

Exogenously Applied Sodium Nitroprusside Alleviated Cadmium Toxicity in Different Aromatic Rice Cultivars by Improving Nitric Oxide Accumulation and Modulating Oxidative Metabolism

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site, Plant Husbandry, and Growth Environments
2.2. Estimating the Photosynthetic Efficacy, Pigment Contents, and Transmission Electron Microscopy
2.3. Estimation of ROS Level, Lipid Peroxidation, Electrolyte Leakage, and Proline Content
2.4. Estimating the Cd Concentrations in Roots, Leaves, and Grains
2.5. Estimation of NO Content
2.6. Estimating the Antioxidant Enzymes
2.7. Estimating the Grain Yield Attributes and Quality Traits of Fragrant Rice
2.8. Quantification of 2-Acetyl-1-Pyrroline (2AP)
2.9. Statistical Analysis
3. Results
3.1. Impacts of SNP Treatments on Cadmium Concentration, Nitric Oxide Content, and Plant Growth
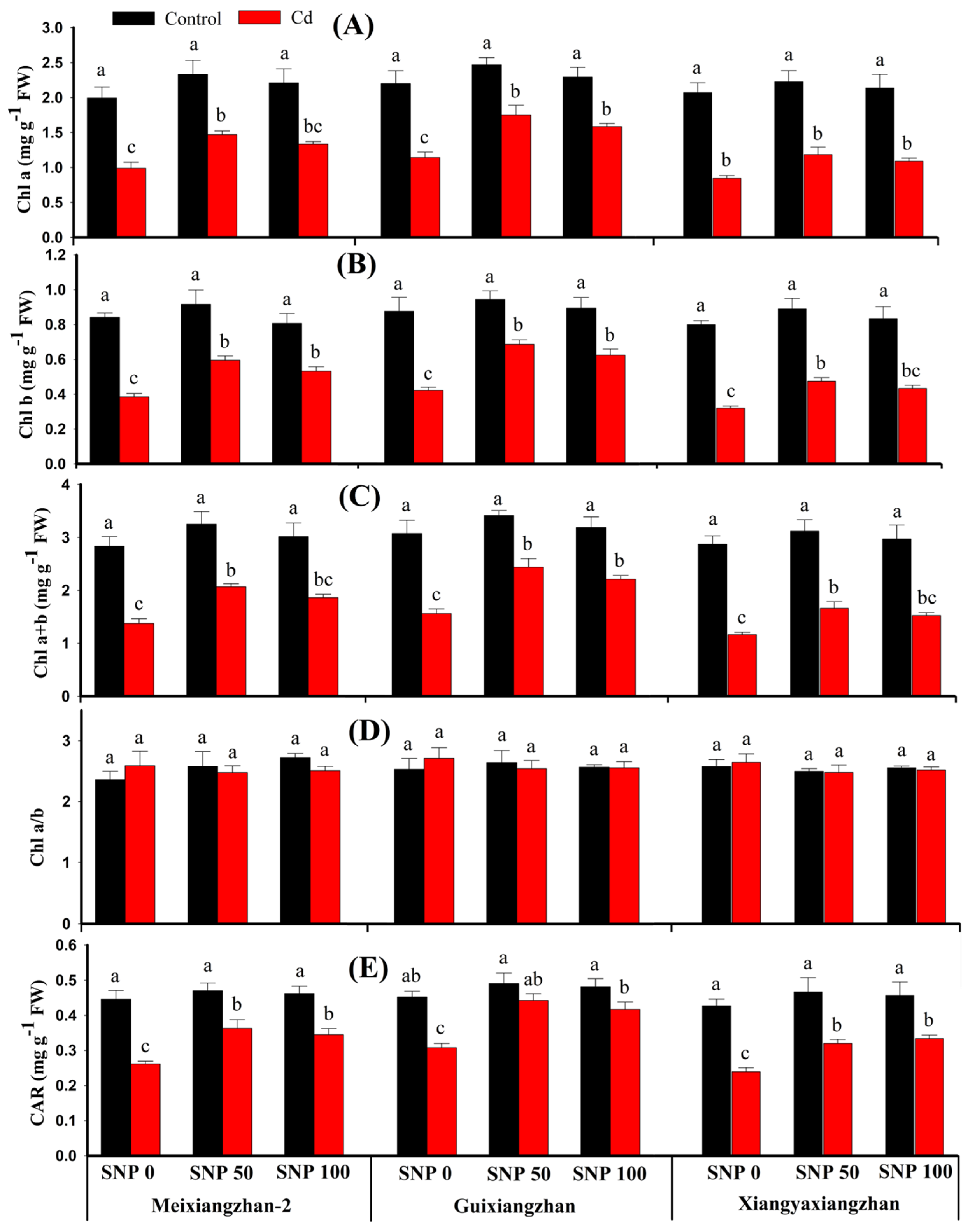
3.2. Effects of SNP Treatments on Photosynthetic Pigments
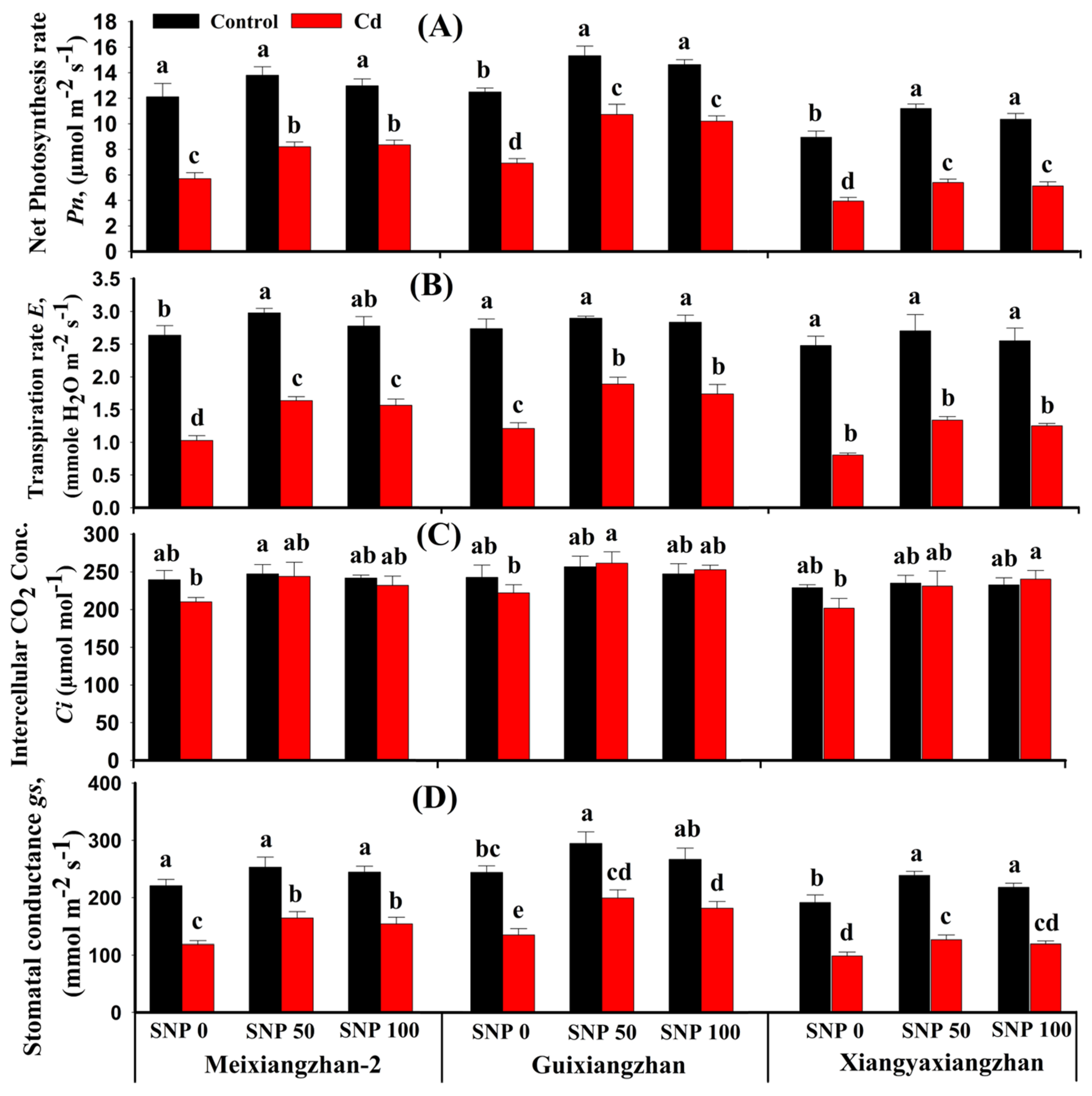
3.3. SNP Application Improved Photosynthetic Apparatus
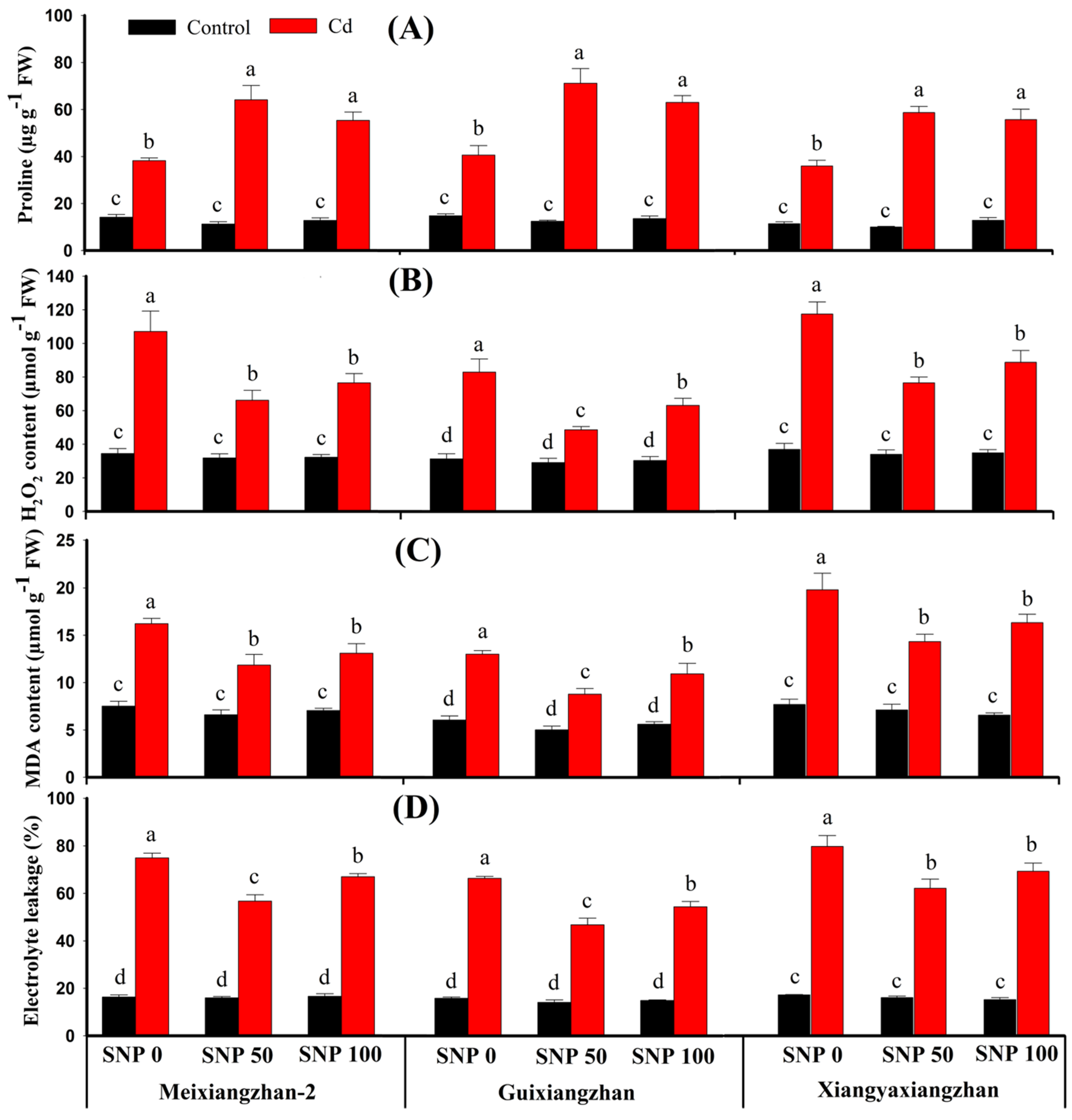
3.4. Impact of Foliar SNP Treatments on Membrane Integrity
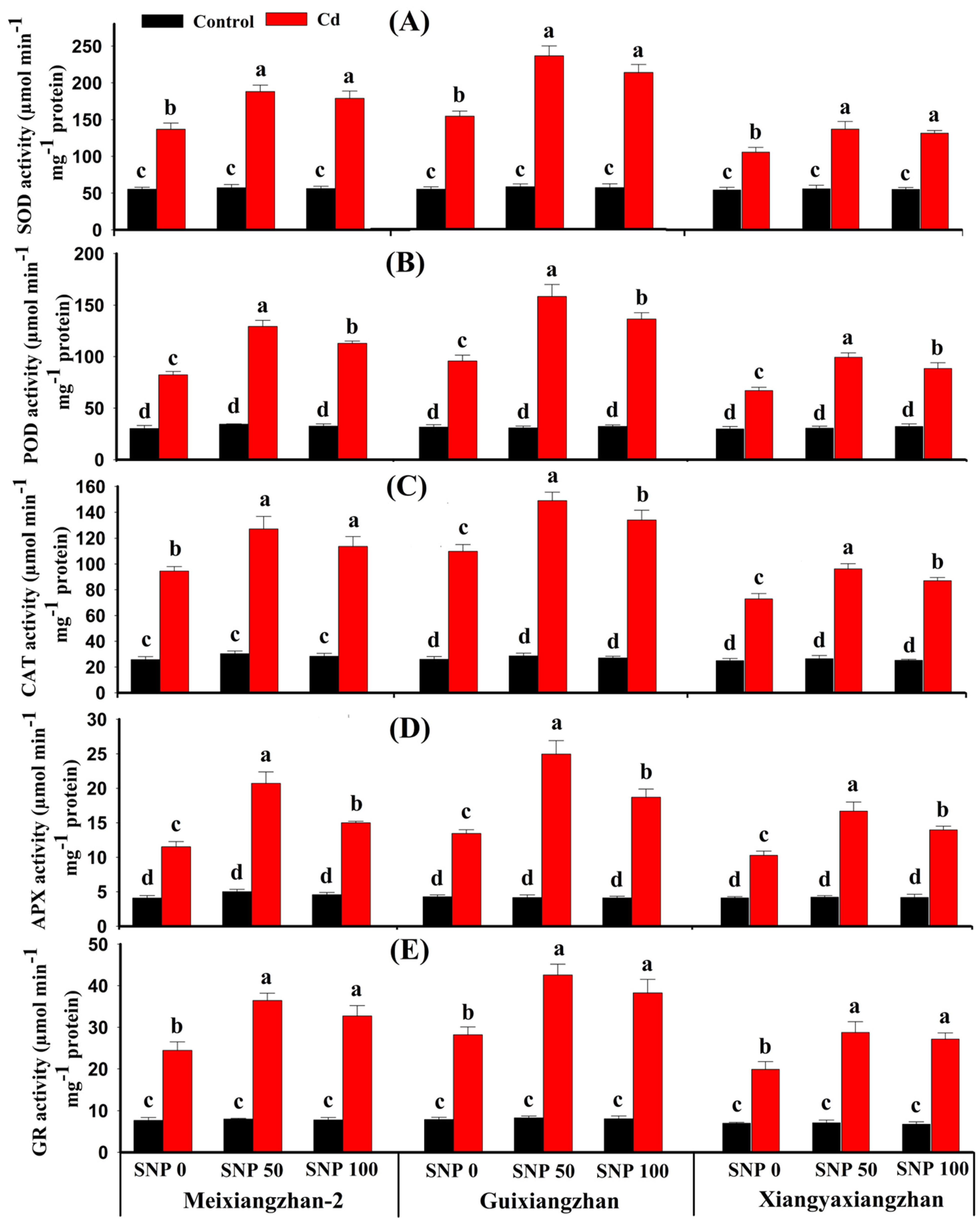
3.5. Foliar Application of SNP-Enhanced Antioxidant Enzyme Activity under Cd Toxicity
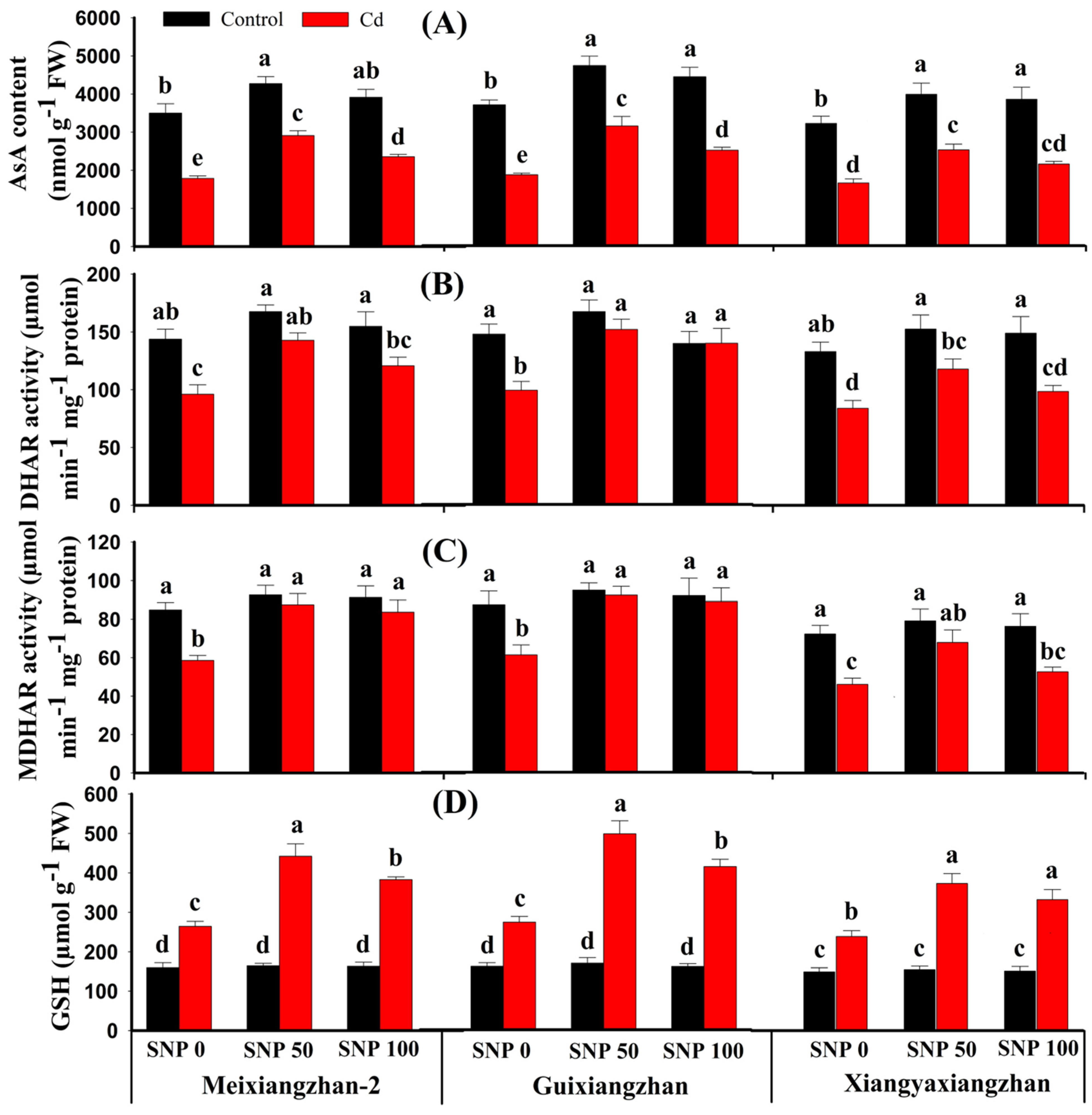
3.6. Foliar Application of SNP Regulated AsA-GSH Cycle
3.7. Influence of SNP Treatments on Aromatic Rice Yield and Grain Attributes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bano, K.; Kumar, B.; Alyemeni, M.N.; Ahmad, P. Exogenously-Sourced Salicylic Acid Imparts Resilience towards Arsenic Stress by Modulating Photosynthesis, Antioxidant Potential and Arsenic Sequestration in Brassica Napus Plants. Antioxidants 2022, 11, 2010. [Google Scholar] [CrossRef] [PubMed]
- Rather, B.A.; Masood, A.; Sehar, Z.; Majid, A.; Anjum, N.A.; Khan, N.A. Mechanisms and Role of Nitric Oxide in Phytotoxicity-Mitigation of Copper. Front. Plant Sci. 2020, 11, 675. [Google Scholar]
- Mir, I.R.; Rather, B.A.; Masood, A.; Anjum, N.A.; Khan, N.A. Nitrogen Sources Mitigate Cadmium Phytotoxicity Differentially by Modulating Cellular Buffers, N-Assimilation, Non-Protein Thiols, and Phytochelatins in Mustard (Brassica juncea L.). J. Soil Sci. Plant Nutr. 2022, 22, 3847–3867. [Google Scholar] [CrossRef]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Tack, F.M.G.; Sebastian, A.; Prasad, M.N.V.; Rinklebe, J. Cadmium Stress in Plants: A Critical Review of the Effects, Mechanisms, and Tolerance Strategies. Crit. Rev. Environ. Sci. Technol. