Ancient Genome of Broomcorn Millet from Northwest China in Seventh Century CE: Shedding New Light to Its Origin and Dispersal Patterns

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Study Area
3. Materials and Methods
4. Bioinformatics and Statistics
5. Results
5.1. Ancient Dataset
5.2. Closest Genetic Affinity of Ancient Samples with Modern Samples from Ningxia
5.3. Gene Flow from Ancient to Modern Samples
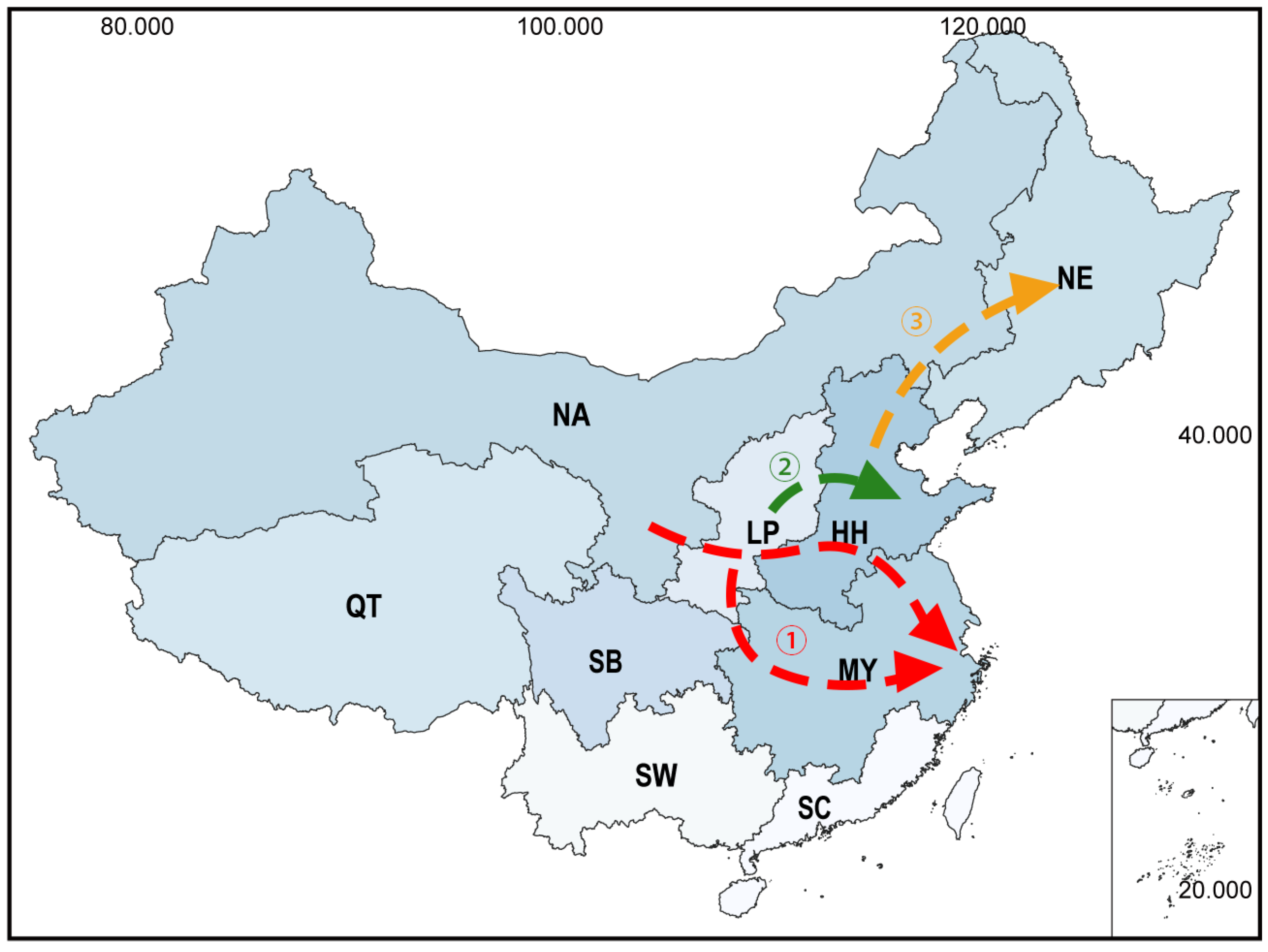
6. Discussion
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, F.H.; Dong, G.H.; Zhang, D.J.; Liu, X.Y.; Jia, X.; An, C.B.; Ma, M.M.; Xie, Y.W.; Barton, L.; Ren, X.Y.; et al. Agriculture Facilitated Permanent Human Occupation of the Tibetan Plateau after 3600 B.P. Science 2015, 347, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T.; et al. Early Millet Use in Northern China. Proc. Natl. Acad. Sci. USA 2012, 109, 3726–3730. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhang, J.; Liu, K.; Wu, N.; Li, Y.; Zhou, K.; Ye, M.; Zhang, T.; Zhang, H.; Yang, X.; et al. Earliest Domestication of Common Millet (Panicum miliaceum) in East Asia Extended to 10,000 Years Ago. Proc. Natl. Acad. Sci. USA 2009, 106, 7367–7372. [Google Scholar] [CrossRef]
- Ventresca-Miller, A.R.; Wilkin, S.; Smithers, R.; Larson, K.; Spengler, R.; Haruda, A.; Kradin, N.; Bazarov, B.; Miyagashev, D.; Odbaatar, T.; et al. Adaptability of Millets and Landscapes: Ancient Cultivation in North-Central Asia. Agronomy 2023, 13, 2848. [Google Scholar] [CrossRef]
- Dong, G.; Yang, Y.; Han, J.; Wang, H.; Chen, F. Exploring the History of Cultural Exchange in Prehistoric Eurasia from the Perspectives of Crop Diffusion and Consumption. Sci. China Earth Sci. 2017, 60, 1110–1123. [Google Scholar] [CrossRef]
- Spengler, R.N.; Ryabogina, N.; Tarasov, P.E.; Wagner, M. The Spread of Agriculture into Northern Central Asia: Timing, Pathways, and Environmental Feedbacks. Holocene 2016, 26, 1527–1540. [Google Scholar] [CrossRef]
- Ma, M.; Dong, J.; Yang, Y.; Jones, M.K.; Wang, J.; Chen, G.; Li, G.; He, L.; Wei, W.; Xu, Y.; et al. Isotopic Evidence Reveals the Gradual Intensification of Millet Agriculture in Neolithic Western Loess Plateau. Fundam. Res. 2023, in press. [Google Scholar] [CrossRef]
- Long, T.; Wagner, M.; Demske, D.; Leipe, C.; Tarasov, P.E. Cannabis in Eurasia: Origin of Human Use and Bronze Age Trans-Continental Connections. Veget. Hist. Archaeobot. 2017, 26, 245–258. [Google Scholar] [CrossRef]
- Flad, R.; Shuicheng, L.; Xiaohong, W.; Zhijun, Z. Early Wheat in China: Results from New Studies at Donghuishan in the Hexi Corridor. Holocene 2010, 20, 955–965. [Google Scholar] [CrossRef]
- Yang, Y.; Dong, G.; Zhang, S.; Cui, Y.; Li, H.; Chen, G.; Dodson, J.; Chen, F. Copper Content in Anthropogenic Sediments as a Tracer for Detecting Smelting Activities and Its Impact on Environment during Prehistoric Period in Hexi Corridor, Northwest China. Holocene 2017, 27, 282–291. [Google Scholar] [CrossRef]
