Abstract
Lignin, an organic compound with a complex structure, is formed through the polymerization of structural units linked by carbon–carbon bonds and ether bonds. The question of whether lignin is labile or resistant to biological and chemical degradation in soil, particularly in alpine ecosystems, remains unresolved. To address this knowledge gap, we analyzed the relationship between phospholipid fatty acid biomarkers and the abundance of lignin components in grassland soils from North Tibet, China. Soil samples were collected from alpine grasslands, including alpine meadows and alpine steppes. The relative abundance of lignin in these alpine grassland soils before and after a 210-day incubation period was measured. Our results indicate that the relative abundance of lignin in the alpine grassland soils decreased during the incubation period. Significant relationships were found between the phospholipid fatty acid biomarkers of bacteria, fungi, Gram-positive bacteria, and Gram-negative bacteria and the relative abundance of lignin components. This research was conducted under laboratory conditions that are optimal for the development of microorganisms but significantly different from the conditions in Tibet. Furthermore, this study contributes to the understanding of soil organic matter degradation and the dynamics of microbial communities in alpine grassland soils in the context of future global warming.
1. Introduction
Lignin is the second most abundant polymer compound in nature, and it is considered the most important aromatic resource in biomass energy [1,2]. It is a type of renewable natural aromatic macromolecule polymer, containing guaiacyl, syringyl, and p-hydroxyphenyl propane monomers [3]. These monomers represent different lignin types, vegetation, and environments. Studies have shown that the syringyl propane monomer is the dominant component of lignin in forest soil and that the guaiacyl propane monomer is the main component of lignin in grassland soil [4,5]. The composition of lignin in soil reflects the degradation of plant debris, and studying the lignin in soils can help us understand the soil organic matter composition in different environments.
Lignin is considered to be resistant to biological and chemical degradation due to its complex structure [6,7]. Fungi, including brown-, white-, and soft-rot fungi, dominate the depolymerization process of lignin macromolecules in soil [8]. These fungi have unique physiological and biochemical mechanisms and strong metabolic abilities, and they mineralize lignin into carbon dioxide and water [9]. However, bacteria can also degrade lignin in the soil through the action of enzymes, such as laccase and peroxidase [10]. Therefore, lignin can be degraded by microbes in some environments, and lignin degradation and the related microbial mechanisms need to be studied in order to obtain a better understanding of the degradation of soil organic matter.
Some studies have reported microbial mechanisms in lignin degradation in soils. These studies have indicated that more than 90% of the known wood-rotting species are white-rot fungi [11]. In the process of lignin degradation, fungi first become adsorbed on the end of lignin and then extend into the interior, where lignin-degrading enzymes are secreted, thus degrading lignin from the inside to the outside [12]. However, white-rot fungi cannot mediate lignin decomposition and transformation alone. Lignin degradation results from the combined action of various bacteria and fungi in the soil [13,14,15]. Lignin-degrading bacteria can be divided into three categories: actinibacteria, α-proteobacteria, and γ-proteobacteria. Among prokaryotes, one of the most commonly identified and studied lignolytic enzymes is laccase [16,17]. Phylogeny has shown that many strains with laccase genes occur in proteobacteria, actinobacteria, and firmicutes [18]. The apical and branching growth of these bacteria and the formation and development of airborne or stromal hyphae lead them to resemble fungi. Recent studies by Rashid et al. [19] showed that superoxide manganese dismutase produced by Sphingobacterium (bacteroides) could oxidize lignin through a hydroxyl radical mechanism. In the composting process, actinibacteria can decompose organic matter in the initial stages and decolorize refractory organic matter such as lignin in the later stages. It appears that actinibacteria can enter the interior of refractory lignin and increase its water solubility.
The Tibetan Plateau is known as the third “pole” of the world. Due to the high altitude, it is colder than anywhere else outside the polar regions. In order to adapt to such an extreme environment, the physiological structure and activity mode and microorganisms in soil may be quite unique [20,21,22]. The composition and transformation of soil organic matter in the alpine grassland in this area may be affected by the extreme climate [23]. However, the relationship between the soil microbial community and lignin, which is part of the chemical composition of soil organic matter, has not been reported in Northern Tibet. The lignin components in soil from alpine meadow and alpine steppe were analyzed using pyrolytic gas chromatography–mass spectrometry (Py-GC/MS). The purpose of this study was to compare the lignin content in soil organic matter in different alpine grasslands under the extreme climatic conditions in the Tibetan Plateau and to explore the relationship between lignin content and soil microorganisms.
2. Materials and Methods
2.1. Study Area
Northern Tibet is located between the Kailas Range and Nyanqentanglala in the Tibet Autonomous Region, China [24]. It covers about 446,000 km2 and approximately one-third of the total area of the Tibet Autonomous Region, with an average elevation of more than 4500 m [25,26]. Due to the high and complex terrain, this region is characterized by cold and drought [27]. The annual average temperature is about 0 °C (the coldest month is January, and the hottest month is July), and the average annual precipitation ranges from 247.3 to 513.6 mm [28,29]. Because of the vast territory and highly divided topography, this area has different climate and vegetation belts [20,30].
Alpine grasslands are the most widely distributed ecosystems in Tibet, China. Alpine grasslands cover more than 70% of the total area of the Tibetan Plateau [31]. This ecosystem is dominated by alpine steppe and alpine meadow. Alpine meadow occupies approximately 31.3% of the total grassland area, in which the main species is Kobresia humilis. Alpine meadows are mainly located in Ando and Damxung in Naqu region, with cold and wet climatic conditions. Alpine steppe, which comprises approximately 38.9% of the total grassland area, is the most widely distributed alpine grassland in Tibet. Alpine steppe is mainly located in Shenza in the Naqu region, with cold and arid/semi-arid climate conditions. The main species in this grassland are Stipa purpura and Carex moorcroftii [23].
