. Legacies at work: Plant–soil–microbiome interactions underpinning agricultural sustainability. Trends Plant Sci. 2022, 27, 961–963. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.L.; Schartiger, R.G.; Daniels, N.K.; Freedman, Z.B.; McDonald, L.M.; Skousen, J.G.; Morrissey, E.M. Bioenergy crop Miscanthus x giganteus acts as an ecosystem engineer to increase bacterial diversity and soil organic matter on marginal land. Soil Biol. Biochem. 2023, 186, 109178. [Google Scholar] [CrossRef]
- Sahu, P.K.; Singh, D.P.; Prabha, R.; Meena, K.K.; Abhilash, P.C. Connecting Microbial Capabilities with the Soil and Plant Health: Options for Agricultural Sustainability. Ecol. Indic. 2019, 105, 601–612. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, X.M.; He, N.; Zhang, J.; Wu, J.; Liu, C.S. Shifts in Microbial Communities and Networks Are Correlated with the Soil Ionome in a Kiwifruit Orchard under Different Fertilization Regimes. Appl. Soil Ecol. 2020, 149, 103517. [Google Scholar] [CrossRef]
- Wang, X.K.; Wang, G.; Guo, T.; Xing, Y.Y.; Mo, F.; Wang, H.D.; Fan, J.L.; Zhang, F.C. Effects of Plastic Mulch and Nitrogen Fertilizer on the Soil Microbial Community, Enzymatic Activity, and Yield Performance in a Dryland Maize Cropping System. Eur. J. Soil Sci. 2021, 72, 400–412. [Google Scholar] [CrossRef]
- Cavard, X.; Bergeron, Y.; Paré, D.; Nilsson, M.C.; Wardle, D.A. Disentangling Effects of Time Since Fire, Overstory Composition and Organic Layer Thickness on Nutrient Availability in Canadian Boreal Forest. Ecosystems 2022, 164, 108495. [Google Scholar] [CrossRef]
- Mensah, A.K. Arsenic Contamination from Gold Mining and Remediation of Active and Abandoned Mine Spoils in Ghana. Ph.D. Thesis, Ruhr-Universität Bochum, Bochum, Germany, 2022. [Google Scholar]
- Yin, Y.N.; Wang, J.L. Enhanced biohydrogen production from macroalgae by zero-valent iron nanoparticles: Insights into microbial and metabolites distribution. Bioresour. Technol. 2019, 282, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Wang, H.D.; Liu, C.; Sun, B.B.; Zheng, J.F.; Bian, R.J.; Drosos, M.; Zhang, X.H.; Li, L.Q.; Pan, G.X. Biochar increases maize yield by promoting root growth in the rainFed region. Arch. Agrono Soil Sci. 2021, 67, 1411–1424. [Google Scholar] [CrossRef]
- Du, T.Y.; He, H.Y.; Zhang, Q.; Lu, L.; Mao, W.J.; Zhai, M.Z. Positive effects of organic Fertilizers and bioFertilizers on soil microbial community composition and walnut yield. Appl. Soil Ecol. 2022, 175, 104457. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.X.; Yuan, Y.Z.; Wang, K.Y.; Chen, Y.; Wang, H.Y. Control efficiency of hexaconazole-lentinan against wheat sharp eyespot and wheat crown rot and the associated effects on rhizosphere soil fungal community. Front. Microbiol. 2022, 13, 1014969. [Google Scholar] [CrossRef]
- Bhunia, S.; Bhowmik, A.; Mallick, R.; Mukherjee, J. Agronomic efficiency of animal-derived organic Fertilizers and their effects on biology and Fertility of soil: A review. Agronomy 2021, 11, 823. [Google Scholar] [CrossRef]
- Cui, H.Y.; Sun, W.; Delgado-Baquerizo, M.; Song, W.Z.; Ma, J.Y.; Wang, K.Y.; Ling, X.L. The effects of mowing and multi-level N Fertilization on soil bacterial and fungal communities in a semiarid grassland are year-dependent. Soil Biol. Biochem. 2020, 151, 108040. [Google Scholar] [CrossRef]
- Fatima, F.; Ahmad, M.M.; Verma, S.R.; Pathak, N. Relevance of phosphate solubilizing microbes in sustainable crop production: A review. Int. J. Environ. Sci. Technol. 2020, 19, 9283–9296. [Google Scholar] [CrossRef]
- Mann, C.; Lynch, D.; Fillmore, S.; Mills, A. Relationships between field management, soil health, and microbial community composition. Appl. Soil Ecol. 2019, 144, 12–21. [Google Scholar] [CrossRef]