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Parmar, P.; Kumari, N.; Sharma, V. Structural and Functional Alterations in Photosynthetic Apparatus of Plants under Cadmium Stress. Bot. Stud. 2013, 54, 45. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of Cadmium Stress on Leaf Chlorophyll Fluorescence and Photosynthesis of Elsholtzia Argyi—A Cadmium Accumulating Plant. Int. J. Phytoremediat. 2015, 17, 85–92. [Google Scholar] [CrossRef]
- Todorenko, D.; Volgusheva, A.; Timofeev, N.; Kovalenko, I.; Matorin, D.; Antal, T. Multiple in Vivo Effects of Cadmium on Photosynthetic Electron Transport in Pea Plants. Photochem. Photobiol. 2021, 97, 1516–1526. [Google Scholar] [CrossRef]
- Mir, I.R.; Rather, B.A.; Sehar, Z.; Masood, A.; Khan, N.A. Nitric Oxide in Co-Ordination with Nitrogen Reverses Cadmium-Inhibited Photosynthetic Activity by Interacting with Ethylene Synthesis, Strengthening the Antioxidant System, and Nitrogen and Sulfur Assimilation in Mustard (Brassica juncea L.). Sci Hortic 2023, 314, 111958. [Google Scholar] [CrossRef]
- Bano, K.; Kumar, B.; Alyemeni, M.N.; Ahmad, P. Protective Mechanisms of Sulfur against Arsenic Phytotoxicity in Brassica Napus by Regulating Thiol Biosynthesis, Sulfur-Assimilation, Photosynthesis, and Antioxidant Response. Plant Physiol. Biochem. 2022, 188, 1–11. [Google Scholar] [CrossRef]
- Yang, Y.; Xiong, J.; Tao, L.; Cao, Z.; Tang, W.; Zhang, J.; Yu, X.; Fu, G.; Zhang, X.; Lu, Y. Regulatory Mechanisms of Nitrogen (N) on Cadmium (Cd) Uptake and Accumulation in Plants: A Review. Sci. Total Environ. 2020, 708, 135186. [Google Scholar] [CrossRef]
- Imran, M.; Shafiq, S.; Ashraf, U.; Qi, J.; Mo, Z.; Tang, X. Biosynthesis of 2-Acetyl-1-Pyrroline in Fragrant Rice: Recent Insights into Agro-Management, Environmental Factors, and Functional Genomics. J. Agric. Food Chem. 2023, 71, 4201–4215. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd_Allah, E.F.; Gucel, S.; Tran, L.-S.P. Nitric Oxide Mitigates Salt Stress by Regulating Levels of Osmolytes and Antioxidant Enzymes in Chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Combined Kinetin and Spermidine Treatments Ameliorate Growth and Photosynthetic Inhibition in Vigna Angularis by Up-Regulating Antioxidant and Nitrogen Metabolism under Cadmium Stress. Biomolecules 2020, 10, 147. [Google Scholar] [CrossRef]
