

Toward the Development of Garlic Varieties: The First Attempts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Statements for the Field Trials
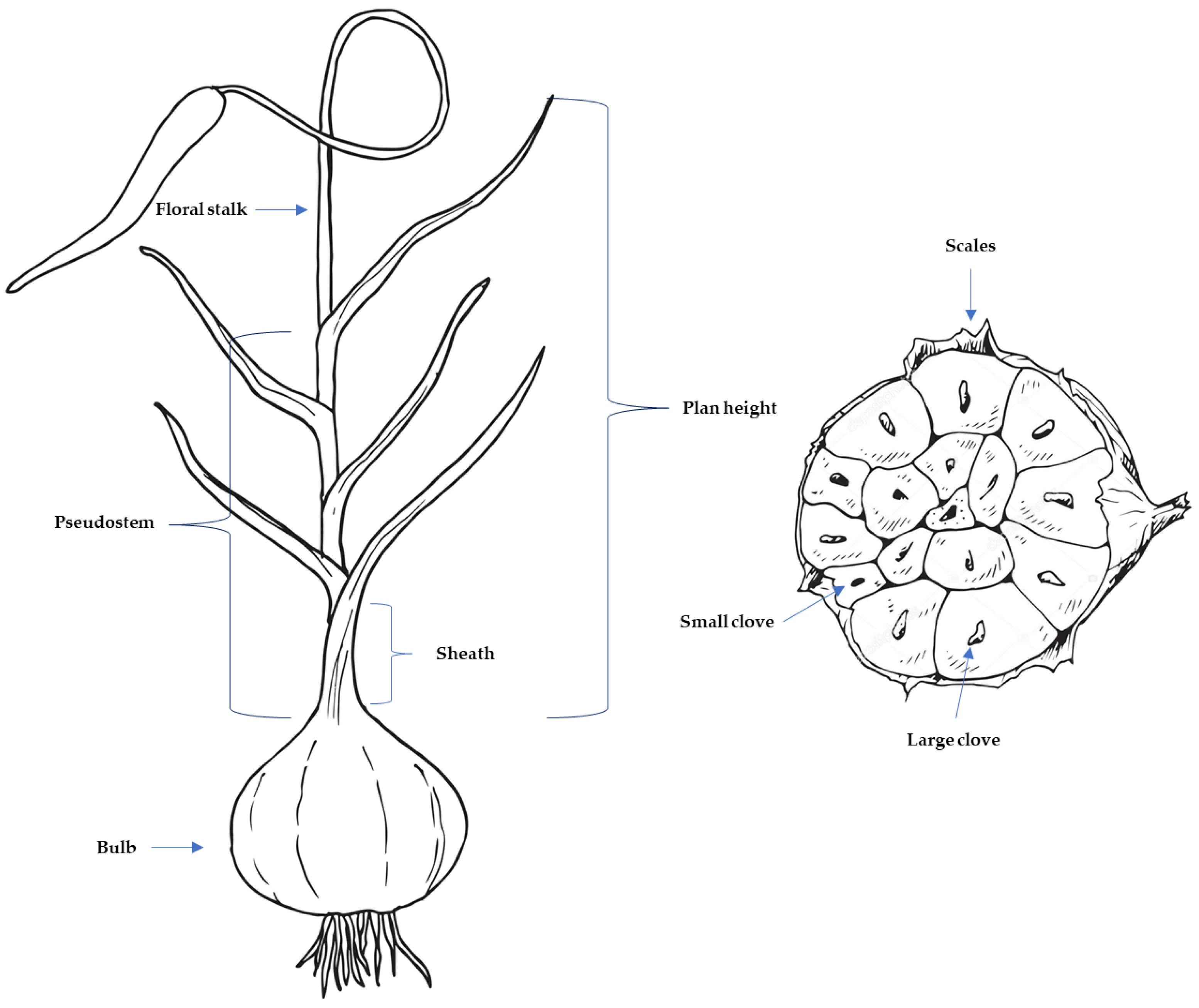
2.2. Phenotypic Selection
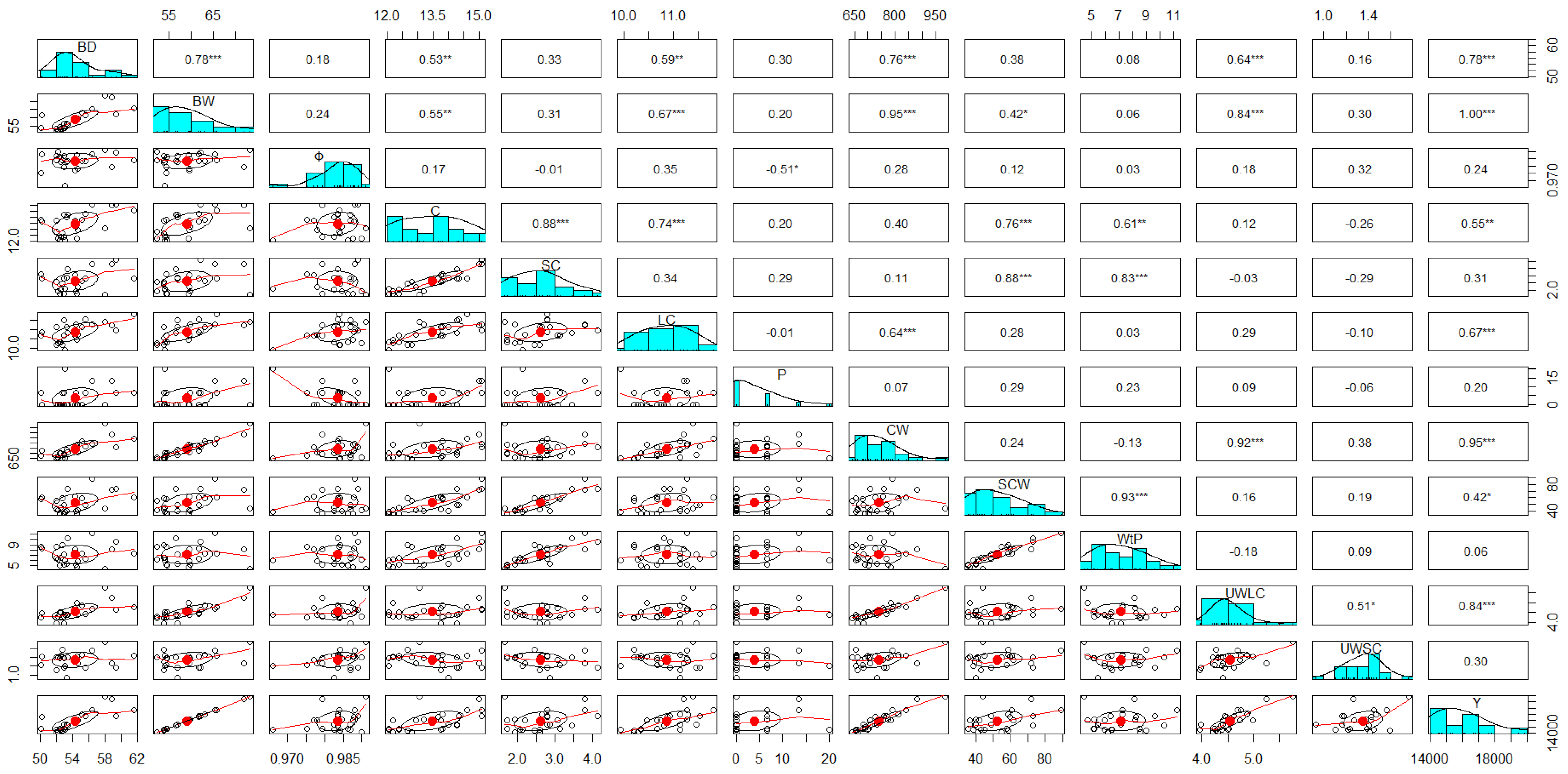
2.3. Statistical Analysis
3. Results
3.1. The First Trial: Season 2021–2022
3.2. The Second Trial: Season 2022–2023
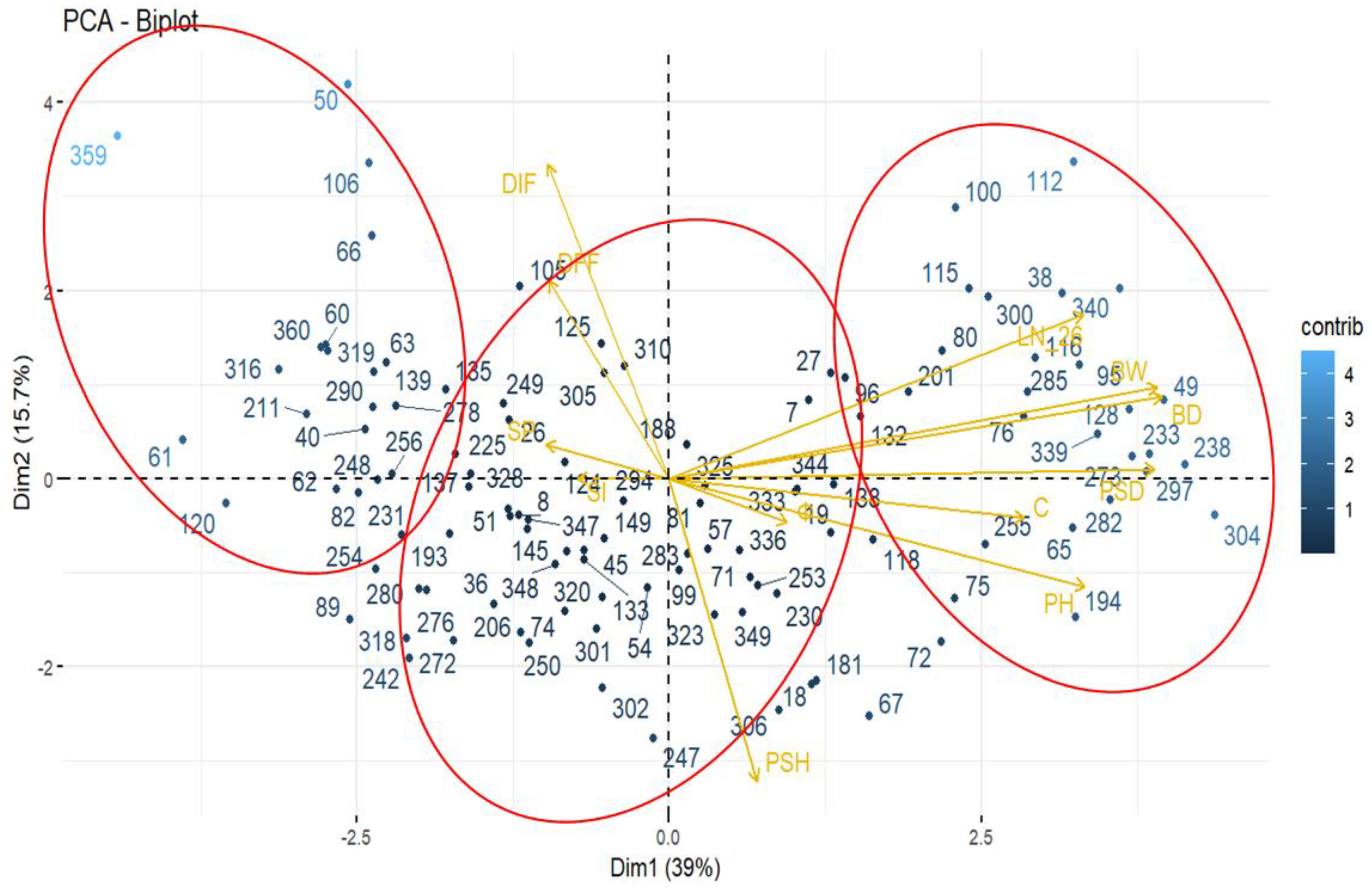
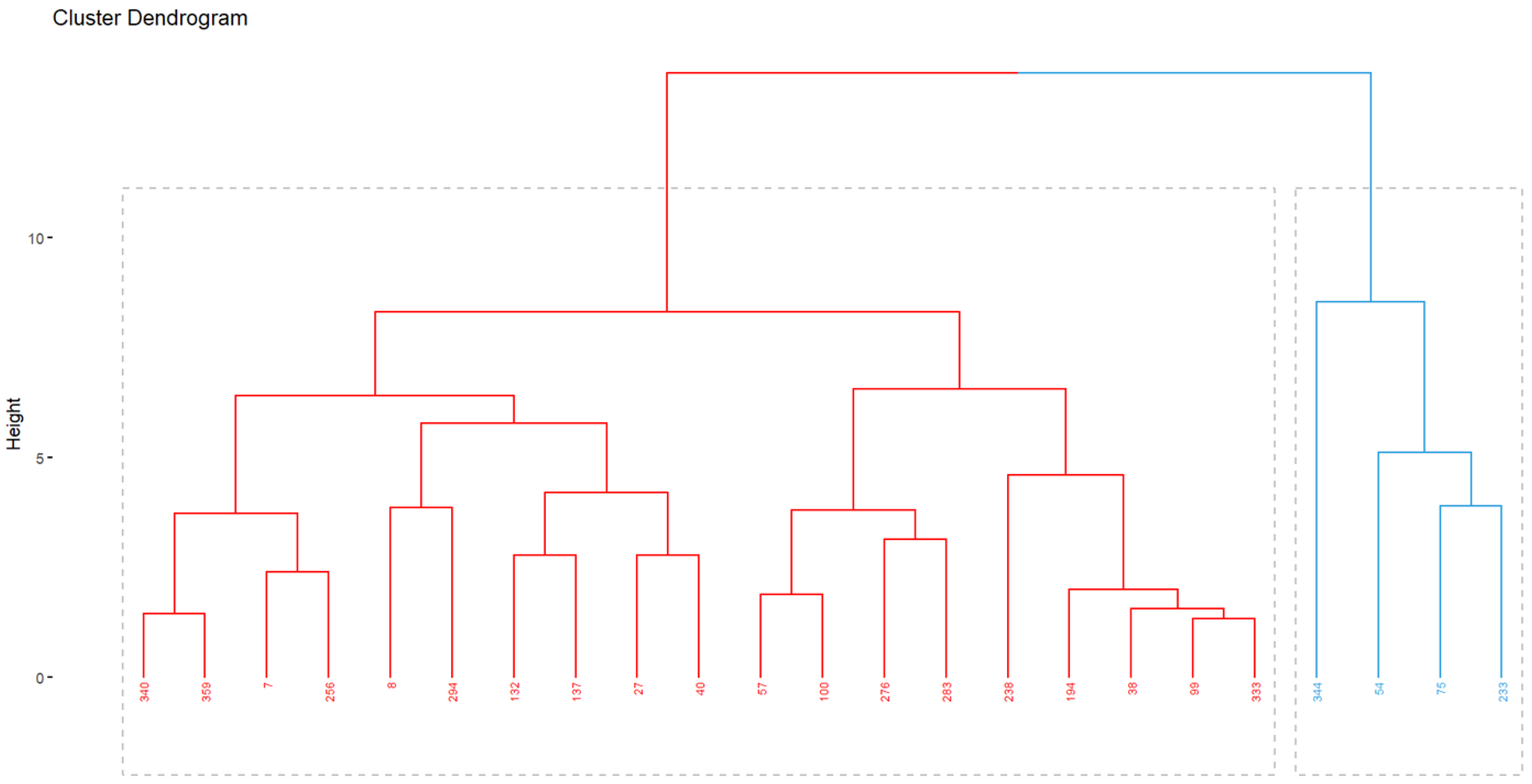
3.3. Interaction Genotype–Environment (G × E)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- APG. