
How Long Should Grasses of South American Campos Grasslands Rest for Stockpiling Forage?

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
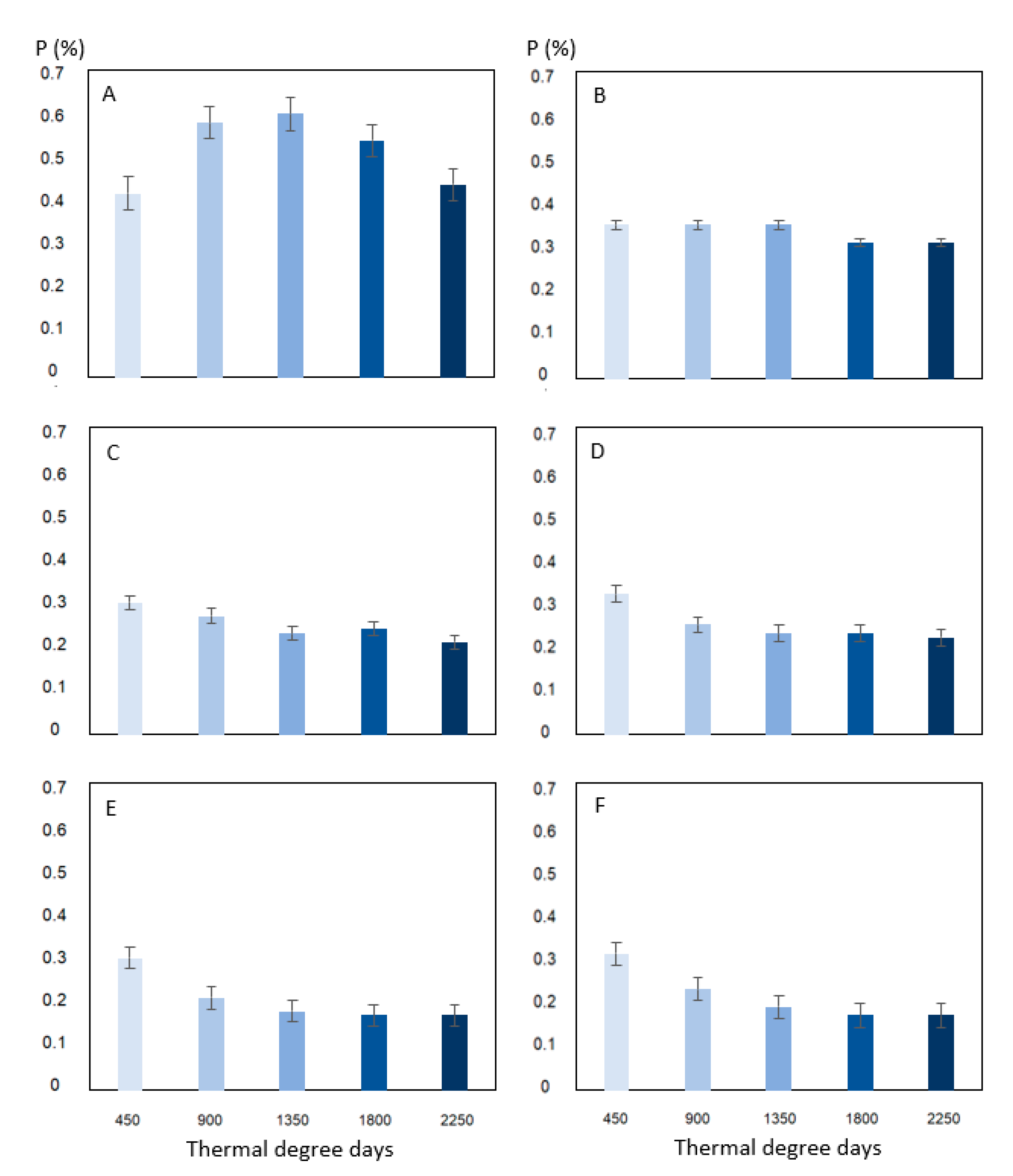
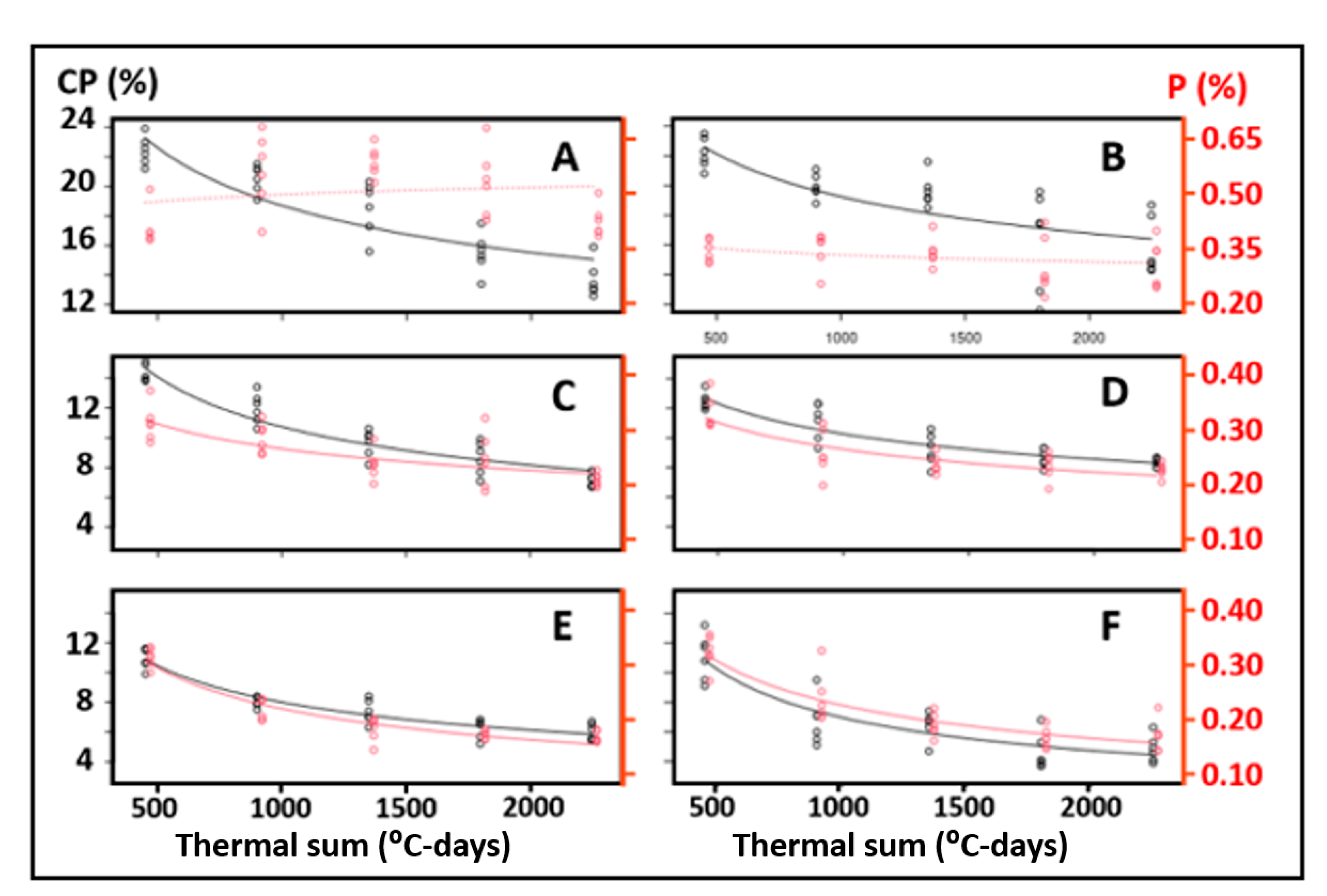
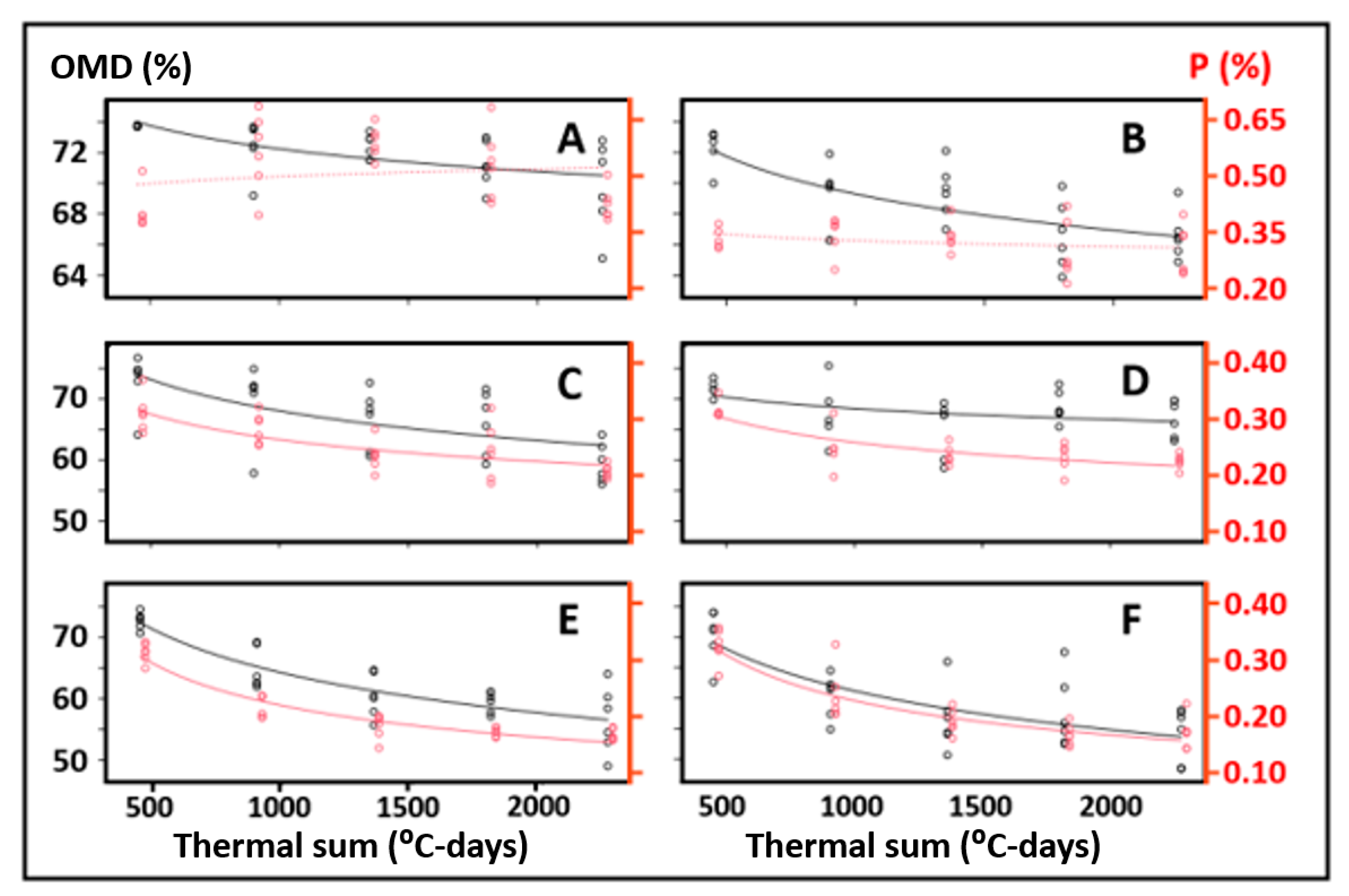
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pallares, O.R.; Berretta, E.J.; Maraschin, G.E. The South American Campos ecosystem. In Grasslands of the World; Suttie, J.M., Reynolds, S.G., Batello, C., Eds.; FAO: Rome, Italy, 2005; Volume 42, pp. 171–219. [Google Scholar]
- Gibson, D.J. Grasses and Grassland Ecology, 1st ed.; Oxford University Press: Oxford, UK, 2009; 313p. [Google Scholar]
- McIvor, J.G.; Guppy, C.; Probert, M.E. Phosphorus requirements of tropical grazing systems: The northern Australian experience. Plant Soil 2011, 349, 55–67. [Google Scholar] [CrossRef]
- Jaurena, M.; Durante, M.; Devincenzi, T.; Savian, J.V.; Bendersky, D.; Moojen, F.G.; Pereira, M.; Soca, P.; Quadros, F.L.F.; Pizzio, R.; et al. Native grasslands at the core: A new paradigm of intensification for the Campos of Southern South America to increase economic and environmental sustainability. Front. Sustain. Food Syst. 2021, 5, 11. [Google Scholar] [CrossRef]
- Mezzalira, J.C.; Carvalho, P.C.d.F.; da Trindade, J.K.; Bremm, C.; Fonseca, L.; do Amaral, M.F.; Reffatti, M.V. Produçao animal e vegetal em pastagem nativa manejada sob diferentes ofertas de forragem por bovinos. Cienc. Rural 2012, 42, 1264–1270. [Google Scholar] [CrossRef]
- Cruz, P.; Lezana, L.; Durante, M.; Jaurena, M.; Figari, M.; Bittencourt, L.; Theau, J.P.; Massa, E.; Viegas, J.; Quadros, F.L.F. A functional classification of 63 common Poaceae in the “Campos” grasslands of South America. Ecol. Austral. 2019, 29, 239–248. [Google Scholar] [CrossRef]