- Beckman, J.S.; Beckman, T.W.; Chen, J.; Marshall, P.A.; Freeman, B.A. Apparent Hydroxyl Radical Production by Peroxynitrite: Implications for Endothelial Injury from Nitric Oxide and Superoxide. Proc. Natl. Acad. Sci. USA 1990, 87, 1620–1624. [Google Scholar] [CrossRef] [PubMed]
- Padmaja, S.; Huie, R.E. The Reaction of Nitric Oxide with Organic Peroxyl Radicals. Biochem. Biophys. Res. Commun. 1993, 195, 539–544. [Google Scholar] [CrossRef]
- Rubbo, H.; Radi, R.; Trujillo, M.; Telleri, R.; Kalyanaraman, B.; Barnes, S.; Kirk, M.; Freeman, B.A. Nitric Oxide Regulation of Superoxide and Peroxynitrite-Dependent Lipid Peroxidation. Formation of Novel Nitrogen-Containing Oxidized Lipid Derivatives. J. Biol. Chem. 1994, 269, 26066–26075. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Leterrier, M.; Valderrama, R.; Airaki, M.; Chaki, M.; Palma, J.M.; Barroso, J.B. Nitric Oxide Imbalance Provokes a Nitrosative Response in Plants under Abiotic Stress. Plant Sci. 2011, 181, 604–611. [Google Scholar]
- Singh, S.; Husain, T.; Kushwaha, B.K.; Suhel, M.; Fatima, A.; Mishra, V.; Singh, S.K.; Bhatt, J.A.; Rai, M.; Prasad, S.M. Regulation of Ascorbate-Glutathione Cycle by Exogenous Nitric Oxide and Hydrogen Peroxide in Soybean Roots under Arsenate Stress. J. Hazard. Mater. 2021, 409, 123686. [Google Scholar] [PubMed]
- Wu, S.; Hu, C.; Tan, Q.; Xu, S.; Sun, X. Nitric Oxide Mediates Molybdenum-Induced Antioxidant Defense in Wheat under Drought Stress. Front. Plant Sci. 2017, 8, 1085. [Google Scholar]
- Sun, C.; Liu, L.; Yu, Y.; Liu, W.; Lu, L.; Jin, C.; Lin, X. Nitric Oxide Alleviates Aluminum-induced Oxidative Damage through Regulating the Ascorbate-glutathione Cycle in Roots of Wheat. J. Integr. Plant Biol. 2015, 57, 550–561. [Google Scholar] [PubMed]
- Rizwan, M.; Mostofa, M.G.; Ahmad, M.Z.; Imtiaz, M.; Mehmood, S.; Adeel, M.; Dai, Z.; Li, Z.; Aziz, O.; Zhang, Y. Nitric Oxide Induces Rice Tolerance to Excessive Nickel by Regulating Nickel Uptake, Reactive Oxygen Species Detoxification and Defense-Related Gene Expression. Chemosphere 2018, 191, 23–35. [Google Scholar] [PubMed]
- Fatma, M.; Masood, A.; Per, T.S.; Khan, N.A. Nitric Oxide Alleviates Salt Stress Inhibited Photosynthetic Performance by Interacting with Sulfur Assimilation in Mustard. Front. Plant Sci. 2016, 7, 521. [Google Scholar]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Elbasan, F.; Kucukoduk, M.; Turkan, I. Hydrogen Sulfide (H2S) and Nitric Oxide (NO) Alleviate Cobalt Toxicity in Wheat (Triticum aestivum L.) by Modulating Photosynthesis, Chloroplastic Redox and Antioxidant Capacity. J. Hazard. Mater. 2020, 388, 122061. [Google Scholar]
