Abstract
Acidovorax citrulli (Ac) is an important pathogenic bacterium causing bacterial fruit blotch (BFB) in Cucurbitaceae plants and is an important quarantine pest in China. This study was conducted to establish a rapid, convenient, and accurate visual method for detecting A. citrulli. A. citrulli-specific primers and a prober were designed based on the conserved region of the YD-repeat protein gene. Loop-mediated isothermal amplification combined with lateral flow dipstick (LAMP-LFD) was used to establish an assay for the rapid visual detection of A. citrulli by optimizing the reaction temperature and time. The specificity, sensitivity, and performance of the optimized LAMP-LFD assay were evaluated using the genomic DNA of the tested isolates, A. citrulli pure culture, infested seeds, commercial seeds, and leaf samples. The optimal assay temperature and time were 64 °C and 60 min, respectively. The assay specifically detected A. citrulli, and no cross-reactions were observed with the genomic DNA of other closely related species. The detection sensitivity of the LAMP-LFD for detecting pure genomic DNA, the bacterial suspension, bacterial amount on seeds (colony-forming units (CFU)·g−1), and infection rate of seeds (%) were 1 fg·μL−1, 8 CFU·mL−1, 5 CFU·g−1, and 0.05% infestation per reaction, respectively. The positive detection rate of the LAMP-LFD assay was 20–100% in seed samples (n = 1000 seeds) with 0.05–0.1% infestation. The LAMP-LFD assay rapidly and accurately detected A. citrulli in seeds and leaf tissues carrying pathogens. This assay thus offers the advantages of easy operation, rapidity, high specificity and sensitivity, low cost (no need for complex and expensive precision instruments), visualization of detection results, good stability, and strong applicability, which can be used for epidemiological studies and disease management.
1. Introduction
Bacterial fruit blotch (BFB), caused by Acidovorax citrulli Schaad et al., is a devastating bacterial disease that endangers the production of cucurbit plants such as watermelons and other melons. It has been listed as a quarantine pest of imported plants by many countries and regions, including China [1,2,3,4,5,6,7]. This pathogen is widely distributed throughout the world and can cause damage to melon crops at all growth stages. It frequently leads to large-scale outbreaks, mainly damaging the leaves and fruits of plants. Typical symptoms of diseased plants include water-soaked dark or olive-green lesions on the leaves or rind surfaces. The rinds may crack in severe cases, eventually causing the fruit to decay, which can affect the yield and quality of melon crops. The incidence of BFB in general fields is 45–75%, with yield losses of up to 50–90%, resulting in substantial economic losses. Furthermore, this disease is increasing, posing a serious threat to the production of cucurbit plants. Therefore, there is an urgent need for effective methods to prevent and control this disease [8,9,10,11,12]. BFB is a typical seedborne bacterial disease, and seeds carrying A. citrulli are the most important source of infection. The disease is characterized by a quick onset, explosive outbreaks, rapid spread, and difficulty in prevention and control. Once infection occurs, the resulting disease can bring about massive yield losses in melon crops [13,14,15]. The rapid and accurate detection of A. citrulli in the early stages of disease and the prompt adoption of effective control measures are important for preventing the occurrence, transmission, and spread of this pathogen. Therefore, establishing a rapid, simple, highly specific, highly sensitive, and applicable technique for A. citrulli detection and the appropriate treatment of infected seeds are crucial to preventing this disease [16,17,18,19,20].
Conventional methods for the detection of A. citrulli mainly include semiselective agar medium plating techniques [10,21,22], the enzyme immunosorbent assay [23,24], immunomagnetic beads/microsphere immunoassay [25,26], serological assays [27,28,29,30], and pathogenicity tests [31,32]. However, these methods have low sensitivity, poor specificity, complexity in their operations, and long detection cycles. Hence, they are not suitable for testing large batches of seeds and latently infected plant tissues [33,34]. With advances in rapid molecular detection technologies for phytopathogens, the classic polymerase chain reaction (PCR) assay [35], LAMP method [36], droplet digital PCR (ddPCR) [34], real-time, fluorescence-based quantitative PCR [37,38,39], padlock probe-based PCR [40], bio-PCR [5,22], immuno-capture PCR [41], cross-priming amplification (CPA) [42], and other nucleic acid amplification techniques have been successfully applied in the rapid detection of A. citrulli. Tian et al. [3] designed a pair of primers targeting the hrpB2 gene for specific PCR detection of A. citrulli. These specific primers only amplified A. citrulli, presenting negative PCR results for all other test strains. Sensitivity testing demonstrated that this method could detect 103 CFU/mL−1 of target bacteria, and the positive detection rate of A. citrulli in commercially available melon seeds was 23.52%. Wang et al. [4] designed three primers targeting the BOX short repeated sequences of A. citrulli and established a nested PCR method with BX-L1/BX-R5 as outer primers and BX-L1/BX-S-R2 as inner primers. The limit of detection (LOD) for pure A. citrulli cultures and extracts of artificially infected seeds was 4.7 CFU/mL−1, which was 1000-fold higher than with classic PCR. When the infestation level of watermelon seeds was 0.1–10%, the positive detection rate of nested PCR was 66.7–100%. Kan et al. [5] combined semiselective agar medium plating with PCR to establish an A. citrulli bio-PCR detection method. The LOD of PCR alone was 102 CFU/mL−1. When two types of semiselective media, ASCM and EBBA, were combined with PCR for bio-PCR detection, the LOD of ASCM-PCR for the bacterial load of infected seeds was 0.01 CFU/g−1, while that of EBBA-PCR was 0.1 CFU/g−1. Zhao et al. [34] employed ddPCR to establish an A. citrulli ddPCR detection method. The LODs for bacterial suspensions and DNA samples were 103 CFU/g−1 and 1 pg/μL−1, respectively, and were 10-fold higher than those of real-time PCR under the same conditions. When testing artificially infected watermelon and melon seeds with an average bacterial load of 105–106 CFU/mL−1, ddPCR allowed for the detection of bacteria in seed samples with an infestation level of 0.2%. Tian et al. used the 16S–23S internal transcribed spacer ribosomal DNA sequence to design specific primers and probes for A. citrulli that were used to perform padlock probe detection of A. citrulli. The detection sensitivity was 100 fg/μL−1, and the detection accuracy for artificially inoculated melon seedlots was 100%, thus demonstrating greater effectiveness, sensitivity, and accuracy than conventional PCR based on the primers WFB1/WFB2 [40]. Zhang et al. established an A. citrulli-CPA detection method using 16S rDNA as the target gene, which yielded a detection sensitivity of 3.7 × 103 CFU/mL−1 and was successfully applied to the detection of watermelon seeds carrying A. citrulli [42]. Cho et al. developed an A. citrulli–SYBR Green-based real-time, fluorescence-based quantitative PCR assay targeting the YD-repeat protein of A. citrulli, which showed a detection sensitivity of 5 fg/μL−1. Thus, this system can be used for the detection of A. citrulli in cucurbit seeds and plant tissues [38].
Compared with conventional A. citrulli identification and detection methods, PCR-based detection techniques have shown several advantages, including good specificity, high specificity, rapid testing times, and high efficiency while also improving the detection accuracy to some extent [16]; however, PCR-based detection techniques pose greater needs for instruments and equipment, experimental environments, and operator proficiency. Hence, these techniques have a number of disadvantages, including high costs, complex operations, low detection sensitivity, heavy reliance on instruments and equipment, need for a certain level of operating proficiency, susceptibility to leaking trace amounts of pathogens, and difficulty in implementing on-site detection, all of which have greatly limited the application of such techniques [33,42,43]. Loop-mediated isothermal amplification (LAMP) is a rapid nucleic acid amplification method. It involves designing two pairs of primers targeting six loci in the conserved regions of a target gene and uses a Bst DNA polymerase with high strand displacement activity to perform amplification at a constant temperature (around 63 °C) for 30 to 60 min of a large amount of target DNA [44,45]. Currently, detection techniques that combine LAMP with a lateral flow dipstick (LFD), i.e., LAMP-LFD, have been successfully applied to rapidly detect various pests such as pathogenic bacteria [46,47,48,49], phytopathogenic viruses [50,51], phytopathogenic fungi [52,53,54], root-knot nematodes [55,56], mycoplasma [57], and harmful algal blooms [58]. However, to date, there have been no reports on the application of LAMP-LFD for the detection of A. citrulli.
Therefore, in this study, we designed specific primers and probes targeting the YD-repeat protein of A. citrulli and optimized the reaction conditions to establish a LAMP-LFD assay that was then subjected to analyses of specificity, sensitivity, and applicability. This LAMP-LFD system can serve as a new method for the rapid, simple, sensitive, and accurate detection of A. citrulli, as well as provide technical support for the effective and convenient prevention and control of BFB.
