Abstract
Oat (Avena sativa) is one of the most important cereal crops and cool-season forage grasses in the world. The calmodulin-binding transcription activator (CAMTA) gene family is one of the largest families in plants, and it plays vital roles in multiple biological processes. However, the CAMTA genes in oats, especially those involved in abiotic stress, have not yet been elucidated. Herein, our findings reveal the presence of 20 distinct AsCAMTA genes, which were clustered into three subfamilies based on their gene structure and conserved motifs, indicating functional similarities within each subgroup. Chromosomal mapping indicated an uneven distribution across 10 chromosomes, suggesting a complex evolutionary history marked by potential gene duplication events. The results showed that most AsCAMTA genes contained stress-related cis-elements. The study further investigated the expression patterns of these genes under abiotic stress conditions utilizing RT-qPCR analysis. The results identified three AsCAMTA genes (AsCAMTA5, AsCAMTA7, and AsCAMTA19) that exhibited significant up-regulation under salt stress, with AsCAMTA7 also showing a marked increase in expression under drought stress. These findings suggest a pivotal role of AsCAMTA5, AsCAMTA7, and AsCAMTA19 genes in mediating the responses to various abiotic stresses by integrating multiple stress signals in oats. This investigation provides valuable insights into the potential functions of AsCAMTA genes in the stress response mechanisms of oats, laying a foundation for further functional studies aimed at enhancing abiotic stress tolerance in crops.
1. Introduction
Avena sativa L., commonly referred to as oats, holds a significant position within the spectrum of cereal crops [1,2]. It is an annual cereal and fodder crop of the Poaceae family that is grown in approximately 42 countries worldwide [3]. Oat is also widely grown in North China due to its high stress resistance and feed value. Not only does it have nutritious and health-enhancing properties, but it also has a good ability to adapt to environments such as salinity, cold and drought [4]. Beyond its foundational role as a food source and a component of livestock nutrition, oats are pivotal in promoting sustainable agricultural methodologies, bolstering soil health, and facilitating ecological equilibrium [5,6]. The cultivation of oats is critical for the rehabilitation of deteriorated natural grasslands and the amelioration of saline soils, positioning it as an integral crop capable of addressing the imperatives of food security, supporting grassland animal husbandry, and preserving environmental integrity [7,8]. With the release of oat genomic data, researchers can now conduct extensive studies of the molecular mechanisms that regulate growth, development, and stress in oats at the genomic level.
Abiotic stress factors, such as drought and salt, seriously affect the growth and development process of crops and severely limit plant growth. Drought and salt stress are among the most important abiotic stress factors that hinder plant growth and development, and they are also among the most common forms of plant damage. Drought causes water loss in plant cells, leading to significant changes in plant physiology and biochemistry [9,10]. Therefore, improving plant stress tolerance has become the focus of the current crop breeding research. When plants are subjected to abiotic stress, the stress induces the expression of plant-related genes, which participate in or activate regulatory networks to respond to the abiotic stress and minimize the damage caused by the outside world in order to protect the plants themselves. Genes that respond to abiotic stress can be divided into two categories: first, functional genes that respond to drought, such as water channel proteins, LEA proteins, etc. Studies have found that the expression of genes related to plant resistance to adversity is elevated under abiotic stress [9]. The second is drought-responsive regulatory genes, such as transcription factors and protein kinases, which can rapidly respond to drought stress and regulate gene expression. Numerous studies have shown that transcription factors are involved in responding to abiotic stresses in plants, while increasing plant stress tolerance, such as WRKY [10], NAC, MYB [11], bZIP [12], etc. These have important biological functions in plant species and can regulate plant resistance to adversity. The intricate web of intracellular signaling pathways, central to cellular communication and response mechanisms across diverse biological contexts, prominently features calcium ions (Ca2+) as pivotal messengers [13]. These ions play an indispensable role in facilitating plant adaptation to both biotic and abiotic stresses, underpinning essential growth and developmental processes [14]. The specificity and versatility of Ca2+ signaling lie in its ability to be intricately decoded by a sophisticated ensemble of sensor proteins within the plant cell. This ensemble comprises three primary types: calmodulin (CaM) and CaM-like proteins, calcium-dependent protein kinases (CDPKs), and calcineurin B-like (CBL) proteins [15]. These sensor proteins are not mere passive conveyors of signals but are deeply integrated into the mechanisms of Ca2+-dependent transcriptional regulation, thus influencing a broad spectrum of physiological outcomes.
