Abstract
Maize is among the most significant crops in the world regarding production and yield, but it is highly sensitive to drought, which reduces the growth, photosynthetic efficiency, grain quality, and yield production of a plant. Quantum yield efficiency of photosystem II is a critical photosynthetic component that is susceptible to drought stress. This study intended to investigate the effects of drought stress on growth and morpho-physiological parameters using three maize hybrids (‘P-3011w’, ‘P-3092’ and ‘iku20’) with contrasting soil moisture contents (100%, 40%) at the pre-flowering stage. The stress treatment (40%) was initiated at stage V7, for a period of 15 days; the experimental units were established in a completely randomized design with split-plot arrangement along with three repetitions in 42 L pots using a substrate of peat moss, black soil and poultry manure (1:2:1). The morphological, growth-related and physiological parameters were assessed, including chlorophyll fluorescence (Fv/Fm), which was measured using a LiCor-6400-40 fluorometer. The results showed that all morphological, growth-related and physiological variables decreased under drought stress during the reproductive stage, with the exception of leaf temperature and intercellular CO2 concentration, which increased by 12% and 54%, respectively. Drought stress significantly reduced the photosynthetic chlorophyll fluorescence (43%), due to damage to photosystem II. The lowest percentage of damage to photosystem II (34%) was observed in the iku20 genotype. In contrast, P-3011w and P-3092 had the highest levels of significantly similar damage (49% and 46%, respectively). The correlation analysis showed a highly positive interaction of chlorophyll fluorescence (Fv/Fm) with net photosynthetic rate and stomatal conductance under drought conditions, and multiple regression analysis revealed that the maximum effect on net photosynthetic rate under drought was due to the damage it caused to photosystem II. Thus, iku20 might have a tendency to be able to withstand drought stress in the dry northeast region of Mexico. Overall, we concluded that the photosystem II was negatively impacted by drought stress thus causing a reduction in all physiological, morphological and growth-related variables.
1. Introduction
Maize is among the most extensively cultivated crops globally grown in tropical, sub-tropical and temperate regions [1]. After rice and wheat, it was the third-most significant crop in the world with a 1222.07 million metric ton production on a cultivated area of 201.98 million hectares [2]. Maize is mostly consumed by the people in Mexico in a form of variety of tejuino, pinole, tortillas, tamales, tostadas, atole and tortilla chips [3]. In 2022, the consumption of maize amounted to 1740 million bushels in Mexico [2]. In the northeastern part of Mexico, a majority of the irrigated maize area is cultivated with hybrids [4]. In addition to having a moisture level of less than 70%, the grains have 18–20% carbs, 5–6% sugars and 2.1–4.5% proteins. Livestock is fed the leftover fodder [5]. Różewicz [6] has mentioned that when compared to other cereal grains, maize offers the best feed value for poultry. Due to its high fat content (44.0 g kg−1) and starch content (64.5% to 71.5%), it has a significant quantity of metabolic energy (14.8 MJ kg−1). Farmers who raise livestock view maize plants as a superior source of nourishment for their animals. Because of its high dry matter content and digestible elements in comparison to other crops, it is the perfect crop to utilize as silage or fodder. Even yet, compared to colder regions, silage is less frequently used in tropical areas. Animals can also be fed the maize stover that is left in the field after the grain is harvested [7]. In Mexico, farmers use their maize production as a family food, feed for animals, for sales and as a seed to grow in the following season; however, some of the production is lost due to crop losses. Framers that grow maize on a small scale use their production for food, feed and sales in similar proportions (23–35%); medium-scale farmers use only 5% of their produce as food, 33% for sales and 25% for feed. Only 0.1% of the maize produced by large-scale farmers is used for food, 8% for feed and the majority, 89%, is sold [8].
In contrast to other grains such as sorghum and millet, maize is more vulnerable to water deficiency. For instance, compared to a 39% yield loss in sorghum, drought stress can result in a 66% loss in maize yield [9,10]. According to a multi-model approach (climate and crop), in the future, the heat will increase due to the current climate change scenarios caused by global warming, which will cause unexpected rainfall events and it would lead to a significant reduction in the yield of maize by the middle of the century [11] due to a decline in the favorable climate in the far future that makes maize farming less viable [12]. The local agricultural economies have been impacted directly by this climate change. For instance, 87% of maize farmers in the northeastern region of Mexico experienced adverse consequences from weather events and climate change [13]. Król-Badziak et al. [12] also projected that the future water shortages brought on by present climate change would reduce the climate’s appropriateness for maize cultivation, perhaps rendering the regions now used for maize production unusable. The largest economic loss of USD 58 billion worldwide is thought to result from drought stress, which affected 124 million hectares of the maize-growing region by cutting 40% of its production [14,15]. The photosynthetic process is majorly impacted by drought, which is among the most critical physiological processes [16,17]. After photosynthesis inhibition, more light energy is absorbed by the plant as compared to the energy required for photosynthetic carbon fixation [18,19]; the electron transport chain between photosystem II and photosystem I is potentially triggered by this excess energy leading to a rise in reactive oxygen species [20]. Since climate change has increased the likelihood of drought stress, soil cultivation techniques like strip-tillage and no-tillage have become more popular as protective measures against it [21]. According to studies by Stępień-Warda [22], in water-deficient soil, maize production was greater when reduced tillage was combined with direct rather than full tillage. Furthermore, with no-tillage farming, photosystem II performed more effectively, as indicated by the maize yield. The lifecycle of maize has many stages whereby water deficiency stress disrupts the crop’s growth and lowers its output. The two crucial maize stages for determining yield are vegetative flowering and grain filling [23,24] and drought stress during these essential periods can cause a yield loss of up to 25%, 50% and 21%, respectively [25]. Furthermore, plants under stress during blooming and early grain filling stages have hindered the supply of starch which limits the grain’s access to assimilates during seed formation [26].
