
Vegetation’s Dynamic Changes, Spatial Trends, and Responses to Drought in the Yellow River Basin, China

Abstract
:1. Introduction
2. Study Area and Dataset
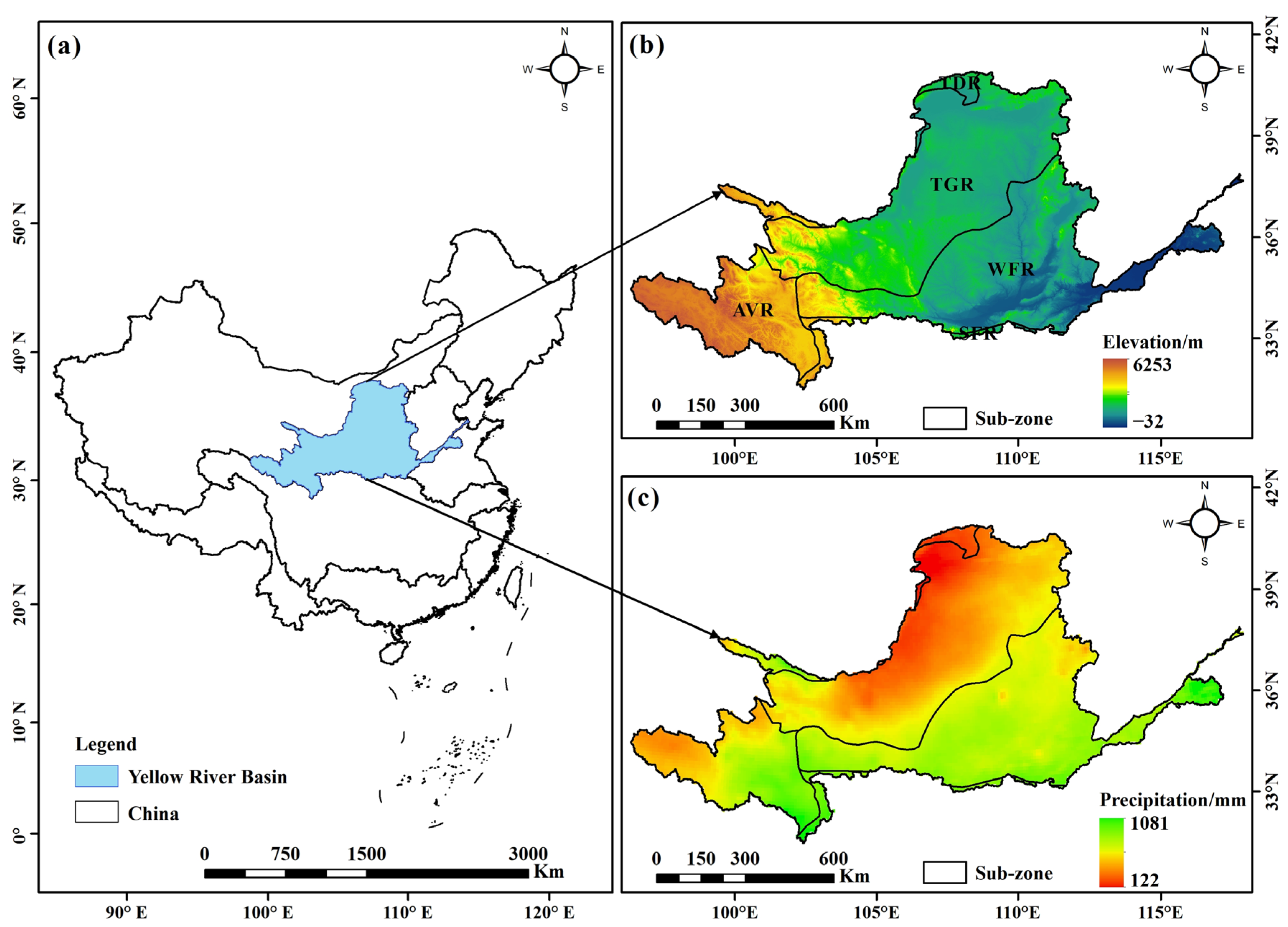
2.1. Study Area Description
2.2. Dataset
3. Methodology
3.1. Construction of Standardized Drought Index
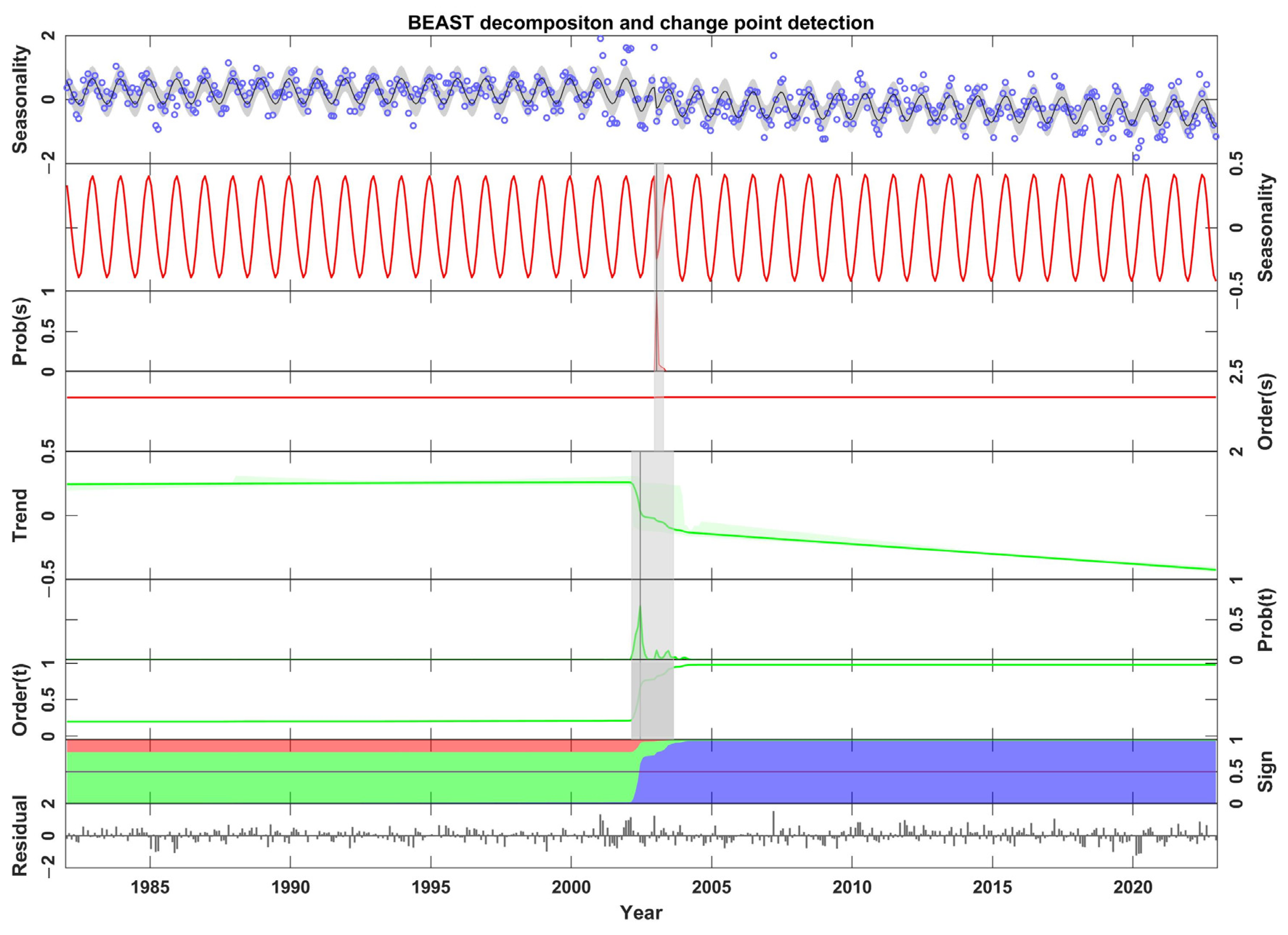