Among the pantheon of Ca2+ sensors, CaM stands out as the most critical receptor, engaging in complex formation with Ca2+ to interact with a wide array of target proteins. This array spans enzymes involved in metabolic processes, protein kinases, phosphatases, ion channels, transporters, transcription factors, and molecular motors, highlighting CaM’s central role in cellular signaling and regulation [16,17,18,19,20]. The interaction between Ca2+ and CaM is a linchpin in the activation of over 90 transcription factors, including the calmodulin-binding transcription activator (CAMTA), underscoring the extensive reach and impact of Ca2+ signaling within plant cellular functions [21,22].
The CAMTA family, characterized by its dependency on the Ca2+/CaM complex for transcription factor activity [23], is pivotal in the orchestration of plant responses to environmental stresses and developmental cues. These transcription factors are distinguished by their unique functional domains, which delineate their indispensable roles in stress response mechanisms across a variety of species [24]. Following its initial discovery in tobacco, a plethora of studies have ventured into the genomic and functional landscape of the CAMTA gene family across different plant species. CAMTAs contain a conserved family of transcription factors in a variety of multicellular eukaryotes that may respond to calcium signaling by directly binding to calmodulin [25]. CAMTA1 and CAMTA2 work synergistically with CAMTA3 at low temperatures (4 °C) to induce multiple gene expressions [26]. These investigations have illuminated the significant roles CAMTA genes play in bolstering stress tolerance and modulating plant immunity, showcasing their essential contributions to plant resilience and adaptability [27,28,29,30].
Despite oats’ agricultural and nutritional significance, there remains untapped potential in understanding the genetic mechanisms underlying their stress tolerance and nutritional content. This research endeavor aims to bridge this knowledge gap by delving into the CAMTA gene family within oats, leveraging bioinformatics tools for comprehensive gene identification and characterization. Through the detailed analyses of phylogenetic relationships, gene structures, and chromosomal distributions, coupled with quantitative assessments of gene expression under drought and salt stress conditions via real-time PCR (qRT-PCR), this study seeks to unravel the functional dynamics of AsCAMTA genes. This study provides insights into the potential functional role of CAMTA genes in oats. The comprehensive analysis was helpful in screening the CAMTA gene of oats to further identify the function and genetic improvement of environmental resistance in oats.
2. Materials and Methods
2.1. Identification of CAMTA Genes
Genomic data and Gene Feature Format (GFF) annotations for hexaploid A. sativa (cultivar ‘Sang’) were acquired from the Ensembl Plants database (https://plants.ensembl.org/index.html, accessed on 22 November 2023, Accession number: GCA 910574605). To better understand the historical dynamics of the CAMTA gene during the evolution of the genus Avena, the whole genome sequences (DNA and Gff3) of two genomes, Avena insularis (tetraploid, CCDD, 2n = 4x = 28,) and Avena longiglumis (diploid, AA, 2n = 2x = 14), were downloaded from the results of a joint sequencing study by PepsiCo and Corteva Agriscience (https://wheat.pw.usda.gov/GG3, accessed on 25 November 2023, Accession number: GCA_910574615.1 and GCA_910589755.1). To identify the potential CAMTA transcription factors in A. sativa, the reference sequences of the Arabidopsis CAMTA gene family were retrieved from the Arabidopsis thaliana database TAIR (https://www.arabidopsis.org, accessed on 25 November 2023). These reference sequences were then subjected to BLAST alignment using the TBtools software v2.112 [31], with the subsequent comparison of the results against the UniProt database (https://www.uniprot.org, accessed on 5 January 2024) for validation through a blastp analysis. The sequences lacking the characteristic CAMTA domain were excluded. Ultimately, 20 AsCAMTA genes were identified and designated AsCAMTA1 through AsCAMTA20 based on their chromosomal positioning. Seven CAMTA genes of A. insularis and 20 CAMTA genes of A. longiglumis were obtained.
2.2. Gene Structure, Motif Composition, and Chromosomal Localization of the AsCAMTA Gene Family
Gene structures, including exons, coding sequences, and untranslated regions (UTRs), were extracted from GFF3 files using TBtools software v2.112. The physicochemical properties of the encoded proteins were predicted using the ExPASy online platform (https://www.expasy.org, accessed on 25 February 2024), and their subcellular localization was determined via Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/, accessed on 25 February 2024) [32]. Conserved motifs within the AsCAMTA proteins were identified using the MEME Suite (https://meme-suite.org/meme/tools/meme, accessed on 6 January 2024) [33]. The phylogenetic tree was constructed by neighbor-joining (NJ) after trimming the sequences that did not align with the shortest sequence using the MEGA11 software, and the full-length sequences of the proteins were constructed using Bootstrap with 1000 repetitions and visualized using TBtools software v2.112. The chromosomal locations of the AsCAMTA genes were derived from the A. sativa genome GFF file and analyzed with TBtools software v2.112 for mapping and nomenclature consistency.