2.2. Soil Sampling
In this study, ten sampling sites were selected in eight counties—Damxung, Naqu, Nyainrong, Ando, Shenza, Baingoin, Nyima, and Gêrzê—in the Tibet Autonomous Region of China. Two alpine grasslands, i.e., alpine meadow and alpine steppe (Figure 1), were selected in this region, and five sampling sites were selected in each grassland type. The coordinates of the sampling sites are listed in Table 1. Three 1 m × 1 m quadrats were laid out at each sample site, with intervals of approximately 50 m. We removed all aboveground plants, litter, and stone on the soil surface before sampling and collected the surface (0–15 cm) soils in each quadrat. Soils from three quadrats were mixed well as a soil sample [30]. In addition, global positioning system (GPS) (Garmin MAP62CSX made in Garmin Ltd., Olathe, KS, USA) was used to measure the location and elevation of each site.

Figure 1.
Photographs of the studied areas.

Table 1.
Coordinates of the sampling sites.
2.3. Incubation Experiment
Alpine meadow and alpine steppe soils (100.0 g dry-weight) were placed in 250 mL triangular flasks for 210 days, and the soil moisture was adjusted to a 40% water-holding capacity and 25 °C to activate the soil microbes. The temperature and moisture were adapted to the optimum temperature and humidity for microorganisms during incubation [32,33]. For incubation, five experimental groups were selected from five alpine meadow sites, and five soil experimental groups were selected from five alpine steppe sites, with three replicates per experimental group. When all samples were ready, the quality of each triangular flask was labeled. To reduce humidity losses while ensuring adequate gas inside, all flasks were covered with a perforated, adherent film. When the quality of the triangular flasks decreased, water was added to the soil along the flask borders every three days to maintain constant soil moisture. After 210 days of incubation, all incubation samples were harvested using the destructive method.
2.4. Pyrolysis Gas Chromatography–Mass Spectrometry
The soil samples were analyzed for lignin components using pyrolysis gas chromatography–mass spectrometry (Py-GC/MS) technology. The Py-GC/MS operational process was conducted with reference to the methods outlined in the previously published literature [5]. The soil samples were passed through a 2 mm mesh sieve and air-dried in the shade. The lignin components were identified using a pyrolyzer (CDS5200, CDS company, Oxford, PA, USA). The procedure involved placing 25 mg of soil into quartz tubes, which had a length of 2 cm and an inside diameter of 2 mm. Then, the soil samples were heated from ambient to 700 °C for 15 s at 20 °C/ms using a Mettler microbalance (Mettler–Toledo, Greifensee, Switzerland). The pyrolyzer was used with a PerkinElmer Clarus680GC-SQ8MS System (PerkinElmer, Santa Clara, CA, USA) to pyrolyze the soil samples at a high temperature in order to obtain different compounds. The operation procedure was as follows: the temperature of the gas chromatograph was maintained at 40 °C for 3 min, increased to 280 °C at a rate of 10 °C/min, and maintained for 5 min. Helium was the carrier gas. The injector temperature was 280 °C. In the electron shock mode, the temperature of the ion source of the mass spectrometry indicator was maintained at 250 °C, and it operated at an electron energy of 70 eV.
The NIST 2014 library was used to identify the pyrolysis products, as well as reports from other researchers [5,34,35]. The pyrolysis products were identified as compounds when the surface of two characteristic ion fragments of the pyrolysis products were the same as the compounds in the NIST 2014 library. The relative percentage of lignin compounds was calculated according to the proportion of the peak area of lignin pyrolysis compounds to all the peaks measured in the chromatogram. Therefore, the relative peak area of lignin compounds was calculated by normalizing results to the all the peaks measured in the chromatogram.
2.5. Phospholipid Fatty Acid Analysis
All soil samples were stored in sealed bags at 4 °C before measurement. Lipids were extracted using one-phase chloroform, water extractant, and methanol from soils. Then, using a silicic acid column, they were fractionated into neutral lipids, phospholipids, and glycolipids. The quantification of phospholipid fatty acids (PLFAs) was conducted using gas chromatography (GC Agilent 6890-Agilent Technology, Santa Clara, CA, USA) technology, coupled with an HP7673 auto-sampler, a split injector, and a flame ionization detector (FID). The carrier gas was helium. It was operated with a pressure of 35 psi and a flow rate of 0.8 mL min−1. The detector temperature was 300 °C, and the injector temperature was 250 °C [36].
X: YωZ were used to designate phospholipid fatty acids. In the formula, X is the number of carbon bonds, Y is the number of unsaturated olefinic bonds, Z represents the keys or cyclopropane chain location, and ω is the end of methyl. In addition, i (iso) and a (anteriso) represent the branching chain; 10 Me represents the methyl group, which is tenth at the end of the pitch molecule carbon chain; and cy is a cyclopropyl group. The biomarkers used for each group of microorganisms were determined based on specific works in the research literature [37]. Bacterial PLFAs were estimated as the sum of i13:0, 14:0, i14:0, i15:0, a15:0, i15:1G, 16:0, i16:0, 16:1 2OH, 16:1G, 16:1ω5c, 16:1ω9c, i17:0, a17:0, cy17:0, 17:1ω8c, 18:1ω5c, 18:0, and cy19:0ω8c. In addition, 18:1ω9c and 20:1ω9c represent fungal PLFA biomarkers and 10Me17:0 and 10Me18:0 represent the actinobacteria groups. The Gram-positive bacterial PLFAs included 13:0 anteiso, 14:0, 14:0 iso, 15:0 anteiso, 15:0 iso, 15:1 iso G, 16:0 iso, 16:1 2OH, 16:1 iso G, 17:0 anteiso, and 17:0 iso. The Gram-negative bacterial PLFAs contained 16:1ω5c, 16:1ω9c, 17:0 cyclo, 17:1ω8c, 18:0, 18:1ω5c, and 19:0 cyclo ω8c. The soil microbial PLFAs are expressed as the absolute abundance of PLFA biomarkers of microorganisms in nmol/g. The microbial biomass was calculated using the sum of the absolute abundances of PLFAs. PLFAs that were present in >3 samples were selected for analysis [27].
2.6. Statistical Analysis
All statistical analyses were conducted using SPSS 20.0 (IBM, Chicago, IL, USA, 2011), and the significance level was p < 0.05. The differences in soil lignin and PLFA in soils from alpine meadow and alpine steppe were determined using an independent sample t test. The lignin relative abundance and soil microbial characteristics were subjected to a correlation analysis. All figures were developed using Sigmaplot® Version 10 software (Systat Software Inc., Chicago, IL, USA, 2007). A principal component analysis was conducted using Canoco 5.