- Andrade, B.O.; Marchesi, E.; Burkart, S.; Setubal, R.B.; Lezama, F.; Perelman, S.; Schneider, A.A.; Trevisan, R.; Overbeck, G.E.; Boldrini, I.I. Vascular plant species richness and distribution in the Río de la Plata grasslands. Bot. J. Linn. Soc. 2018, 188, 250–256. [Google Scholar] [CrossRef]
- Claramunt, M.; Fernández-Foren, A.; Soca, P. Effect of herbage allowance on productive and reproductive responses of primiparous beef cows grazing on Campos grassland. Anim. Prod. Sci. 2017, 58, 1615–1624. [Google Scholar] [CrossRef]
- Poppi, D.P.; Quigley, S.P.; Silva, T.A.C.C.D.; McLennan, S.R. Challenges of beef cattle production from tropical pastures. Rev. Bras. Zoot. 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Ansquer, P.; Al Haj Kahled, R.; Cruz, P.; Theau, J.P.; Thérond, O.; Duru, M. Characterizing and predicting plant phenology in species rich grasslands. Grass Forage Sci. 2009, 64, 57–70. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Derner, J.D.; Augustine, D.J. Adaptive management for drought on rangelands. Rangelands 2016, 38, 211–215. [Google Scholar] [CrossRef]
- Scasta, J.D.; Lalman, D.L.; Henderson, L. Drought mitigation for grazing operations: Matching the animal to the environment. Rangelands 2016, 38, 204–210. [Google Scholar] [CrossRef]
- Berretta, E.J.; Risso, D.F.; Montossi, F.; Pigurina, G. Campos in Uruguay. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., Moraes, A., Carvalho, P.C.F., Nabinger, C., Eds.; CAB International: Wallingford, UK, 2000; pp. 377–394. [Google Scholar] [CrossRef]
- Giménez, A.; Castaño, J.P.; Baethgen, W.; Lanfranco, B. Cambio Climático en Uruguay, Posibles Impactos y Medidas de Adaptación en el Sector Agropecuario; Serie Técnica 151; INIA: Montevideo, Uruguay, 2009; 50p. [Google Scholar]
- Buttolph, L.P.; Coppock, D.L. Influence of deferred grazing on vegetation dynamics and livestock productivity in an Andean pastoral system. J. Appl. Ecol. 2004, 41, 664–674. [Google Scholar] [CrossRef]