2.3. Collinearity and Phylogenetic Analysis of CAMTA Genes
A comprehensive phylogenetic analysis was conducted comparing the full sequence set of predicted AsCAMTA genes against those of A. thaliana, Oryza sativa, A. insularis, and A. longiglumis. The phylogenetic tree was constructed by the bootstrap of 1000 repetitions of the sequences after trimming their ends that do not align with the shorter sequences using the MEGA11 software by the maximum likelihood (ML) method. The phylogenetic tree was analyzed by iTOL (http://itol.embl.de, accessed on 18 February 2024). These sequences are also provided in a fasta file as Supplementary Material. The intraspecific sequence alignments of the AsCAMTA gene family were executed using BLAST, followed by collinearity analysis via MCScanX [34], with collinearity diagrams generated by TBtools software v2.112 to elucidate gene duplication events.
2.4. Plant Material and Stress Treatments
The ‘Qinghai No. 444’ oat cultivar (Qinghai Academy of Animal Husbandry and Veterinary Sciences) with good comprehensive stress resistance was selected. We sterilized the seeds with 75% ethanol for 10 min and rinsed twice with deionized water. The seeds were sown in plastic containers (21 × 15 × 6 cm3) lined with double-layer filter paper. The container was placed in a growth chamber and selected seeds were germinated for 7 days, after which seedlings were transferred to a hydroponic system under controlled conditions (25 °C/18 °C day/night cycle, 65–75% relative humidity, and a light intensity of 200–250 μmol·m−2·s−1). Stress treatments involved culturing in 15% PEG-6000 dissolved in Hoagland’s solution for drought simulation and 150 mM NaCl for salt stress, with controls grown in standard Hoagland’s solution. After 7 days, stressed and control seedlings were harvested for RNA extraction and stored at −80 °C. Three biological replicates, each of which consisted of three single plants, were included in this study.
2.5. Analysis of Cis-Acting Elements in the AsCAMTA Gene Family
The 1000 bp upstream base of the start codon of the oat CAMTA gene family was used as the sequence in the PLACE (http://www.dna.affrc.go.jp/PLACE, accessed on 15 January 2024) online software. The TBtools software v2.112 was then used to analyze the sequence with the extracted elements used to perform predictive analysis on the cis-acting elements of this gene family. The cis-elements of the AsCAMTA gene family were visualized by the ‘Simple BioSequence Viewer’ function of the TBtools software v2.112.
2.6. RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR Expression Analyses
Total RNA was isolated using the RNAprep Pure Plant Plus Kit (Chengdu Baifeite Technology Co., Ltd., Chengdu, China) and cDNA was then synthesized using a TransScript II One-Step gDNA Removal and cDNA Synthesis SuperMix kit (Vazyme, Nanjing, China). The RNA concentrations and quality were evaluated using a UV Spectrophotometer (N50 Touch; Implen, Munich, Bavaria, Germany). RT-qPCR analyses were performed using the 2 × SYBR Green Premix (Shanghai Tolo Port Biotechnology Co., Ltd., Shanghai, China) and the QuantStudioTM 3 Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA). The primer sequences for the 20 AsCAMTA genes were generated using the Primer Premier 5.0 software (Table S1). The Actin1 (Forward primer: 5′-GTGGTGGTGAATGAGTAGCC-3′, Reverse primer: 5′-TTGGATTCTGGTGATGGTGTC-3′) gene was used as an internal reference gene to calculate the expression of 20 AsCAMTA genes [35]. The RT-qPCR was performed using three biological replicates and three technical replicates for each gene. The relative expression of the target genes was determined using the 2−ΔΔCt method.
3. Results
3.1. Identification of CAMTA Family Numbers in A. sativa
A total of 20 AsCAMTA genes were identified in A. sativa, denominated AsCAMTA1 to AsCAMTA20, based on their chromosomal positioning. The analysis of their physicochemical properties revealed protein lengths ranging from 403 to 1506 amino acids. The theoretical isoelectric points (pI) spanned from 5.36 (AsCAMTA18) to 8.46 (AsCAMTA5), with molecular weights from 62.24 kDa (AsCAMTA17) to 166.98 kDa (AsCAMTA3). Except for AsCAMTA10, AsCAMTA11, AsCAMTA14, and AsCAMTA15, which had an instability index below 40, suggesting stability [35], the remaining members showed greater values, indicating potential instability. Subcellular localization predictions suggested a nuclear predominance for the AsCAMTA proteins, aligning with their expected transcriptional regulatory roles. These findings are summarized in Table 1.