Water deficiency had a significantly negative impact on stomatal conductance in plants [27]. A notable decrease in transpiration rate and leaf water potential was seen because of water deficiency, leading to increased leaf and canopy temperatures. Because stomatal closure caused by drought stress decreases CO2 accessibility, plants are more vulnerable to light damage [28]. Damage to chlorophyll content and thylakoid membrane is caused by drought stress [25], whereas a decreased chlorophyll content reduces photosynthesis [29]. The quantum efficiency of photosystem II (PSII) is a fundamental component of photosynthetic processes, which is represented as a ratio between variable and maximum leaf chlorophyll fluorescence (Fv/Fm) [30]. Analysis of chlorophyll fluorescence is a very effective and widely used approach for estimating the effect of stress on photosynthetic equipment, as indicated by the availability of several non-invasive, easy-to-use, and portable fluorometers. Although the use of these fluorometers is rather easy, the science underlying it and interpreting the data received from these measurements remains challenging [31,32]. The Fv/Fm ratio was the first significant parameter to be extracted from the Kautsky curve [33]. Later on, it was used as a crucial parameter to identify photosystem II photoinhibition caused by stress [34]. This ratio is calculated by measuring the yield of minimal fluorescence (Fo) in a leaf adapted to dark using a weakly modulated measuring beam and then inducing the maximum production of chlorophyll fluorescence (Fm) by superimposing a saturating flux. A measure of PSII’s maximum photochemical efficiency is performed using the ratio Fv/Fm [(Fm − Fo)/Fm, to detect the photosystem II reaction center’s loss of function. Fv/Fm values usually fall between 0.75 and 0.85, and this ratio is directly related to photochemistry’s quantum yield [35]. The damage to PSII by water deficiency stops the main photochemistry by interfering with the photosynthetic electron transport [36,37]. A decrease in the Fv/Fm value is indicative of reduced photosystem II efficiency, i.e., photoinhibition [38]. Despite the fact that occasionally Fv/Fm is mistakenly interpreted as a sign of PS II photoinactivation, it ought to be mentioned that this ratio also decreases when competing processes, like the thermal dissipation of absorbed light, interfere with the separation of charge [39]. The measurement of effective chlorophyll fluorescence in fields under natural light conditions has been supported by the advancements made in fluorometer technology. Due to a lack of dark adaptation, this parameter has been considered a useful indication of stress in plants. However, it is still challenging to measure the fluorescence under natural light, an advanced MONITORING-PAM would be a reliable instrument for measuring the effective quantum efficiency of photosystem II during daylight hours [40].
We hypothesized that drought stress might induce negative impacts on morpho-physiological parameters among maize cultivars by causing damage to photosystem II. Therefore, the comparative study presented here was conducted to report the morpho-physiological responses to drought stress in maize hybrids during the reproductive stage and to evaluate if water deficiency results in statistically significant damage to photosystem II.
2. Materials and Methods
2.1. Plant Material and Site Description
The morphological, growth-related and physiological response of three maize hybrids, P-3011w, P-3092 and iku20, under drought stress, was assessed. The seeds of P-3011w and P-3092 were obtained from the local farmers in Miguel Aleman, Tamaulipas, Mexico whereas the seeds of iku20 were obtained from AgroRivaz, Ocotlan, Jalisco, Mexico. Among these three hybrids, P-3011w and P-3092 are the most commonly used hybrids in the northeastern part of Mexico and iku20, a newly developed single cross hybrid was used to see its behavior in the northeastern region of Mexico. The study was conducted at the Facultad de Agronomía of the Universidad Autonoma de Nuevo León in Marín, Nuevo León, Mexico (at 24°19′16.71″ N and 99°54′58.06″ W) from February to May 2023. Thirty seeds of each hybrid were sown in germination trays on 17 February 2023. Subsequently, the seedlings were transplanted into 42 L cylindrical pots (length = 56 cm and diameter = 30 cm) with a substrate mixture of peat moss, black soil and poultry manure (1:2:1) having a field capacity of 45%. Each pot contained 26 kg substrate with one plant per pot. All pots were optimally watered (100% irrigation) until the V7 (pre-tasseling) stage to avoid drought stress.
2.2. Stress Treatment
For the application of stress treatment, plants were divided into two irrigation levels, i.e., normal irrigation (100% field capacity) and reduced irrigation (40% field capacity) [41]. The daytime and nighttime temperatures were recorded at 35 °C and 22 °C, respectively. When plants reached the V7 (Pre-tasseling) stage, drought stress treatment was initiated for fifteen days by providing enough water to maintain a humidity percentage of 40%. The water quantity required to maintain the humidity levels at 100% and 40% was estimated by a weighing method [42]. After fifteen days of drought stress, when the plants were at reproductive stage, the physiological, morphological and plant growth related measurements were made.
2.3. Measurement of Morphological and Growth-Related Variables
Morphological (height of plant, diameter of the stem) and growth-related (area of leaf; length, fresh and dry weight of root) measurements were taken. To determine the plant height, measurement was taken in cm from the base to the apex of the plant by a meter scale. The total leaf area was computed using the following equation [43].
where, LA = Leaf area, L = leaf length, W = leaf width and 0.75 is the correction factor for maize leaf area estimation.
A Vernier caliper (14,388 6 PuLG, Truper, Mexico City, Mexico) was used to measure the stem diameter of the region 15 cm above the ground [44]. In order to measure the length and fresh and dry weight of the roots, they were first sprayed with water to wash away any dirt particles. Then, they were placed on a level surface and, by using a meter scale, their length was measured in cm. An electronic weighing balance (Nimbus NBL 124E, AE Adam, CT, USA) was used to determine the fresh and dry weight of the roots. After measuring fresh weight, the roots were dried in an oven at 70 °C until constant weight [45] and were measured again for dry weight.