- Mukherjee, S. Recent Advancements in the Mechanism of Nitric Oxide Signaling Associated with Hydrogen Sulfide and Melatonin Crosstalk during Ethylene-Induced Fruit Ripening in Plants. Nitric Oxide 2019, 82, 25–34. [Google Scholar]
- Wakte, K.; Zanan, R.; Hinge, V.; Khandagale, K.; Nadaf, A.; Henry, R. Thirty-three Years of 2-acetyl-1-pyrroline, a Principal Basmati Aroma Compound in Scented Rice (Oryza sativa L.): A Status Review. J. Sci. Food Agric. 2017, 97, 384–395. [Google Scholar] [PubMed]
- Mo, Z.; Li, Y.; Nie, J.; He, L.; Pan, S.; Duan, M.; Tian, H.; Xiao, L.; Zhong, K.; Tang, X. Nitrogen Application and Different Water Regimes at Booting Stage Improved Yield and 2-Acetyl-1-Pyrroline (2AP) Formation in Fragrant Rice. Rice 2019, 12, 74. [Google Scholar]
- Kanu, A.S.; Ashraf, U.; Mo, Z.; Fuseini, I.; Mansaray, L.R.; Duan, M.; Pan, S.; Tang, X. Cadmium Uptake and Distribution in Fragrant Rice Genotypes and Related Consequences on Yield and Grain Quality Traits. J. Chem. 2017, 2017, 1405878. [Google Scholar]
- Imran, M.; Junaid, M.; Shafiq, S.; Liu, S.; Chen, X.; Wang, J.; Tang, X. Multiomics Analysis Reveals a Substantial Decrease in Nanoplastics Uptake and Associated Impacts by Nano Zinc Oxide in Fragrant Rice (Oryza sativa L.). J. Hazard. Mater. 2024, 474, 134640. [Google Scholar] [CrossRef]
- Imran, M.; Hu, C.; Hussain, S.; Rana, M.S.; Riaz, M.; Afzal, J.; Aziz, O.; Elyamine, A.M.; Ismael, M.A.F.; Sun, X. Molybdenum-Induced Effects on Photosynthetic Efficacy of Winter Wheat (Triticum aestivum L.) under Different Nitrogen Sources Are Associated with Nitrogen Assimilation. Plant Physiol. Biochem. 2019, 141, 154–163. [Google Scholar]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of Osmotic Stress on Compatible Solutes Content, Membrane Stability and Water Relations in Two Maize Cultivars. Plant Soil Environ. 2006, 52, 184. [Google Scholar]
- Imran, M.; Sun, X.; Hussain, S.; Ali, U.; Rana, M.S.; Rasul, F.; Shaukat, S.; Hu, C. Molybdenum Application Regulates Oxidative Stress Tolerance in Winter Wheat under Different Nitrogen Sources. J. Soil Sci. Plant Nutr. 2020, 20, 1827–1837. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar]
- Filipiak-Szok, A.; Kurzawa, M.; Szłyk, E. Determination of Toxic Metals by ICP-MS in Asiatic and European Medicinal Plants and Dietary Supplements. J. Trace Elem. Med. Biol. 2015, 30, 54–58. [Google Scholar] [PubMed]