Table 1.
Biophysical properties and subcellular localization of the oat CAMTA properties.
3.2. Gene Structure and Motif Composition of the AsCAMTA Gene Family
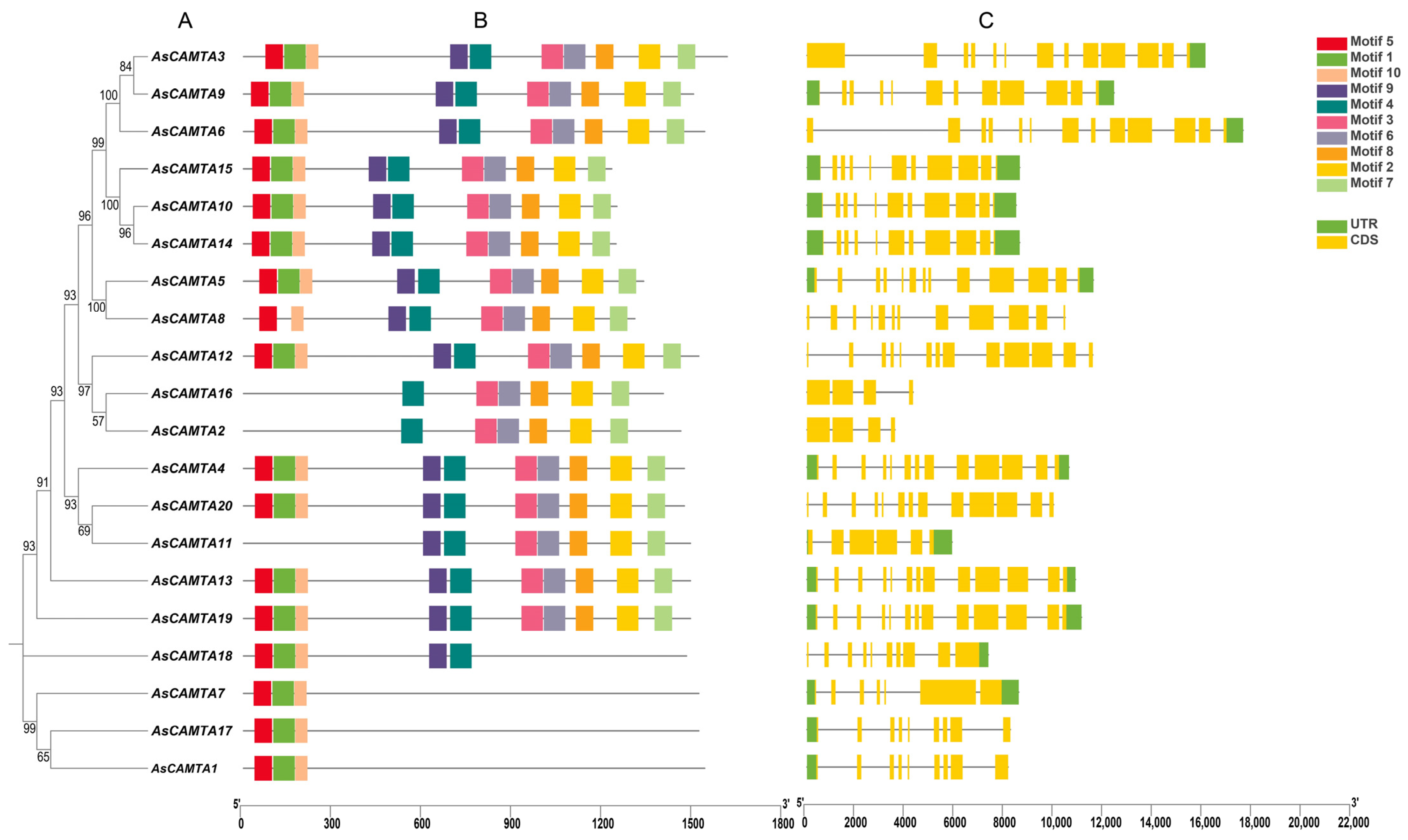
The gene structure and motif composition of the AsCAMTA family were explored through the analysis of their amino acid sequences using the MEME software (Figure 1). This led to the identification of 10 conserved motifs across the 20 AsCAMTA proteins, with individual proteins containing between 1 and 9 of these motifs. Specifically, AsCAMTA3, AsCAMTA9, AsCAMTA6, AsCAMTA15, AsCAMTA10, AsCAMTA14, AsCAMTA5, AsCAMTA12, AsCAMTA4, AsCAMTA20, AsCAMTA13, and AsCAMTA19 each contained all ten identified motifs. AsCAMTA8 had nine motifs; AsCAMTA11, seven; AsCAMTA16 and AsCAMTA2, six; AsCAMTA18, five; and AsCAMTA7, AsCAMTA17, and AsCAMTA1. Except for three members, AsCAMTA2, AsCAMTA11, and AsCAMTA16, all the other members have motifs 5 and 10. Additionally, the analysis revealed that genes grouped together on the evolutionary tree shared structural similarities and coding sequences (CDSs) with comparable numbers of introns. The positioning and lengths of these introns were consistent within groups but varied significantly between different groups, suggesting a correlation between gene structure and evolutionary lineage. In addition, genes on the same branch of the evolutionary tree are structurally similar and their CDSs have a similar number of introns. The position and length of the introns did not vary much within groups but differed significantly between groups.