2. Materials and Methods
2.1. Test Strains
A total of 32 strains were tested, including 6 strains of A. citrulli and 26 strains of common closely related species. Details on the strain names, hosts, and sources are listed in Table 1.

Table 1.
Bacterial strains used in this study.
2.2. Reagents and Seeds
The main test reagents were as follows: Luria–Bertani (LB) broth medium (tryptone, 10.00 g/L−1; yeast extract, 5.00 g/L−1, NaCl 10.00 g/L−1), pH 7.2, autoclaved at 121 °C for 15 min; solid LB medium (17 g of agar powder added per 1000 mL of LB broth); a LAMP DNA amplification kit and fluorescent detection reagent (FD), which were purchased from EIKEN China Co., Ltd. (Shanghai, China); a lateral flow dipstick (Milenia GenLine HybriDetect Kit, Milenia GenLine HybriDetect-MGHD 1), which was purchased from Milenia Biotec GmbH (Gieben, Germany); and a bacterial genomic DNA extraction kit (spin column type, DP302-02), which was purchased from Tiangen Biotech Co., Ltd. (Beijing, China). All reagents were analytically pure and could be used without further purification.
The seeds for testing were purchased from the local seed market in Fuzhou City, Fujian, China, included 18 watermelon seedlots and 12 melon seedlots. Details such as the seed names and production companies are listed in Table 2.
Table 2.
Results of LAMP-LFD assay for detecting A. citrulli in commercially available seeds.
2.3. DNA Extraction
All test strains were streaked onto solid LB medium plates and cultured at 28 °C for 24 to 48 h. Single colonies were picked, inoculated into LB broth medium, and cultured for 24 h with shaking at 28 °C and 150 rpm to obtain bacterial suspensions. Each bacterial suspension was collected in a 1.5 mL centrifuge tube for centrifugation at 13,800× g for 3 min, the supernatant was discarded, and the pellet was retained. The genomic DNA of each test strain was extracted using a bacterial genomic DNA extraction kit (DP302-02, Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer’s protocol and dissolved in 60 μL of ddH2O. Purified DNA was measured using a spectrophotometer (K5800C, Beijing Kaiao Technology Developmemt Co., Ltd., Beijing, China), diluted to 50 ng/μL−1, and then stored at –20 °C for later use.
2.4. Primer and Probe Design
Based on the gene sequences of six loci in the Aave_2049 conserved region of the A. citrulli YD-repeat protein (accession number: CP029373.0), the online LAMP primer design software Primer Explorer V5 [https://primerexplorer.jp/lampv5/index.html (accessed 20 September 2012); Eiken Chemical Co., Japan] was used to design two pairs of specific primers (one pair of outer primers, F3 and B3, and one pair of inner primers, FIP and BIP, with FIP composed of F2 and F1c, and BIP composed of B2 and B1c) for the A. citrulli LAMP reaction. In addition, Primer Premier 6.0 software [http://www.premierbiosoft.com (accessed on 30 January 2011)] was used to design a probe (HP) that could specifically hybridize with LAMP products based on the sequences between primers F1c and B1c for the LFD assay. Details on the loci and sequences of the primers and probes are shown in Figure 1 and Table 3. The 5′ end of primer FIP was labeled with biotin, and the 5′ end of probe HP was labeled with 6-carboxyfluorescein (6-FAM). The primers and probes were synthesized by Sangon Biotech (Shanghai, China) Co., Ltd.
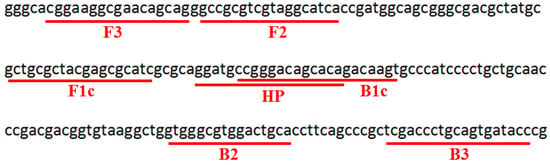
Figure 1.
Primers and DNA probe designed based on YD-repeat protein gene of LAMP-LFD assay for detection of A. citrulli. F3, F2, and F1c: the forward primers in the LAMP reaction, with the reverse complement sequence of F1c and F2 composed of the forward inner primer FIP. B3, B2, and B1c: the reverse primers in the LAMP reaction, with B2 and B1c composed of the reverse inner primer. HP: the DNA probe used in LFD detection. The sequences and sites of the primer and probe are underlined in red.
Table 3.
Primers and DNA probe designed for detecting A. citrulli causing BFB disease via LAMP-LFD assay.
2.5. Optimization of LAMP-LFD Reaction Conditions
LAMP reaction system: The LAMP reaction mixture was prepared according to the instructions of the LAMP DNA Amplification Kit (LAMP), with reference to the methods described by Lan et al. [43]. The LAMP assay was conducted using a total volume of 25 μL, which contained 2× reaction buffer (RM), 12.5 μL; 0.2 μM outer primers (F3 and B3) and 1.6 μM inner primers (FIP and BIP), 1.0 μL each; Bst DNA polymerase (8 U/μL−1), 1.0 μL; nucleic acid fluorescent dye (FD), 1.0 μL; A. citrulli genomic DNA, 1.0 μL; and sterilized ddH2O to reach 25 μL. A. citrulli genomic DNA served as the positive control and sterilized ddH2O as the negative control.
Optimization of LAMP reaction conditions: To obtain the optimal LAMP reaction conditions, the prepared LAMP reaction mixture was incubated in a water bath (DK-8D, Shanghai Yiheng Technology Co., Ltd., Shanghai, China) at different temperatures (60, 61, 62, 63, 64, 65, 66, and 67 °C) for 60 min, and the appropriate reaction temperature was determined based on the amplification results. Under the optimal reaction temperature (64 °C, see Section 3.1), amplification was carried out for 15, 30, 45, 60, and 75 min, and the optimal reaction time was determined based on the amplification results. The final LAMP reaction mixture was heated at 80 °C for 5.0 min to inactivate the enzymes and terminate the reaction. All experiments were performed in triplicate.
Detection of LAMP amplification products: Two methods were used to detect LAMP amplification products, namely visual inspection of color change in the amplification products and agarose gel electrophoresis. If the reaction mixture changed from orange (pre-amplification) to yellow-green under normal, natural light and yielded a white turbid precipitate under ultraviolet light (wavelength: 365 nm), then the amplification product was positive. If the reaction mixture remained orange or orange-yellow under normal, natural light before and after the amplification and was colorless and transparent under ultraviolet light (wavelength: 365 nm), then the LAMP reaction was negative. To confirm the accuracy of the visual inspection method, 5.0 μL of LAMP amplification products were obtained for 2.0% agarose gel electrophoresis. If typical ladder-like DNA bands were observed, the reaction was positive; if typical ladder-like DNA bands were absent, the reaction was negative. The LAMP detection results were only considered accurate if the results of visual inspection and agarose gel electrophoresis agreed.
LFD assay: DNA was amplified using the optimized LAMP method, but the reaction was not terminated at 80 °C for 5 min. Instead, 2.0 μL of 10 μmol/L−1 probe HP was added to the amplification reaction mixture and hybridized for 5 min before the reaction was terminated by heating at 80 °C for 5 min. After hybridization, the LFD assay was performed on the amplification products, with 10.0 μL of the hybridization mixture mixed evenly with 90 μL of HybriDetect Assay Buffer (Milenia Biotec GmbH, Gieben, Germany). The detection end of the LFD dipstick was immersed vertically into the mixture and left to react at room temperature for 5 to 10 min. The results were observed visually on the dipstick: if two purple-red bands appeared on the dipstick, with one in the quality control area (C) and one in the test area (T), then the result was positive, indicating that the amplification product was positive. If only one purple-red band appeared in the quality control area and none in the test area, then the result was negative. All experiments were performed in triplicate.
2.6. Specificity Assay
To verify the specificity of LAMP-LFD for A. citrulli, the genomes of six strains of A. citrulli, five closely related Acidovorax strains, and twenty-one strains from another eight genera (Table 1) were used as templates. Specificity validation was performed using the LAMP-LFD method optimized in Section 2.5, with the genomic DNA of A. citrulli used as the positive control and sterilized ddH2O used as the negative control. The detection of the LAMP amplification products was performed through visual inspection, 2% agarose gel electrophoresis, and LFD. All experiments were performed in triplicate.
2.7. Sensitivity Assay
Sensitivity analysis using genomic DNA: A. citrulli Ac-1 genomic DNA was subjected to a 10-fold gradient dilution using sterilized ddH2O. Genomic DNA samples at 10 concentrations (1 ng/μL−1, 100 μg/μL−1, 10 μg/μL−1, 1 pg/μL−1, 100 fg/μL−1, 10 fg/μL−1, 1 fg/μL−1, 100 ag/μL−1, 10 ag/μL−1, and 1 ag/μL−1) were used as LAMP-LFD reaction templates, and sterile water was used as the negative control. The LAMP-LFD assay was performed as previous optimized (Section 2.5). The experiments were conducted in triplicate under the same conditions.