- Lemaire, G.; Belanger, G. Allometries in plants as drivers of forage nutritive value: A review. Agriculture 2019, 10, 5. [Google Scholar] [CrossRef]
- Andrade, I.F.; Atkinson, L.G.; Sollenberger, L.E.; Ruegsegger, G.J.; Mislevy, P.; Kalmbacher, R.S. Stockpiling herbaceous tropical legumes for dry season feed in Jamaica. Trop. Grassl. 1998, 32, 166–172. [Google Scholar]
- Karn, J. Phosphorus nutrition of grazing cattle: A review. Anim Feed Sci. Tech. 2001, 89, 133–153. [Google Scholar] [CrossRef]
- Malafaia, P.; Barbosa, J.D.; Brito, M.F.; Souza, V.C.D.; Costa, D.F. A Phosphorus for Cattle and Buffaloes in Brazil: Clinical Signs and Diagnosis of Its Deficiency and Relevance, and Recommended Strategies to Alleviate Issues Observed under Grazing Conditions. Ruminants 2023, 3, 55–75. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Wiles, P.G.; Gray, I.K.; Kissling, R.C.; Delahanty, C.; Evers, J.; Greenwood, K.; Grimshaw, K.; Hibbert, M.; Kelly, K.; Luckin, H.; et al. Routine analysis of proteins by Kjeldahl and Dumas methods: Review and interlaboratory study using dairy products. J. AOAC Int. 1998, 81, 620–632. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 17th ed.; Association of official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- Kozloski, G.V.; Zilio, E.M.C.; Ongarato, F.; Kuinchtner, B.C.; Saccol, A.G.; Genro, T.C.M.; Oliveira, L.; Faria, B.M.; Cezimbra, I.M.; Quadros, F.L.F. Faecal N excretion as an approach for estimating organic matter intake by free-ranging sheep and cattle. J. Agric. Sci. 2018, 156, 443–449. [Google Scholar] [CrossRef]
- Roberts, D. The statistical program SYSTAT: Wilkinson, L. SYSTAT, The System for Statistics. SYSTAT, Inc. 2902 Central Street, Evanston, IL 60201. J. Sch. Psychol. 1987, 25, 313–318. [Google Scholar] [CrossRef]