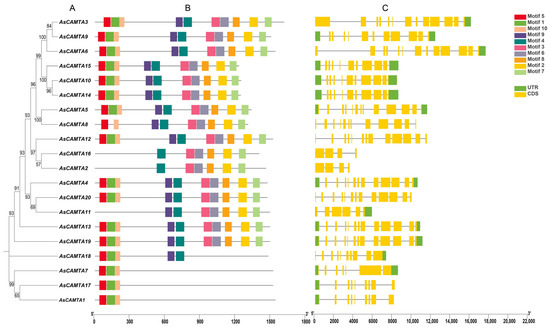
Figure 1.
Phylogenetic tree, motif analysis, and gene structure of AsCAMTA: (A) The phylogenetic tree analysis of the AsCAMTA protein. (B) The motif composition of AsCAMTA. (C) The gene structure of the AsCAMTA genes in oats.
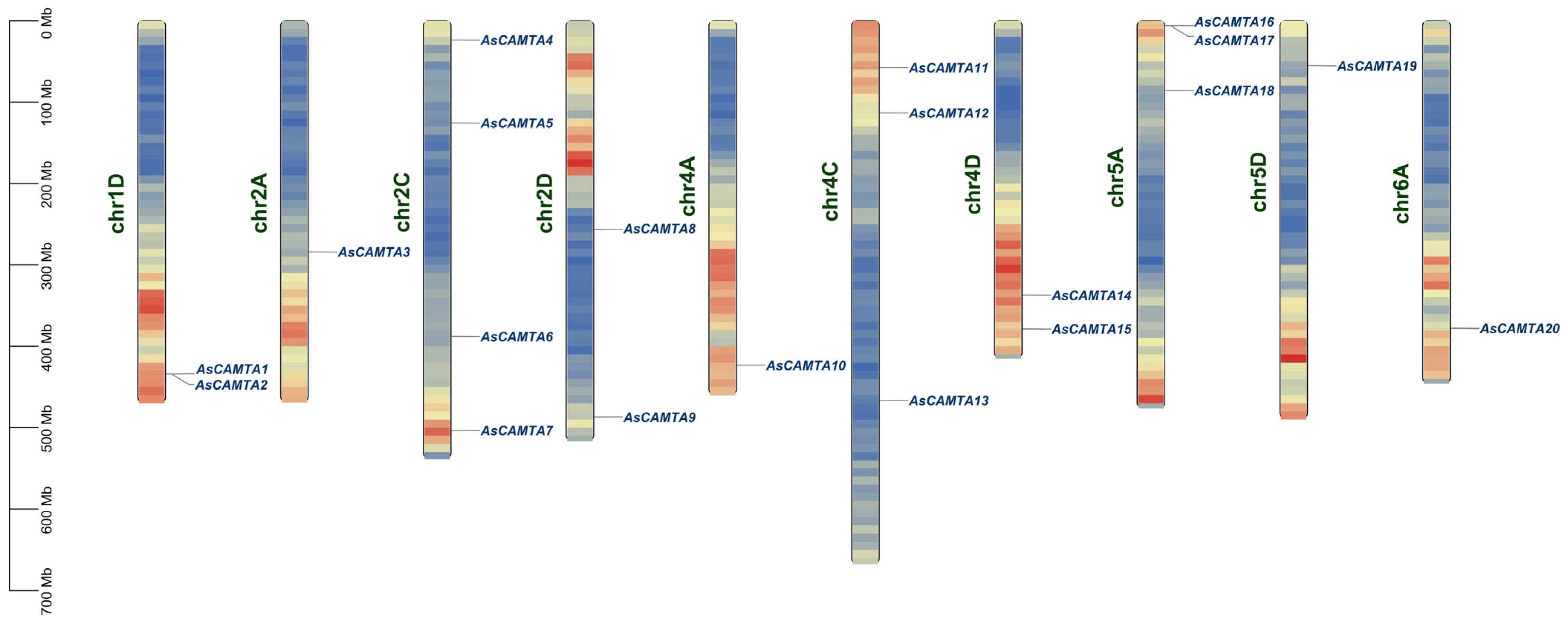
3.3. Chromosomal Location of AsCAMTAs
Through the utilization of A. sativa genome annotation and the AsCAMTA gene IDs, we mapped twenty AsCAMTA genes across various chromosomes, as illustrated in Figure 2. The analysis revealed a predominant distribution of these genes on chromosome 2A, with each of the remaining chromosomes hosting at least one of the identified genes. Notably, the chromosomes 3A/C/D, 7A/C/D, 1A/C, 6C/D, and 5C were devoid of any CAMTA genes, indicating a non-uniform distribution across the A. sativa genome. Furthermore, gene clusters were specifically