Sensitivity analysis using bacterial cultures: To determine the lowest bacterial concentration required for LAMP-LFD to detect A. citrulli, the A. citrulli Ac-1 strain that was stored at –80 °C was streaked onto solid LB medium plates and cultured at 28 °C for 24 to 48 h. Single colonies were picked, inoculated into LB broth medium, and cultured with shaking at 28 °C and 150 rpm for 24 h to obtain bacterial LB broth. The bacterial LB broth was then centrifuged at 9580 g for 1 min, the supernatant discarded, and the bacterial pellet was resuspended in ddH2O to prepare the bacterial suspension. The concentration of the resulting bacterial suspension was measured using a spectrophotometer (OD600) combined with a conventional plate count method. The absorbance value measured using the spectrophotometer (OD600) served as a reference for subsequent bacterial suspension concentrations. The concentration of the bacterial suspension was first adjusted to 1.0 × 107 CFU/mL−1 (OD600 = 0.5), followed by 10- or 5-fold serial dilutions with ddH2O. The concentrations corresponding to the different series of bacterial suspensions were 103, 102, 50, 10, 8, 4, 2, and 1 CFU/mL−1, respectively. With reference to the methods of Kan et al. [5] and Hui et al. [59], 1.0 mL of bacterial suspension was pipetted into a 1.5 mL centrifuge tube, vortexed, and placed in boiling water for 10 min to obtain the bacterial suspension template DNA for LAMP-LFD sensitivity testing. The experiments were conducted in triplicate under the same conditions.
2.8. Evaluation of LAMP-LFD Assay for Detection of A. citrulli in Seeds
Preparation of artificially infected seeds: Referring to the methods described by Walcott et al. [2,60], 1000 healthy watermelon seeds previously confirmed to be free of A. citrulli (watermelon cv. ‘Zhaojia 84-24′) were placed in 50 mL of an A. citrulli Ac-1 bacterial suspension with a cell concentration of 107 CFU/mL−1. After incubation at 28 °C for 2 h, the seeds were removed and air dried for use as artificially infected seeds. Different amounts of artificially infected and healthy seeds were mixed to produce watermelon seedlots with infestation levels of 0.01–0.1%. For example, mixing 1 artificially infected seed with 999 healthy seeds would give a watermelon seedlot with 0.1% infestation. In addition, to verify the detection sensitivity of LAMP-FLD for a seed’s bacterial load, 20 g of healthy watermelon seeds (watermelon cv. ‘Zhaojia 84-24′) previously confirmed to be free from A. citrulli were immersed in A. citrulli Ac-1 bacterial suspensions with different cell concentrations (106 CFU/mL−1, 105 CFU/mL−1, 104 CFU/mL−1, 103 CFU/mL−1, 102 CFU/mL−1, 10 CFU/mL−1, 5 CFU/mL−1, and 1 CFU/mL−1). After incubation at 28 °C for 2 h, the seeds were removed and air-dried for use as artificially infected seeds with different bacterial loads. The bacterial load per gram of seeds (CFU/g−1) was calculated using the plate counting method described by Yan et al. [33] and Lu et al. [61].
Detection of A. citrulli in artificially infected seeds: Artificially infected seeds were immersed in an appropriate volume of ddH2O (e.g., 50 mL of ddH2O for 1000 seeds) and shaken at 28 °C and 200 rpm for 4 h. Then, the seeds and other residues were removed via filtering through two layers of sterile gauze to obtain the seed leachate. With reference to the methods described by Kan et al. [5] and Hui et al. [59], DNA in the seed leachate was extracted as a template for the LAMP-LFD assay of infected seeds, with sterile ddH2O used as the negative control and 1 ng/μL−1 A. citrulli Ac-1 genomic DNA used as the positive control. The procedure and observation methods for the LAMP-LFD assay were the same as those described in Section 2.5. When testing seeds with different infestation levels (%) and bacterial loads (CFU/g−1), each treatment was replicated 10 times. The positive detection rate was determined based on each test result. As long as one replicate was positive, the treatment result was considered positive.
Detection of A. citrulli in commercially available seeds: To evaluate the feasibility of using LAMP-LFD to detect A. citrulli in commercially available seeds, 30 watermelon/melon seedlots (including 18 watermelon seedlots and 12 melon seedlots) were purchased from a seed market in Fujian, with each seedlot containing at least 500 seeds. The seeds were not coated with bactericides. One sample was obtained from each seedlot by randomly selecting 100 seeds. The same detection methods used for the artificially infected seeds were used to extract the seed leachate DNA and perform the LAMP-LFD assay. Sterile ddH2O served as the negative control, and 1 ng/μL−1 A. citrulli Ac-1 genomic DNA served as the positive control. Each variety was tested five times in triplicate. As long as one result was positive, the sample was considered infected.
2.9. Detection of A. citrulli in Leaf Tissues
Collection of artificially infected and naturally diseased in-field leaf tissues: An A. citrulli Ac-1 bacterial suspension (1 × 108 CFU/mL−1) was inoculated via needle punch onto the true leaves of 14-day-old watermelon seedlings (watermelon cv. ‘Zhaojia 84-24′). At 7 d after inoculation, the symptomatic parts of diseased leaves were collected for use as artificially infected leaf tissues. In addition, diseased leaves suspected of BFB were collected from the field for use as naturally diseased in-field samples.
Detection of A. citrulli in leaves: The artificially infected leaf tissues and diseased in-field leaf tissues suspected of BFB were placed in separate mortars, frozen with liquid nitrogen, and ground to a fine powder. Genomic DNA was then extracted using a bacterial genomic DNA extraction kit (DP302-02) and detected using the LAMP-LFD method optimized in Section 2.5. Sterile ddH2O served as the negative control and 1 ng/μL−1 A. citrulli Ac-1 genomic DNA as the positive control.
3. Results
3.1. Optimization of A. citrulli LAMP-LFD Conditions and Establishment of Detection Method
The optimization results of the LAMP reaction temperature and time indicated that when the reaction time was 60 min, the LAMP reaction mixture showed a color change from orange to yellow-green at 62, 63, 64, and 65 °C (Figure 2a). Furthermore, a white turbid precipitate was produced, typical ladder-line DNA bands were observed following gel electrophoresis, and the LFD results were all positive at these temperatures (Figure 2b–d). Among them, the electrophoresis DNA bands were the brightest at 64 °C, while the LFD positive line was the clearest at 63 and 64 °C (Figure 2c,d). Given that non-specific amplification may occur if the temperature is too low and the target band may not be amplified if the temperature is too high, we selected 64 °C as the optimal reaction temperature.
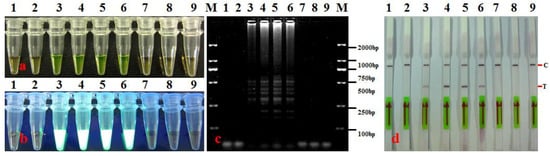
Figure 2.
Optimization of LAMP-LFD reaction temperature for detection of A. citrulli. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1–8: 60 °C, 61 °C, 62 °C, 63 °C, 64 °C, 65 °C, 66 °C, and 67 °C, respectively; 9: negative control (sterile water); M: DL 2000 DNA marker.
At the optimal reaction temperature, the optimization results for the reaction time indicated that the LAMP reaction showed positive results at 30 min, 45 min, 60 min, and 75 min (Figure 3). However, the electrophoresis DNA bands and LFD lines were the clearest at 60 min and 75 min (Figure 3c,d).
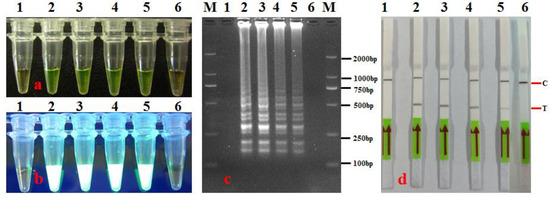
Figure 3.
Optimization of LAMP-LFD reaction time for detection of A. citrulli. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1: negative control (sterile water); 2–6: 75 min, 60 min, 45 min, 30 min, and 15 min, respectively; M: DL 2000 DNA marker.
By comprehensively comparing the amplification effects of different reaction temperatures and times, the optimal reaction conditions were ultimately determined to be 64 °C for 60 min. The LAMP-LFD was conducted in 25 μL, which included 12.5 μL of 2× RM, 1.0 μL each of 0.2 μM outer primers (F3 and B3) and 1.6 μM inner primers (FIP and BIP), 1.0 μL of Bst DNA polymerase (8 U·μL−1), 1.0 μL of fluorescent detection reagent FD, 1.0 μL of genomic DNA template, and sterile ddH2O to reach 25 μL. The results of testing through visual inspection, agarose gel electrophoresis, and the LFD assay were all in agreement (Figure 4).

Figure 4.