3. Results
3.1. The Relative Abundance of Lignin in Alpine Grassland Soils
A total of 20 samples, comprising 5 alpine meadow soil samples and 5 alpine meadow soil samples before and after incubation, were analyzed using Py-GC/MS. The results showed that the relative abundance of lignin in the alpine meadow soil was significantly higher than that in the alpine steppe soil (Table 2). A comparison of the lignin before and after the experiment showed that its relative abundance decreased significantly in the alpine meadow soil, with values of 4.71% ± 1.04% and 1.32% ± 0.58%, respectively (p < 0.05) (Table 2). However, the relative abundance of lignin showed no significant change in the alpine steppe soil before and after the experiment (p > 0.05) (Table 2). The lignin components detected in the two types of alpine grassland soils are shown in Figure 2. Seven compounds with lignin as their precursor material were detected in the alpine grassland soils: 1-butanol, 2-ethyl-; 1-pentanol, 2-methyl-; 3-hexyne-2,5-diol; 3,5-hexadien-2-ol; 2-methoxy-4-vinylphenol; ethanol, 2-[2-(2-butoxyethoxy)ethoxy]-; and benzenemethanol, α-ethyl-4-methoxy- (Figure 2). The result showed that the relative abundance of 2-methoxy-4-vinylphenol increased and that the relative abundance of 1-pentanol, 2-methyl-, and benzenemethanol, α-ethyl-4-methoxy- decreased in the alpine grassland soils (Table 3; Figure 3).
Table 2.
The relative abundance of lignin in alpine grassland soils before and after incubation.
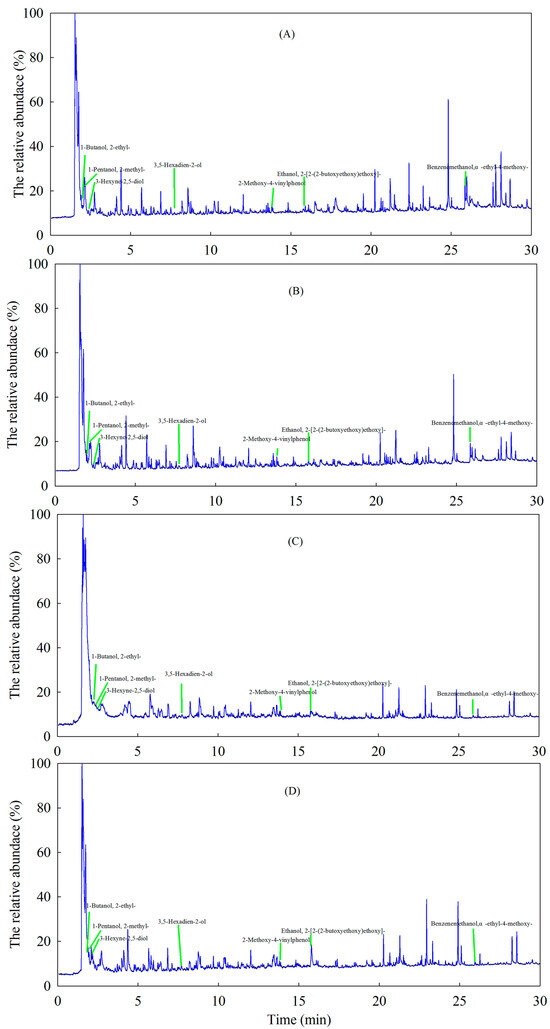
Figure 2.
The dominant compositions of lignin in alpine grassland soils based on pyrolysis gas chromatography–mass spectrometry. (A) The alpine meadow soil before incubation; (B) the alpine steppe soil before incubation; (C) the alpine meadow soil after incubation; (D) the alpine steppe soil after incubation.
Table 3.
Lignin components in alpine grassland soils before and after incubation.
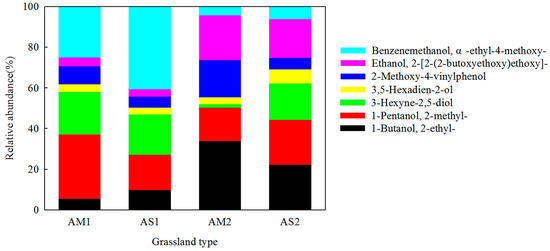
Figure 3.
The compositions of lignin in alpine grassland soils before and after incubation. AM1 and AS1 are alpine meadow and alpine steppe before incubation; AM2 and AS2 are alpine meadow and alpine steppe after incubation.
3.2. Phospholipid Fatty Acid Analysis
The absolute abundance of phospholipid fatty acids (PLFAs) in the alpine grassland soils is presented in Table 4. A significant difference was observed in the microbial PLFA biomarkers in the alpine meadow soil and alpine steppe soil, as illustrated in Figure 4. The levels of PLFA biomarkers of bacteria, fungi, actinibacteria, Gram-positive bacteria, and Gram-negative bacteria were significantly higher in the alpine meadow soils than in the alpine steppe soils (Table 4). Similarly, the fungal PLFA biomarker content in the alpine meadow soils was higher than that in the alpine steppe soils (Table 4). Overall, the total PLFA content, which serves as a proxy for the total microbial biomass, was approximately twice as high in the alpine meadow soils than in the alpine steppe soils (Table 4). The alpine grassland soil PLFAs mainly contained 18:1ω9c (fungi) and 16:0 (bacteria), with a relative abundance of more than 10% in the total PLFA. Also, 19:0 and 15:0 anteiso (bacteria) were more abundant in the alpine meadow than in the alpine steppe (Figure 4).
Table 4.
The phospholipid fatty acid biomarkers in alpine grassland soils.