observed on chromosomes 1D and 5A, attributed to tandem duplication events. For instance, AsCAMTA1 and AsCAMTA2 on chromosome 1D, and AsCAMTA16 and AsCAMTA17 on chromosome 5A, formed tandem duplication clusters. This suggests an evolutionary relationship among these AsCAMTA members, highlighting a common origin from chromosomal duplications and similar structural and functional properties.
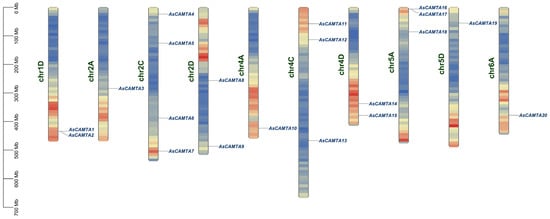
Figure 2.
Chromosomal localization of the members of the oat CAMTA gene family. Blue to yellow colors within the chromosomes indicate increased gene density. Chromosome numbers are shown at the right of the vertical bar; gene locations are shown at the left of the vertical bar.
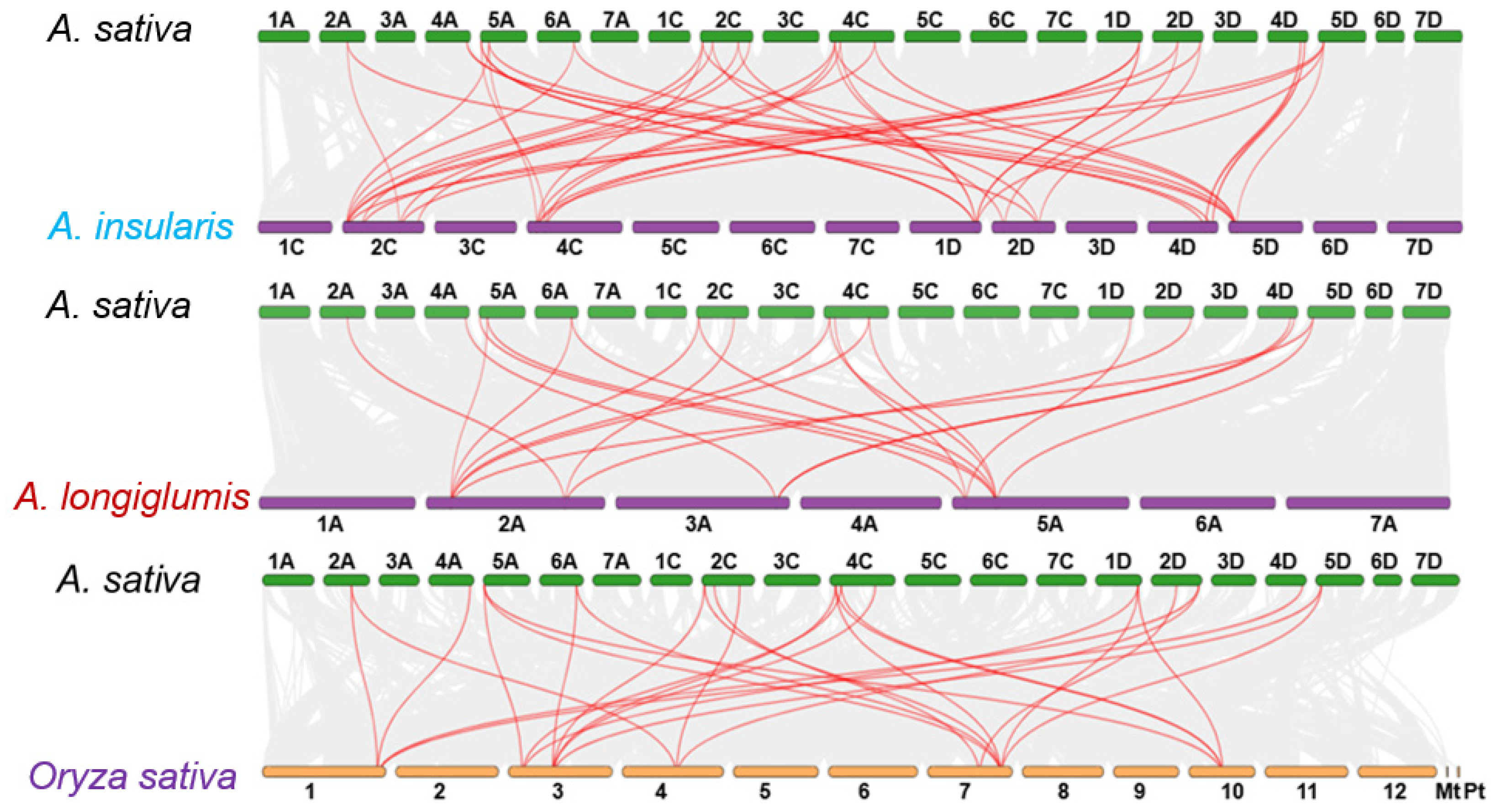
3.4. Gene Duplication and Collinearity Analysis of CAMTA Genes