- McIvor, J.G. Seasonal changes in dry matter distribution and herbage quality of Urochloa species in north-eastern Queensland. Aust. J. Exp. Agric. 1990, 30, 523–528. [Google Scholar] [CrossRef]
- McDowell, L.R. Feeding minerals to cattle on pasture. Anim. Feed Sci. Technol. 1996, 60, 247–271. [Google Scholar] [CrossRef]
- McIvor, J.G. Seasonal changes in the growth, dry matter distribution and herbage quality of three native grasses in northern Queensland. Aust. J. Exp. Agric. 1981, 21, 600–609. [Google Scholar] [CrossRef]
- Dixon, R.M.; Sullivan, M.T.; O’Connor, S.N.; Mayer, R.J. Diet quality, liveweight change and responses to N supplements by cattle grazing Astrebla spp. (Mitchell grass) pastures in the semi-arid tropics in north-western Queensland, Australia. Rangel. J. 2022, 44, 97–113. [Google Scholar] [CrossRef]
- Grings, E.E.; Haferkamp, M.R.; Heitschmidt, R.K.; Karl, M.G. Mineral dynamics in forages of the Northern Great Plains. J. Range Manag. 1996, 49, 234–240. [Google Scholar] [CrossRef]
- Elejalde, D.A.G.; Nabinger, C.; Pascual, M.G.C.; Ferreira, E.T.; Missio, R.L.; Kunrath, T.R.; Devincenzi, T.; Cardoso, R.R. Quality of the forage apparently consumed by beef calves in natural grassland under fertilization and oversown with cool season forage species. Rev. Bras. Zoot. 2012, 41, 1360–1368. [Google Scholar] [CrossRef]
- Heringer, I.; Jacques, A.V.A. Qualidade da forragem de pastagem nativa sob distintas alternativas de manejo. Pesq. Agrop. Bras. 2002, 37, 399–406. [Google Scholar] [CrossRef]
- Silveira, V.C.P.; Vargas, A.F.D.C.; Oliveira, J.O.R.; Gomes, K.E.; Motta, A.F. Qualidade da pastagem nativa obtida por diferentes métodos de amostragem e em diferentes solos na Apa do Ibirapuitã, Brasil. Cienc. Rural 2005, 35, 582–588. [Google Scholar] [CrossRef]
- Van Soest, P. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: New York, NY, USA, 1989; 528p. [Google Scholar]