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Chase, M.W.; Reveal, J.L.; Fay, M.F.A. Subfamilial classification for the expanded asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Bot. J. Linn. Soc. 2009, 161, 132–136. [Google Scholar] [CrossRef]
- Haston, E.; Richardson, J.E.; Stevens, P.F.; Chase, M.W.; Harris, D.J. The Linear Angiosperm Phylogeny Group (LAPG) III: A linear sequence of the families in APG III. Bot. J. Linn. Soc. 2009, 161, 128–131. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhou, S.D.; He, X.J.; Yu, Y.; Zhang, Y.C.; Wei, X.Q. Phylogeny and biogeography of Allium (Amaryllidaceae: Allieae) based on nuclear ribosomal internal transcribed spacer and chloroplast rps16 sequences, focusing on the inclusion of species endemic to China. Ann. Bot. 2010, 106, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Giussani, L.M.; Jang, C.G.; Oh, B.U.; Cota-Sánchez, J.H. Systematics of disjunct northeastern Asian and northern north American Allium (Amaryllidaceae). Botany 2012, 90, 491–508. [Google Scholar] [CrossRef]
- Fritsch, R.M.; Friesen, N. Evolution, domestication and taxonomy. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI publishing: Wallingford, UK, 2002; pp. 5–30. [Google Scholar]
- Kamenetsky, R.; Khassanov, F.; Rabinowitch, H.D.; Auger, J.; Kik, C. Garlic biodiversity and genetic resources. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 1–5. [Google Scholar]
- Simon, P.W.; Jenderek, M.M. Flowering, seed production, and the genesis of garlic breeding. In Plant Breeding Reviews, 23; Janik, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003; pp. 211–244. [Google Scholar]
- Dhall, R.K.; Cavagnaro, P.F.; Singh, H.; Mandal, S. History, evolution and domestication of garlic: A review. Plant Syst. Evol. 2023, 309, 33. [Google Scholar] [CrossRef]
- Simon, P.W. The Origins and Distribution of Garlic: How Many Garlics Are There? Available online: https://www.ars.usda.gov/midwest-area/madison-wi/vegetable-crops-research/docs/simon-garlic-origins/ (accessed on 3 March 2020).