To further investigate the evolutionary lineage of the A. sativa CAMTA gene family, three comparative homology maps between A. sativa CAMTA members and three representative species were constructed, including two putative A. sativa ancestors (A. insularis and A. longiglumis) and rice, as depicted in Figure 3. These maps revealed extensive collinearity among the genes, indicating a close evolutionary relationship. Specifically, 102,298 collinearity pairs were detected between A. sativa and A. insularis, representing 69.66% of the total oat gene count. Similarly, A. sativa and A. longiglumis shared 180,373 collinearity pairs, accounting for the same percentage of A. sativa genes. In comparison, a total of 60,264 collinearity pairs were identified between A. sativa and rice, covering 32.03% of the A. sativa genome.
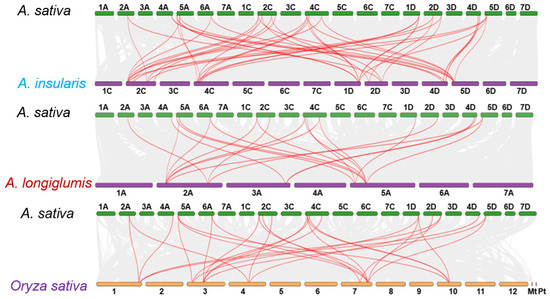
Figure 3.
Synteny analysis of CAMTAs in A. sativa and three representative plants.