Detection of A. citrulli using optimized LAMP-LFD assay. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1–3: genomic DNA of A. citrulli; 4: negative control (sterile water); M: DL 2000 DNA marker.
3.2. LAMP-LFD Specificity
Genomic DNA from 6 strains of A. citrulli and 26 strains from the genera Acidovorax, Bacillus, Burkholderia, Escherichia, Lysinibacillus, Paenibacillus, Pseudomonas, Ralstonia, and Xanthomonas were used to analyze the specificity of the established LAMP-LFD method. The detection results (Table 1, Figure 5) indicate that only A. citrulli DNA was amplified. The amplification products turned yellow-green and showed a white turbid precipitate; agarose gel electrophoresis showed typical ladder-like bands; LFD showed clear quality control and test lines (Figure 5). In contrast, DNA from the other strains, genera, and ddH2O were negative. For example, strains such as A. delafieldii UK-1, A. caeni UK-1, A. avenae AS-1, A. avenae subsp. avenae AS-1, Pseudomonas syringae pv. actinidiae AC-1, Ralstonia solanacearum DE-1, and Xanthomonas axonopodis pv. citri CR-1 were all negative (Table 1). This implies that our established LAMP-LFD method was highly specific to A. citrulli and was able to distinguish between A. citrulli and strains of closely related species, achieving specific detection.
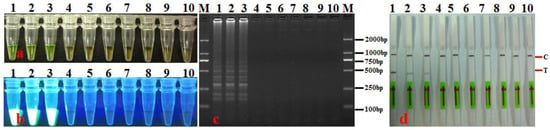
Figure 5.
Specificity of LAMP-LFD for detection of A. citrulli. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1: negative control (SDW); 1–3: A. citrulli Ac-1, A. citrulli Ac-4, A. citrulli Ac-6; 4–9: A. oryzae, A. delafieldi, A. caeni, Pseudomonas syringae pv. actinidiae, Xanthomonas axonopodis pv. citri, and Ralstonia solanacearum, respectively; 10: negative control (sterile water); M: DL 2000 DNA marker.
3.3. LAMP-LFD Sensitivity
The sensitivity of the established LAMP-LFD method was evaluated using a 10-fold gradient dilution series of A. citrulli genomic DNA. Our findings (Figure 6) revealed that when the concentration of A. citrulli genomic DNA was >1 fg·μL−1, the detection results were all positive, i.e., the LAMP product appeared yellow-green, a white turbid precipitate was observed under ultraviolet light, typical ladder-like bands appeared with gel electrophoresis, and quality control and test lines were clearly visible on the LFD assay (Figure 6b–d). This suggests that the detection sensitivity of the established LAMP-LFD method for A. citrulli genomic DNA was 1 fg·μ−1.
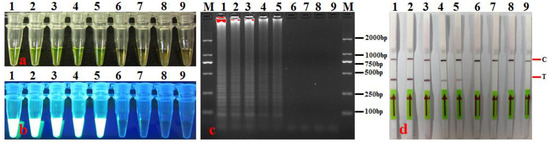
Figure 6.
Sensitivity of LAMP-LFD using serially diluted A. citrulli genomic DNA (μg.μL-1/fg·μL−1). (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1–8: 10 pg.μL−1, 1 pg·μL−1, 100 fg·μL−1, 10 fg·μL−1, 1 fg·μL−1, 100 ag.μL−1, 10 ag·μL−1, and 1 ag·μL−1, respectively; 9: negative control (SDW); M: DL 2000 DNA marker.
The original concentration of the Ac-1 bacterial suspension measured via plate counting was 9.4 × 107 CFU·mL−1 (approximately 1 × 108 CFU·mL−1, hereinafter referred to as 108 CFU·mL−1) (OD600 = 0.7). Template DNA was prepared using 1.0 mL each of Ac-1 bacterial suspension at concentrations of 107, 106, 105, 104, 103, 102, 50, 8, 4, 2, and 1 CFU·mL−1 and tested using the established LAMP-LFD assay. The results show that when the bacterial count was 8 CFU·mL−1, LAMP-LFD still yielded positive results, with negative results at lower bacterial counts (Figure 7). This implies that the detection sensitivity of the LAMP-LFD assay for A. citrulli was 8 CFU·mL−1. The triplicate results were all the same.
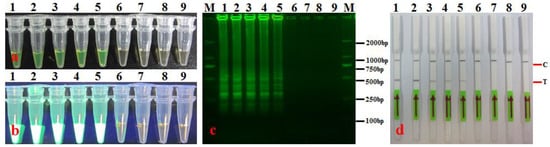
Figure 7.
Sensitivity of LAMP-LFD in detecting bacterial suspensions (CFU·mL-1) of A. citrulli. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick, C: control bank; T: test bank; 1–8: 104, 103, 102, 50, 8, 4, 2, and 1 CFU·mL−1 suspension, respectively; 9: negative control (SDW); M: DL 2000 DNA marker.
3.4. Artificially and Commercially Available Seeds Assay
Detection of A. citrulli in artificially infected seeds: DNA was extracted from the leachates of artificially infected seeds and assayed using LAMP-LFD. The results for artificially infected seeds from watermelon seedlots with different A. citrulli bacterial loads (CFU·g−1) and infection levels (%) that were tested using LAMP-LFD are shown in Figure 8 and Table 4. When the bacterial loads of the artificially infected seeds were 104, 103, 102, 101, and 5 CFU·g−1, the results were all positive (Figure 8, Table 4). When the infection level of watermelon seeds was 0.05–0.1%, the detection rate using LAMP-LFD was 20–100% (Table 4). The infection level of seeds was higher. The positive detection rate was correspondingly higher. The detection sensitivities (LODs) of LAMP-LFD for seeds with different bacterial loads (CFU·g−1) and infestation levels (%), therefore, were 5 CFU·g−1 and 0.05% (Table 4), respectively. These findings suggest that LAMP-LFD showed good repeatability and stability, even for seed samples with relatively low bacterial loads and infection levels.
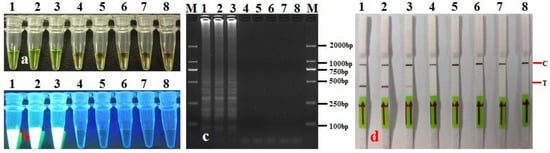
Figure 8.
Detecting A. citrulli from seed samples using LAMP-LFD assay. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1: artificially infested seeds; 2–6: commercially available seeds; 7: healthy seeds (disease-free seeds); 8: negative control (sterile water); M: DL 2000 DNA marker.
Table 4.
Results of LAMP-LFD for detecting A. citrulli from seeds with different bacterial cells (CFU· g 1) and different artificial infestation rates (%).
Detection of A. citrulli in commercially available seeds: Seedlots from 18 watermelon cultivars and 12 melon cultivars were randomly purchased at the market (sold by seed companies), and the leachates of these watermelon/melon seeds were used as templates for the LAMP-LFD assay. Five random samples were selected from each cultivar as replicates. The results (Figure 8, Table 2) indicate that 25 samples (14 watermelon cultivars and 11 melon cultivars) were negative, with a negative rate of 83.34%. Four watermelon cultivars (cv. ‘Xiaodelei’, cv. ‘Lanhanteida’, cv. ‘Hongxiaoyu’, and cv. ‘Lvlongbawang’) and one melon cultivar (cv. ‘Tianla 008’) were positive for A. citrulli, giving a positive rate of 16.66% (Table 2).
3.5. LAMP-LFD Validation with Leaf Samples
The LAMP-LFD assay was used to test artificially infected leaves and in-field leaves suspected of BFB. The results (Figure 9) show that all artificially infected leaves were positive, whereas healthy watermelon leaves not inoculated with A. citrulli were negative. Among the ten leaf samples collected from Xiapu, Ningde, two tested positive and eight tested negative (Table 5). All 12 leaf samples collected from Sanming were negative (Table 5). Among the eight leaf samples collected from Nanping, only one tested positive (Table 5). These findings indicate that some watermelon plants in Fujian were infected with A. citrulli while also demonstrating that the established LAMP-LFD method can be used to detect A. citrulli in plant leaves.
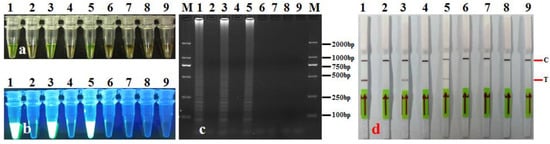
Figure 9.
Detecting A. citrulli from watermelon leaves using LAMP-LFD assay. (a) LAMP products were visualized with the naked eye under natural (normal) light (color change, yellow-green indicates positive reaction); (b) LAMP products were visualized with the naked eye under UV light (bright white turbidity indicates positive reaction); (c) LAMP products were analyzed using electrophoresis on 2% agarose gel (ladder-like pattern bands indicate positive reaction); (d) LAMP products were detected with lateral flow dipstick; C: control bank; T: test bank; 1: positive control (genomic DNA of A. citrulli); 2: negative control (sterile water); 3: artificially inoculated leaves; 4–9: leaves collected from fields; M: DL 2000 DNA marker.