- Hunt, H.V.; Campana, M.G.; Lawes, M.C.; Park, Y.-J.; Bower, M.A.; Howe, C.J.; Jones, M.K. Genetic Diversity and Phylogeography of Broomcorn Millet (Panicum miliaceum L.) across Eurasia. Mol. Ecol. 2011, 20, 4756–4771. [Google Scholar] [CrossRef]
- Zou, C.; Li, L.; Miki, D.; Li, D.; Tang, Q.; Xiao, L.; Rajput, S.; Deng, P.; Peng, L.; Jia, W.; et al. The Genome of Broomcorn Millet. Nat. Commun. 2019, 10, 436. [Google Scholar] [CrossRef]
- Li, C.; Liu, M.; Sun, F.; Zhao, X.; He, M.; Li, T.; Lu, P.; Xu, Y. Genetic Divergence and Population Structure in Weedy and Cultivated Broomcorn Millets (Panicum miliaceum L.) Revealed by Specific-Locus Amplified Fragment Sequencing (SLAF-Seq). Front. Plant Sci. 2021, 12, 688444. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, M.; Li, C.; Sun, F.; Lu, P.; Meng, F.; Zhao, X.; He, M.; Wang, F.; Zhu, X.; et al. Domestication and Spread of Broomcorn Millet (Panicum miliaceum L.) Revealed by Phylogeography of Cultivated and Weedy Populations. Agronomy 2019, 9, 835. [Google Scholar] [CrossRef]
- Chen, N.; Zhang, Z.; Hou, J.; Chen, J.; Gao, X.; Tang, L.; Wangdue, S.; Zhang, X.; Sinding, M.-H.S.; Liu, X.; et al. Evidence for Early Domestic Yak, Taurine Cattle, and Their Hybrids on the Tibetan Plateau. Sci. Adv. 2023, 9, eadi6857. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Fu, W.; Cai, D.; Heller, R.; Zheng, Z.; Wen, J.; Li, H.; Wang, X.; Alshawi, A.; Sun, Z.; et al. Ancient Genomes Reveal the Evolutionary History and Origin of Cashmere-Producing Goats in China. Mol. Biol. Evol. 2020, 37, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Mascher, M.; Schuenemann, V.J.; Davidovich, U.; Marom, N.; Himmelbach, A.; Hübner, S.; Korol, A.; David, M.; Reiter, E.; Riehl, S.; et al. Genomic Analysis of 6,000-Year-Old Cultivated Grain Illuminates the Domestication History of Barley. Nat. Genet. 2016, 48, 1089–1093. [Google Scholar] [CrossRef]
- Swarts, K.; Gutaker, R.M.; Benz, B.; Blake, M.; Bukowski, R.; Holland, J.; Kruse-Peeples, M.; Lepak, N.; Prim, L.; Romay, M.C.; et al. Genomic Estimation of Complex Traits Reveals Ancient Maize Adaptation to Temperate North America. Science 2017, 357, 512–515. [Google Scholar] [CrossRef]
- Ramos-Madrigal, J.; Runge, A.K.W.; Bouby, L.; Lacombe, T.; Samaniego Castruita, J.A.; Adam-Blondon, A.-F.; Figueiral, I.; Hallavant, C.; Martínez-Zapater, J.M.; Schaal, C.; et al. Palaeogenomic Insights into the Origins of French Grapevine Diversity. Nat. Plants 2019, 5, 595–603. [Google Scholar] [CrossRef]
- Wu, X.; Ding, B.; Zhang, B.; Feng, J.; Wang, Y.; Ning, C.; Wu, H.; Zhang, F.; Zhang, Q.; Li, N.; et al. Phylogenetic and Population Structural Inference from Genomic Ancestry Maintained in Present-Day Common Wheat Chinese Landraces. Plant J. 2019, 99, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Green, R.E.; Krause, J.; Briggs, A.W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H.; Zhai, W.; Fritz, M.H.-Y.; et al. A Draft Sequence of the Neandertal Genome. Science 2010, 328, 710–722. [Google Scholar] [CrossRef]