- Cao, F.; Cai, Y.; Liu, L.; Zhang, M.; He, X.; Zhang, G.; Wu, F. Differences in Photosynthesis, Yield and Grain Cadmium Accumulation as Affected by Exogenous Cadmium and Glutathione in the Two Rice Genotypes. Plant Growth Regul. 2015, 75, 715–723. [Google Scholar]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in Plants: Uptake, Toxicity, and Its Interactions with Selenium Fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of Photosynthesis to Different Concentrations of Heavy Metals in Davidia Involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar]
- Liu, Z.; Gao, J.; Gao, F.; Liu, P.; Zhao, B.; Zhang, J. Photosynthetic Characteristics and Chloroplast Ultrastructure of Summer Maize Response to Different Nitrogen Supplies. Front. Plant Sci. 2018, 9, 576. [Google Scholar]
- Khan, M.N.; Siddiqui, M.H.; AlSolami, M.A.; Alamri, S.; Hu, Y.; Ali, H.M.; Al-Amri, A.A.; Alsubaie, Q.D.; Al-Munqedhi, B.M.A.; Al-Ghamdi, A. Crosstalk of Hydrogen Sulfide and Nitric Oxide Requires Calcium to Mitigate Impaired Photosynthesis under Cadmium Stress by Activating Defense Mechanisms in Vigna Radiata. Plant Physiol. Biochem. 2020, 156, 278–290. [Google Scholar]
- Misra, A.N.; Vladkova, R.; Singh, R.; Misra, M.; Dobrikova, A.G.; Apostolova, E.L. Action and Target Sites of Nitric Oxide in Chloroplasts. Nitric Oxide 2014, 39, 35–45. [Google Scholar]
- Hasanuzzaman, M.; Inafuku, M.; Nahar, K.; Fujita, M.; Oku, H. Nitric Oxide Regulates Plant Growth, Physiology, Antioxidant Defense, and Ion Homeostasis to Confer Salt Tolerance in the Mangrove Species, Kandelia Obovata. Antioxidants 2021, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Kohli, S.K.; Thukral, A.K.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Protective Role of Selenium against Chromium Stress Involving Metabolites and Essential Elements in Brassica juncea L. Seedlings. 3 Biotech 2018, 8, 66. [Google Scholar]
- Sirhindi, G.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic Acid Modulates the Physio-Biochemical Attributes, Antioxidant Enzyme Activity, and Gene Expression in Glycine Max under Nickel Toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar]