3.2. Bayesian Estimator of Abrupt Seasonal and Trend Change Algorithm (BEAST)
3.3. Spatial Trend Identification Method
3.4. Cross-Wavelet Transform Technology
4. Results
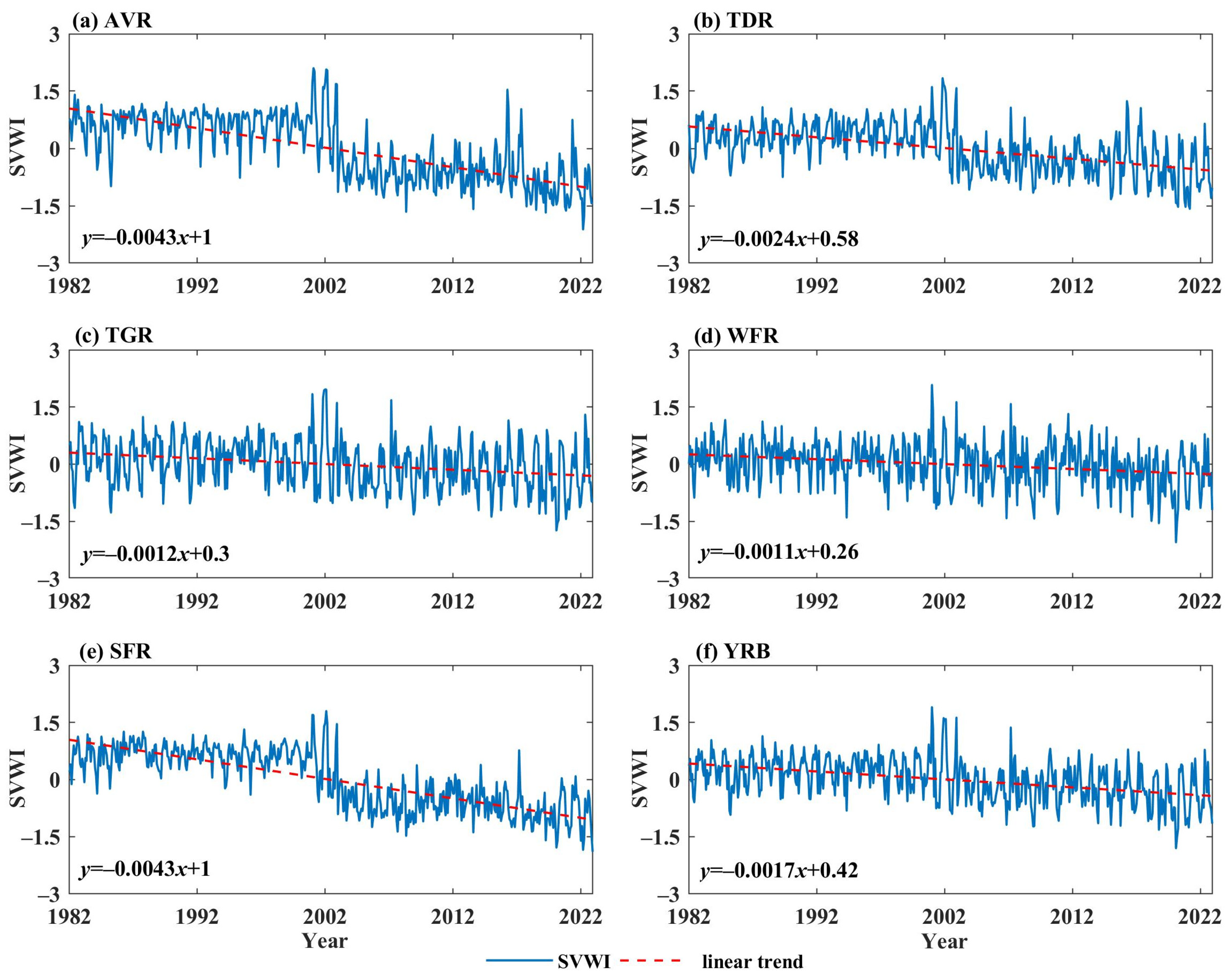
4.1. Changes in Vegetation Dynamics
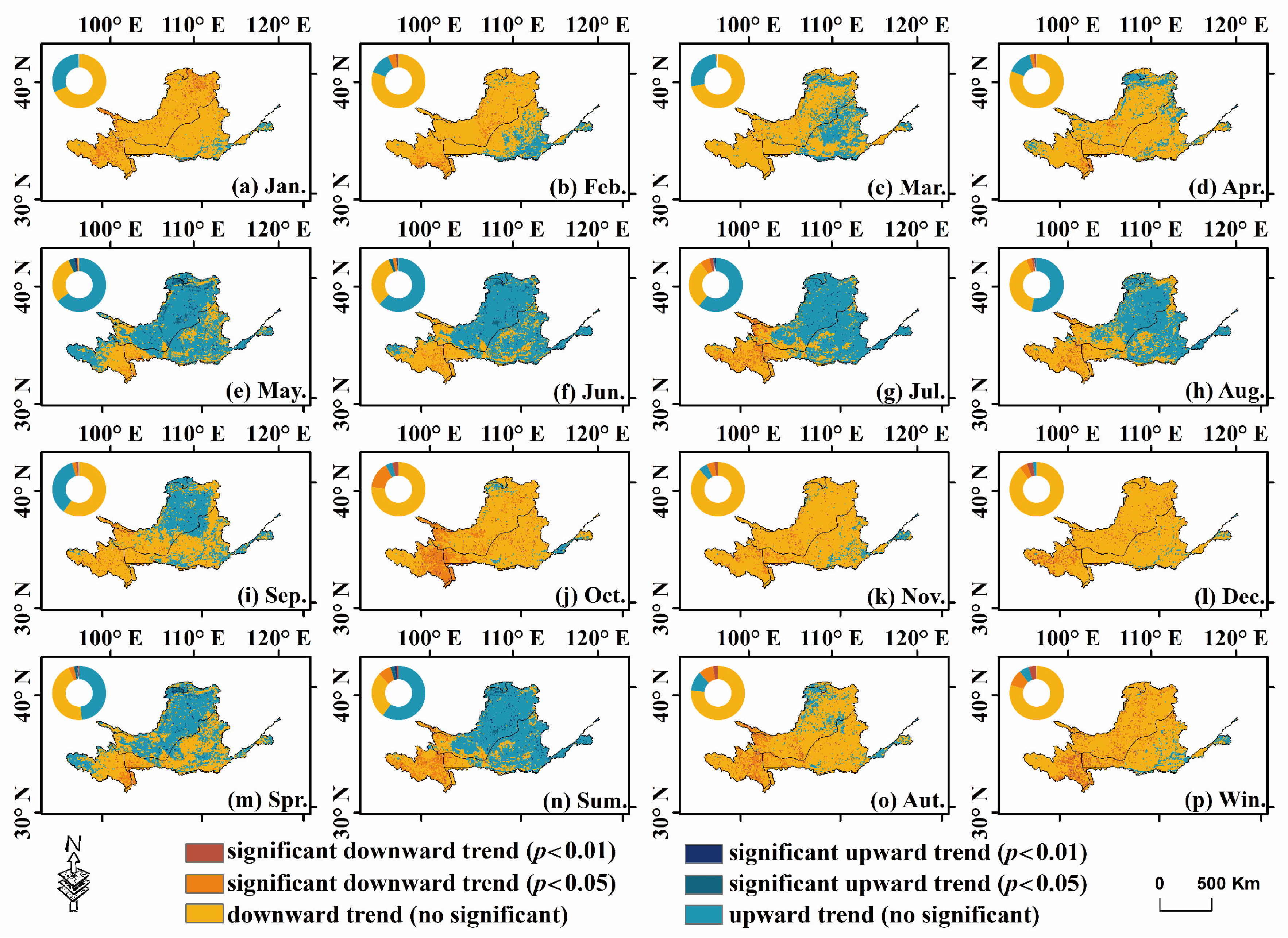
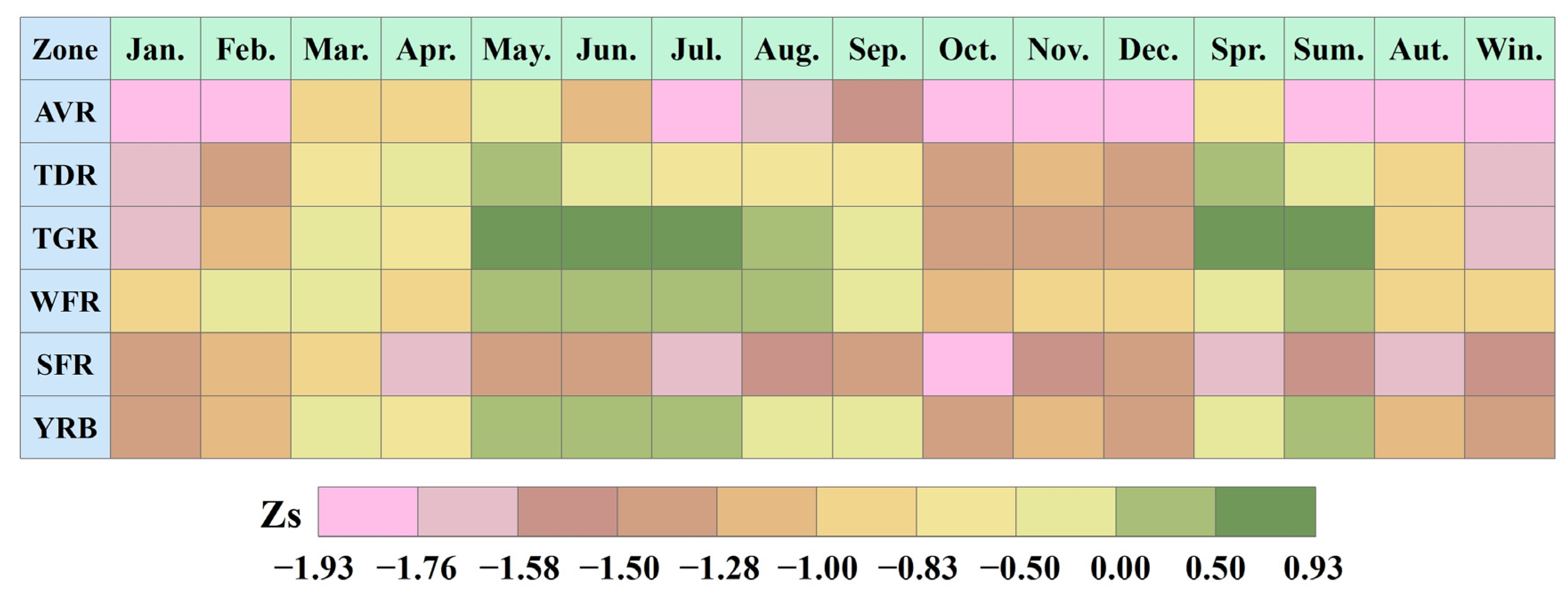