- Kjær, K.H.; Winther Pedersen, M.; De Sanctis, B.; De Cahsan, B.; Korneliussen, T.S.; Michelsen, C.S.; Sand, K.K.; Jelavić, S.; Ruter, A.H.; Schmidt, A.M.A.; et al. A 2-Million-Year-Old Ecosystem in Greenland Uncovered by Environmental DNA. Nature 2022, 612, 283–291. [Google Scholar] [CrossRef]
- Richards, S.M.; Li, L.; Breen, J.; Hovhannisyan, N.; Estrada, O.; Gasparyan, B.; Gilliham, M.; Smith, A.; Cooper, A.; Zhang, H. Recovery of Chloroplast Genomes from Medieval Millet Grains Excavated from the Areni-1 Cave in Southern Armenia. Sci. Rep. 2022, 12, 15164. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Dong, Y.; Liu, M.; Lu, P.; Li, W.; Wang, Y.; Cui, X.; Zhou, H.; Xu, Y. Ancient DNA Analysis of Panicum miliaceum (Broomcorn Millet) from a Bronze Age Cemetery in Xinjiang, China. Veget. Hist. Archaeobot. 2016, 25, 469–477. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, B.; Liu, R.; Jiang, H.; Yang, Y.; Ye, Q.; Li, R.; Wei, W.; Chen, G.; Dong, G. The Lifestyle of Tuyuhun Royal Descendants: Identification and Chemical Analysis of Buried Plants in the Chashancun Cemetery, Northwest China. Front. Plant Sci. 2022, 13, 972891. [Google Scholar] [CrossRef]
- Gansu Provincial Institute of Cultural Relics and Archaeology; Wuwei Municipal Institute of Cultural Relics and Archaeology; Tianzhu Tibetan Autonomous County Museum. Tang Dynasty Tuyuhun Royal Tombs in Wuwei, Gansu. Chin. Archaeol. 2023, 23, 87–105. [Google Scholar] [CrossRef]
- Ge, S.; Wang, J.; Jiang, C. Projection of the Precipitation-Induced Landslide Risk in China by 2050. Environ. Res. Lett. 2024, 19, 024048. [Google Scholar] [CrossRef]
- Meyer, M.; Kircher, M. Illumina Sequencing Library Preparation for Highly Multiplexed Target Capture and Sequencing. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5448. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Lindgreen, S.; Orlando, L. AdapterRemoval v2: Rapid Adapter Trimming, Identification, and Read Merging. BMC Res. Notes 2016, 9, 88. [Google Scholar] [CrossRef]
- Chen, S. Ultrafast One-Pass FASTQ Data Preprocessing, Quality Control, and Deduplication Using Fastp. iMeta 2023, 2, e107. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Tobler, R.; Llamas, B.; Souilmi, Y. Additional Evaluations Show That Specific BWA-Aln Settings Still Outperform BWA-Mem for Ancient DNA Data Alignment. Ecol. Evol. 2021, 11, 18743–18748. [Google Scholar] [CrossRef]
- Jónsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.F.; Orlando, L. mapDamage2.0: Fast Approximate Bayesian Estimates of Ancient DNA Damage Parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Picard Tools—By Broad Institute. Available online: https://broadinstitute.github.io/picard/ (accessed on 15 January 2024).