Table 5.
Results of detecting A. citrulli in watermelon leaves using LAMP-LFD.
4. Discussion
The simple, rapid, and accurate detection of A. citrulli is critical for making decisions on whether to apply control measures before symptoms of BFB appear in cucurbit plants [6,18,32,35]. Using a combination of LAMP and LFD, we successfully designed primers and probes specific to A. citrulli, determined the optimal reaction temperature and time, and determined the specificity, sensitivity, and applicability of the LAMP-LFD assay to establish a rapid and accurate A. citrulli LAMP-LFD detection technique with a visual readout. This technique can be applied directly to rapidly diagnose BFB caused by infection with A. citrulli.
LAMP products can be determined by color change or UV observation of white precipitate methods. Although these methods are simple and visual, they have the following disadvantages: (i) LAMP detection methods involve the design of primers. If the quality of the designed primers is not ideal, dimers are easily generated during LAMP amplification. When the concentration of dimers reaches a certain amount, the detection results based on color change or UV methods will be positive, which can easily lead to false positives and incorrect detection results. (ii) Using DNA-intercalating dyes to induce color changes in LAMP amplification products can easily lead to aerosol deposition and false positives [43]. To overcome the above shortcomings, an LFD step for detecting LAMP amplification products was added in this study. An LFD has the following advantages: the probe (HP) of the LFD is designed based on conserved sites in the target gene sequence of the detection pathogen and highly matches the sequence of the corresponding site of the target gene. Positive reactions can only occur when the probe (HP) specifically hybridizes with LAMP products. The combination and confirmation of four LMAP amplification product detection methods, including color change, ultraviolet white precipitation observation, 2% agarose gel electrophoresis, and LFD, effectively overcome the false positive defect caused by aerosol deposition and dimers, and thus further improve the specificity and accuracy of the LAMP detection method.
The selection of target genes and the design and use of specific primers are the keys to developing assays for the accurate detection of pathogens. Most primers used in PCR-based A. citrulli detection techniques were designed based on the 16S rDNA sequence [2,60]. However, sequence analysis has shown that there is a high level of sequence homology (up to 90%) between the genera Acidovorax and Pseudomonas and between the various subspecies within the genus Acidovorax. Thus, it is not possible to distinguish among closely related species based on this target gene [62,63,64,65,66]. Tian et al. [40] designed A. citrulli-specific primers targeting the hrpB2 gene, HB2F2/HB2R2, which were able to effectively distinguish A. citrulli from A. cattleyae, A. konjaci, and A. facilis. However, a specific 290 bp fragment could also be detected in some A. avenae strains. Hence, the specificity of these primers required further improvement [40]. The YD-repeat protein (Aave_2049) is one of the rhs genes, and there are many homologous genes among bacteria in the family Enterobacteriaceae. There is a certain degree of conservation in the YD-repeat protein gene sequences between different species [67]. A real-time fluorescent, quantitative PCR targeting the YD-repeat protein gene has been successfully applied for the detection of A. citrulli [30]. The results of this study once again confirm that primers and probes designed based on conserved sites (specific sites) of the YD-repeat protein gene have high specificity for A. citrulli, thus ensuring the accuracy of the results in detecting A. citrulli. In addition to the effects of primer and probe conservation on the specific detection of pathogens, the optimization of reaction conditions can also have a crucial impact on specificity. Under suboptimal reaction conditions, primers and probes may hybridize with non-target pathogens, thereby producing false positives. Therefore, the optimization of reaction conditions is vital to ensuring high detection specificity [68,69]. In this study, specific primers and probes were designed based on conserved loci in the YD-repeat protein gene sequence of the BFB pathogen, and the reaction conditions (i.e., temperature and time) were optimized, with the aim of establishing a LAMP-LFD assay for the BFB pathogen. The optimal reaction conditions for LAMP-LFD were 64 °C and 60 min. The results of an examination of specificity indicated that the designed LAMP primers and probes only amplified A. citrulli DNA, with no amplification signals observed in non-target strains, including five strains that are closely related to A. citrulli (i.e., A. delafieldii UK-1, A. caeni UK-1, A. avenae AS-1, A. avenae subsp. avenae AS-1, and A. oryzae OS-1). These findings suggest that the established A. citrulli LAMP-LFD method showed high specificity. Hence, this method can be used to effectively distinguish A. citrulli from other bacteria.
Detection sensitivity is another key indicator for evaluating the applicability of different detection techniques. A higher sensitivity or lower LOD indicates that this technique has greater applicability [70,71]. There are differences in the sensitivities of different nucleic acid detection methods for A. citrulli. Song et al. [72] performed immuno-concentration PCR and direct PCR for the detection of BFB pathogens. They found that for purified bacterial suspensions, both techniques achieved a detection sensitivity of 3 × 104 CFU/mL; for unpurified seed suspensions, the detection sensitivity of direct PCR was 3 × 105 CFU/mL, whereas that of immuno-concentration PCR was still 3 × 104 CFU/mL. Zhou et al. established a ddPCR technique capable of simultaneously detecting A. citrulli and Pseudomonas syringae pv. lachrymans that demonstrated a detection sensitivity of 1 pg/μL [34]. Based on the hrpB2 and ITS sequences, Nurhan and Hüseyin established an A. citrulli locked nucleic acid real-time PCR assay with an LOD of 12 fg/μL [73]. Lu et al. found an LOD for ddPCR of A. citrulli of 2 fg/μL−1 and a sensitivity 10-fold higher than that of real-time quantitative PCR [61]. Yan et al. used the non-ribosomal peptide synthetase (NRPS) gene to establish an A. citrulli LAMP assay and reported a detection sensitivity of 1 fg/μL, which was 100-fold higher than that of conventional PCR [33]. By targeting the YD-repeat protein gene, Cho et al. established an A. citrulli–SYBR Green-based real-time quantitative, fluorescent PCR technique with a detection sensitivity of 5 fg/μL for A. citrulli genomic DNA, which was equivalent to 6.5 CFU/mL−1 [38]. Tian et al. designed primers based on the 16S-23S internal transcribed spacer ribosomal DNA sequence and established an A. citrulli padlock probe assay with a detection sensitivity of 100 fg/μL−1 [40]. Ha et al. combined quantitative PCR with magnetic capture hybridization to achieve the simultaneous detection of two pathogens, A. avenae subsp. citrulli and Didymella bryoniae, among which the detection sensitivity for A. avenae reached 10 CFU/mL−1 [74]. Bahar et al. designed a pair of primers, BX-L1/BX-S-R2, and, for a PCR assay based on these primers, reported a relatively high specificity and sensitivity for A. citrulli: in a real-time assay of melon seed samples, A. citrulli could be detected if more than 0.02% of 5000 seeds were infected [35]. Compared to the methods mentioned above, the detection sensitivity of our A. citrulli LAMP-LFD assay for genomic DNA, the bacterial suspension, seed bacterial load, and seed infection level were 1 fg/μL−1, 8 CFU/mL−1, 5 CFU/g−1, and 0.05%, respectively. These findings are 100–10,000-fold higher than the sensitivity of A. citrulli immuno-concentration PCR [72], conventional PCR [63], nested PCR [4], ddPCR [34], and padlock probe PCR [40] assays previously established by researchers. The method of extracting bacterial genomic DNA plays an important role in improving the sensitivity of pathogen detection. Optimizing and improving the extraction method of bacterial genomic DNA are some of the ways to enhance the detection sensitivity. Under the same concentration of bacterial suspension, excellent or advanced genomic DNA extraction methods can obtain DNA with a better quality and higher concentration, which is crucial for improving the detection sensitivity. For example, the genomic DNA extracted from a bacterial suspension of 6.5 CFU·mL−1 was 5 fg·μL−1 using the method of Cho et al. [38]. However, the genomic DNA from a bacterial suspension of 8 CFU·mL−1 was only 1.0 fg·μL−1 in this study. Obviously, the bacterial genomic DNA extraction method adopted by Cho et al. [38] is more effective than the method used in this study. Moreover, the sensitivity of a plant pathogen detection assay is not only closely related to the detection method, reaction temperature, reaction time, and genomic DNA extraction methods but also influenced by factors such as the selection of the target gene, reagent concentration, primers, and probe sequences [49,71,74]. Therefore, in order to further improve the sensitivity of the A. citrulli LAMP-LFD assay, it is necessary to re-optimize and combine the above-mentioned factors in future research.