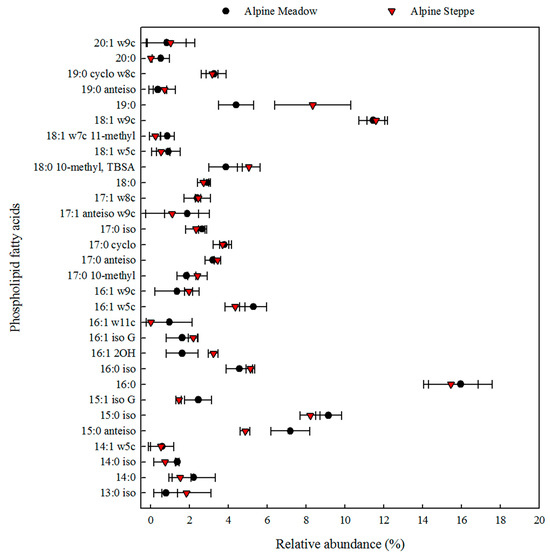
Figure 4.
The relative abundance of phospholipid fatty acids in alpine grassland soils.
3.3. The Relationship between Lignin and Phospholipid Fatty Acids
The correlation analysis showed that the relative abundance of lignin components had a significant effect on the PLFA biomarkers (Figure 5 and Figure 6). The results showed that the relative abundance of lignin was significantly correlated with the PLFA biomarkers of bacteria, fungi, Gram-positive bacteria, and Gram-negative bacteria (Figure 5). Nevertheless, the relative abundance of lignin components was not correlated with the PLFA biomarkers of actinibacteria (Figure 6). The results showed that 2-methoxy-4-vinylphenol was significantly correlated with the total PLFAs; ethanol, 2-[2-(2-butoxyethoxy)ethoxy]- and 3-hexyne-2,5-diol were significantly correlated with the PLFA biomarkers of fungi and bacteria; and 3,5-hexadien-2-ol and 1-pentanol, 2-methyl- were significantly correlated with the PLFA biomarkers of Gram-positive bacteria and Gram-negative bacteria (Figure 6).
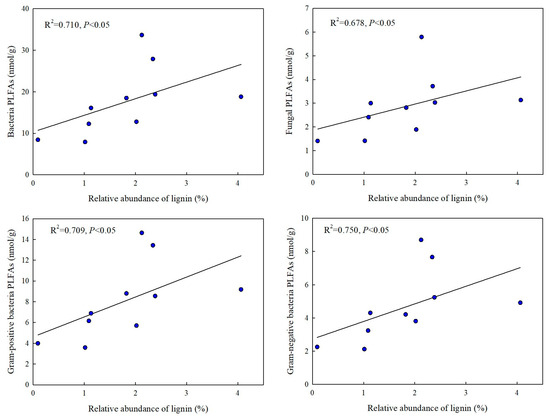
Figure 5.
The correlations between lignin and soil PLFA biomarkers in alpine grassland soils.
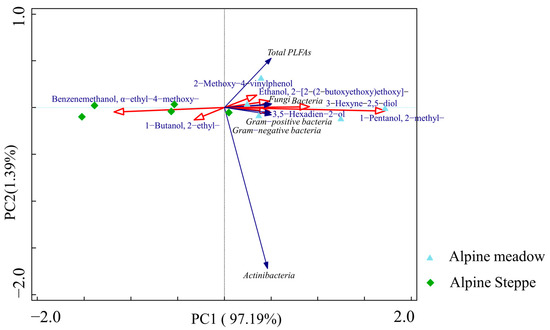
Figure 6.
Principal component analysis of lignin and PLFAs in alpine grassland soils. PLFA, phospholipid fatty acid.
4. Discussion
Lignin is a major component of plants and contributes significantly to the soil organic carbon pool [38]. In addition, lignin biomarkers can reflect the vegetation type due to plant-released monomers with a vanillyl or syringyl group (depending on the lignin type), and differences between gymnosperms, angiosperms, and grasses can be detected [15,16]. Studies have indicated that the syringyl propane monomer is the dominant component of lignin in soil in forest ecosystems and that the guaiacyl propane monomer is the main component in grassland ecosystems [5]. Vancampenhout et al. [39] provided similar evidence regarding the role of biomarkers in terms of lignin. They showed that the lignin compounds in soil under coniferous vegetation were mainly dominated by guaiacol (coniferyl-derived), while both guaiacol and syringol (sinapyl-derived) compounds were present in broadleaved forests. In this study, guaiacols, such as 2-methoxy-4-vinylphenol, were detected as lignin components, but no syringyl groups were detected (Table 3 and Figure 3). In fact, guaiacyl-type lignins have a higher resistance to biodegradation than syringyl-type lignins due to the higher degree of internal cross-linking [39]. Because of its recalcitrant nature and biomarkers, lignin is useful for studying land-use history, as reflected in the soil. Therefore, 2-methoxy-4-vinylphenol, which is a member of the guaiacol group, was preserved in the soil as a stable lignin material, and it may be an indicator of alpine grassland soil.
Scientists have found that lignin differs considerably between litters and soils, and elevation and vegetation can affect the abundance of lignin in soils [40,41]. For instance, the syringyl propane monomer is the dominant component in lignin in soil in forest ecosystems, and the guaiacyl propane monomer is the main component in lignin in soil in grassland ecosystems [35]. Lignin is dominated by guaiacol (up to 3%) in secondary succession in Imperata grassland soil [5]. In this study, the relative abundance of lignin was lower in the alpine steppe soils than in the alpine meadow soils. This may be due to the lower number of plant species and plant biomass in the alpine steppe [20,24]. In addition, 2-methoxy-4-vinylphenol, a type of guaiacyl propane lignin, was the dominant product in the alpine grassland soils. This finding is supported by the findings of Yassir and Buurman [5] and Nierop et al. [42]. During the incubation experiment, the relative abundance of 2-methoxy-4-vinylphenol increased in most samples (Table 3 and Figure 3). Therefore, this result suggests that 2-methoxy-4-vinylphenol can be stored in soil for a long time and is not easily broken down, thus indicating lignin in alpine grassland soils.