2.4. Measurement of Physiological Variables
2.4.1. Potential Quantum Yield Efficiency of PSII (Fv/Fm)
Measurements of Fv/Fm began 73 days following the date of sowing. A Li-6400-40 fluorometer (LI-COR Inc., Lincoln, NE, USA) was used on a flag leaf to measure the Fv/Fm [46]. Before making the measurement, each plant’s flag leaf was covered with a plastic bag for 20 minutes to open all the photosystem II reaction centers, which allowed the measurement of the minimal fluorescence (Fo) [47]. In order to prevent any light from getting to the dark-acclimated leaf while taking the readings, the whole leaf and the fluorometer chamber were kept covered. Fv/Fm was calculated as follows:
where: Fo = Minimal chlorophyll fluorescence after dark adaptation; Fm = Maximum chlorophyll fluorescence after dark adaptation; Fv = Variable leaf chlorophyll fluorescence after dark adaptation.
2.4.2. Photosynthetic Parameters
Photosynthesis and gas exchange variables including net photosynthetic rate (PN, µmol CO2 m−2 s−1), stomatal conductance (gs, mmol H2O m−2 s−1) transpiration rate (Tr, mmol H2O m−2 s−1), leaf temperature (Tleaf, °C) and intercellular CO2 concentration (Ci, µmol CO2 m−2 s−1) were measured on the flag leaf with a LiCor-6400 apparatus using a 6400-02B LED light source [10]. The calibration for taking the readings by the LiCor-6400 were [46]: Rate of flow: 700 µmol s−1, a constant CO2 level: 400 µmol CO2 and a 6400-02 LED light source: 1500 µmol m−2 s−1. The instantaneous water-use efficiency (WUEi) of the plant was estimated by PN/Tr [48].
2.5. Statistical Analysis
This study was designed as a split plot in a completely randomized design with three replications comprising a total of fifty-four pots. The replicated data normality was assessed using the Shapiro–Wilk test. The data that followed the normal distribution were then subjected to a two-way analysis of variance (ANOVA) by assuming the treatments to be the main plot factor and genotypes to be the sub-plot factor using the Statistix 10 software (Analytical Software, Tallahassee, FL, USA). The significant differences among the treatments with regard to genotypes were calculated at a 5% level of significance, and the mean comparison through the Tukey test (p < 0.05). Correlation and stepwise multiple regression analyses were conducted using SPSS (IBM SPSS 22.0, IBM Corporation, New York, NY, USA).
3. Results
All the morphological, growth-related and physiological variables varied significantly (p < 0.05) for the treatments and genotypes except for instantaneous water-use efficiency, which was significant for treatments only, whereas the genotype-treatment interaction was significant only in the quantum yield efficiency of photosystem II, leaf temperature and intercellular CO2 concentration (Table 1).

Table 1.
Mean squares of morphological, growth-related and physiological parameters in maize hybrids.
3.1. Morphological and Growth-Related Variables
Drought stress hampered the morphological growth of all genotypes studied. Compared with control, drought stress significantly reduced the plant height, leaf area, stem diameter, root length, and root fresh and dry weight as shown in Table 2. A reduction in the average values of plant height (13%), leaf area (5%), stem diameter (13%), root length (21%), and root fresh weight (12%) and root dry weight (26%) was recorded under drought compared to the control. This decrease in the average values of all variables indicates a decline in the plant’s vegetative development and root attributes. Under stress conditions, all the morphological and growth-related parameters were least affected in iku20 compared to P-3011w and P-3092.
Table 2.
Effect of drought stress on maize growth and morphology-related attributes.
3.2. Physiological Parameters
3.2.1. Potential Quantum Yield Efficiency of PSII (Fv/Fm) under Drought
Drought reduced Fv/Fm compared to control (Table 3). It showed significant (p < 0.05) variations for treatments, genotypes, and genotype-treatment interaction. The Fv/Fm values recorded varied from 0.77 to 0.80 with an average of 0.78 in control and 0.35 to 0.55 with an average of 0.44 in drought conditions. All the hybrids studied showed a significant decrease in Fv/Fm in drought compared to the control with the maximum reduction recorded in genotype P-3011w (0.39 ± 0.03), followed by P-3092 (0.42 ± 0.05) and iku20 (0.52 ± 0.05).
Table 3.
Mean Fv/Fm under control and drought stress.
A reduction of 43% in Fv/Fm was recorded in drought compared to the control. The genotype iku20 was least affected, showing a reduction of 34% as compared to P-3011w (49%) and P-3092 (46%) (Figure 1).
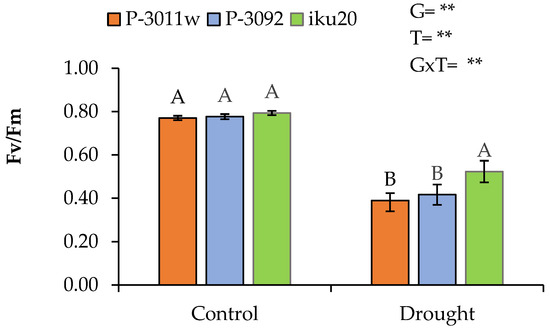
Figure 1.
Effects of drought stress on maize hybrids (P-3011w, P-3092, iku20) during reproductive stage on Fv/Fm. Means sharing same letters are not significantly different according to Tukey test (p < 0.05). ** Significance at p < 0.05.