BFB caused by A. citrulli is a typical seedborne disease, and infected seeds are the most important source of primary infection in fields. Therefore, stringent seed testing is a critical step in the prevention and control of this disease [13,75]. With the developed LAMP-LFD assay, artificially infected seeds with a concentration of A. citrulli greater than 5 CFU/g−1 tested positive, while those with a bacterial load less than 5 CFU/g−1 tested negative. Furthermore, positive results were obtained with a seed infestation level of 0.05% and above, although the frequency of positive results was less when the infection levels were low. These results are consistent with those reported by Walccot et al. [60]. This may be because due to interference from other bacteria or inhibitory compounds, the seed bacterial load may not have been sufficient for the amount of DNA template required for detection using the LAMP-LFD assay, which may limit the application of the A. citrulli LAMP-LFD assay in seed phytosanitary detection [76,77]. Therefore, further exploration is needed to determine whether the target bacteria should first be enriched via selective plating to detect A. citrulli in seeds to ensure the accuracy of the detection results [72,78]. Moreover, in the detection of A. citrulli in seeds, the sample size and number of replicates should be increased as much as possible. Apart from excellent specificity and sensitivity, the practicality and convenience of a detection method can also play a decisive role in its actual application. The LAMP-LFD method established in this study can be used to detect the presence of A. citrulli in seeds and leaf tissues, which suggests that this is a feasible and practical technique for the detection of A. citrulli. Our study demonstrated that LAMP-LFD can be used for the rapid, sensitive, and visually readable detection of phytopathogens.
5. Conclusions
In this study, primers and probes targeting the YD-repeat protein gene were designed for the specific detection of A. citrulli, and LAMP was combined with LFD to establish an A. citrulli LAMP-LFD assay. This assay showed a relatively high specificity and sensitivity for A. citrulli, was able to rapidly and accurately detect A. citrulli in seeds and leaf tissues, and showed good applicability. In conclusion, the A. citrulli LAMP-LFD assay established in this study offers several advantages, including easy operation, rapidity, high specificity and sensitivity, low cost (no need for complex and expensive precision instruments), visualization of detection results, good stability, and strong applicability. Therefore, it can be employed in the rapid detection of A. citrulli, thereby providing technical support for the accurate diagnosis and timely adoption of control measures for BFB to prevent the spread of this disease.
Author Contributions
Conceptualization (designed the study), C.L. and X.Y.; Methodology (performed the experiments), C.L., M.L., X.L. and Y.D.; Data curation (date analysis and discussion), C.L., M.L., L.G. and Y.D.; Software, C.L., L.G., M.H., H.R. and Y.D.; Visualization, X.Y.; Writing—original draft preparation, C.L., M.L. and X.Y.; Formal analysis: M.H. and H.R.; Funding acquisition and Project administration, X.Y. and L.G.; Investigation, C.L. and M.L.; Resources, C.L. and X.Y.; Writing—review and editing, C.L., M.L. and X.Y.; Supervision, X.Y.; Validation, M.L. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by funds from the National Key Research and Development Program of China (2021YFC2600402), the Agricultural High-Quality Development “5511” Collaborative Innovation Project of the Fujian Provincial People’s Government—Chinese Academy of Agricultural Sciences (XTCXGC2021011), and the Science and Technology Innovation Foundation of the Fujian Academy of Agricultural Sciences (CXTD2021002-1).
Data Availability Statement
Sequence data from this article can be found in GenBank at https://www.ncbi.nlm.nih.gov/nuccore/CP029373.1/ (accessed on 5 June 2019) with the accession number CP029373.0.
Acknowledgments
The authors would like to thank Xueqing Cai (College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou, China) and Yixuan Xie (Plant Protection and Phytosanitary Station of Xiamen, Xiamen, China) for kindly providing the Acidovorax citrulli strains. Also, we would like to thank the Xiapu Huajiu Family Farm for providing the watermelon and muskmelon leaf samples.
Conflicts of Interest
The authors confirm that there are no conflicts of interest for this manuscript and agree to the submission of this paper.
References
- Crall, J.M.; Schenck, N.C. Bacterial fruit rot of watermelon in Florida. Plant Dis. Rep. 1969, 53, 74–75. [Google Scholar]
- Walcott, R.R.; Gitaitis, R.D. Detection of Acidovorax avenae subsp. citrulli in watermelon seed using immunomagnetic separation and the polymerase chain reaction. Plant Dis. 2000, 84, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.L.; Xu, J.; Zhao, Y.Q.; Li, X.H.; Hu, B.S.; Liu, F.Q. Specific detection of Acidovorax avenae subsp. citrulli by PCR. Jiangsu J. Agric. Sci. 2010, 26, 512–516. [Google Scholar]
- Wang, J.; Bi, Y.; Zhu, Y.; Han, S.Y.; Zhu, X.; Sheng, W.J.; Li, M. Nested-PCR rapidly detect Acidovorax avenae subsp. citrulli from watermelon seeds. Sci. Agric. Sin. 2014, 47, 284–291. [Google Scholar]
- Kan, Y.M.; Yun, X.M.; Li, Y.W.; Li, J.Q.; Lou, L.X. Screening of specific primers for detection of Acidovorax citrulli from cucurbits seed by Bio-PCR. Acta Phytopath. Sin. 2018, 48, 263–270. [Google Scholar]
- Husni, A.A.A.; Ismail, S.I.; Jaafar, N.; Zulperi, D. Etiology, diagnostic approaches and management strategies of Acidovorax citrulli, a bacterial fruit blotch pathogen of cucurbits. Plant Prot. Sci. 2021, 57, 75–94. [Google Scholar] [CrossRef]
- Takashi, S. Studies on control of seed borne bacterial vegetable disease: Studies on epidemiology and control of cucurbits bacterial fruit blotch. J. Gen. Plant Pathol. 2021, 87, 408–412. [Google Scholar]
- Cameron Somodi, C.; Jones, J.B.; Hopkins, D.L.; Stall, R.E.; Kucharer, T.A.; Hodge, N.C.; Watterson, J.C. Occurrence of a bacterial watermelon fruit blotch in Florida. Plant Dis. 1991, 75, 1053–1056. [Google Scholar] [CrossRef]
- Latin, R.X.; Hopkins, D.L. Bacterial fruit blotch of watermelon. Plant Dis. 1995, 79, 761–765. [Google Scholar] [CrossRef]
- Cai, X.Q.; Huang, Y.Y.; Yang, J.Z.; Chen, J.; Cai, G.L.; Hu, F.P. Pathogen identification of bacterial fruit blotch of watermelon in Fujian. J. Fujian Agric. For. Univ. 2005, 34, 434–437. [Google Scholar]
- Burdman, S.; Walcott, R. Acidovorax citrulli: Generating basic and applied knowledge to tackle a global threat to the cucurbit industry. Mol. Plant Pathol. 2012, 13, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Toshio, M.; Nakayama, E.; Nanaka, N.; Tabuchi, M. Host-specific activation of a pathogen effector Aave_4606 from Acidovorax citrulli, the causal agent for bacterial fruit blotch. Biochem. Biophys. Res. Commun. 2022, 616, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.L.; Thompson, C.M. Seed transmission of Acidovarax avenae subsp. citrulli in cucurbits. HortScience 2002, 37, 924–926. [Google Scholar] [CrossRef]
- Burdman, S.; Kots, N.; Kritzman, G.; Kopelowitz, J. Molecular, physiological, and host-range characterization of Acidovorax avenae subsp. citrulli isolates from watermelon and melon in Israel. Plant Dis. 2005, 89, 1339–1347. [Google Scholar] [CrossRef]