Lignin has been considered to be more recalcitrant than other organic matter (cellulose) in soils because it has a complex structure that includes multiple monomers connecting carbon–carbon and ether bonds [41]. However, new evidence has indicated that lignin can be degraded by microbes under some pedoclimatic conditions [43,44]. Martin and Haider [45,46] studied lignin decomposition in soils with different properties and vegetation types, and they showed that the initial amount of synthetic 14C-DHP lignin decreased by 19~60% after 13 weeks to 2 years. The degradation of lignin was also observed in field studies [6,47]. These studies indicated that the degraded lignin in litter constituted between 48% and 87% of the initial content after five years [6]. A decrease in lignin has been observed in agricultural soil. For example, lignin was more rapidly degraded than bulk soil organic matter [48,49], especially under low acidic pH conditions [39,50,51]. Kögel [52] found that lignin is significantly decreased at the transition to mineral soil in forests. Kiem and Kögel-Knabner [53] indicated that lignin is not part of the hard-to-degrade pool in temperate arable soils. A study on secondary succession in Imperata grassland soil also indicated that lignin underwent decomposition [5], and it showed that lignin, including 4-vinylphenol and 4-vinylguaiacol, is significantly higher in litter than in soils. These reports indicate that 4-vinylphenol can be retained in soil and is a component of plant matter. Is lignin recalcitrant or decomposed in soil? Based on this question, we incubated alpine meadow and alpine steppe soils and studied the changes in the lignin components in them. The result showed that the relative abundance of lignin decreased significantly in the alpine meadow soils (Table 2). In addition, the relative abundance of lignin components, including benzenemethanol, α-ethyl-4-methoxy-, and 1-pentanol, 2-methyl-, significantly decreased in the alpine meadow soils. However, the relative abundance of ethanol 2-[2-(2-butoxyethoxy)ethoxy]- increased during incubation (Table 3 and Figure 3). This is similar to the reports by Dao et al. [54]. They focused on lignin decomposition in the soil of the Northern Siberian tundra and discovered that the neutral sugars contained within lignin were more susceptible to degradation. Therefore, perhaps benzenemethanol, α-ethyl-4-methoxy-, and 1-pentanol, 2-methyl- are decomposed and utilized by microorganisms. They were not recalcitrant in the alpine meadow soil.
The activity of soil microbes, which produce necessary extracellular enzymes, is an important factor regulating the biological decomposition of lignin [41]. In general, white-rot fungi degrade lignin by secreting extracellular enzymes into the soil. Grandy et al. [55] found that b-1,4-N-acetyl-glucosaminidase was positively related to lignin in the light fraction in alpine and forest ecosystems, and they indicated that fungi could degrade lignin by secreting enzymes. Lignin degradation is, in fact, the result of combined fungal and bacterial activities. Some studies have shown that bacteria, including α-proteobacteria, actinibacteria, and γ-proteobacteria, can degrade lignin by secreting laccase [40]. How do the different types of bacteria, including aerobic, anaerobic, and extreme bacteria, degrade lignin? The complexity and heterogeneity of the lignin structure hinder its efficient utilization. In this study, we found that the PLFA biomarkers of bacteria, fungi, Gram-positive bacteria, and Gram-negative bacteria were significantly correlated with lignin in alpine grassland soils (Figure 5). 1-pentanol, 2-methyl decreased (Figure 3) and was significantly correlated with the PLFA biomarkers of bacteria, Gram-positive bacteria, and Gram-negative bacteria (Figure 6). This indicates that 1-pentanol, 2-methyl may have been used by bacteria in the alpine grassland soil during incubation. Researchers have found that bacteria can secrete enzymes, such as 3,4-dioxygenase, hydroxyquinol 1,2-dioxygenase, and 1,2-dioxygenase, which can reveal the lignin components [56,57]. While this study did not measure the relevant enzymes as direct evidence of the bacterial decomposition of lignin compounds, the correlation between the PLFA biomarkers of microorganisms and lignin components, as well as previous research [58], implicitly demonstrates the potential for lignin components to be utilized by microorganisms. Thus, 1-pentanol, 2-methyl could have been degraded and provided important food for microbes by secreting enzymes in the alpine meadow and alpine steppe soils. In addition, there was no correlation between the PLFA biomarkers of actinobacteria and the relative abundance of lignin components in this study (Figure 6). Although actinobacteria play an important role in soil ecosystems, they do not directly participate in the decomposition process of lignin [59]. The decomposition of lignin mainly relies on microorganisms such as bacteria, which possess specific decomposing enzymes. This finding is similar to that of Song et al. [60], Nagaraju et al. [61], and Thitinun et al. [62]. Focus should be placed on the abundance of functional bacteria and fungi, the composition of functional fungal communities, and the structure of the overall microbial community to jointly regulate the rate of lignin decomposition.
This study indicates that lignin components can be decomposed by microorganisms, as revealed by an analysis of lignin components in alpine grassland soils using Py-GC/MS. Studying the microbial degradation mechanism of lignin in the unique ecological environment of North Tibet, China, not only helps us gain a deeper understanding of soil nutrient characteristics but also provides important clues for exploring nutrient cycling patterns. Specifically, the question of whether lignin, as the second most abundant organic matter in the world [1], can be used by microorganisms has been a focus of study. However, lignin and soil organic matter degradation are highly impacted by microorganisms and environmental conditions, such as litter type, incubation temperature, and moisture [41]. We only described the factor of microorganisms in this study. Previous studies have also reported on environmental conditions. For example, on grasslands in the Mediterranean, it has been found that microbial communities have adapted to harsher conditions, and future climate change will affect plant biomass and enzyme activity, thereby influencing soil lignin decomposition and soil carbon storage mechanisms through temperature and humidity deviations [63]. So, what is the mechanism underlying the relationship between lignin decomposition and microorganisms in the alpine grassland in North Tibet, China? This will be the direction of our future research.
5. Conclusions
Lignin is known to be a recalcitrant compound. This study found that the relative abundance of lignin was significant and decreased in alpine grassland soils from North Tibet, China, after 210 days of incubation in a laboratory. The relative abundance of lignin was significantly correlated with the abundance of the PLFA biomarkers of bacteria, fungi, Gram-positive bacteria, and Gram-negative bacteria. Our findings suggest that lignin can be degraded and form an important food source for microbes in alpine grassland soils.
The present study carried out preliminary research on lignin biodegradation in alpine grassland soils using incubation. However, it is difficult to determine which specific species or populations of microorganisms decompose lignin or fully understand the process of lignin degradation based on the data obtained in this study. In the future, extracellular enzyme activity around the microbes needs to be measured, and sufficient evidence of microbes degrading lignin needs to be provided. In addition, Py-GC/MS is an analytic technique that is used to calculate the relative abundance of lignin in soil organic matter. The absolute content of lignin in soil organic matter needs to be determined in combination with other methods, including an authenticated external lignin standard. Thus, quantitative analyses and the biodegradation mechanisms of lignin in the Tibetan Plateau need to be studied further.