- Poppi, D.; McLennan, S. Protein and Energy Utilization by Ruminants at pasture. J. Anim. Sci. 1995, 73, 278–290. [Google Scholar] [CrossRef]
- Cano, C.C.P.; Cecato, U.; Canto, M.W.D.; Santos, G.T.D.; Galbeiro, S.; Martins, E.N.; Mira, R.T. Valor nutritivo do capim-Tanzânia (Panicum maximum Jacq. cv. Tanzânia-1) pastejado em diferentes alturas. Rev. Bras. Zoot. 2004, 33, 1959–1968. [Google Scholar] [CrossRef]
- Azambuja Filho, J.C.R.; Carvalho, P.C.F.; Bonnet, O.J.F.; Bastianelli, D.; Jouven, M. Functional classification of feed items in pampa grassland, based on their near-infrared spectrum. Rangel. Ecol. Manag. 2020, 73, 358–367. [Google Scholar] [CrossRef]
- Hacker, J.B.; Minson, D.J. The digestibility of plant parts. Herb. Abstr. 1981, 51, 459–482. [Google Scholar]
- Insua, J.R.; Agnusdei, M.G.; Utsumi, S.A.; Berone, G.D. Morphological, environmental and management factors affecting nutritive value of tall fescue (Lolium arundinaceum). Crop Pasture Sci. 2018, 69, 1165–1172. [Google Scholar] [CrossRef]
- Baudracco, J.; Lopez-Villalobos, N.; Holmes, C.W.; Macdonald, K.A. Effects of stocking rate, supplementation, genotype and their interactions on grazing dairy systems: A review. N. Z. J. Agric. Res. 2010, 53, 109–133. [Google Scholar] [CrossRef]
- Jackson, D.; Rolfe, J.; English, B.; Mathews, R.; Dixon, R.M.; Smith, P.; MacDonald, M. Phosphorus Management of Beef Cattle in Northern Australia, 1st ed; Meat and Livestock Australia: Sydney, Australia, 2012; 44p. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. auleticus | N. neesiana | |||||||||
| TS | L | S | D | FM | TS | L | S | D | FM | |
| P | 0.01 | 0.09 | −0.11 | −0.06 | 0.06 | −0.32 | 0.14 | −0.19 | −0.06 | −0.6 |
| TS | −0.88 * | 0.26 | 0.91 * | 0.94 * | −0.70 * | 0.27 | 0.65 | 0.72 * | ||
| S | −0.52 | −0.98 * | −0.83 * | −0.39 | −0.92 * | −0.38 | ||||