- Shemesh-Mayer, E.; Kamenetsky-Goldstein, R. Traditional and Novel Approaches in Garlic (Allium sativum L.) Breeding. In Advances in Plant Breeding Strategies: Vegetable Crops. Bulb, Roots and Tubers; Al-Khairy, J.M., Mohan Jain, S., Jhonson, D.V., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; Volume 8, pp. 3–49. [Google Scholar]
- Etoh, T. Fertility of the garlic clones collected in Soviet Central Asia. Hort. J. 1986, 55, 312–319. [Google Scholar]
- Lallemand, J.; Messian, C.M.; Briand, F.; Etoh, T. Delimitation of varietal groups in garlic (Allium sativum L.) by morphological, physiological and biochemical. characters. In Proceedings of the I International Symposium on Edible Alliaceae, Mendoza, Argentina, 14–18 March 1994; Burba, J.L., Galmarini, C.R., Eds.; ISHS Acta Horticulturae: Leuven, Belgium, 1997; Volume 433, pp. 123–132. [Google Scholar]
- Chovelon, V.; Souche, S.; Delecolle, B.; Etoh, T.; Messiaen, C.M.; Lot, H. Resistance to onion yellow dwarf virus and leek yellow stripe virus found in a fertile garlic clone. In Proceeding of the II International Symposium on Edible Alliaceae, Adelaide, Australia, 10–13 November 1997; Armstrong, J., Ed.; ISHS Acta Horticulturae: Leuven, Belgium, 2001; Volume 555, pp. 243–246. [Google Scholar]
- Al-Safadi, B.; Arabi, M.I.E.; Ayyoubi, Z. Differences in quantitative and qualitative characteristics of local and introduced cultivars and mutated lines of garlic. J. Veg. Crop Prod. 2003, 9, 21–31. [Google Scholar] [CrossRef]
- Burba, J.L.; Portela, J.A.; Lanzavechia, S. Argentine garlic I: A wide offer of clonal cultivars. In Proceedings of the IV International Symposium on Edible Alliaceae, Beijing, China, 21–26 April 2004; Guangshu, L., Ed.; ISHS Acta Horticulturae: Leuven, Belgium, 2005; Volume 688, pp. 291–296. [Google Scholar]
- Volk, G.M.; Stern, D. Phenotypic characteristics of ten garlic cultivars grown at different North American locations. HortScience 2009, 44, 1238–1247. [Google Scholar] [CrossRef]
- García-Lampasona, S.; Asprelli, P.; Burba, J.L. Genetic analysis of a garlic (Allium sativum L.) germplasm collection from Argentina. Sci. Hortic. 2012, 138, 183–189. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Y.; Liu, X.; Du, Z.; Qiu, Y.; Song, J.; Zhang, X.; Li, X. Resistance and clonal selection among Allium sativum L. germplasm resources to Delia antiqua M. and its correlation with allicin content. Pest. Manag. Sci. 2019, 75, 2830–2839. [Google Scholar] [CrossRef]
- Barboza, K.; Salinas, M.C.; Acuña, C.V.; Bannoud, F.; Beretta, V.; Garcia-Lampasona, S.; Burba, J.L.; Galmarini, C.R.; Cavagnaro, P.F. Assessment of genetic diversity and population structure in a garlic (Allium sativum L.) germplasm collection varying in bulb content of pyruvate, phenolics, and solids. Sci. Hortic. 2020, 261, 108900. [Google Scholar] [CrossRef]
- Ferreira de Carvalho, J.; de Jager, V.; van Gurp, T.P.; Wagemaker, N.C.A.M.; Verhoeven, K.J.F. Recent and dynamic transposable elements contribute to genomic divergence under asexuality. BMC Genom. 2016, 17, 884. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhu, S.; Li, N.; Cheng, Y.; Zhao, J.; Qiao, X.; Lu, L.; Liu, S.; Wang, Y.; Liu, C.; et al. A chromosome-level genome assembly of garlic (Allium sativum) provides insights into genome evolution and allicin biosynthesis. Mol. Plant 2020, 13, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Ajo Morado de Las Pedroñeras. Description. EC No: ES/PGI/005/0228/12.03.2002. GI 1204. The International Appellations of Origin; World Intellectual Property Organization (WIPO): Geneva, Switzerland, 2023; pp. 1–18. [Google Scholar]
- Peña-Iglesias, A. El ajo: Virosis, fisiopatías y selección clonal y sanitaria. II: Científico-experimental. Boletín Sanid. Vegetal. Plagas 1988, 14, 493–553. [Google Scholar]
- RAEA. Ensayo de variedades comerciales de ajo. Campaña 2005–2006. Instituto de Investigación de Formación Agraria y Pesquera. Consejería de Innovación, Ciencia y Empresa. Consejería de Agricultura y Pesca. Junta de Andalucía, 2006; pp. 1–92. Available online: https://www.juntadeandalucia.es/export/drupaljda/1337163241RAEA_Ajo_2006.pdf (accessed on 16 April 2024).