3.2.2. Photosynthetic Variables under Drought Stress
The PN, gs, Tr and WUEi of all genotypes were negatively influenced by drought stress; therefore they showed reduced values compared to the control as shown in Table 4. All of these parameters were significantly different under control and drought stress for genotypes and treatments except WUEi, which was statistically different only for treatments. All these parameters did not have significant differences for the genotype-treatment interaction. The PN values ranged from 12.53 to 16.53 CO2 µmolm−2 s−1 under control conditions with an average of 14.18 CO2 µmolm−2 s−1 recorded whereas a range of 5.12–8.08 CO2 µmolm−2 s−1 was observed under drought stress with an average of 6.58 CO2 µmolm−2 s−1 recorded. The gs values varied from 0.07 to 0.10 H2O mmolm−2 s−1 and 0.03 to 0.07 H2O mmolm−2 s−1 under control and drought stress, respectively, with an average of 0.09 H2O mmolm−2 s−1 recorded under control conditions and 0.05 H2O mmolm−2 s−1 recorded in drought. The WUEi values ranged from 4.50 to 5.72 CO2 µmol H2O mmol with an average of 4.98 CO2 µmol H2O mmol whereas under drought stress the values varied from 4.05 to 4.96 CO2 µmol H2O mmol with an average of 4.36 CO2 µmol H2O mmol. The recorded values for Tr under control and drought stress were 2.32–3.56 H2O mmolm−2 s−1 and 1.21–1.88 H2O mmolm−2 s−1, respectively. Mean values recorded for transpiration were 2.88 H2O mmolm−2 s−1 under control conditions and 1.50 H2O mmolm−2 s−1 under drought stress. Among all maize hybrids, iku20 showed maximum values for photosynthesis (15.86), stomatal conductance (0.10) and transpiration (3.23) under the control conditions.
Table 4.
Means of net photosynthetic rate (PN), stomatal conductance (gs) and transpiration (Tr) under control and drought stress.
The drought stress decreased the PN (54%), gs (42%), Tr (48%) and WUEi (12%). Under stress conditions, genotype iku20 had reduced PN by 50%, gs by 31%, Tr by 47% and WUEi by 5%; genotype P-3092 had reduced PN by 55%, gs by 44%, Tr by 47% and WUEi by 18%; and genotype P-3011w had reduced PN, gs, Tr and WUEi by 57%, 52%, 50% and 14%, respectively (Figure 2a–d).


Figure 2.
Effects of drought stress on maize hybrids (P-3011w, P-3092, iku20) during pre-flowering stage on (a) photosynthetic rate (PN) (µmol CO2 m−2 s−1), (b) stomatal conductance (gs) (mmol H2O m−2 s−1), (c) transpiration (Tr) (mmol H2O m−2 s−1), (d) instantaneous water-use efficiency (WUEi) (CO2 µmol H2O mmol), (e) leaf temperature (Tleaf) (°C) and (f) intercellular CO2 concentration (Ci) (µmol CO2 m−2 s−1). Means sharing same letters do not have significant differences according to Tukey test (p < 0.05). ** Significance at p < 0.05.
Leaf temperature and intercellular CO2 concentration increased under drought conditions compared to the control as shown in Table 5. These parameters were influenced significantly by treatments and genotypes and had significant genotype–treatment interaction. The leaf temperature values measured varied from 28.80 to 30.87 °C with an average of 30.04 °C under control conditions whereas, in the case of drought stress, the values varied from 31.62 to 36.24 °C with an average of 33.67 °C. In the case of intercellular CO2 concentration, the values ranged from 80.52 to 91.07 CO2 µmolm−2 s−1 under control conditions and 129.98 to 135.77 CO2 µmolm−2 s−1 under drought stress with an average of 86.22 CO2 µmolm−2 s−1 and 132.99 CO2 µmolm−2 s−1 recorded under control and drought stress, respectively.
Table 5.
Means of leaf temperature (Tleaf) and intercellular CO2 concentration (Ci) under control and drought stress.
In all genotypes studied, the stress conditions increased leaf temperature and intercellular CO2 concentration. The leaf temperature and intercellular CO2 concentration increased by 12% and 54%, respectively. Iku20 had the lowest rise in leaf temperature (8%) when compared to P-3092 (11%) and P-3011w (17%). The greatest rise in intercellular CO2 concentration during drought stress was observed in P-3011w (64%), followed by P-3092 (56%), and iku20 (44%) (Figure 2e,f).
3.2.3. Correlation and Multiple Regression Analyses
The results of the correlation analysis of the selected morpho-physiological parameters under control conditions are shown in Table 6. All the parameters had significant correlation with Fv/Fm except for stem diameter and root dry weight. According to the thumb rule of correlation coefficient interpretation [49], under control conditions, the net photosynthetic rate showed a high positive correlation, and stomatal conductance, transpiration, plant height and leaf area had a moderate positive correlation with Fv/Fm. The only parameter that had a negative significant correlation with photosystem II was intercellular CO2 concentration (−0.422). Significantly high correlations were found between other variables as can be seen in Table 6.
Table 6.
Pearson correlation analysis of selected growth and morpho-physiological parameters of maize hybrids under control conditions.
The correlation analysis of selected morpho-physiological parameters under drought stress showed that all variables had a significant correlation with Fv/Fm except for intercellular CO2 concentration. Under stress conditions, stomatal conductance showed a remarkably high positive correlation with Fv/Fm followed by PN, whereas Tr, plant height, leaf area and root dry weight had moderate positive correlation with Fv/Fm (Table 7). Significantly high correlations were also found between other variables as can be seen in Table 7 [49].
Table 7.
Pearson correlation analysis of selected growth and morpho-physiological parameters of maize hybrids under drought stress.
The multiple regression analysis (stepwise method) of physiological parameters under control conditions showed that Fv/Fm, stomatal conductance and WUEi had significant impacts on the net photosynthetic rate as shown in Table 8. The analysis showed that almost 85% of the variation in net photosynthetic rate was due to Fv/Fm, stomatal conductance and WUEi (Table 9). Other variables had low contributions so they were not included in the analysis. The predicted equation for the net photosynthetic rate under control conditions is also mentioned in Table 9.
Table 8.
Analysis of variance of stepwise multiple regression of physiological variables under control conditions.
Table 9.
The regression coefficients (b), standard error and R square values of the estimated variables in predicting maize net photosynthetic rate under control conditions by stepwise multiple regression analysis.