- Wu, P.Y.; Ho, L.C.; Chang, J.J.; Tzeng, K.C.; Deng, W.L.; Lin, Y.H. Development of a TaqMan probe-based insulated isothermal PCR (TiiPCR) for the detection of Acidovorax citrulli, the bacterial pathogen of watermelon fruit blotch. Eur. J. Plant Pathol. 2017, 147, 869–875. [Google Scholar] [CrossRef]
- Feng, J.J.; Li, J.Q.; Wacott, R.R.; Zhang, G.M.; Luo, L.X.; Kang, L.; Zheng, Y.; Schaad, N.W. Advances in detection of Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbits. Seed Sci. Technol. 2013, 41, 1–15. [Google Scholar] [CrossRef]
- Tian, Q.; Feng, J.J.; Hu, J.; Zhao, W.J. Selective detection of viable seed-borne Acidovorax citrulli by real-time PCR with propidium monoazide. Sci. Rep. 2016, 6, 35457. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Hossain, R.H.; Kim, H.T.; Jesse, D.M.I.; Abuyusuf, M.; Jung, H.J.; Park, J.I.; Nou, I.S. Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon. Int. J. Mol. Sci. 2019, 20, 2715. [Google Scholar] [CrossRef]
- Fei, N.Y.; Chen, H.M.; Yang, Y.W.; Guan, W.; Liu, B.Y.; Zhao, T.C. Advances of cucurbit bacterial fruit blotch abroad. China Cucurb. Veget. 2022, 35, 1–5. [Google Scholar]
- Wang, Z.L.; Cheng, W.H.; Dong, Z.Y.; Yao, X.M.; Deng, X.; Ou, C. A CCRSPR/LbCas12a-based method for detection of bacterial fruit blotch pathogens in watermelon. Microbiol. Spectr. 2024, 12, 1–11. [Google Scholar]
- Schaad, N.W.; Postnikova, E.; Sechler, A.; Claflin, L.E.; Vidaver, A.K.; Jones, J.B.; Agarkova, I.; Ignatov, A.; Dickstein, E.; Ramundo, B.A. Reclassification of subspecies of Acidovorax avenae as A. avenae (Manns 1905) emend., A. cattleyae (Pavarino 1911) comb. nov., A. citrulli (Schaad et al., 1978) comb. nov., and proposal of A. oryzae sp. nov. Syst. Appl. Microbiol. 2008, 31, 434–446. [Google Scholar] [CrossRef]
- Zhao, T.; Feng, J.J.; Sechler, A. An Improved Assay for detection of Acidovorax avenae subsp. citrulli in watermelon and melon seed. Seed Sci. Technol. 2009, 37, 337–349. [Google Scholar] [CrossRef]
- Xiong, L.B.; Liu, Q.; Wang, T.C.; Gao, L.P.; Wang, J.; Liu, S.T.; Song, R.; Shi, Y.B.; Wang, J.P.; Wen, Z.H. An improved DAS Dot ELISA method for detection of Acidovorax avenae subsp. citrulli. Microbiol. China 2010, 37, 1551–1556. [Google Scholar]
- Himananto, O.; Thummabenjapone, P.; Luxananil, P. Novel and highly specific monoclonal antibody to Acidovorax citrulli and development of ELISA-based detection in cucurbit leaves and seeds. Plant Dis. 2011, 95, 1172–1178. [Google Scholar] [CrossRef]
- Charlermroj, R.; Himananto, O.; Seepiban, C. Multiplex detection of plant pathogens using a microsphere immunoassay technology. PLoS ONE 2013, 8, e62344. [Google Scholar] [CrossRef]
- Charlermroj, R.; Makornwattana, M.; Himananto, O.; Seepidan, C.; Phuengwas, S.; Warin, N.; Gajanandana, O.; Karoonuthaisiri, K. An accurate, specific, sensitive, high-throughput method based on a microsphere immunoassay for multiplex detection of three viruses and bacterial fruit blotch bacterium in cucurbits. J. Virol. Methods 2017, 247, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hu, J. Preliminary research on serological method for detecting fruit blotch bacterium in hemi melon seed. J. Inner Mong. Agric. Univ. 2005, 26, 20–23. [Google Scholar]
- Puttharugsa, C.; Wangkam, T.; Huangkamhang, N.; Gajanandana, O.; Himananto, O.; Sutapun, B.; Amarit, R.; Somboonkaew, A.; Srikhirin, T. Development of surface plasmon resonance imaging for detection of Acidovorax avenae subsp. citrulli (Aac) using specific monoclonal antibody. Biosens. Bioelectron. 2011, 26, 2341–2346. [Google Scholar] [CrossRef]
- Kuo, S.Y.; Lin, Y.C.; Lai, Y.C. Production of fluorescent antibody-labeling proteins in plants using a viral vector and the application in the detection of Acidovorax citrulli and Bamboo mosaic virus. Pub. Libr. Sci. 2018, 13, e0192455. [Google Scholar] [CrossRef]
- Saisin, L.; Amarit, R.; Somboonkaew, A. Significant sensitivity improvement for camera-based lateral flow immunoassay readers. Sensors 2019, 18, 4026. [Google Scholar] [CrossRef]
- Venette, J.R.; Lamppa, R.S.; Albaugh, D.A.; Nayes, J.B. Presumptive procedure (dome test) for detection of seedborne bacterial pathogens in dry beans. Plant Dis. 1987, 71, 984–990. [Google Scholar] [CrossRef]
- Zhao, T.C.; Sun, F.Z.; Wang, B.W.; Hui, W.G. Pathogen identification of hami melon bacterial fruit blotch. Acta Phytopathol. Sin. 2001, 31, 357–364. [Google Scholar]
- Yan, L.C.; Zhao, Y.Q.; Zhou, J.J.; Chen, S.; Bai, S.; Tian, Y.L.; Gong, W.R.; Hu, B.S. Rapid and sensitive detection of Acidovorax citrulli in cucurbit seeds by visual loop-mediated isothermal amplification assay. J. Phytopath. 2019, 167, 10–18. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Lu, Y.; Tian, W.; Wang, Y.X.; Wen, C.L.; Li, J.Q.; Xu, X.L.; Luo, L.X. Detection of Acidovorax citrulli and Pseudoonas syringae pv. lachrymans from cucurbit seeds by multiplex droplet digital PCR. Plant Prot. 2021, 47, 156–163. [Google Scholar]
- Bahar, O.; Efrat, M.; Hadar, M. New subspecies-specific polymerase chain reaction-based assay for the detection of Acidovorax avenae subsp. citrulli. Plant Pathol. 2008, 57, 754–763. [Google Scholar] [CrossRef]
- Oya, H.; Nakagawa, H.; Saito, N.; Uematsu, H.; Ohara, T. Detection of Acidovorax avenae subsp. citrulli from seed using LAMP method. Jpn. J. Phythol. 2008, 74, 304–310. [Google Scholar]
- Yang, G.; Erdman, D.; Tondella, M.L.; Fields, B.S. Evaluation tetramethylrhodamine and black hole quencher 1 labeled probes and five commercial amplification mixes in TaqMan® real-time RT-PCR assays for respiratory pathogens. J. Virol. Methods 2009, 162, 288–290. [Google Scholar] [CrossRef]
- Cho, M.S.; Park, D.H.; Ahn, T.Y.; Park, D.S. Rapid and specific detection of Acidovorax avenae subsp. citrulli using SYBR green-based real-time PCR amplification of the YD-repeat protein gene. J. Microbiol. Biotechnol. 2015, 25, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Slovareva, O.Y.; Starikova, E.V. A novel qPCR-based test system for Acidovorax citrulli based on the PAS domain S-Box protein gene. Mol. Genet. Microbiol. Virol. 2021, 36, 100–103. [Google Scholar] [CrossRef]
- Tian, Y.L.; Zhao, Y.Q.; Bai, S.; Walcott, R.R.; Hu, B.S.; Liu, F.Q. Reliable and sensitive detection of Acidovorax cirulli in cucurbit seed using padlock-probe-based assay. Plant Dis. 2013, 97, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Xu, F.S.; Zhao, L.H.; Xie, G.L. Immuno-capture PCR method for detecting Acidovorax avenae subsp. citrulli from watermelon. Chin. J. Agric. Biotechnol. 2007, 4, 173–179. [Google Scholar]
- Zhang, J.; Tian, Q.; Zhu, S.F.; Zhao, W.J.; Liu, F.Q. Rapid on-sit detection of Acidovorax citrulli by cross-priming amplification. Mol. Cell. Probes 2012, 26, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.Z.; Gan, L.; Dai, Y.L.; Liu, X.F.; Yang, X.J. Development of loop-mediated isothermal amplification (LAMP) assay for specific and sensitive detection of Mycocentrospora acerina (Hart.) causing round leaf spot disease in Sanqi (Panax notogenseng). Horticulturae 2022, 18, 1060. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rigano, L.A.; Malamud, F.; Orce, I.G.; Filippone, M.P.; Marano, M.R.; Amaral, A.D.; Castagnaro, A.P.; Vojnov, A.A. Rapid and sensitive detection of Candidatus Liberibacter asiaticus by loop mediated isothermal amplification combined with a lateral flow dipstick. BMC Microbiol. 2014, 14, 86. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wang, R.N.; Zhou, Q.J.; Chen, J.R. Rapid Detection of Vibrio vulnificus by Loop-mediated Isothermal Amplification Combined with Lateral Flow Dipstick Assay. Biotechnol. Bull. 2014, 6, 81–87. [Google Scholar]