- Bali, S.; Kaur, P.; Kohli, S.K.; Ohri, P.; Thukral, A.K.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Jasmonic Acid Induced Changes in Physio-Biochemical Attributes and Ascorbate-Glutathione Pathway in Lycopersicon Esculentum under Lead Stress at Different Growth Stages. Sci. Total Environ. 2018, 645, 1344–1360. [Google Scholar] [PubMed]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Leterrier, M.; Airaki, M.; Palma, J.M.; Chaki, M.; Barroso, J.B.; Corpas, F.J. Arsenic Triggers the Nitric Oxide (NO) and S-Nitrosoglutathione (GSNO) Metabolism in Arabidopsis. Environ. Pollut. 2012, 166, 136–143. [Google Scholar] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The Relationship between Metal Toxicity and Cellular Redox Imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar]
- Chang, L.; Sun, H.; Yang, H.; Wang, X.; Su, Z.; Chen, F.; Wei, W. Over-Expression of Dehydroascorbate Reductase Enhances Oxidative Stress Tolerance in Tobacco. Electron. J. Biotechnol. 2017, 25, 1–8. [Google Scholar]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric Oxide: A Multitasked Signaling Gas in Plants. Mol. Plant 2015, 8, 506–520. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Cultivars | Treatments | Total Plant DW (g) | NO Content in Roots (nmol g−1 FW) | NO Content in Leaves (nmol g−1 FW) | Cd Conc. in Root (mg kg−1 DW) | Cd Conc. in Leaves (mg kg−1 DW) | Cd Conc. in Grains (mg kg−1 DW) | |
|---|---|---|---|---|---|---|---|---|
| MXZ | SNP0 | Cd0 | 3.21 ± 0.18 bc | 8.34 ± 0.56 b | 15.69 ± 1.03 b | – | – | – |
| Cd50 | 2.17 ± 0.10 d | 5.16 ± 0.23 c | 9.65 ± 0.55 c | 27.06 ± 1.16 a | 19.97 ± 1.27 a | 1.21 ± 0.10 a | ||
| SNP50 | Cd0 | 4.69 ± 0.35 a | 15.42 ± 1.26 a | 28.84 ± 1.52 a | – | – | – | |
| Cd50 | 2.98 ± 0.18 c | 8.14 ± 0.21 b | 15.53 ± 1.03 b | 26.87 ± 0.69 a | 14.86 ± 0.41 b | 0.59 ± 0.05 b | ||
| SNP100 | Cd0 | 3.79 ± 0.22 b | 13.49 ± 0.86 a | 26.24 ± 1.45 a | – | – | – | |
| Cd50 | 2.65 ± 0.17 cd | 6.83 ± 0.15 bc | 13.53 ± 0.66 b | 25.62 ± 1.22 a | 15.21 ± 0.49 b | 0.68 ± 0.04 b | ||
| GXZ | SNP0 | Cd0 | 3.70 ± 0.42 c | 8.78 ± 0.51 c | 16.99 ± 0.94 c | – | – | – |
| Cd50 | 2.64 ± 0.10 d | 5.70 ± 0.47 d | 11.26 ± 0.80 d | 27.94 ± 1.31 a | 18.42 ± 0.92 a | 0.98 ± 0.07 a | ||
| SNP50 | Cd0 | 5.66 ± 0.38 a | 17.25 ± 1.24 a | 31.92 ± 1.60 a | – | – | – | |
| Cd50 | 3.71 ± 0.19 c | 9.64 ± 0.45 c | 19.62 ± 0.93 c | 25.08 ± 0.91 a | 12.93 ± 0.95 b | 0.35 ± 0.02 c | ||
| SNP100 | Cd0 | 4.86 ± 0.19 b | 12.44 ± 0.90 b | 24.56 ± 1.80 b | – | – | – | |
| Cd50 | 3.51 ± 0.10 c | 8.67 ± 0.64 c | 17.04 ± 1.31 c | 26.91 ± 1.12 a | 14.40 ± 0.46 b | 0.48 ± 0.04 b | ||
| XYXZ | SNP0 | Cd0 | 2.81 ± 0.13 b | 8.28 ± 0.66 bc | 14.19 ± 0.80 b | – | – | – |
| Cd50 | 1.88 ± 0.07 c | 4.84 ± 0.32 e | 8.08 ± 0.42 d | 28.86 ± 1.27 a | 20.55 ± 0.82 a | 1.47 ± 0.11 a | ||