The multiple regression analysis of physiological parameters under drought stress using net photosynthetic rate as a dependent variable showed that Fv/Fm, transpiration and WUEi had a significant impact on net photosynthetic rate as shown in Table 10. According to the results, 94% of the variation in net photosynthetic rate was attributed to Fv/Fm, transpiration and WUEi (Table 11). The other variables that had low relative contributions were not included in the analysis. The predicted equation for the net photosynthetic rate under drought stress is also mentioned in Table 11.
Table 10.
Analysis of variance of stepwise multiple regression of physiological variables under drought stress.
Table 11.
The regression coefficients (b), standard error and R square values of the estimated variables in predicting maize net photosynthetic rate under drought stress by stepwise multiple regression analysis.
4. Discussion
4.1. Morphological and Growth-Related Variables
As one of the abiotic factors, drought limits the production and development of many field crops [50]. Lack of water causes changes in morphology and anatomy, as well as in physio-chemical processes, changing many functions in a plant [51]. Under such conditions, the plant loses its turgidity causing a reduction in the growth of the plant, which is dependent on cell expansion and division and involves intricate interactions among processes of physiology, morphology and genetics [52], elongation of the stem, leaf expansion, and opening of stomata, thus changing the sink–source relationship, which has an impact on the translocation of photosynthates [53]. The plant’s metabolic activity is also limited by water deficiency in soil causing a reduction in biomass and leaf area, and reduced photosynthesis due to the loss of chlorophyll in the leaves, all of which contribute to a low maize yield [54]. The height reduction in plants might be associated with inefficient mitosis, high leaf abscission rate under drought and reduced development of a cell [55]. In the current study, plant height, leaf area, stem diameter, root length, root fresh and dry weight were reduced by drought as shown in Table 1. This stress treatment has more severe negative effects on P-3011w as compared to P-3092 and iku20. Previously, it has been reported that a significant decline occurred in the plant height, leaf area and stem diameter of maize hybrids subjected to drought than control conditions [10,56]. In another study, a significant decrease in the plant height, leaf area, root length, and root fresh and dry weight in maize genotypes under drought stress in comparison to control has been reported [57]. Laskari et al. [54] also showed a significant difference in the height of the maize plants under different irrigation levels with the tallest being observed under normal irrigation and the shortest under reduced irrigation.
4.2. Physiological Variables
4.2.1. Potential Quantum Yield Efficiency of PSII (Fv/Fm)
Drought affected all the physiological parameters of maize such as Fv/Fm, PN, gs, Tr, leaf temperature and intercellular CO2 concentration. Under control conditions, iku20 had the maximum Fv/Fm value (0.79 ± 0.01) followed closely by P-3092 (0.78 ± 0.01) and P-3011w (0.77 ± 0.01), which indicates an undamaged photosystem II and these results are similar to the ones reported earlier [33], where they registered a value range of 0.76–0.80 for Fv/Fm under normal irrigation. Under drought stress, all of our genotypes showed values below 0.75, which is considered a cutoff point for completely functional PSII reaction centers [58]. Changes in stomatal conductance, leaf temperature, chlorophyll and respiration in a plant are caused by drought, and it also causes damage to photosystem II [59]. The reduced values under drought stress show damage to PSII, which is probably the result of damage to thylakoid membranes; increased permeability of the membrane or increased oxidative stress response, which probably led to proton leakage and a drop in the production of ATP and NADPH; and decreased photochemical efficiency [60]. Results found in our study were also reported previously [61], where a significant reduction in Fv/Fm under drought than the control was observed. Other studies also reported a significantly decreased Fv/Fm in severe drought stress conditions [30,62,63].
Based on the assumption that the damage caused by drought stress to photosystem II is less in tolerant genotypes, the presence of significant genotype–treatment interaction allowed us to identify the tolerant genotype, which was iku20.
4.2.2. Photosynthetic Parameters
Certain elements such as leaf water potential, canopy and leaf temperature, transpiration, and stomatal conductance have an influence on plant–water relations. During drought, the stomata are closed reducing transpiration and ultimately impairing the photosynthesis [50]. Drought tolerance and water-use efficiency are used often as synonymous terms. Water usage efficiency is frequently used as a direct indicator of the fundamental physiological response of a plant to water deficiency [48]. In the present study, the genotypes showed a negative response to drought stress for photosynthetic parameters. The drought decreased the stomatal conductance and transpiration to prevent excessive water loss which resulted in an increased leaf temperature and water-use efficiency, and reduced photosynthesis because of less availability of CO2. The intercellular CO2 concentration was increased due to damage caused by drought stress to photosynthetic activity. Temperatures above 35 °C are known to reduce the RuBiscO activity, reducing transpiration and photosynthesis processes [53]. The results recorded in this study are in accordance with several studies conducted in the past. A study conducted on the effects of drought stress during the reproductive stage of maize hybrids found that drought stress reduced stomatal conductance (52%) and transpiration (47%) [41]. Another study observed higher PN, gs and Tr reduction by drought stress compared to control conditions [64]. In a study about drought influence on the physiological parameters of maize, stomatal conductance values of 0.05–0.07 H2O mmolm−2 s−1 have been observed under drought stress due to reduced relative water content, along with a significant increase in the intercellular CO2 concentration compared to control conditions [10]. A significant increase has been recorded in the leaf temperature of maize when subjected to drought stress [63]. Yousaf et al. [65] reported a significant reduction in instantaneous water-use efficiency in maize hybrids under drought stress during the reproductive stage. Salgado-Aguilar et al. [53] observed a significant elevation in the leaf temperature of maize hybrids and even after restoring the irrigation, they did not return to their initial temperature values. Stomatal regulation maintains an ideal internal CO2 concentration level under water availability conditions, enabling the Calvin cycle to proceed. In case of water deficiency, the plant will make a compromise between its need to produce carbohydrates and maintain a proper hydric balance [66,67].