4.2. Spatial Trend Variations of Vegetation
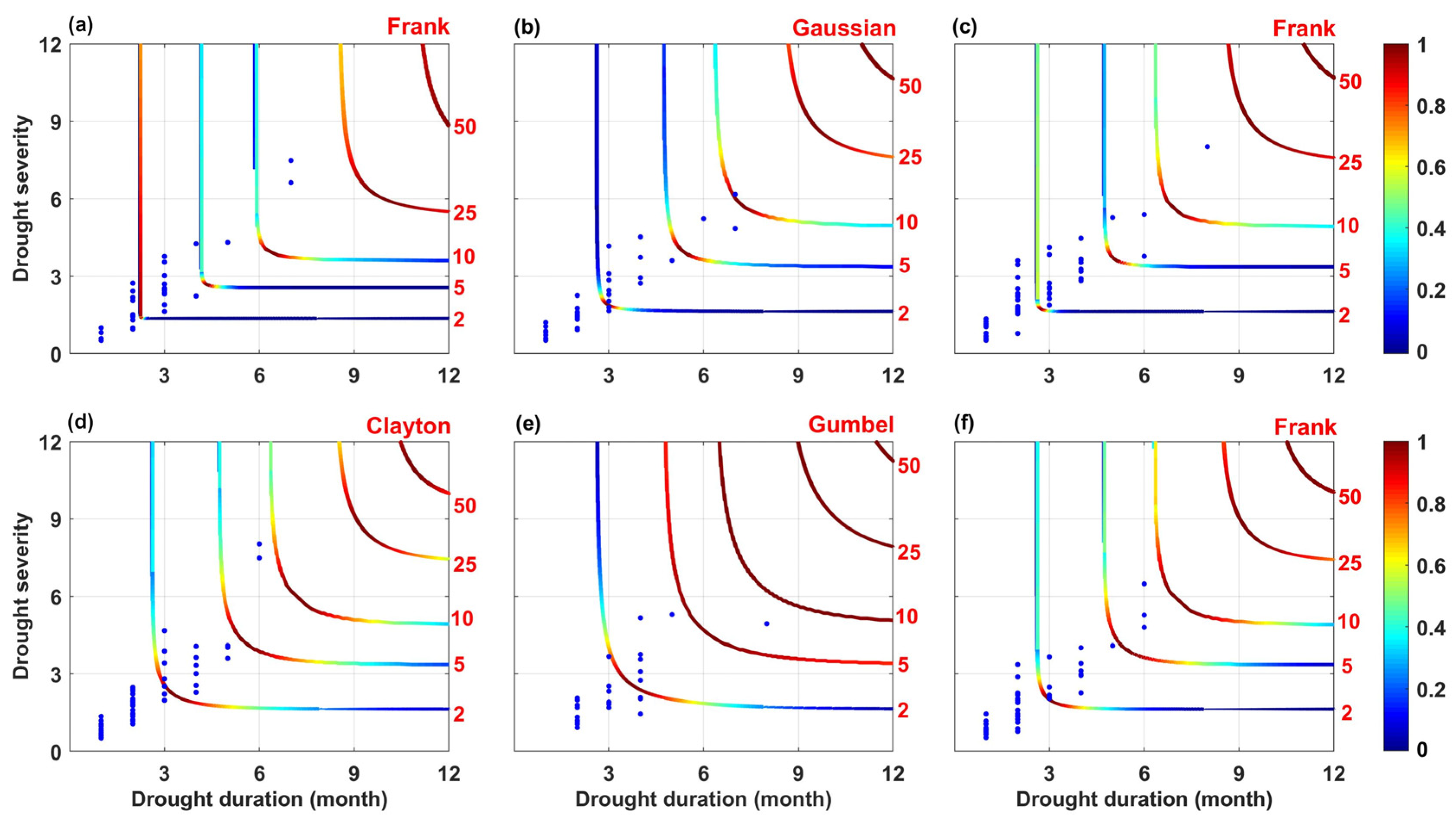
4.3. The Spatial Distribution and Recurrence Period of Drought
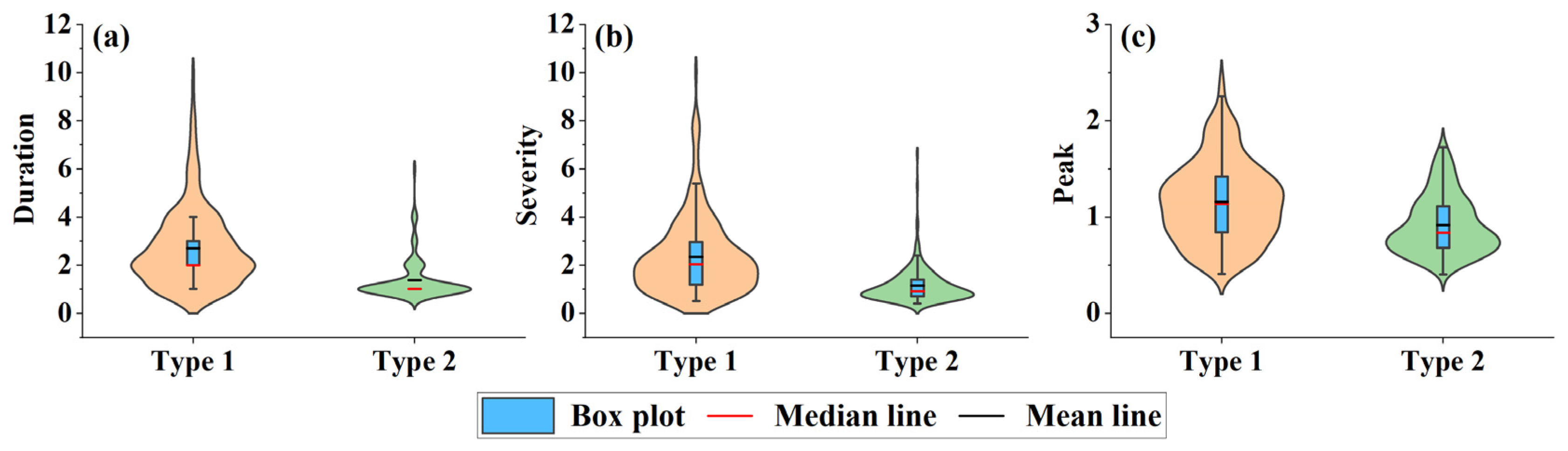
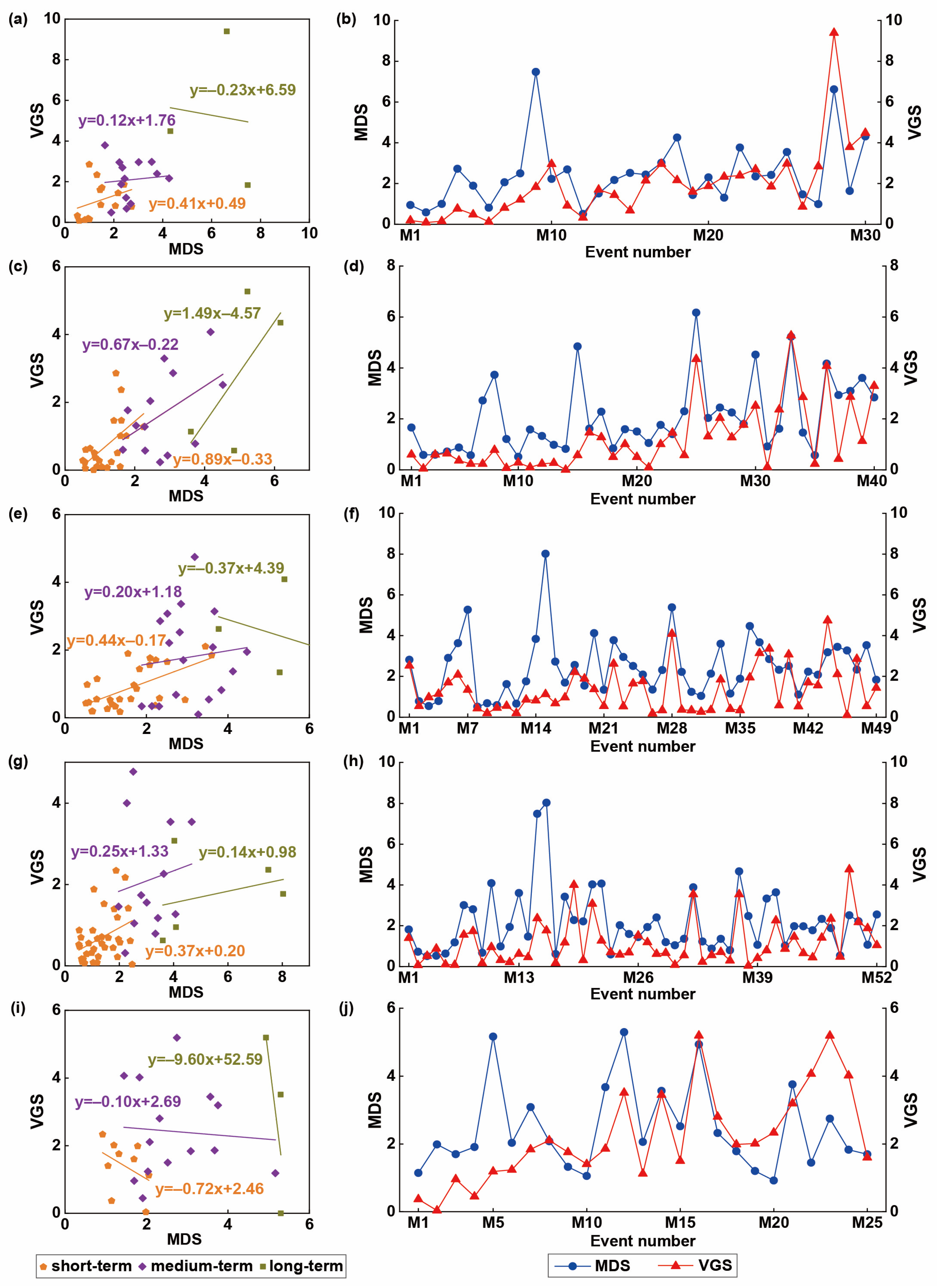
4.4. The Impacts of Drought Events on Vegetation Growth
5. Discussion
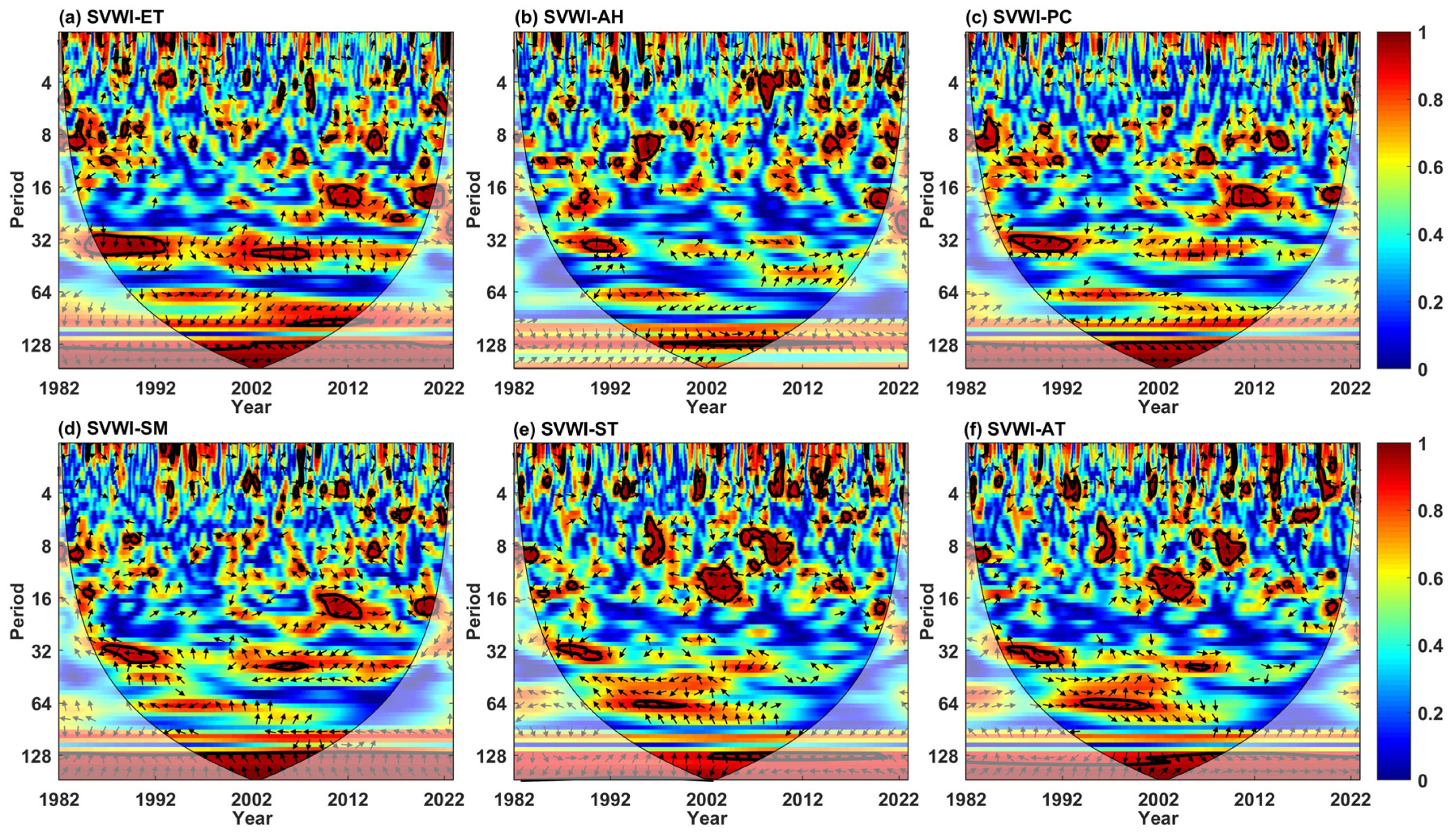