- Sridapan, T.; Tangkawsakul, W.; Janvilisri, T.; Kiatpathomchai, W.; Dangtip, S.; Ngamwongsatit, N.; Nacapricha, D.; Ounjai, P.; Chankhamhaengdecha, S. Rapid detection of Clostridium perfringens in food by loop-mediated isothermal amplification combined with a lateral flow biosensor. PLoS ONE 2021, 16, e0245144. [Google Scholar] [CrossRef]
- Liu, W.W.; Lu, G.; Wang, Y.; Cheng, Z.H.; Gao, Y.Y.; Yin, Z.P.; Wu, Y.; Lv, X.Q.; Guo, P.B.; Zhao, Y.H. A novel loop-mediated isothermal amplification-lateral flow dipstick method for Helicobacter pylori detection. Front. Microbiol. 2023, 14, 1094600. [Google Scholar] [CrossRef]
- Peng, Q.D.; Ning, J.C.; Xu, Q.Y.; Yang, T.; Wang, Y.R.; Zheng, T.R.; Zhuang, Q.G.; Xi, D.H. Development and application of a reverse transcription loop-mediated isothermal amplification combined with lateral flow dipstick for rapid and visual detection of Citrus leaf blotch virus in kiwifruit. Crop Prot. 2021, 143, 105555. [Google Scholar] [CrossRef]
- Kim, N.K.; Lee, H.J.; Kim, S.M.; Jeong, R.D. Rapid and visual detection of barley yellow dwarf virus by reverse transcription recombinase polymerase amplification with lateral flow strips. Plant Pathol. J. 2022, 38, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.S.; Brennan, M.S.; Khan, A.; Ali, G.S. Implementation of loop-mediated isothermal amplification methods in lateral flow devices for the detection of Rhizoctonia solani. Can. J. Plant Pathol. 2015, 37, 118–129. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, H.L.; Li, X.J.; Shan, C.L.; Li, X.S.; Chen, Y.; Shao, W.D.; Zhu, P. Establishment of Leptosphaeria maculans detection by loop-mediated isothermal amplification combined with a lateral flow dipstick. Plant Quar. 2016, 30, 32–37. [Google Scholar]
- Jiang, L.; Gu, R.; Li, X.; Mu, D. Simple and rapid detection Aspergillus fumigatus by loop-mediated isothermal amplification coupled with lateral flow biosensor assay. J. Appl. Mcriobiol. 2021, 131, 2352–2360. [Google Scholar] [CrossRef]
- Cai, Y.; Zhou, Q.J.; Gu, J.F.; Chen, X.F.; Chen, J. Rapid and Sensitive Detection of Meloidogyne camelliae by LAMP-LFD. J. Agric. Biotechnol. 2016, 24, 770–780. [Google Scholar]
- Zhou, Q.J.; Cai, Y.; Gu, J.F.; Wang, X.; Chen, J. Rapid and sensitive detection of Meloidogyne mali by loop-mediated isothermal amplification combined with a lateral flow dipstick. Eur. J. Plant Pathol. 2017, 148, 755–769. [Google Scholar] [CrossRef]
- Sun, P.P.; Shi, J.L.; Peng, Z.; Wang, S.; Wang, Y.; Wu, X.Y.; Xu, S.J.; Li, J. Rapid and sensitive diagnosis of Mycoplasma hyopneumoniae by loop-mediated isothermal amplification combined with a lateral flow dipstic. Acta Vet. Zootech. Sin. 2020, 51, 1419–1428. [Google Scholar]
- Han, X.T.; Zhao, T.; Yan, T.; Yu, R.C. Rapid and sensitive detection of Karenia mikimotoi by loop-mediated isothermal amplification combined with lateral flow dipstick. Environ. Sci. Pollut. Res. 2022, 29, 24696–24703. [Google Scholar] [CrossRef]
- Hui, W.G.; Zhao, T.C.; Schaad, N.W.; Sun, F.Z.; Wang, J.R. Establishment of rapid detection method for the pathogen of hami melon fruit blotch. Sci. Agric. Sin. 2007, 10, 2495–2501. [Google Scholar]
- Walcott, R.R.; Castro, A.C.; Fessehaie, A.; Ling, K. Progress towards a commercial PCR-based seed assay for Acidovorax avenae subsp. citrulli. Seed Sci. Technol. 2006, 34, 101–116. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, H.J.; Zhao, Z.J.; Wen, C.L.; Wu, P.; Song, S.H.; Yu, S.C.; Lou, L.X.; Xu, X.L. Application of droplet digital PCR in detection of seed-transmitted pathogen Acidovorax citrulli. J. Integr. Agric. 2020, 19, 561–569. [Google Scholar]
- Walcott, R.R.; Langston, D.B.; Sanders, J.F.H.; Gitatitis, R.D. Investigating intraspecific variation of Acidovorax avenae subsp. citrulli using DNA fingerprinting and whole cell fatty acid analysis. Phytopathology 2000, 90, 191–196. [Google Scholar] [CrossRef]
- Ren, Y.Z.; Li, H.; Li, G.Y.; Wang, X.D.; Wan, G.; Fang, L. Detection of Acidovorax avenae subsp. citrulli in melon seed using the polymerase chain reaction. Xinjiang Agric. Sci. 2004, 41, 329–332. [Google Scholar]
- Zhang, X.L.; Wu, Y.M.; Wang, C.; Li, B.J. Sequence analysis of 16S rDNA and specified primers design of Acidovorax avenae subsp. citrulli. Acta Phytopathol. Sin. 2007, 37, 225–231. [Google Scholar]
- Eckshtain-Levi, N.; Shkedy, D.; Gershovits, M.; Da Silva, G.M.; Tamir-Ariel, D.; Walcott, R.; Pupko, T.; Burdman, S. Insights from the genome sequence of Acidovorax citrulli M6, a group I strain of the causal agent of bacterial fruit blotch of cucurbits. Front. Microbiol. 2016, 7, 430. [Google Scholar]
- Song, J.Y.; Oo, M.M.; Park, S.Y.; Seo, M.W.; Lee, S.C.; Jeon, N.B.; Nam, M.H.; Lee, Y.S.; Kim, H.G.; Oh, S.K. Analysis of intraspecific genetic diversity in Acidovorax citrulli causing bacterial fruit blotch on cucurbits in Korea. Korean J. Agric. Sci. 2018, 45, 575–582. [Google Scholar]
- Jackson, A.P.; Thomas, G.H.; Parkhill, J.; Thomson, N.R. Evolutionary diversification of an ancient gene family (rhs) through C-terminal displacement. BMC Genom. 2009, 10, 584. [Google Scholar] [CrossRef]
- Innis, M.A.; Gelfand, D.H. Optimization of PCR’s. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 3–12. [Google Scholar]
- Infantino, A.; Pucci, N. A PCR-based assay for the detection and identification of Pyrenochaeta lycopersici. Eur. J. Plant Pathol. 2005, 112, 337–347. [Google Scholar] [CrossRef]
- Zeng, H.J.; Zhang, D.Q.; Zhai, X.Z.; Wang, S.J.; Liu, Q. Enhancing the immunofluorescent sensitivity for detection of Acidovorax citrulli using fluorescein isothiocyanate labeled antigen and antibody. Anal. Bioanal. Chem. 2018, 410, 71–77. [Google Scholar]
- Wang, R.B.; Chen, S.Z.; Zhao, Y.M.; Li, B.J.; Liu, P.Q.; Chen, Q.H. Development of a recombinase polymerase amplification-lateral flow dipstick assay for rapid detection of the taro leaf blight pathogen Phytophthora colocasiae. J. Plant Prot. 2022, 49, 1654–1662. [Google Scholar]
- Song, Q.L.R. Rapid detection of seed borne Acidovorax avenae subsp. citrulli by “pathogen immune-concentration” PCR. J. Plant Pathol. 2009, 90, 600. [Google Scholar]
- Nurhan, Ö.; Hüseyi, B. A real-time PCR assay using locked nucleic acid probe for detection of Acrdovarax citrulli. J. Plant Dis. Prot. 2022, 129, 395–409. [Google Scholar]
- Ha, Y.; Fessehaie, A.; Ling, K.S. Simultaneous detection of Acidovarax avenae subsp. citrulli and Didymella bryoniae in cucurbit seedlots using magnetic capture hybridization and real-time polymerase chain reaction. Phytopathology 2009, 99, 666–678. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kirshner, B.; Kritzman, G. Sweat boxes and selective media for the detection of Acidovorax avenae subsp. citrulli in melon and watermelon seeds. Phytoparasitica 2008, 36, 134. [Google Scholar]
- DeBoer, S.H.; Ward, L.J.; Li, X.; Chittaranjan, S. Attenuation of PCR inhibition in the presence of plant compounds by addition of Blotto. Nucleic Acids Res. 1995, 23, 2567–2568. [Google Scholar] [CrossRef] [PubMed]
- Kontanis, E.J.; Reed, F.A. Evaluation of real-time PCR amplification efficiencies to detect PCR inhibitors. J. Forensic Sci. 2006, 51, 795–804. [Google Scholar] [CrossRef]
- Ozakman, M.; Schaad, N.W. A real-time BIO-PCR assay for detection of Ralstonia solanacearum race 3, biovar 2, in asymptomatic potato tubers. Can. J. Plant Pathol. 2003, 25, 232–239. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).