| D | 0.35 | 0.17 | 0.01 | 0.26 | ||||||
| FM | 0.88 * | 0.26 | ||||||||
| A. laterallis | P. notatum | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TS | L | S | D | FM | TS | L | S | D | FM | |
| P | −0.74 * | 0.6 | −0.39 | −0.43 | −0.81 * | −0.72 * | 0.88 * | −0.71 * | −0.39 | −0.80 * |
| TS | −0.53 | 0.26 | 0.67 | 0.92 * | −0.54 | 0.27 | 0.42 | 0.84 * | ||
| S | −0.91 * | −0.24 | −0.55 | −0.73 * | −0.54 | −0.76 * | ||||
| D | −0.1 | 0.3 | −0.17 | 0.47 | ||||||
| FM | 0.56 | 0.51 | ||||||||
| M. selloana | P. dilatatum | |||||||||
| TS | L | S | D | FM | TS | L | S | D | FM | |
| P | −0.85 * | 0.61 | −0.05 | −0.59 | −0.81 * | −0.82 * | 0.74 * | −0.11 | −0.48 | −0.80 * |
| TS | −0.46 | −0.23 | 0.75 * | 0.83 * | −0.78 * | −0.16 | 0.71 * | 0.86 * | ||
| S | −0.62 | −0.39 | −0.59 | −0.19 | −0.65 | −0.79 * | ||||
| D | −0.47 | −0.07 | −0.59 | −0.01 | ||||||
| FM | 0.61 | 0.61 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Risso Barbosa, M.; Durante, M.; Marin, L.; Cazzuli, F.; Ferreira de Quadros, F.L.; Dixon, R.M.; Riet Correa, F.; Jaurena, M. How Long Should Grasses of South American Campos Grasslands Rest for Stockpiling Forage? Agronomy 2024, 14, 1790. https://doi.org/10.3390/agronomy14081790
Risso Barbosa M, Durante M, Marin L, Cazzuli F, Ferreira de Quadros FL, Dixon RM, Riet Correa F, Jaurena M. How Long Should Grasses of South American Campos Grasslands Rest for Stockpiling Forage? Agronomy. 2024; 14(8):1790. https://doi.org/10.3390/agronomy14081790
Chicago/Turabian StyleRisso Barbosa, Marlon, Martín Durante, Luciana Marin, Fiorella Cazzuli, Fernando Luiz Ferreira de Quadros, Rob M. Dixon, Franklin Riet Correa, and Martin Jaurena. 2024. "How Long Should Grasses of South American Campos Grasslands Rest for Stockpiling Forage?" Agronomy 14, no. 8: 1790. https://doi.org/10.3390/agronomy14081790