- Bhojwani, S.S.; Cohen, D.; Fry, P.R. Production of virus-free garlic and field performance of micropropagated plants. Sci. Hortic. 1982, 18, 39–43. [Google Scholar] [CrossRef]
- R Core Team. R. A language and environment for statistical computing. In R Foundation for Statistical Computing; R Foundation: Vienna, Austria, 2013. [Google Scholar]
- Peña, E.A.; Slate, E.H. Global validation of linear model assumptions. J. Am. Stat. Assoc. 2006, 101, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Tomczak, M.; Tomczak, E. The need to report effect size estimates revisited. An overview of some recommended measures of effect size. Trends Sport. Sci. 2014, 1, 19–25. [Google Scholar]
- Sawilowsky, S.S. New effect size rules of thumb. J. Mod. Appl. Stat. Methods 2009, 8, 26. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: A Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Gimenez, M.D.; García Lampasona, S. Before-after analysis of genetic and epigenetic markers in garlic: A 13-year experiment. Sci. Hortic. 2018, 240, 23–28. [Google Scholar] [CrossRef]
- Figliuolo, G.; Candido, V.; Logozzo, G.; Miccolis, V.; Spagnoletti Zeuli, P.L. Genetic evaluation of cultivated garlic germplasm (Allium sativum L. and A. ampeloprasum L.). Euphytica 2001, 121, 325–334. [Google Scholar] [CrossRef]
- Ragas, R.E.G.; Padron, F.K.J.R.; Ruedas, M.Y.A.D. Analysis of the morpho-anatomical traits of four major garlic (Allium sativum L.) cultivars in the Philippines. Appl. Ecol. Environ. Res. 2019, 17, 1143–1157. [Google Scholar] [CrossRef]
- Benke, A.P.; Khar, A.; Mahajan, V.; Gupta, A.; Singh, M. Study on dispersion of genetic variation among Indian garlic ecotypes using agro morphological traits. Indian. J. Gen. Plant Breed. 2020, 80, 94–102. [Google Scholar] [CrossRef]
- Khar, A.; Hirata, S.; Abdelrahman, M.; Shigyo, M.; Singh, H. Breeding and genomic approaches for climate-resilient garlic. In Genomic Designing of Climate-Smart Vegetable Crops; Chittaranjan, K., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 359–383. [Google Scholar]
- Baghalian, K.; Ziai, S.A.; Naghavi, M.R.; Badi, H.N.; Khalighi, A. Evaluation of allicin content and botanical traits in Iranian garlic (Allium sativum L.) ecotypes. Sci. Hortic. 2005, 103, 155–166. [Google Scholar] [CrossRef]
- Panthee, D.R.; Kc, R.B.; Regmi, H.N.; Subedi, P.P.; Bhattarai, S.; Dhakal, J. Diversity analysis of garlic (Allium sativum L.) germplasms available in Nepal based on morphological characters. Genet. Resour. Crop Evol. 2006, 53, 205–212. [Google Scholar] [CrossRef]
- Jabbes, N.; Arnault, I.; Auger, J.; Dridi, B.A.M.; Hannachi, C. Agro-morphological markers and organo-sulphur compounds to assess diversity in Tunisian garlic landraces. Sci. Hortic. 2012, 148, 47–54. [Google Scholar] [CrossRef]
- Tchorzewska, D.; Bocianowski, J.; Najda, A.; Dąbrowska, A.; Winiarczyk, K. Effect of environment fluctuations on biomass and allicin level in Allium sativum (cv. Harnas, Arkus) and Allium ampeloprasum var. ampeloprasum (GHG-L). J. Appl. Bot. Food Qual. 2017, 90, 106–114. [Google Scholar]
- Casals, J.; Rivera, A.; Campo, S.; Aymerich, E.; Isern, H.; Fenero, D.; Garriga, A.; Palou, A.; Monfort, A.; Howad, W.; et al. Phenotypic diversity and distinctiveness of the Belltall garlic landrace. Front. Plant Sci. 2023, 13, 1004069. [Google Scholar] [CrossRef]
- Nasir, S.; Regasa, T.; Yirgu, M. Influence of clove weight and planting depth on yield and yield components of garlic (Allium sativum L.). American-Eurasian J. Agric. Environ. Sci. 2017, 17, 315–319. [Google Scholar]
- IPGRI; ECP/GR; AVRDC. Descriptors for Allium (Allium spp.); International Plant Genetic Resources Institute: Rome, Italy; European Cooperative Programme for Crop Genetic Resources Networks (ECP/GR), Asian Vegetable Research and Development Center: Taiwan, China, 2001; pp. 1–42. [Google Scholar]
- Quintero, J.J. El Cultivo del Ajo; Hojas Divulgadoras, Ministerio de Agricultura: Pesca y Alimentación, España, 1984; Volume 1/84, pp. 1–16. Available online: https://www.mapa.gob.es/ministerio/pags/biblioteca/hojas/hd_1984_01.pdf (accessed on 16 April 2024).