5.1. Driving Factor Identification
5.1.1. Univariate Influencing Factors
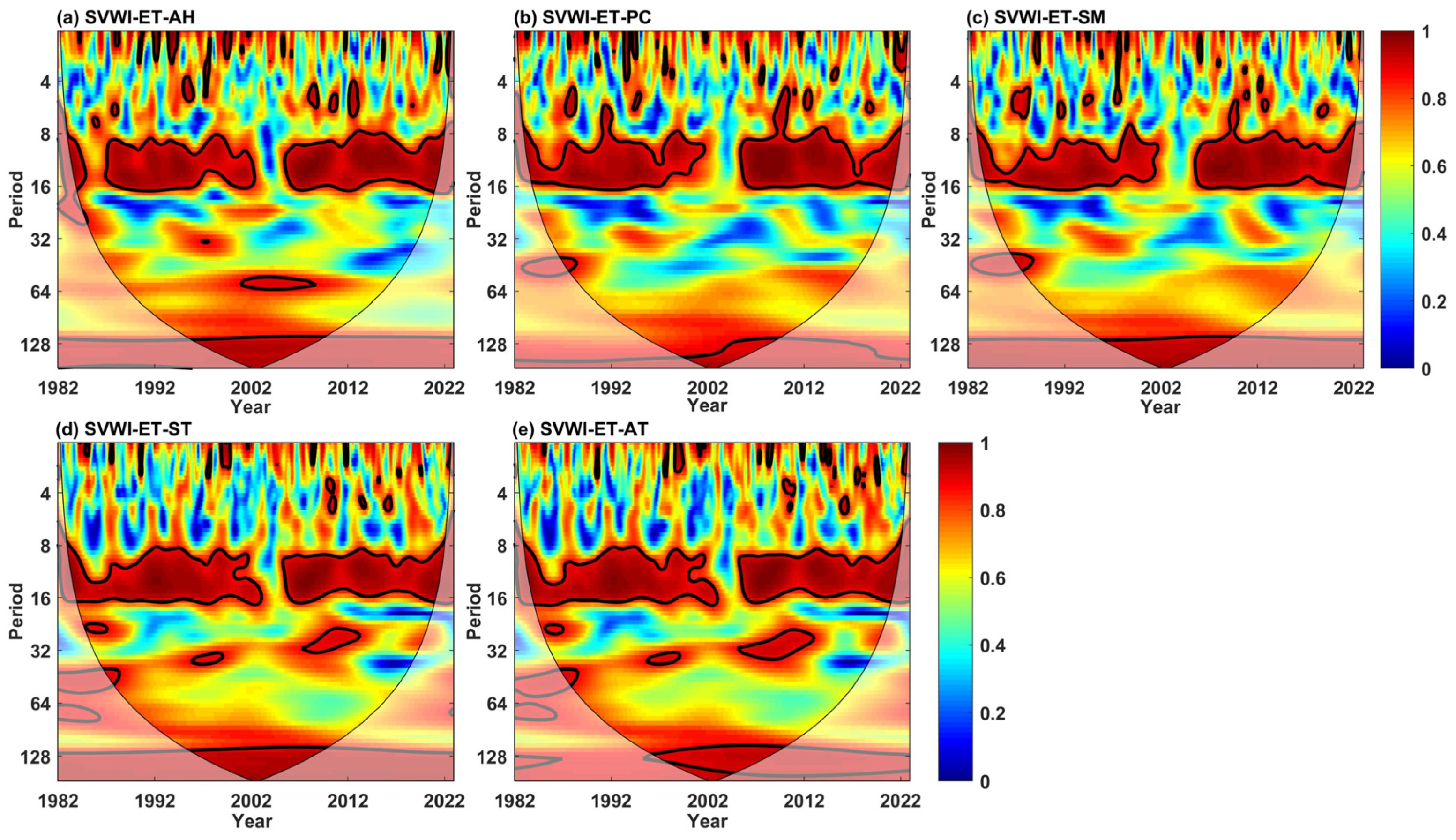
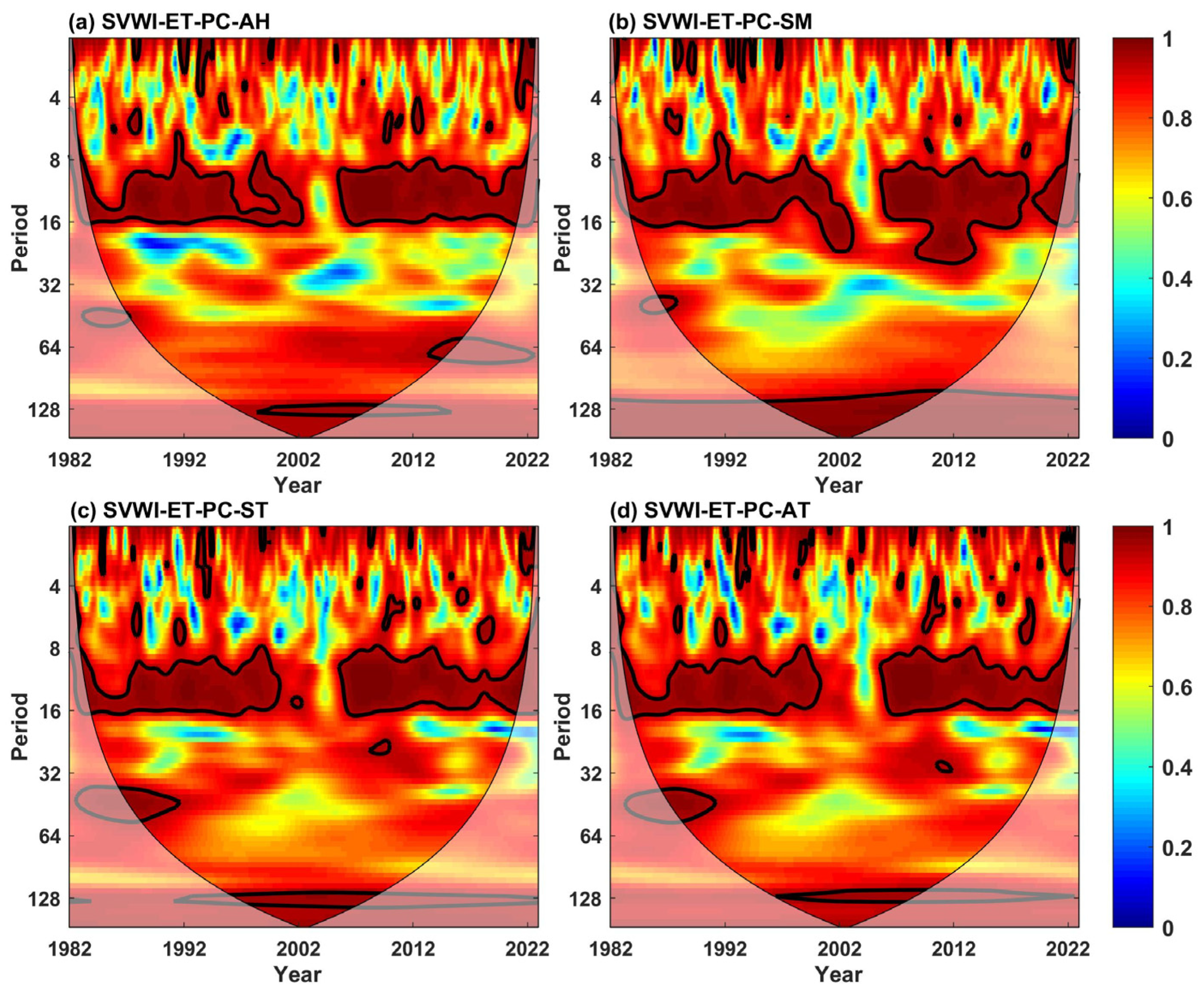
5.1.2. Multivariate Influencing Factors
5.2. Advantages and Limitations
5.3. Future Prospects
6. Conclusions
- (1)
- From 1982 to 2022, the SVWI showed a decreasing trend in the YRB, but the magnitude of the decline varied across different sub-regions. At the decadal scale, the SVWI exhibited a gradual decline in the 1990s and 2010s. Based on the BEAST algorithm, the seasonal change point of the SVWI occurred with a probability of 99.52% in January 2003, while the trend change point had a probability of 72.15% in June 2002.