4.2.3. Correlation and Multiple Regression Analyses
In this study, the Pearson correlation analysis of selected morphological and physiological parameters showed significant positive and negative correlations under both water regimes. Intercellular CO2 concentration had a negative significant correlation with Fv/Fm under control conditions because, when the photosystem II is functioning properly, plants are using the intercellular CO2 for photosynthesis, and thus the concentration will be low and vice versa [10]. Under both water regimes, the correlations between PN and gs; PN and Tr and gs and Tr were significantly positive indicating a positive linear relationship among them [68,69]. Additionally, a positive significant correlation was found among Fv/Fm, plant height and stem diameter. These findings were also reported previously [70]. These results therefore demonstrated the importance of these characteristics in choosing drought-tolerant genotypes. When combined, the strong positive and negative correlations between the many variables under study and the water stress further support the use of these correlations to find prospective maize lines that are tolerant to drought. On the other hand, distinct patterns of correlations between the same traits under various water regimes ought to be heavily employed, as certain interrelationships among certain influential parameters under particular water conditions might serve as selection criteria for genotypes that have promising traits responsive to drought. The stepwise multiple regression analysis showed that the maximum variation in net photosynthetic rate under both water regimes was attributed to the Fv/Fm, which indicates the importance of this variable. A proper functioning photosystem II increases the photosynthesis, which will ultimately enhance the overall plant yield [71].
5. Conclusions
In this study, we screened three maize hybrids under different water regimes for various traits. Overall, drought stress brought a remarkable negative impact on morphological and physiological attributes and caused statistically significant damage to photosystem II as well. The genotypes P-3011w, P-3092 and iku20 varied in their responses to drought stress treatment. Overall, the morpho-physiological performance of iku20 was better than that of P-3011w and P-3092. As a result, we conclude that iku20 might have a tendency to tolerate drought stress in the arid northeast of Mexico.
Author Contributions
Conceptualization, F.Z.-G.; Data curation, S.Y.; Formal analysis, S.Y.; Investigation, S.Y.; Project administration, S.Y.; Validation, G.N.-M., P.A.R.-S., A.G.-D., S.R.S.-G. and E.L.-C.; Writing—original draft, S.Y.; Writing—review and editing, F.Z.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by CONAHCYT, as a scholarship awarded to Saba Yasin (CVU: 1193352), for a doctoral program at Facultad de Agronomía, Universidad Autónoma de Nuevo León. The funding source has no involvement in conducting this study or in an article submission.
Data Availability Statement
All the data generated during this study are mentioned in the article in the form of tables and figures.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sahoo, S.; Adhikari, S.; Joshi, A.; Singh, N.K. Use of wild progenitor teosinte in maize (Zea mays subsp. mays) improvement: Present status and future prospects. Trop. Plant Biol. 2021, 14, 156–179. [Google Scholar] [CrossRef]
- USDA. Corn Production. 2022. Available online: https://fas.usda.gov/data/mexico-grain-and-feed-update-22 (accessed on 29 January 2024).
- Ramírez-Vega, H.; Vázquez-Carrillo, G.; Muñóz-Rosales, G.M.; Martínez-Loperena, R.; Heredia-Nava, D.; Martínez-Sifuentes, J.Á.; Anaya-Esparza, L.M.; Gómez-Rodríguez, V.M. Physical and chemical characteristics of native maize from the Jalisco highlands and their influence on the nixtamalization process. Agriculture 2022, 12, 1293. [Google Scholar] [CrossRef]
- Ureta, C.; González, E.J.; Espinosa, A.; Trueba, A.; Piñeyro-Nelson, A.; Álvarez-Buylla, E.R. Maize yield in Mexico under climate change. Agric. Syst. 2020, 177, 102697. [Google Scholar] [CrossRef]
- Revilla, P.; Anibas, C.M.; Tracy, W.F. Sweet corn research around the world 2015–2020. Agronomy 2021, 11, 534. [Google Scholar] [CrossRef]
- Różewicz, M. Production, use and efficiency of utilising grains of various cereal species as feed resources for poultry production. Pol. J. Agron. 2019, 38, 66–74. [Google Scholar]
- Sánchez Hernández, M.Á.; Cruz Vázquez, M.; Sánchez Hernández, C.; Morales Terán, G.; Rivas Jacobo, M.A.; Villanueva Verduzco, C. Forage yield of maize adapted to the humid tropic of Mexico. Rev. Mex. Cienc. Agric. 2019, 10, 699–712. [Google Scholar]
- Ibarrola-Rivas, M.J.; Castillo, G.; González, J. Social, economic and production aspects of maize systems in Mexico. Investig. Geográficas 2020, 102, 1–8. [Google Scholar]
- Safian, N.; Naderi, M.R.; Torabi, M.; Soleymani, A.; Salemi, H.R. Corn (Zea mays L.) and sorghum (Sorghum bicolor (L.) Moench) yield and nutritional quality affected by drought stress. Biocatal. Agric. Biotechnol. 2022, 45, 102486. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef]
- Yasin, M.; Ahmad, A.; Khaliq, T.; Habib-ur-Rahman, M.; Niaz, S.; Gaiser, T.; Ghafoor, I.; Hassan, H.S.U.; Qasim, M.; Hoogenboom, G. Climate change impact uncertainty assessment and adaptations for sustainable maize production using multi-crop and climate models. Environ. Sci. Pollut. Res. 2022, 29, 18967–18988. [Google Scholar] [CrossRef]
- Król-Badziak, A.; Kozyra, J.; Rozakis, S. Assessment of Suitability Area for Maize Production in Poland Related to the Climate Change and Water Stress. Sustainability 2024, 16, 852. [Google Scholar] [CrossRef]