- Hoopes, R.W.; Plaisted, R.L. Potato. In Principles of Cultivar Development: Crop Species; Fehr, W.R., Ed.; Macmillan Publishing Company: New York, NY, USA, 1987; Volume 2, pp. 385–436. [Google Scholar]
- Tascón Rodríguez, C.; Morales, D.A.; Ríos Mesa, D.J. Caracterización morfológica preliminar de un grupo de ajos de la Isla de Tenerife. In XXXVI Seminario de Técnicos y Especialistas en Horticultura: Ibiza, 2006; Centro de Publicaciones Agrarias: Pesqueras y Alimentarias, España, 2007; pp. 53–58. [Google Scholar]
- Chen, S.; Chen, W.; Shen, X.; Yang, Y.; Qi, F.; Liu, Y.; Meng, H. Analysis of the genetic diversity of garlic (Allium sativum L.) by simple sequence repeat and inter simple sequence repeat analysis and agro-morphological traits. Biochem. Syst. Ecol. 2014, 55, 260–267. [Google Scholar] [CrossRef]
- Papaioannou, C.; Fassou, G.; Petropoulos, S.A.; Lamari, F.N.; Bebeli, P.J.; Papasotiropoulos, V. Evaluation of the genetic diversity of Greek garlic (Allium sativum L.) accessions using DNA markers and association with phenotypic and chemical variation. Agriculture 2023, 13, 1408. [Google Scholar] [CrossRef]
- Mengistu, F.G.; Mossie, G.A.; Fita, G.T. Evaluation of garlic genotypes for yield performance and stability using GGE biplot analysis and genotype by environment interaction. Plant Genet. Resour. 2023, 21, 490–496. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dependent Variables | F Value | df | Signif. (p) | ω2 | 95% CI ω2 ** | Interpretation of Effect Size |
|---|---|---|---|---|---|---|
| PSH | 6.631 | 102 | <0.001 | 0.54 | (0.41–1.00) | Medium |
| PSD | 8.216 | 102 | <0.001 | 0.60 | (0.50–1.00) | Medium |
| LN_26 | 5.030 | 102 | <0.001 | 0.45 | (0.29–1.00) | Small |
| PH | 7.843 | 102 | <0.001 | 0.58 | (0.48–1.00) | Medium |
| BD | 12.230 | 102 | <0.001 | 0.70 | (0.63–1.00) | Medium |
| Φ * | 148.100 | 102 | <0.01 | 0.27 | (0.36–0.48) | Small |
| BW | 12.540 | 102 | <0.001 | 0.70 | (0.64–1.00) | Medium |
| C | 6.296 | 102 | <0.001 | 0.52 | (0.39–1.00) | Medium |
| Dependent Variables | F Value | df | Signif. (p) | ω2 | 95% CI ω2 * | Interpretation of Effect Size |
|---|---|---|---|---|---|---|
| BD | 8.254 | 23 | <0.001 | 0.32 | (0.22, 1.00) | Small |
| Φ | 2.326 | 23 | <0.001 | 0.08 | (0.00, 1.00) | Very Small |
| BW | 5.330 | 23 | <0.001 | 0.22 | (0.11, 1.00) | Small |
| C | 8.883 | 23 | <0.001 | 0.33 | (0.24, 1.00) | Small |
| SC | 8.018 | 23 | <0.001 | 0.31 | (0.21, 1.00) | Small |
| LC | 3.225 | 23 | <0.001 | 0.12 | (0.02, 1.00) | Small |
| Family | Yield (kg ha−1) | Grouping of Families by Yield | Bulb Size Category (%) | |||||
|---|---|---|---|---|---|---|---|---|
| F < 37 mm | E 37–45 mm | D 45–50 mm | C 50–55 mm | B 55–60 mm | A >60 mm | |||
| 38 | 14,112 | I | 0.0 | 0.0 | 20.0 | 60.0 | 20.0 | 0.0 |
| 57 | 14,490 | 0.0 | 0.0 | 20.0 | 53.3 | 26.7 | 0.0 | |