- (2)
- Using the spatial trend identification method, at the grid scale, the SVWI generally increased from May to July and decreased in the other months. Additionally, the minimum value of the trend characteristic Zs, −1.45, occurred in October. Across the different quarters, the range of mean Zs values was −1.46 (winter) to 0.20 (summer), indicating a more pronounced declining trend in winter, with an area percentage reaching 94.26%.
- (3)
- The most severe drought occurred in 1997 within the study period, with an SPEI value of −1.09. In October of that year, the SPEI reached its minimum value of −1.44. Furthermore, the optimal copula function connecting drought variables in the AVR, TGR, and YRB regions was the Frank copula. In the entire basin, the duration of the drought that occurred from September 1998 to February 1999 was 6 months, with a severity of 6.48 and a return period of 8.54 years.
- (4)
- Overall, there was a close relationship between drought and vegetation growth in the YRB. The maximum number of drought events (52 times) occurred in the WFR, while the minimum number of drought events (25 times) occurred in the SFR. Particularly, the MDS of the short-term, medium-term, and long-term drought events in the AVR was 32.21%, 28.97%, and 17.17% higher than the mean of VGS.
- (5)
- The accuracy of the vegetation water consumption and demand used to calculate the degree of vegetation water shortage may introduce some uncertainty into the results. In the future, we will adopt higher-resolution land-use data to accurately obtain the vegetation water consumption and demand, which will also be our main research focus in the next work.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, M.C.; Zolin, C.A.; Sentelhas, P.C.; Hain, C.R.; Semmens, K.; Tugrul Yilmaz, M.; Gao, F.; Otkin, J.A.; Tetrault, R. The Evaporative Stress Index as an indicator of agricultural drought in Brazil: An assessment based on crop yield impacts. Remote Sens. Environ. 2016, 174, 82–99. [Google Scholar] [CrossRef]
- Bradford, J.B.; Schlaepfer, D.R.; Lauenroth, W.K.; Palmquist, K.A. Robust ecological drought projections for drylands in the 21st century. Glob. Change Biol. 2020, 26, 3906–3919. [Google Scholar] [CrossRef]
- Su, B.D.; Huang, J.L.; Mondal, S.K.; Zhai, J.Q.; Wang, Y.J.; Wen, S.S.; Gao, M.N.; Lv, Y.R.; Jiang, S.; Li, A.W. Insight from CMIP6 SSP-RCP scenarios for future drought characteristics in China. Atmos. Res. 2021, 250, 105375. [Google Scholar] [CrossRef]
- Zhao, S.D.; Jiang, Y.; Wen, Y.; Jiao, L.; Li, W.Q.; Xu, H.; Ding, M.H. Frequent locally absent rings indicate increased threats of extreme droughts to semi-arid Pinus tabuliformis forests in North China. Agric. For. Meteorol. 2021, 308–309, 108601. [Google Scholar] [CrossRef]
- Ding, Y.J.; Zhang, L.F.; He, Y.; Cao, S.P.; Wei, X.; Guo, Y.; Ran, L.; Filonchyk, M. Spatiotemporal evolution of agricultural drought and its attribution under different climate zones and vegetation types in the Yellow River Basin of China. Sci. Total Environ. 2024, 914, 169687. [Google Scholar] [CrossRef]
- Otkin, J.A.; Anderson, M.C.; Hain, C.; Svoboda, M.; Johnson, D.; Mueller, R.; Tadesse, T.; Wardlow, B.; Brown, J. Assessing the evolution of soil moisture and vegetation conditions during the 2012 United States flash drought. Agric. For. Meteorol. 2016, 218–219, 230–242. [Google Scholar] [CrossRef]
- Kang, W.P.; Wang, T.; Liu, S.L. The response of vegetation phenology and productivity to drought in semi-arid regions of northern China. Remote Sens. 2018, 10, 727. [Google Scholar] [CrossRef]
- Khatri-Chhetri, P.; Hendryx, S.M.; Hartfield, K.A.; Crimmins, M.A.; Leeuwen, W.J.D.V.; Kane, V.R. Assessing vegetation response to multi-scalar drought across the mojave, sonoran, chihuahuan deserts and apache highlands in the Southwest United States. Remote Sens. 2021, 13, 1103. [Google Scholar] [CrossRef]
- Huete, A. Ecology: Vegetation’s responses to climate variability. Nature 2016, 531, 181–182. [Google Scholar] [CrossRef]
- Pei, F.S.; Wu, C.J.; Liu, X.P.; Li, X.; Yang, K.Q.; Zhou, Y.; Wang, K.; Xu, L.; Xia, G.R. Monitoring the vegetation activity in China using vegetation health indices. Agric. For. Meteorol. 2018, 248, 215–227. [Google Scholar] [CrossRef]
- Chen, Q.; Timmermans, J.; Wen, W.; van Bodegom, P.M. Ecosystems threatened by intensified drought with divergent vulnerability. Remote Sens. Environ. 2023, 289, 113512. [Google Scholar] [CrossRef]
- Zhong, S.B.; Sun, Z.H.; Di, L.P. Characteristics of vegetation response to drought in the CONUS based on long-term remote sensing and meteorological data. Ecol. Indic. 2021, 127, 107767. [Google Scholar] [CrossRef]
- Weng, Z.; Niu, J.; Guan, H.D.; Kang, S.Z. Three-dimensional linkage between meteorological drought and vegetation drought across China. Sci. Total Environ. 2023, 859, 160300. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2012, 110, 52–57. [Google Scholar] [CrossRef]
- Zhao, X.; Wei, H.; Liang, S.L.; Zhou, T.; He, B.; Tang, B.; Wu, D.H. Responses of natural vegetation to different stages of extreme drought during 2009–2010 in southwestern China. Remote Sens. 2015, 7, 14039–14054. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Zhao, C.Y.; Yang, X.M. Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric. For. Meteorol. 2018, 262, 1–13. [Google Scholar] [CrossRef]
- Geldenhuys, C.; van der Merwe, H.; van Rooyen, M.W. Vegetation response to grazing and drought (13 yr) in a conservation area in the Succulent Karoo, South Africa. J. Arid Environ. 2023, 219, 105093. [Google Scholar] [CrossRef]
- Kogan, F. World droughts in the new millennium from AVHRR-based vegetation health indices. Eos Trans. Am. Geophys. Union. 2011, 83, 557–563. [Google Scholar] [CrossRef]
- Prăvălie, R.; Sîrodoev, I.; Nita, I.-A.; Patriche, C.; Dumitraşcu, M.; Roşca, B.; Tişcovschi, A.; Bandoc, G.; Săvulescu, I.; Mănoiu, V.; et al. NDVI-based ecological dynamics of forest vegetation and its relationship to climate change in Romania during 1987–2018. Ecol. Indic. 2022, 136, 108629. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Abubakar, A.; Ishak, M.Y.; Danhassan, S.S.; Zhang, J.H.; Alatalo, J.M. Modeling the influence of daily temperature and precipitation extreme indices on vegetation dynamics in Katsina State using statistical downscaling model (SDM). Ecol. Indic. 2023, 155, 110979. [Google Scholar] [CrossRef]