| SNP50 | Cd0 | 4.36 ± 0.22 a | 13.67 ± 0.78 a | 24.44 ± 1.64 a | – | – | – | |
| Cd50 | 2.53 ± 0.17 b | 7.20 ± 0.57 cd | 12.47 ± 0.55 bc | 25.20 ± 1.05 a | 16.82 ± 1.17 b | 0.94 ± 0.07 b | ||
| SNP100 | Cd0 | 3.77 ± 0.30 a | 9.43 ± 0.70 b | 21.41 ± 1.74 a | – | – | – | |
| Cd50 | 2.36 ± 0.21 bc | 6.51 ± 0.10 de | 10.61 ± 1.12 cd | 26.93 ± 1.10 a | 17.54 ± 0.66 b | 0.84 ± 0.04 b | ||
| Yield Traits | Grain Quality Attributes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rice Cultivars | Treatment | Productive Tiller per Pot | Grains per Panicle | 1000-Grains Weight | Grain Yield per Pot | Browning Rate (%) | Milling Rate (%) | Chalkiness Degree (%) | 2AP Content (ng g−1 FW) | ||
| MXZ | SNP0 | Cd0 | 25.10±1.27 b | 137.66 ± 2.10 bc | 18.49 ± 0.47 ab | 53.35 ± 1.51 b | 70.08 ± 3.58 a | 56.52 ± 1.74 bc | 13.96 ± 0.30 bc | 136.70 ± 6.17 bc | |
| Cd50 | 19.93 ± 0.61 c | 119.76 ± 5.62 c | 17.40 ± 0.99 b | 40.42 ± 2.86 c | 67.53 ± 1.69 a | 51.32 ± 2.35 c | 18.91 ± 1.35 a | 98.18 ± 7.11 c | |||
| SNP50 | Cd0 | 36.63 ± 2.47 a | 175.04 ± 11.37 a | 20.95 ± 0.92 a | 80.79 ± 5.53 a | 76.43 ± 1.84 a | 65.77 ± 2.56 a | 10.92 ± 0.86 d | 276.00 ± 25.26 a | ||
| Cd50 | 28.44 ± 1.41 b | 159.34 ± 8.06 ab | 18.88 ± 0.89 ab | 57.42 ± 2.45 b | 71.35 ± 2.19 a | 54.91 ± 1.82 bc | 14.87 ± 0.64 b | 140.53 ± 8.82 b | |||
| SNP100 | Cd0 | 34.85 ± 1.29 a | 159.68 ± 9.11 ab | 19.04 ± 0.67 ab | 71.85 ± 5.54 a | 73.35 ± 3.78 a | 62.96 ± 5.73 ab | 12.10 ± 0.19 cd | 259.26 ± 10.96 a | ||
| Cd50 | 25.85 ± 1.80 b | 149.94 ± 5.04 b | 18.57 ± 1.61 ab | 52.87 ± 2.14 b | 71.29 ± 4.30 a | 53.33 ± 1.53 c | 16.26 ± 0.86 b | 148.27 ± 9.17 b | |||
| GXZ | SNP0 | Cd0 | 30.75 ± 2.00 b | 140.33 ± 8.85 c | 19.15 ± 0.65 b | 64.36 ± 4.16 b | 81.52 ± 1.61 ab | 66.39 ± 2.05 a | 11.55 ± 0.20 bc | 158.47 ± 11.30 bc | |
| Cd50 | 21.24 ± 0.76 c | 120.99 ± 4.16 c | 17.98 ± 0.89 b | 46.08 ± 2.76 c | 77.28 ± 2.90 b | 57.28 ± 2.17 b | 15.31 ± 0.44 a | 115.32 ± 4.30 c | |||
| SNP50 | Cd0 | 43.80 ± 3.12 a | 195.44 ± 10.82 a | 21.94 ± 1.28 a | 99.23 ± 6.75 a | 90.06 ± 1.68 a | 69.14 ± 1.47 a | 9.58 ± 0.53 d | 302.23 ± 29.26 a | ||
| Cd50 | 31.57 ± 0.98 b | 166.81 ± 7.64 b | 19.90 ± 0.14 ab | 67.30 ± 1.17 b | 83.35 ± 1.93 ab | 64.79 ± 2.73 ab | 12.64 ± 0.41 b | 179.72 ± 14.47 b | |||
| SNP100 | Cd0 | 38.09 ± 2.11 a | 182.69 ± 11.96 ab | 20.76 ± 0.98 ab | 90.72 ± 6.22 a | 86.65 ± 3.29 ab | 68.82 ± 4.17 a | 10.58 ± 0.58 cd | 283.60 ± 21.02 a | ||