| 99 | 14,724 | 0.0 | 6.7 | 13.3 | 66.7 | 13.3 | 0.0 | |
| 100 | 14,112 | 0.0 | 13.3 | 26.7 | 26.7 | 26.7 | 6.7 | |
| 194 | 14,652 | 0.0 | 0.0 | 13.3 | 73.3 | 6.7 | 6.7 | |
| 238 | 14,562 | 0.0 | 0.0 | 26.7 | 53.3 | 13.3 | 6.7 | |
| 256 | 15,372 | 0.0 | 0.0 | 20.0 | 46.7 | 33.3 | 0.0 | |
| 276 | 15,228 | 0.0 | 0.0 | 6.7 | 60.0 | 33.3 | 0.0 | |
| 283 | 14,670 | 0.0 | 6.7 | 53.3 | 26.7 | 13.3 | 0.0 | |
| 333 | 14,544 | 0.0 | 0.0 | 6.7 | 73.3 | 20.0 | 0.0 | |
| 340 | 15,336 | 0.0 | 0.0 | 13.3 | 46.7 | 40.0 | 0.0 | |
| 359 | 14,508 | 0.0 | 6.7 | 13.3 | 66.7 | 13.3 | 0.0 | |
| 7 | 16,668 | II | 0.0 | 0.0 | 20.0 | 20.0 | 53.3 | 6.7 |
| 8 | 16,092 | 0.0 | 0.0 | 20.0 | 26.7 | 46.7 | 6.7 | |
| 27 | 16,650 | 0.0 | 13.3 | 26.7 | 53.3 | 6.7 | 0.0 | |
| 40 | 17,028 | 0.0 | 0.0 | 6.7 | 40.0 | 46.7 | 6.7 | |
| 132 | 16,056 | 0.0 | 0.0 | 13.3 | 46.7 | 26.7 | 13.3 | |
| 137 | 16,146 | 0.0 | 0.0 | 6.7 | 53.3 | 40.0 | 0.0 | |
| 233 | 16,776 | 0.0 | 0.0 | 0.0 | 20.0 | 33.3 | 46.7 | |
| 294 | 16,092 | 0.0 | 0.0 | 0.0 | 53.3 | 46.7 | 0.0 | |
| 54 | 17,640 | III | 0.0 | 0.0 | 13.3 | 20.0 | 40.0 | 26.7 |
| 75 | 19,458 | IV | 0.0 | 0.0 | 0.0 | 0.0 | 66.7 | 33.3 |
| 344 | 19,800 | 0.0 | 0.0 | 0.0 | 20.0 | 66.7 | 13.3 | |
| Dependent Variable | Variation Factor | F Value | df | Signif. (p) | ω2 | 95% CI ω2 * | Interpretation of Effect Size |
|---|---|---|---|---|---|---|---|
| Bulb Diameter | |||||||
| F | 8.750 | 22 | <0.001 | 0.27 | (0.18, 1.00) | Small | |
| L | 824.089 | 1 | <0.001 | 0.64 | (0.60, 1.00) | Medium | |
| F × L | 9.725 | 22 | <0.001 | 0.30 | (0.21, 1.00) | Small | |
| Sphericity | |||||||
| F | 1.507 | 22 | n.s. | 0.02 | (0.00, 1.00) | Very small | |
| L | 69.445 | 1 | <0.001 | 0.13 | (0.08, 1.00) | Small | |
| F × L | 2.256 | 22 | <0.01 | 0.06 | (0.00, 1.00) | Very small | |
| Bulb Weight | |||||||
| F | 6.296 | 22 | <0.001 | 0.20 | (0.12, 1.00) | Small | |
| L | 429.649 | 1 | <0.001 | 0.49 | (0.43, 1.00) | Small | |
| F × L | 8.750 | 22 | <0.001 | 0.27 | (0.18, 1.00) | Small | |
| Number of Cloves | |||||||
| F | 7.859 | 22 | <0.001 | 0.25 | (0.16, 1.00) | Small | |
| L | 256.415 | 1 | <0.001 | 0.36 | (0.30, 1.00) | Small | |
| F × L | 5.255 | 22 | <0.001 | 0.17 | (0.08, 1.00) | Small | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Licea-Moreno, R.J.; Rodríguez-Haro, Á.; Marín-Martínez, J.A. Toward the Development of Garlic Varieties: The First Attempts. Agronomy 2024, 14, 1812. https://doi.org/10.3390/agronomy14081812
Licea-Moreno RJ, Rodríguez-Haro Á, Marín-Martínez JA. Toward the Development of Garlic Varieties: The First Attempts. Agronomy. 2024; 14(8):1812. https://doi.org/10.3390/agronomy14081812
Chicago/Turabian StyleLicea-Moreno, Ricardo Julián, Ángeles Rodríguez-Haro, and Juan Antonio Marín-Martínez. 2024. "Toward the Development of Garlic Varieties: The First Attempts" Agronomy 14, no. 8: 1812. https://doi.org/10.3390/agronomy14081812