- Harvey, C.A.; Saborio-Rodríguez, M.; Martinez-Rodríguez, M.R.; Viguera, B.; Chain-Guadarrama, A.; Vignola, R.; Alpizar, F. Climate change impacts and adaptation among smallholder farmers in Central America. Agric. Food Secur. 2018, 7, 57. [Google Scholar] [CrossRef]
- Ndlovu Kim, W.; Iizumi, T.; Nishimori, M. Global patterns of crop production losses associated with droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Lukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Jedmowski, C.; Ashoub, A.; Brüggemann, W. Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiol. Plant. 2013, 35, 345–354. [Google Scholar] [CrossRef]
- Liu, Y.; Yue, L.; Wang, C.; Zhu, X.; Wang, Z.; Xing, B. Photosynthetic response mechanisms in typical C3 and C4 plants upon La 2 O 3 nanoparticle exposure. Environ. Sci. Nano 2020, 7, 81–92. [Google Scholar] [CrossRef]
- Foyer, C.H.; Harbinson, J. Oxygen metabolism and the regulation of photosynthetic electron transport. In Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants, 1st ed.; Foyer, C.H., Mullineaux, P.M., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–42. [Google Scholar]
- Różewicz, M.; Grabiński, J.; Wyzińska, M. Effect of strip-till and cultivar on photosynthetic parameters and grain yield of winter wheat. Int. Agrophys. 2024, 38, 279–291. [Google Scholar] [CrossRef]
- Stępień-Warda, A. Effect of soil cultivation system on the efficiency of the photosynthetic apparatus in maize leaves (Zea mays L.). Pol. J. Agron. 2020, 43, 57–62. [Google Scholar]
- Ndlovu, E.; Van Staden, J.; Maphosa, M. Morpho-physiological effects of moisture, heat and combined stresses on Sorghum bicolor [Moench (L.)] and its acclimation mechanisms. Plant Stress 2021, 2, 100018. [Google Scholar] [CrossRef]
- Liu, J.; Guo, Y.; Bai, Y.W.; Camberato, J.J.; Xue, J.Q.; Zhang, R.H. Effects of drought stress on the photosynthesis in maize. Russ. J. Plant Physl. 2018, 65, 849–856. [Google Scholar] [CrossRef]
- Wang, B.; Liu, C.; Zhang, D.; He, C.; Zhang, J.; Li, Z. Effects of maize organ-specific drought stress response on yields from transcriptome analysis. BMC Plant Biol. 2019, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, Y.; Huang, S.; Xu, C.; Wang, X.; Gao, J.; Meng, Q.; Wang, P. The impact of drought and heat stress at flowering on maize kernel filling: Insights from the field and laboratory. Agric. For. Meteorol. 2022, 312, 108733. [Google Scholar] [CrossRef]
- Basal, O.; Szabó, A.; Veres, S. Physiology of soybean as affected by PEG-induced drought stress. Curr. Plant Biol. 2020, 22, 100135. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Singh, A.K.; Ansari, M.I. Effect of drought stress on crop production. In New Frontiers in Stress Management for Durable Agriculture; Rakshit, A., Singh, H.B., Singh, A.K., Singh, U.S., Fraceto, L., Eds.; Springer: Singapore, 2020; pp. 35–47. [Google Scholar]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Lee, J.G. Effect of drought stress on chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities in lettuce seedlings. Horticulturae 2021, 7, 238. [Google Scholar] [CrossRef]
- Badr, A.; Brüggemann, W. Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica 2020, 58, 38–645. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Krause, G.H. Photoinhibition of photosynthesis. An evaluation of damaging and protective mechanisms. Physiol. Plant. 1988, 74, 566–574. [Google Scholar] [CrossRef]
- Krause, A.G.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Öquist, G.; Chow, W.S.; Anderson, J.M. Photoinhibition of photosynthesis represents a mechanism for the long-term regulation of photosystem II. Planta 1992, 186, 450–460. [Google Scholar] [CrossRef]
- Mehta, P.; Allakhverdiev, S.I.; Jajoo, A. Characterization of photosystem II heterogeneity in response to high salt stress in wheat leaves (Triticum aestivum). Photosyn. Res. 2010, 105, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Zushi, K.; Matsuzoe, N. Using of chlorophyll a fluorescence OJIP transients for sensing salt stress in the leaves and fruits of tomato. Sci. Hortic. 2017, 219, 216–221. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Malnoë, A. Photoinhibition or photoprotection of photosynthesis? Update on the (newly termed) sustained quenching component qH. Environ. Exp. Bot. 2018, 154, 123–133. [Google Scholar] [CrossRef]
- He, W.; Yoo, G.; Ryu, Y. Evaluation of effective quantum yields of photosystem II for CO2 leakage monitoring in carbon capture and storage sites. PeerJ 2021, 9, e10652. [Google Scholar] [CrossRef] [PubMed]
- Bheemanahalli, R.; Ramamoorthy, P.; Poudel, S.; Samiappan, S.; Wijewardane, N.; Reddy, K.R. Effects of drought and heat stresses during reproductive stage on pollen germination, yield, and leaf reflectance properties in maize (Zea mays L.). Plant Direct 2022, 6, e434. [Google Scholar] [CrossRef]
- Imakumbili, M.L. Making Water Stress Treatments in Pot Experiments: An Illustrated Step-by-Step Guide; Sokoine University of Agriculture: Morogoro, Tanzania, 2019; pp. 1–17. [Google Scholar]
- Montgomery, E. Correlation studies in corn. Neb. Agric. Exp. Stn. Annu. Rep 1911, 24, 108–159. [Google Scholar]
- Zhang, Y.; Liu, P.; Zhang, X.; Zheng, Q.; Chen, M.; Ge, F.; Li, Z.; Sun, W.; Guan, Z.; Liang, T.; et al. Multi-locus genome-wide association study reveals the genetic architecture of stalk lodging resistance-related traits in maize. Front. Plant Sci. 2018, 9, 611. [Google Scholar] [CrossRef]