- Chi, D.K.; Wang, H.; Li, X.B.; Liu, H.H.; Li, X.H. Estimation of the ecological water requirement for natural vegetation in the Ergune River basin in Northeastern China from 2001 to 2014. Ecol. Indic. 2018, 92, 141–150. [Google Scholar] [CrossRef]
- Won, J.; Seo, J.; Lee, J.; Lee, O.; Kim, S. Vegetation drought vulnerability mapping using a copula model of vegetation index and meteorological drought index. Remote Sens. 2021, 13, 5103. [Google Scholar] [CrossRef]
- Cai, Y.T.; Liu, S.T.; Lin, H. Monitoring the vegetation dynamics in the Dongting lake wetland from 2000 to 2019 using the BEAST algorithm based on dense Landsat time series. Appl. Sci. 2020, 10, 4209. [Google Scholar] [CrossRef]
- White, J.H.R.; Walsh, J.E.; Thoman Jr, R.L. Using Bayesian statistics to detect trends in Alaskan precipitation. Int. J. Climatol. 2021, 41, 2045–2059. [Google Scholar] [CrossRef]
- Quiring, S.M.; Ganesh, S. Evaluating the utility of the Vegetation Condition Index (VCI) for monitoring meteorological drought in Texas. Agric. For. Meteorol. 2010, 150, 330–339. [Google Scholar] [CrossRef]
- Sheffield, J.; Wood, E.F.; Roderick, M.L. Little change in global drought over the past 60 years. Nature 2012, 491, 435–438. [Google Scholar] [CrossRef]
- Zeng, J.; Zhou, T.; Qu, Y.; Bento, V.A.; Qi, J.; Xu, Y.; Li, Y.; Wang, Q. An improved global vegetation health index dataset in detecting vegetation drought. Sci. Data. 2023, 10, 338. [Google Scholar] [CrossRef]
- Swann, A.L.; Hoffman, F.M.; Koven, C.D.; Randerson, J.T. Plant responses to increasing CO2 reduce estimates of climate impacts on drought severity. Proc. Natl. Acad. Sci. USA 2016, 113, 10019–10024. [Google Scholar] [CrossRef]
- Zhang, P.; Jeong, J.H.; Yoon, J.H.; Kim, H.; Wang, S.Y.S.; Linderholm, H.W.; Fang, K.; Wu, X.; Chen, D. Abrupt shift to hotter and drier climate over inner East Asia beyond the tipping point. Science 2020, 370, 1095–1099. [Google Scholar] [CrossRef]
- Wang, F.; Lai, H.X.; Li, Y.B.; Feng, K.; Zhang, Z.Z.; Tian, Q.Q.; Zhu, X.M.; Yang, H.B. Dynamic variation of meteorological drought and its relationships with agricultural drought across China. Agr. Water Manag. 2022, 261, 107301. [Google Scholar] [CrossRef]
- Wang, F.; Lai, H.X.; Li, Y.B.; Feng, K.; Tian, Q.Q.; Guo, W.X.; Qu, Y.P.; Yang, H.B. Spatio-temporal evolution and teleconnection factor analysis of groundwater drought based on the GRACE mascon model in the Yellow River Basin. J. Hydrol. 2023, 626, 130349. [Google Scholar] [CrossRef]
- Jiang, T.L.; Su, X.L.; Qu, Y.P.; Singh, V.P.; Zhang, T.; Chu, J.D.; Hu, X.X. Determining the response of ecological drought to meteorological and groundwater droughts in Northwest China using a spatio-temporal matching method. J. Hydrol. 2024, 633, 130753. [Google Scholar] [CrossRef]
- Mao, J.F.; Shi, X.Y.; Thornton, P.E.; Hoffman, F.M.; Zhu, Z.C.; Myneni, R.B. Global latitudinal-asymmetric vegetation growth trends and their driving mechanisms: 1982–2009. Remote Sens. 2013, 5, 1484–1497. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Yang, T.B. Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Remote Sens. Environ. 2017, 579, 1658–1674. [Google Scholar] [CrossRef]
- Tietjen, B.; Schlaepfer, D.R.; Bradford, J.B.; Lauenroth, W.K.; Hall, S.A.; Duniway, M.C.; Hochstrasser, T.; Jia, G.; Munson, S.M.; Pyke, D.A.; et al. Climate change-induced vegetation shifts lead to more ecological droughts despite projected rainfall increases in many global temperate drylands. Global Change Biol. 2017, 23, 2743–2754. [Google Scholar] [CrossRef]
- Du, J.; He, Z.B.; Piatek, K.B.; Chen, L.F.; Lin, P.F.; Zhu, X. Interacting effects of temperature and precipitation on climatic sensitivity of spring vegetation green-up in arid mountains of China. Agr. Forest Meteorol. 2019, 269–270, 71–77. [Google Scholar] [CrossRef]
- Chu, H.S.; Venevsky, S.; Wu, C.; Wang, M.H. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef]
- Nzabarinda, V.; Bao, A.; Xu, W.; Uwamahoro, S.; Xiaoran, H.; Habiyakare, T.; Sindikubwabo, C.; Habumugisha, J.M.; Itangishaka, A.C. A simple model to predict the spatiotemporally vegetation dynamics in terms of precipitation and temperature. Environ. Dev. 2022, 44, 100769. [Google Scholar] [CrossRef]
- Nanzad, L.; Zhang, J.; Tuvdendorj, B.; Nabil, M.; Zhang, S.; Bai, Y. NDVI anomaly for drought monitoring and its correlation with climate factors over Mongolia from 2000 to 2016. J. Arid Environ. 2019, 164, 69–77. [Google Scholar] [CrossRef]
- Zhan, C.; Liang, C.; Zhao, L.; Jiang, S.Z.; Niu, K.J.; Zhang, Y.L. Drought-related cumulative and time-lag effects on vegetation dynamics across the Yellow River Basin, China. Ecol. Indic. 2022, 143, 109409. [Google Scholar] [CrossRef]
- Niwa, H.; Kamada, M.; Morisada, S.; Ogawa, M. Assessing the impact of storm surge flooding on coastal pine forests using a vegetation index. Landsc. Ecol. Eng. 2023, 19, 151–159. [Google Scholar] [CrossRef]
- Jiang, T.L.; Su, X.L.; Singh, V.P.; Zhang, G.X. A novel index for ecological drought monitoring based on ecological water deficit. Ecol. Indic. 2021, 129, 107804. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.Z.; Li, J.; Yang, X.J.; Zhang, N.; Chen, H. Diurnal Variations in Summer Precipitation over the Yellow River Basin. Adv. Meteorol. 2019, 2019, 2482656. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.M.; Yang, H.B.; Zhao, Y. Study of the temporal and spatial patterns of drought in the Yellow River basin based on SPEI. Sci. China Earth Sci. 2018, 61, 1098–1111. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.M.; Yang, H.B.; Di, D.Y.; Liang, Q.H.; Hussain, Z. Comprehensive evaluation of hydrological drought and its relationships with meteorological drought in the Yellow River basin, China. J. Hydrol. 2020, 584, 124751. [Google Scholar] [CrossRef]
- Li, J.; Xi, M.F.; Pan, Z.W.; Liu, Z.Z.; He, Z.L.; Qin, F. Response of NDVI and SIF to Meteorological Drought in the Yellow River Basin from 2001 to 2020. Water 2022, 14, 2978. [Google Scholar] [CrossRef]
- Qin, G.; Meng, Z.; Fu, Y. Drought and water-use efficiency are dominant environmental factors affecting greenness in the Yellow River Basin, China. Sci. Total Environ. 2022, 834, 155479. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yan, D.H.; Yang, G.Y.; Wang, X.G.; Xiao, W.H.; Zhang, H.T. Assessment on agricultural drought vulnerability in the Yellow River basin based on a fuzzy clustering iterative model. Nat. Hazards 2013, 67, 919–936. [Google Scholar] [CrossRef]