Author Contributions
Conceptualization, D.Y. and X.L.; methodology, S.M. and X.L.; software, S.M. and X.L.; validation, D.Y., S.M. and X.L.; writing—original draft preparation, D.Y. and S.M.; writing—review and editing, D.Y., S.M. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant No. 41877338) and the Natural Science Foundation of the Tibet Autonomous Region.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Acknowledgments
We acknowledge the Yanting Agro-ecological Experimental Station of Purple Soil for the incubation experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kubicek, C.; Druzhinina, I. Degradation of Plant Cell Wall Polymers by Fungi. Environmental and Microbial Relationships. Mycota 2007, 4, 325–340. [Google Scholar]
- Jönsson, A.S. 5-Membranes for lignin and hemicellulose recovery in pulp mills. In Membrane Technologies for Biorefining; Alberto Figoli, A., Cassano, A., Basile, A., Eds.; Woodhead Publishing: Cambridge UK, 2016; pp. 105–133. [Google Scholar]
- Otto, A.; Simpson, M.J. Evaluation of CuO oxidation parameters for determining the source and stage of lignin degradation in soil. Biogeochemistry 2006, 80, 121–142. [Google Scholar] [CrossRef]
- Vancampenhout, K.; De Vos, B.; Wouters, K.; Van Calster, H.; Swennen, R.; Buurman, P.; Deckers, J. Determinants of soil organic matter chemistry in maritime temperate forest ecosystems. Soil Biol. Biochem. 2010, 42, 220–233. [Google Scholar] [CrossRef]
- Yassir, I.; Buurman, P. Soil organic matter chemistry changes upon secondary succession in Imperata Grasslands, Indonesia: A pyrolysis–GC/MS study. Geoderma 2012, 173–174, 94–103. [Google Scholar] [CrossRef]
- Thevenot, M.; Dignac, M.; Rumpel, C. Fate of lignins in soils: A review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar] [CrossRef]
- He, Y.C.; Li, X.L.; Ben, H.X.; Xue, X.Y.; Yang, B. Lipid production form Dilute Alkali Corn Stover Lignin by Strains. ACS Sustain. Chem. Eng. 2017, 5, 2302–2311. [Google Scholar] [CrossRef]
- Filley, T.R.; Cody, G.D.; Goodell, B.; Jellison, J.; Noser, C.; Ostrofsky, A. Lignin demethylation and polysaccharide decomposition in spruce sapwood degraded by brown rot fungi. Org. Geochem. 2002, 33, 111–124. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Zhao, X.Y.; Yang, J.J.; Li, S.K.; Lu, X.X.; Li, X. Research progress on lignin degradation mechanism and influencing factors during composting. Environ. Eng. 2021, 39, 128–136. (In Chinese) [Google Scholar]
- Bugg, T.D.H.; Ahmad, M.M.; Hardiman, E.; Singh, R. The emerging role for bacteria in lignin degradation and bio-product formation. Curr. Opin. Biotechnol. 2011, 22, 394–400. [Google Scholar] [CrossRef]
- Sánchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef]
- ten Have, R.; Teunissen, P.J.M. Oxidative mechanisms involved in lignin degradation by white-rot fungi. Chem. Rev. 2001, 101, 3397–3413. [Google Scholar] [CrossRef]
- Masai, E.; Katayama, Y.; Fukuda, M. Genetic and biochemical investigations on bacterial catabolic pathways for ligninderived aromatic compounds. Biosci. Biotechnol. Biochem. 2007, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Taylor, C.R.; Pink, D.; Burton, K.; Eastwood, D.; Bending, G.D.; Bugg, T.D. Development of novel assays for lignin degradation: Comparative analysis of bacterial and fungal lignin degraders. Mol. Biosyst. 2010, 6, 815–821. [Google Scholar] [CrossRef]
- Furukawa, T.; Bello, F.O.; Horsfall, L. Microbial enzyme systems for Lignin degradation and their transcriptional regulation. Front. Biol. 2014, 9, 448–471. [Google Scholar] [CrossRef]
- Janusz, G.; Anna, P.; Justyna, S.; Urszula, W.B.; Anna, J.W.; Andrzej, P. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 6, 941–962. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wu, D.; Wei, D.; Zhao, Y.; Wu, J.; Xie, X.; Zhang, R.; Wei, Z. Improved lignocellulose-degrading performance during straw composting from diverse sources with actinomycetes inoculation by regulating the key enzyme activities. Bioresour. Technol. 2019, 271, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Pourcher, A.; Bouchez, T.; Gelhaye, E.; Peu, P. Occurrence of lignin degradation genotypes and phenotypesamong prokaryotes. Appl. Microbiol. Biot. 2014, 98, 9527–9544. [Google Scholar] [CrossRef]
- Rashid, G.M.; Taylor, C.R.; Liu, Y.; Zhang, X.; Rea, D.; Fülöp, V.; Bugg, T.D. Identification of manganese superoxide dismutase fromSphingobacterium sp. T2 as a novel bacterial enzyme for lignin oxidation. ACS Chem. Biol. 2015, 10, 2286–2294. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Liu, S.; Zhou, H.; Gao, Q.; Cao, G.; Wang, X.; Su, X.; Zhang, Y.; Tang, L.; et al. Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai-Tibetan Plateau of China. Soil Biol. Biochem. 2015, 80, 306–314. [Google Scholar] [CrossRef]
- Guo, G.; Kong, W.; Liu, J.; Zhao, J.; Du, H.; Zhang, X.; Xia, P. Diversity and distribution of autotrophic microbial community along environmental gradients in grassland soils on the Tibetan Plateau. Appl. Microbiol. Biotechnol. 2015, 99, 8765–8776. [Google Scholar] [CrossRef]