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced Multi-Sample Quality Control for High-Throughput Sequencing Data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Jun, G.; Wing, M.K.; Abecasis, G.R.; Kang, H.M. An Efficient and Scalable Analysis Framework for Variant Extraction and Refinement from Population Scale DNA Sequence Data. Genome Res. 2015, 25, 918–925. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. rMVP: A Memory-Efficient, Visualization-Enhanced, and Parallel-Accelerated Tool for Genome-Wide Association Study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef] [PubMed]
- He, W. 2023 “VCF2Dis” Github. 2023. Available online: https://github.com/BGI-Shenzhen/VCF2Dis (accessed on 21 August 2024).
- Baum, B.R. PHYLIP: Phylogeny Inference Package. Version 3.2. Joel Felsenstein. Q. Rev. Biol. 1989, 64, 539–541. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Petr, M.; Vernot, B.; Kelso, J. Admixr—R Package for Reproducible Analyses Using ADMIXTOOLS. Bioinformatics 2019, 35, 3194–3195. [Google Scholar] [CrossRef]
- Deng, Z.; Hung, H.; Fan, X.; Huang, Y.; Lu, H. The Ancient Dispersal of Millets in Southern China: New Archaeological Evidence. Holocene 2018, 28, 34–43. [Google Scholar] [CrossRef]
- Wang, T.; Wei, D.; Chang, X.; Yu, Z.; Zhang, X.; Wang, C.; Hu, Y.; Fuller, B.T. Tianshanbeilu and the Isotopic Millet Road: Reviewing the Late Neolithic/Bronze Age Radiation of Human Millet Consumption from North China to Europe. Natl. Sci. Rev. 2019, 6, 1024–1039. [Google Scholar] [CrossRef]
- Wei, W.; Shi, Z.; Lu, Y.; Du, L.; Zhang, J.; Zheng, G.; Ma, M. Alteration in Plant-Based Subsistence and Its Influencing Factors from Late Neolithic to Historical Periods in Hexi Corridor, Northwestern China: Archaeobotanical Evidence. Land 2024, 13, 419. [Google Scholar] [CrossRef]
- Hunt, H.V.; Jiang, H.; Liu, X.; Lister, D.L.; Abuduresule, Y.; Li, W.; Wang, C.; Jones, M.K. Did Crops Expand in Tandem with Culinary Practices from Their Region of Origin? Evidence from Ancient DNA and Material Culture. Antiquity 2024, 98, 12–29. [Google Scholar] [CrossRef]
- Gu, Z.; Gao, Y.; Wang, Y.; Yang, J.; Ran, J.; Yang, X.; Shargan, W.; Pedersen, M.W.; Sheng, G.; Wang, Y.; et al. Food Resources of the Khog Gzung Site on the Tibetan Plateau Revealed by Sedimentary Ancient DNA. Sci. China Earth Sci. 2023, 66, 840–851. [Google Scholar] [CrossRef]
- Jinsun, L. The Refuge and Migration of Tuyuhun’s Refugees in Wu Zetian Period. Hist. Stud. Anc. Mediev. China 2020, 56, 237–260. [Google Scholar] [CrossRef]
- A Study of Livelihood and City Policy in Chang’an, the Capital of Tang Dynasty (618–907)-All Databases. Available online: https://webofscience.clarivate.cn/wos/alldb/full-record/PQDT:61425067 (accessed on 21 August 2024).