- Jiang, W.G.; Yuan, L.H.; Wang, W.J.; Cao, R.; Zhang, Y.F.; Shen, W.M. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Gao, S.K.; Lai, H.X.; Wang, F.; Qiang, X.M.; Li, H.; Di, D.Y. An Analysis of Spatial–Temporal Evolution and Propagation Features of Vegetation Drought in Different Sub-Zones of China. Agronomy 2023, 13, 2101. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Wen, Z.M.; Chen, Y.M.; Cao, Y.; Ren, J.Y. Spatiotemporal change and trend analysis of potential evapotranspiration over the Loess Plateau of China during 2011–2100. Agric. For. Meteorol. 2017, 233, 183–194. [Google Scholar] [CrossRef]
- Elnashar, A.; Wang, L.J.; Wu, B.F.; Zhu, W.W.; Zeng, H.W. Synthesis of global actual evapotranspiration from 1982 to 2019. Earth Syst. Sci. Data. 2021, 13, 447–480. [Google Scholar] [CrossRef]
- Gebrechorkos, S.H.; Peng, J.; Dyer, E.; Miralles, D.G.; Vicente-Serrano, S.M.; Funk, C.; Beck, H.E.; Asfaw, D.T.; Singer, M.B.; Dadson, S.J. Global high-resolution drought indices for 1981–2022. Earth Syst. Sci. Data 2023, 15, 5449–5466. [Google Scholar] [CrossRef]
- Albarakat, R.; Le, M.H.; Lakshmi, V. Assessment of drought conditions over Iraqi transboundary rivers using FLDAS and satellite datasets. J. Hydrol. -Reg. Stud. 2022, 41, 101075. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, S.Z.; Huang, Q.; Wang, H.; Fang, W.; Yang, Y.Y.; Wang, L. Assessing socioeconomic drought based on an improved Multivariate Standardized Reliability and Resilience Index. J. Hydrol. 2019, 568, 904–918. [Google Scholar] [CrossRef]
- Nalley, D.; Adamowski, J.; Biswas, A.; Gharabaghi, B.; Hu, W. A multiscale and multivariate analysis of precipitation and streamflow variability in relation to ENSO, NAO and PDO. J. Hydrol. 2019, 574, 288–307. [Google Scholar] [CrossRef]
- Hu, W.; Si, B.C. Technical Note: Multiple wavelet coherence for untangling scalespecific and localized multivariate relationships in geosciences. Hydrol. Earth Syst. Sci. 2016, 20, 3183–3191. [Google Scholar] [CrossRef]
- Iqbal, N.; Fareed, Z.; Shahzad, F.; He, X.; Shahzad, U.; Lina, M. The nexus between COVID-19, temperature and exchange rate in Wuhan city: New findings from partial and multiple wavelet coherence. Sci. Total Environ. 2020, 729, 138916. [Google Scholar] [CrossRef] [PubMed]
- Won, J.; Kim, S. Ecological Drought Condition Index to Monitor Vegetation Response to Meteorological Drought in Korean Peninsula. Remote Sens. 2023, 15, 337. [Google Scholar] [CrossRef]
- Shi, Y.; Jin, N.; Ma, X.L.; Wu, B.Y.; He, Q.S.; Yue, C.; Yu, Q. Attribution of climate and human activities to vegetation change in China using machine learning techniques. Agr. Forest Meteorol. 2020, 294, 108146. [Google Scholar] [CrossRef]
- Yu, M.X.; Li, Q.F.; Hayes, M.J.; Svoboda, M.D.; Heim, R.R. Are droughts becoming more frequent or severe in China based on the Standardized Precipitation Evapotranspiration Index: 1951–2010? Int. J. Climatol. 2014, 34, 545–558. [Google Scholar] [CrossRef]
- Zhang, Q.; Peng, J.T.; Singh, V.P.; Li, J.F.; Chen, Y.Q. Spatio-temporal variations of precipitation in arid and semiarid regions of China: The Yellow River basin as a case study. Global Planet. Chang. 2014, 114, 38–49. [Google Scholar] [CrossRef]
- Miao, C.Y.; Sun, Q.H.; Duan, Q.Y.; Wang, Y.F. Joint analysis of changes in temperature and precipitation on the Loess Plateau during the period 1961–2011. Clim. Dyn. 2016, 47, 3221–3234. [Google Scholar] [CrossRef]
- Huo, S.Q.; Wang, H.B.; Peng, M.X. Analysis on No-flow in the Lower Yellow River in 1997. Yellow River 1998, 20, 1–3. [Google Scholar]
- Harris, B.L.; Taylor, C.M.; Weedon, G.P.; Talib, J.; Dorigo, W.; van der Schalie, R. Satellite-Observed Vegetation Responses to Intraseasonal Precipitation Variability. Geophys. Res. Lett. 2022, 49, e2022GL099635. [Google Scholar] [CrossRef]
- Wen, X.; Shao, H.Y.; Wang, Y.; Lv, L.F.; Xian, W.; Shao, Q.F.; Shu, Y.; Yin, Z.Q.; Liu, S.H.; Qi, J.G. Assessment of the Spatiotemporal Impact of Water Conservation on the Qinghai–Tibet Plateau. Remote Sens. 2023, 15, 3175. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Barnes, M.L.; Bowling, D.R.; Driscoll, A.W.; Guo, G.S.; Anderegg, W.R.L. Quantifying the drivers of ecosystem fluxes and water potential across the soil-plant-atmosphere continuum in an arid woodland. Agr. Forest Meteorol. 2023, 329, 109269. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PWC | Scale | ET | AH | PC | SM | ST | AT |
|---|---|---|---|---|---|---|---|
| AWC | Small | 0.91 | 0.91 | 0.91 | 0.91 | 0.91 | 0.91 |
| Medium | 0.91 | 0.90 | 0.90 | 0.90 | 0.91 | 0.91 | |
| Large | 0.97 | 0.92 | 0.96 | 0.96 | 0.95 | 0.96 | |
| Total | 0.96 | 0.91 | 0.95 | 0.95 | 0.92 | 0.94 | |
| PASC (%) | Small | 7.22 | 5.89 | 2.75 | 4.42 | 6.92 | 5.41 |
| Medium | 5.82 | 4.48 | 6.33 | 4.21 | 5.34 | 5.06 | |
| Large | 26.27 | 3.02 | 21.98 | 24.67 | 6.21 | 19.40 | |
| Total | 13.56 | 4.35 | 10.94 | 12.14 | 6.47 | 11.24 |
| Univariate | AWC | PASC (%) | Bivariate | AWC | PASC (%) | Trivariate | AWC | PASC (%) |
|---|---|---|---|---|---|---|---|---|
| ET | 0.96 | 13.56 | ET-AH | 0.93 | 22.57 | ET-PC-AH | 0.96 | 14.44 |
| AH | 0.91 | 4.35 | ET-PC | 0.93 | 23.85 | ET-PC-SM | 0.97 | 24.49 |
| PC | 0.95 | 10.94 | ET-SM | 0.93 | 22.29 | ET-PC-ST | 0.96 | 15.48 |
| SM | 0.95 | 12.14 | ET-ST | 0.93 | 18.80 | ET-PC-AT | 0.96 | 14.76 |
| ST | 0.92 | 6.47 | ET-AT | 0.92 | 19.33 | |||
| AT | 0.94 | 11.24 |
| Copula Families | Mathematical Descriptions | Parameter Settings |
|---|---|---|
| Gaussian copula | ||
| t copula | ||
| Clayton copula | \0 | |
| Frank copula | ||
| Gumbel copula |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Men, R.; Yan, S.; Lai, H.; Wang, Z.; Feng, K.; Gao, S.; Li, Y.; Guo, W.; Qu, Y. Vegetation’s Dynamic Changes, Spatial Trends, and Responses to Drought in the Yellow River Basin, China. Agronomy 2024, 14, 1724. https://doi.org/10.3390/agronomy14081724
Wang F, Men R, Yan S, Lai H, Wang Z, Feng K, Gao S, Li Y, Guo W, Qu Y. Vegetation’s Dynamic Changes, Spatial Trends, and Responses to Drought in the Yellow River Basin, China. Agronomy. 2024; 14(8):1724. https://doi.org/10.3390/agronomy14081724
Chicago/Turabian StyleWang, Fei, Ruyi Men, Shaofeng Yan, Hexin Lai, Zipeng Wang, Kai Feng, Shikai Gao, Yanbin Li, Wenxian Guo, and Yanping Qu. 2024. "Vegetation’s Dynamic Changes, Spatial Trends, and Responses to Drought in the Yellow River Basin, China" Agronomy 14, no. 8: 1724. https://doi.org/10.3390/agronomy14081724