- Fang, Y.; Xu, B.C.; Turner, N.C.; Li, F.M. Grain yield, dry matter accumulation and remobilization, and root respiration in winter wheat as affected by seeding rate and root pruning. Eur. J. Agron. 2010, 33, 257–266. [Google Scholar] [CrossRef]
- Galicia-Juárez, M.; Zavala-García, F.; Sinagawa-García, S.R.; Gutiérrez-Diez, A.; Williams-Alanís, H.; Cisneros-López, M.E.; Valle-Gough, R.E.; Flores-Garivay, R.; Santillano-Cázares, J. Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico. Plants 2021, 10, 2265. [Google Scholar] [CrossRef] [PubMed]
- Padhi, B.; Chauhan, G.; Kandoi, D.; Stirbet, A.; Tripathy, B.C.; Govindjee, G. A comparison of chlorophyll fluorescence transient measurements, using Handy PEA and FluorPen fluorometers. Photosynthetica 2021, 59, 399–408. [Google Scholar] [CrossRef]
- Hasan, S.A.; Rabei, S.H.; Nada, R.M.; Abogadallah, G.M. Water use efficiency in the drought-stressed sorghum and maize in relation to expression of aquaporin genes. Biol. Plant. 2017, 61, 127–137. [Google Scholar] [CrossRef]
- Mukaka, M.M. A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar] [PubMed]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.J.; Reddy, V.R.; Timlin, D. Drought-induced responses in maize under different vapor pressure deficit conditions. Plants 2022, 11, 2771. [Google Scholar] [CrossRef] [PubMed]
- Wach, D.; Skowron, P. An overview of plant responses to the drought stress at morphological, physiological and biochemical levels. Pol. J. Agron. 2022, 50, 25–34. [Google Scholar]
- Salgado-Aguilar, M.; Molnar, T.; Pons-Hernández, J.L.; Covarrubias-Prieto, J.; Ramírez-Pimentel, J.G.; Raya-Pérez, J.C.; Hearne, S.; Iturriaga, G. Physiological and biochemical analyses of novel drought-tolerant maize lines reveal osmoprotectant accumulation at silking stage. Chil. J. Agric. Res. 2020, 80, 241–252. [Google Scholar] [CrossRef]
- Laskari, M.; Menexes, G.; Kalfas, I.; Gatzolis, I.; Dordas, C. Water stress effects on the morphological, physiological characteristics of maize (Zea mays L.), and on environmental cost. Agronomy 2022, 12, 2386. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ physio-biochemical and phyto-hormonal responses to alleviate the adverse effects of drought stress: A comprehensive review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef] [PubMed]
- Wada, S.; Takagi, D.; Miyake, C.; Makino, A.; Suzuki, Y. Responses of the photosynthetic electron transport reactions stimulate the oxidation of the reaction center chlorophyll of photosystem I, P700, under drought and high temperatures in rice. Int. J. Mol. Sci. 2019, 20, 2068. [Google Scholar] [CrossRef] [PubMed]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of pre-anthesis drought, heat and their combination on the growth, yield and physiology of diverse wheat (Triticum aestivum L.) genotypes varying in sensitivity to heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [PubMed]
- Kohzuma, K.; Cruz, J.A.; Akashi, K.; Hoshiyasu, S.; Munekage, Y.N.; Yokota, A.; Kramer, D.M. The long-term responses of the photosynthetic proton circuit to drought. Plant Cell Environ. 2009, 32, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Chiango, H.; Figueiredo, A.; Sousa, L.; Sinclair, T.; da Silva, J.M. Assessing drought tolerance of traditional maize genotypes of Mozambique using chlorophyll fluorescence parameters. S. Afr. J. Bot. 2021, 138, 311–317. [Google Scholar] [CrossRef]
- Liu, M.; Qi, H.; Zhang, Z.P.; Song, Z.W.; Kou, T.J.; Zhang, W.J.; Yu, J.L. Response of photosynthesis and chlorophyll fluorescence to drought stress in two maize cultivars. Afr. J. Agric. Res. 2012, 7, 4751–4760. [Google Scholar]
- Ghassemi-Golezani, K.; Heydari, S.; Dalil, B. Field performance of maize (Zea mays L.) cultivars under drought stress. Acta Agric. Slov. 2018, 111, 25–32. [Google Scholar] [CrossRef][Green Version]
- Cai, F.; Zhang, Y.; Mi, N.; Ming, H.; Zhang, S.; Zhang, H.; Zhao, X. Maize (Zea mays L.) physiological responses to drought and rewatering, and the associations with water stress degree. Agric. Water Manag. 2020, 241, 106379. [Google Scholar] [CrossRef]
- Yousaf, M.I.; Riaz, M.W.; Shehzad, A.; Jamil, S.; Shahzad, R.; Kanwal, S.; Ghani, A.; Ali, F.; Abdullah, M.; Ashfaq, M.; et al. Responses of maize hybrids to water stress conditions at different developmental stages: Accumulation of reactive oxygen species, activity of enzymatic antioxidants and degradation in kernel quality traits. PeerJ 2023, 11, e14983. [Google Scholar] [CrossRef]
- Cornic, G. Drought stress inhibits photosynthesis by decreasing stomatal aperture–not by affecting ATP synthesis. Trends Plant Sci. 2000, 5, 187–188. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed]
- Miner, G.L.; Bauerle, W.L. Seasonal variability of the parameters of the Ball–Berry model of stomatal conductance in maize (Zea mays L.) and sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Plant Cell Environ. 2017, 40, 1874–1886. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Han, Z.; Feng, H.; Wang, Y.; Kang, J.; Han, X.; Wang, L.; Wang, C.; Li, H.; et al. Analysis of physiological indicators associated with drought tolerance in wheat under drought and re-watering conditions. Antioxidants 2022, 11, 2266. [Google Scholar] [CrossRef] [PubMed]
- Holá, D.; Benešová, M.; Honnerová, J.; Hnilička, F.; Rothová, O.; Kočová, M.; Hniličková, H. The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 2010, 48, 545–558. [Google Scholar] [CrossRef]
- Djemel, A.; Álvarez-Iglesias, L.; Santiago, R.; Malvar, R.A.; Pedrol, N.; Revilla, P. Algerian maize populations from the Sahara desert as potential sources of drought tolerance. Acta Physiol. Plant. 2019, 41, 12. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).