- Zhang, H.; Cai, Y.; Tan, L.; Cheng, H.; Qin, S.; An, Z.; Edwards, R.L.; Ma, L. Large Variations of 13C Values in Stalagmites from Southeastern China during Historical Times: Implications for Anthropogenic Deforestation. Boreas 2015, 44, 511–525. [Google Scholar] [CrossRef]
- Jia, D.; Li, Y.; Fang, X. Complexity of Factors Influencing the Spatiotemporal Distribution of Archaeological Settlements in Northeast China over the Past Millennium. Quat. Res. 2018, 89, 413–424. [Google Scholar] [CrossRef]
- Li, R.; Li, B.; Chen, W.; Liu, P.; Xie, M.; Zhang, Y.; Wang, S.; Li, Y.; Dong, G. The Alteration from Agricultural to Nomadic Regimes Resulted in Human Livelihood Transformation in North-Central China during the 12th Century: The Archaeobotanical Evidence. Front. Plant Sci. 2022, 13, 978147. [Google Scholar] [CrossRef] [PubMed]
- The Study of the Migration of the Big Locust Tree in Hongtong. Chin. Local Chron. 2003, S1, 26–30.
- Liu, J. Beyond Categories and Boundaries: Transmarine Mobility and Coastal Governance of Northeast Asia in the Sixteenth Century. J. R. Asiat. Soc. 2023, 33, 133–154. [Google Scholar] [CrossRef]
- Ye, Y.; Fang, X.; Aftab U Khan, M. Migration and Reclamation in Northeast China in Response to Climatic Disasters in North China over the Past 300 Years. Reg. Environ. Change 2012, 12, 193–206. [Google Scholar] [CrossRef]
- Fang, X.; Ye, Y.; Zeng, Z. Extreme Climate Events, Migration for Cultivation and Policies: A Case Study in the Early Qing Dynasty of China. Sci. China Ser. D 2007, 50, 411–421. [Google Scholar] [CrossRef]
- Campbell, C. Swallows and Settlers: The Great Migration from North China to Manchuria. J. Asian Stud. 2005, 64, 1004–1005. [Google Scholar] [CrossRef]
- Kistler, L.; Bieker, V.C.; Martin, M.D.; Pedersen, M.W.; Ramos Madrigal, J.; Wales, N. Ancient Plant Genomics in Archaeology, Herbaria, and the Environment. Annu. Rev. Plant Biol. 2020, 71, 605–629. [Google Scholar] [CrossRef] [PubMed]
- Meiri, M.; Bar-Oz, G. Unraveling the Diversity and Cultural Heritage of Fruit Crops through Paleogenomics. Trends Genet. 2024, 40, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Burbano, H.A.; Gutaker, R.M. Ancient DNA Genomics and the Renaissance of Herbaria. Science 2023, 382, 59–63. [Google Scholar] [CrossRef]
- Motuzaitė Matuzevičiūtė, G.; Laužikas, R. A Brief History of Broomcorn Millet Cultivation in Lithuania. Agronomy 2023, 13, 2171. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Wang, Y.; Lu, Y.; Xu, Y.; Liu, B.; Yang, Y.; Chen, G.; Wang, H.; Huang, Z.; Cai, Y.; et al. Ancient Genome of Broomcorn Millet from Northwest China in Seventh Century CE: Shedding New Light to Its Origin and Dispersal Patterns. Agronomy 2024, 14, 2004. https://doi.org/10.3390/agronomy14092004
Sun X, Wang Y, Lu Y, Xu Y, Liu B, Yang Y, Chen G, Wang H, Huang Z, Cai Y, et al. Ancient Genome of Broomcorn Millet from Northwest China in Seventh Century CE: Shedding New Light to Its Origin and Dispersal Patterns. Agronomy. 2024; 14(9):2004. https://doi.org/10.3390/agronomy14092004
Chicago/Turabian StyleSun, Xiaolan, Yifan Wang, Yongxiu Lu, Yongxiang Xu, Bingbing Liu, Yishi Yang, Guoke Chen, Hongru Wang, Zihao Huang, Yuanyang Cai, and et al. 2024. "Ancient Genome of Broomcorn Millet from Northwest China in Seventh Century CE: Shedding New Light to Its Origin and Dispersal Patterns" Agronomy 14, no. 9: 2004. https://doi.org/10.3390/agronomy14092004