- Pan, S.; Zhang, T.; Rong, Z.; Hu, L.; Gu, Z.; Wu, Q. Population transcriptomes reveal synergistic responses of DNA polymorphism and RNA expression to extreme environments on the Qinghai-Tibetan Plateau in a predatory bird. Mol. Ecol. 2017, 26, 2993–3010. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.Y.; Ma, S.Q.; Chen, Y.C.; Degyi, Y.Z.; Jiang, H.M. Squalene found in alpine grassland soils under a harsh environment in the Tibetan Plateau, China. Biomolecules 2018, 154, 154. [Google Scholar] [CrossRef]
- Lu, X.Y.; Yan, Y.; Sun, J.; Zhang, X.; Chen, Y.C.; Wang, X.D. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion. Ecol. Evol. 2015, 5, 4492–4504. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J. China: The third pole. Nature 2008, 454, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Wang, L.; Li, X.; Zhou, J.; Chen, D.; Yao, T. Climatic and associated cryospheric, biospheric, and hydrological changes on the Tibetan Plateau: A review. Int. J. Climatol. 2018, 38, 1–17. [Google Scholar] [CrossRef]
- Ma, S.Q.; Fan, J.H.; Chen, Y.C.; Lu, X.Y. Studying greenhouse gas emissions through interactions between phospholipid fatty acid content and soil properties of alpine grassland soil in Northern Tibet, China. Glob. Ecol. Conserv. 2021, 27, e01558. [Google Scholar] [CrossRef]
- Lin, X.; Wang, S.; Ma, X.; Xu, G.; Luo, C.; Li, Y.; Jiang, G.; Xie, Z. Fluxes of CO2, CH4, and N2O in an alpine meadow affected by yak excreta on the Qinghai-Tibetan plateau during summer grazing periods. Soil Biol. Biochem. 2009, 41, 718–725. [Google Scholar] [CrossRef]
- Du, Z.Y.; Wang, X.D.; Liu, X.P.; Cai, J.J. Effects of rock fragments on yak dung greenhouse gas emissions on the Qinghai-Tibetan Plateau. J. Mt. Sci. 2016, 13, 2006–2014. [Google Scholar] [CrossRef]
- Chen, Y.C.; Ma, S.Q.; Jiang, H.M.; Degyi, Y.Z.; Cheng, G.W.; Lu, X.Y. Decomposition time, chemical traits and climatic factors determine litter-mixing effects on decomposition in an alpine steppe ecosystem in Northern Tibet. Plant Soil 2021, 459, 23–34. [Google Scholar] [CrossRef]
- Gai, J.P.; Christie, P.; Cai, X.B.; Fan, J.Q.; Zhang, J.L.; Feng, G.; Li, X.L. Occurrence and distribution of arbuscular mycorrhizal fungal species in three types of grassland community of the Tibetan Plateau. Ecol. Res. 2009, 24, 1345–1350. [Google Scholar] [CrossRef]
- Zhang, S.W.; Gong, W.; Wan, X.; Li, J.Y.; Li, Z.G.; Chen, P.; Xing, S.; Li, Z.; Liu, Y. Influence of organic matter input and temperature change on soil aggregate-associated respiration and microbial carbon use efficiency in alpine agricultural soils. Soil Ecol. Lett. 2024, 6, 230220. [Google Scholar] [CrossRef]
- Chen, Y.C.; Ma, S.Q.; Jiang, H.M.; Hu, Y.; Lu, X.Y. Influences of litter diversity and soil moisture on soil microbial communities in decomposing mixed litter of alpine steppe species. Geoderma 2020, 377, 114577. [Google Scholar] [CrossRef]
- Dorado, J.; Almendros, G.; González-Vila, F.J. Response of humic acid structure to soil tillage management as revealed by analytical pyrolysis. J. Anal. Appl. Pyrol. 2016, 117, 56–63. [Google Scholar] [CrossRef]
- Ma, S.Q.; Chen, Y.C.; Lu, X.Y.; Wang, X.D. Soil organic matter chemistry: Based on pyrolysis-gas chromatography-mass spectrometry (Py-GC/MS). Mini-Rev. Org. Chem. 2018, 15, 389–403. [Google Scholar] [CrossRef]
- Yang, B.; Pang, X.Y.; Hu, B.; Bao, W.K.; Tian, G.L. Does thinning-induced gap size result in altered soil microbial community in pine plantaton in eastern Tibetan Plateau? Ecol. Evol. 2017, 7, 2986–2993. [Google Scholar] [CrossRef]
- Jilkova, V.; Cajthaml, T.; Frouz, J. Relative importance of honeydew and resin for the microbial activity in wood ant nest and forest floor substrate—A laboratory study. Soil Biol. Biochem. 2018, 117, 1–4. [Google Scholar] [CrossRef]
- Grunewald, G.; Kaiser, K.; Jahn, R.; Guggenberger, G. Organic matter stabilization in young calcareous soils as revealed by density fractionation and analysis of lignin-derived constituents. Org. Geochem. 2006, 37, 1573–1589. [Google Scholar] [CrossRef]
- Vancampenhout, K.; De Vos, B.; Wouters, K.; Swennen, R.; Buurman, P.; Deckers, J. Organic matter of subsoil horizons under broadleaved forest: Highly processed or labile and plant-derived? Soil Biol. Biochem. 2012, 50, 40–46. [Google Scholar] [CrossRef]
- Huang, X.F.; Santhanam, N.; Badri, D.V.; Hunter, W.J.; Manter, D.K.; Decker, S.R.; Vivanco, J.M.; Reardon, K.F. Isolation and characterization of lignin-degrading bacteria from rainforest soils. Biotechnol. Bioeng. 2013, 110, 1616–1626. [Google Scholar] [CrossRef]
- Yang, S.; Yao, F.; Ye, J.; Fang, S.; Wang, Z.; Wang, R.; Zhang, Q.; Ma, R.; Wang, X.; Jiang, Y.; et al. Latitudinal pattern of soil lignin/cellulose content and the activity of their degrading enzeymes across a temperate forest ecosystem. Ecol. Indic. 2019, 102, 557–568. [Google Scholar] [CrossRef]