| Cd50 | 30.04 ± 1.92 b | 174.54 ± 4.64 ab | 18.28 ± 1.18 b | 63.51 ± 4.68 b | 82.95 ± 6.90 ab | 66.09 ± 1.82 a | 14.42 ± 0.35 a | 158.10 ± 8.91 bc | |||
| XYXZ | SNP0 | Cd0 | 22.68 ± 1.18 bc | 128.20 ± 2.49 bc | 18.19 ± 0.85 bc | 47.61 ± 3.76 b | 64.10 ± 3.28 abc | 52.05 ± 1.34 c | 19.75 ± 0.64 c | 117.31 ± 8.59 b | |
| Cd50 | 17.63 ± 1.29 c | 111.54 ± 5.23 c | 15.99 ± 0.70 c | 35.51 ± 2.68 c | 59.23 ± 1.48 c | 49.11 ± 1.99 c | 23.90 ± 0.88 a | 80.10 ± 6.38 c | |||
| SNP50 | Cd0 | 35.08 ± 2.67 a | 162.53 ± 9.84 a | 20.63 ± 1.07 a | 77.42 ± 5.76 a | 69.81 ± 2.22 ab | 61.34 ± 2.32 ab | 12.33 ± 0.55 d | 245.17 ± 23.62 a | ||
| Cd50 | 23.88 ± 1.67 b | 145.68 ± 7.37 ab | 16.90 ± 0.43 c | 46.59 ± 3.86 bc | 64.12 ± 3.54 abc | 55.52 ± 1.24 bc | 22.42 ± 0.74 ab | 108.72 ± 8.72 bc | |||
| SNP100 | Cd0 | 31.74 ± 2.71 a | 144.26 ± 6.96 ab | 20.34 ± 0.77 ab | 78.17 ± 4.04 a | 71.96 ± 5.48 a | 64.04 ± 5.01 a | 11.64 ± 0.47 d | 212.55 ± 6.99 a | ||
| Cd50 | 22.15 ± 1.19 bc | 136.41 ± 2.55 b | 16.40 ± 0.89 c | 44.68 ± 0.88 bc | 61.32 ± 1.80 bc | 56.98 ± 2.15 abc | 21.64 ± 1.03 bc | 101.36 ± 3.70 bc | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imran, M.; Hussain, S.; Rana, M.S.; Iqbal, A.; Rehman, N.U.; Chen, X.; Tang, X. Exogenously Applied Sodium Nitroprusside Alleviated Cadmium Toxicity in Different Aromatic Rice Cultivars by Improving Nitric Oxide Accumulation and Modulating Oxidative Metabolism. Agronomy 2024, 14, 1857. https://doi.org/10.3390/agronomy14081857
Imran M, Hussain S, Rana MS, Iqbal A, Rehman NU, Chen X, Tang X. Exogenously Applied Sodium Nitroprusside Alleviated Cadmium Toxicity in Different Aromatic Rice Cultivars by Improving Nitric Oxide Accumulation and Modulating Oxidative Metabolism. Agronomy. 2024; 14(8):1857. https://doi.org/10.3390/agronomy14081857
Chicago/Turabian StyleImran, Muhammad, Saddam Hussain, Muhammad Shoaib Rana, Anas Iqbal, Naveed Ur Rehman, Xiaoyuan Chen, and Xiangru Tang. 2024. "Exogenously Applied Sodium Nitroprusside Alleviated Cadmium Toxicity in Different Aromatic Rice Cultivars by Improving Nitric Oxide Accumulation and Modulating Oxidative Metabolism" Agronomy 14, no. 8: 1857. https://doi.org/10.3390/agronomy14081857
APA StyleImran, M., Hussain, S., Rana, M. S., Iqbal, A., Rehman, N. U., Chen, X., & Tang, X. (2024). Exogenously Applied Sodium Nitroprusside Alleviated Cadmium Toxicity in Different Aromatic Rice Cultivars by Improving Nitric Oxide Accumulation and Modulating Oxidative Metabolism. Agronomy, 14(8), 1857. https://doi.org/10.3390/agronomy14081857