- Xiao, G.F.; Cheng, X.J.; Zhu, D.T.; Li, Z.F.; Feng, L.J.; Peng, X.M.; Lu, Z.Y.; Xie, J. Exploring the mechanism of a novel recirculating aquaculture system based on water quality parameters and bacterial communities. Environ. Sci. Pollut. Res. 2023, 30, 34760–34774. [Google Scholar] [CrossRef]
- Ge, Z.; Li, S.Y.; Bol, R.; Zhu, P.; Peng, C.; An, T.T.; Chen, N.; Liu, X.; Li, T.Y.; Xu, Z.Q.; et al. Differential Long-Term Fertilization Alters Residue-Derived Labile Organic Carbon Fractions and Microbial Community during Straw Residue Decomposition. Soil Tillage Res. 2021, 213, 105120. [Google Scholar] [CrossRef]
- Giri, S.; Waschina, S.; Kaleta, C.; Kost, C. Defining Division of Labor in Microbial Communities. J. Mol. Biol. 2019, 431, 4712–4731. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC uS cm−1 | SOM g kg−1 | TN g kg−1 | TP g kg−1 | TK g kg−1 | TF mg kg−1 | |
|---|---|---|---|---|---|---|---|
| content | 8.52 | 59.51 | 59.24 | 0.33 | 0.42 | 2.17 | 95.24 |
| Sample Amendments | Fertilization Measures |
|---|---|
| ck | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1 |
| Fe | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1 |
| OF0.5 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; organic: 0.5 t/1000 m2 |
| OF1 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; organic: 1 t/1000 m2 |
| OF2 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; organic: 2 t/1000 m2 |
| OF5 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; organic: 5 t/1000 m2 |
| C0.5 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; Bio-charcoal: 26 g kg−1 |
| C1 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; Bio-charcoal: 52 g kg−1 |
| C2 | 60 N kg ha−1; 75 P kg ha−1; 75 K kg ha−1; FeSO4: 15 g kg−1; Bio-charcoal: 104 g kg−1 |
| Sample | Bacteria | Fungi | ||||
|---|---|---|---|---|---|---|
| Shannon | Ace | Chao1 | Shannon | Ace | Chao1 | |
| ck | 6.82 b | 3948.82 c | 3885.41 c | 2.98 ab | 180.92 b | 182.40 b |
| Fe | 6.86 b | 4149.32 b | 4222.43 b | 2.88 b | 194.41 a | 191.18 a |
| OF0.5 | 6.75 bc | 4100.96 b | 4135.97 b | 3.19 ab | 159.85 d | 158.07 c |
| OF1 | 6.89 b | 4291.27 ab | 4265.37 b | 3.29 a | 184.80 b | 183.56 b |
| OF2 | 7.01 a | 4503.23 a | 4588.71 a | 2.99 ab | 173.38 c | 172.71 b |
| OF5 | 6.99 b | 4613.84 a | 4545.74 a | 3.31 a | 197.87 a | 210.08 a |
| C0.5 | 4.54 c | 3567.28 d | 3582.38 d | 2.75 c | 197.77 a | 198.16 a |
| C1 | 6.89 b | 4271.97 ab | 4304.24 b | 3.10 ab | 184.52 b | 195.50 a |
| C2 | 6.83 b | 4205.12 b | 4204.11 b | 3.20 a | 173.66 c | 178.27 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazl, U.; Wang, J.; Yin, J.; Jiang, X.; Meng, F.; Zhang, W.; Zhang, L.; Zhao, H. Effects of FeSO4 and Organic Additives on Soil Properties and Microbiota during Model Soybean Planting in Saline-Alkali Soil. Agronomy 2024, 14, 1553. https://doi.org/10.3390/agronomy14071553
Fazl U, Wang J, Yin J, Jiang X, Meng F, Zhang W, Zhang L, Zhao H. Effects of FeSO4 and Organic Additives on Soil Properties and Microbiota during Model Soybean Planting in Saline-Alkali Soil. Agronomy. 2024; 14(7):1553. https://doi.org/10.3390/agronomy14071553
Chicago/Turabian StyleFazl, Ullah, Jian Wang, Jiamin Yin, Xinbo Jiang, Fangang Meng, Wei Zhang, Liqiang Zhang, and Hongyan Zhao. 2024. "Effects of FeSO4 and Organic Additives on Soil Properties and Microbiota during Model Soybean Planting in Saline-Alkali Soil" Agronomy 14, no. 7: 1553. https://doi.org/10.3390/agronomy14071553
APA StyleFazl, U., Wang, J., Yin, J., Jiang, X., Meng, F., Zhang, W., Zhang, L., & Zhao, H. (2024). Effects of FeSO4 and Organic Additives on Soil Properties and Microbiota during Model Soybean Planting in Saline-Alkali Soil. Agronomy, 14(7), 1553. https://doi.org/10.3390/agronomy14071553