- Nierop, K.G.J.; Pulleman, M.M.; Marinissen, J.C.Y. Management induced organic matter differentiation in grassland and arable soil: A study using pyrolysis techniques. Soil Biol. Biochem. 2001, 33, 755–764. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Abelenda, M.S.; Buurman, P.; Camps Arbestain, M.; Kaal, J.; Martinez-Cortizas, A.; Gartzia-Bengoetxea, N.; Macías, F. Comparing NaOH-extractable organic matter of acid forest soils that differ in their pedogenic trends: A pyrolysis-GC/MS study. Eur. J. Soil Sci. 2011, 62, 834–848. [Google Scholar] [CrossRef]
- Martin, J.P.; Haider, K. Effect of concentration on decomposition of some 14C-labeled phenolic compounds, benzoic acid, glucose, cellulose, wheat straw, and Chlorella protein in soil. Soil Sci. Soc. Am. J. 1979, 43, 917–920. [Google Scholar] [CrossRef]
- Martin, J.P.; Haider, K. Microbial degradation and stabilization of 14C-labeled lignins, phenols, and phenolic polymers in relation to soil humus formation. In Lignin Biodegradation: Microbiology, Chemistry and Applications; Kirk, T.K., Higuchi, T., Chang, H.-M., Eds.; CRC Press: Boca Raton, FL, USA, 1980; Volume I, pp. 77–100. [Google Scholar]
- Osono, T.; Takeda, H. Effects of organic mineral chemical quality and mineral nitrogen addition on lignin and holocellulose decomposition of beech leaf litter by Xylaria sp. Eur. J. Soil Biol. 2001, 37, 17–23. [Google Scholar] [CrossRef]
- Dignac, M.F.; Bahri, H.; Rumpel, C.; Rasse, D.P.; Bardoux, G.; Balesdent, J.; Girardin, C.; Chenu, C.; Mariotti, A. Carbon-13 natural abundance as a tool to study the dynamics of lignin monomers in soil: An appraisal at the Closeaux experimental field (France). Geoderma 2005, 128, 3–17. [Google Scholar] [CrossRef]
- Heim, A.; Schmidt, M.W.I. Lignin turnover in arable soil and grassland analysed with two different labelling approaches. Eur. J. Soil Sci. 2007, 58, 599–608. [Google Scholar] [CrossRef]
- Bracewell, J.M.; Robertson, G.W. Quantitative comparison of the nitrogen-containing pyrolysis products and amino acid composition of soil humic acids. J. Anal. Appl. Pyrolysis 1984, 6, 19–29. [Google Scholar] [CrossRef]
- Buurman, P.; Roscoe, R. Different chemical compostion of free light, occluded light and extractable SOM fractions in soils of Cerrade and tilled and untilled fields, Minas Gerais Brazil, a pyrolysis-GC/MS study. Ruropean J. Soil Sci. 2011, 62, 253–266. [Google Scholar] [CrossRef]
- Kögel, I. Estimation and decomposition pattern of the lignin component in forest humus layers. Soil Biol. Biochem. 1986, 18, 589–594. [Google Scholar] [CrossRef]
- Kiem, R.; Kögel-Knabner, I. Contribution of lignin and polysaccharides to the refractory carbon pool in C-depleted arable soils. Soil Biol. Biochem. 2003, 35, 101–118. [Google Scholar] [CrossRef]
- Dao, T.T.; Mikutta, R.; Sauheitl, L.; Gentsch, N.; Shibistova, O.; Wild, B.; Schnecker, J.; Bárta, J.; Čapek, P.; Gittel, A.; et al. Lignin Preservation and Microbial Carbohydrate Metabolism in Permafrost Soils. J. Geophys. Res-Biogeo. 2022, 127, e2020JG006181. [Google Scholar] [CrossRef]
- Grandy, A.S.; Neff, J.C.; Weintraub, M.N. Carbon structure and enzyme activities in alpine and forest ecosystems. Soil Biol. Biochem. 2007, 39, 2701–2711. [Google Scholar] [CrossRef]
- Xu, C.F.; Su, X.; Wang, J.H.; Zhang, F.Z.; Shen, G.N.; Yuan, Y.; Yan, L.; Tang, H.Z.; Song, F.Q.; Wang, W.D. Characteristics and functional bacteria in a microbial consortium for rice straw lignin-degrading. Bioresour. Technol. 2021, 331, 125066. [Google Scholar] [CrossRef]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.T. Lignin degradation by microorganisms: A review. Biotechnol. Progr. 2021, 38, e3226. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.Y.; Fan, Z.; Peng, W.X.; Du, H.; Hu, P.L. Source to Sink of Lignin Phenols in a Subtropical Forest of Southwest China. Forests 2023, 14, 1701. [Google Scholar] [CrossRef]
- Abdellatif Aya, A.M.; Gebily Doha, A.S.; Maissara, E. Transforming Roles of Actinobacteria in Sustainable Agriculture: From Soil Health and Plant Productivity Perspective; Sahu, P.K., Thakur, B., Kaur, S., Eds.; Springer Nature: Singapore, 2024; pp. 299–338. [Google Scholar]
- Song, A.; Zhang, J.; Xu, D.; Wang, E.; Bi, J.; Asante-Badu, B.; Njyenawe, M.C.; Sun, M.; Xue, P.; Wang, S.; et al. Keystone microbial taxa drive the accelerated decompositions of cellulose and lignin by long-term resource enrichments. Sci. Total Environ. 2022, 842, 156814. [Google Scholar] [CrossRef]
- Nagaraju, Y.; Mahadevaswamy; Gowder, S.B.; Triveni, S. Lignin-Degrading Microorganisms from Organic Soils. Int. J. Plant Soil Sci. 2021, 33, 78–85. [Google Scholar] [CrossRef]
- Thitinun, S.; Esther, A.; Surang, C.; Tanaporn, P.; Wethaka, L.; Puey, O.; Phurt, H. Evaluating lignin degradation under limited oxygen conditions by bacterial isolates from forest soil. Sci. Rep. 2024, 14, 13350. [Google Scholar]
- San-Emeterio, L.M.; Hidalgo-Galvez, M.D.; de la Rosa, J.M.; P’erez-Ramos, I.; González-Pérez, J.A. Impact of future scenarios of climate change on lignin dynamics in soil: A case study in a Mediterranean savannah. Sci. Total Environ. 2024, 946, 174317. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).