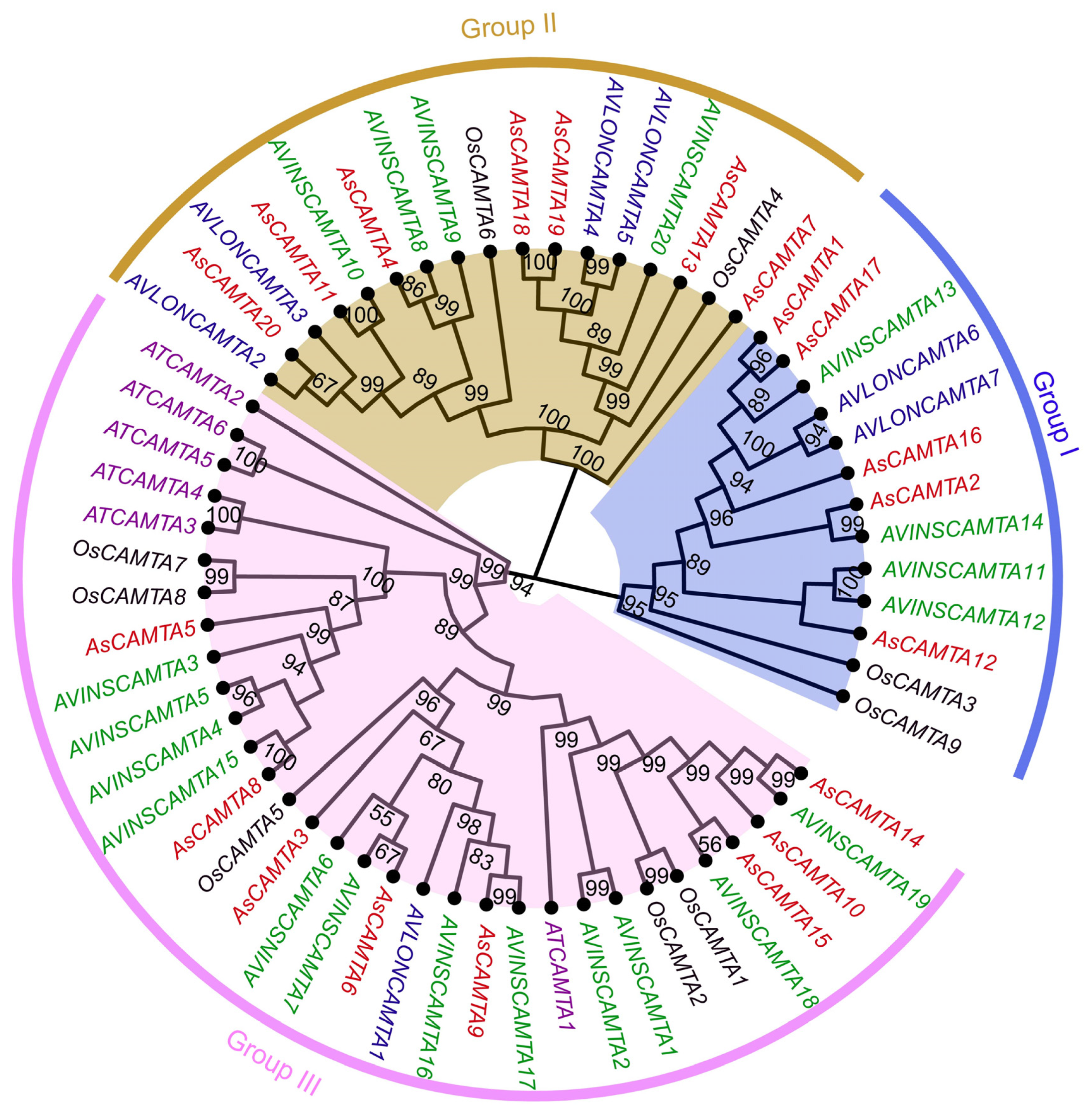
3.5. Phylogenetic Analysis of CAMTA Members
Utilizing the ML method via the MEGA11 software, a phylogenetic tree was constructed to elucidate the evolutionary relationships among CAMTA members, including 7 from Arabidopsis, 20 from A. sativa, 9 from rice, 7 from A. insularis, and 20 from A. longiglumis (Figure 4). The tree stratified the CAMTA protein sequences into three main groups, with Group I comprising the smallest number (13 members), followed by Group II (13 members). Group III is the most numerous (32 members). This classification emphasizes the diversity of the evolutionary pathways of CAMTA proteins among different species, and all the CAMTA members of Arabidopsis were classified in Group III.
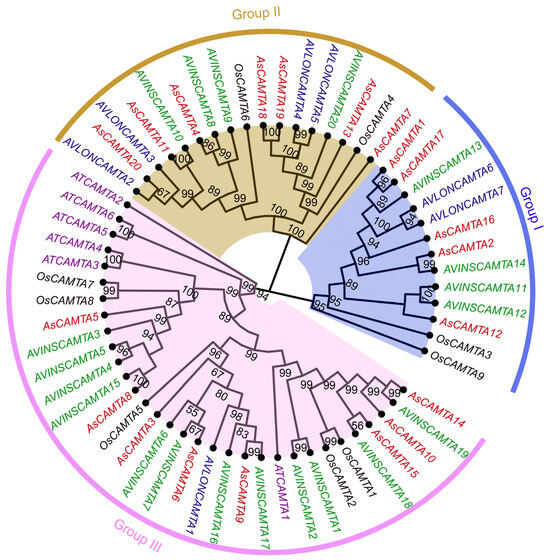
Figure 4.
Phylogenetic tree analysis of the CAMTA proteins from A. sativa, A. thaliana, O. sativa, A. insularis, and A. longiglumis. The CAMTAs were divided into three clades (Group I–III) based on the clustering of the protein sequence.
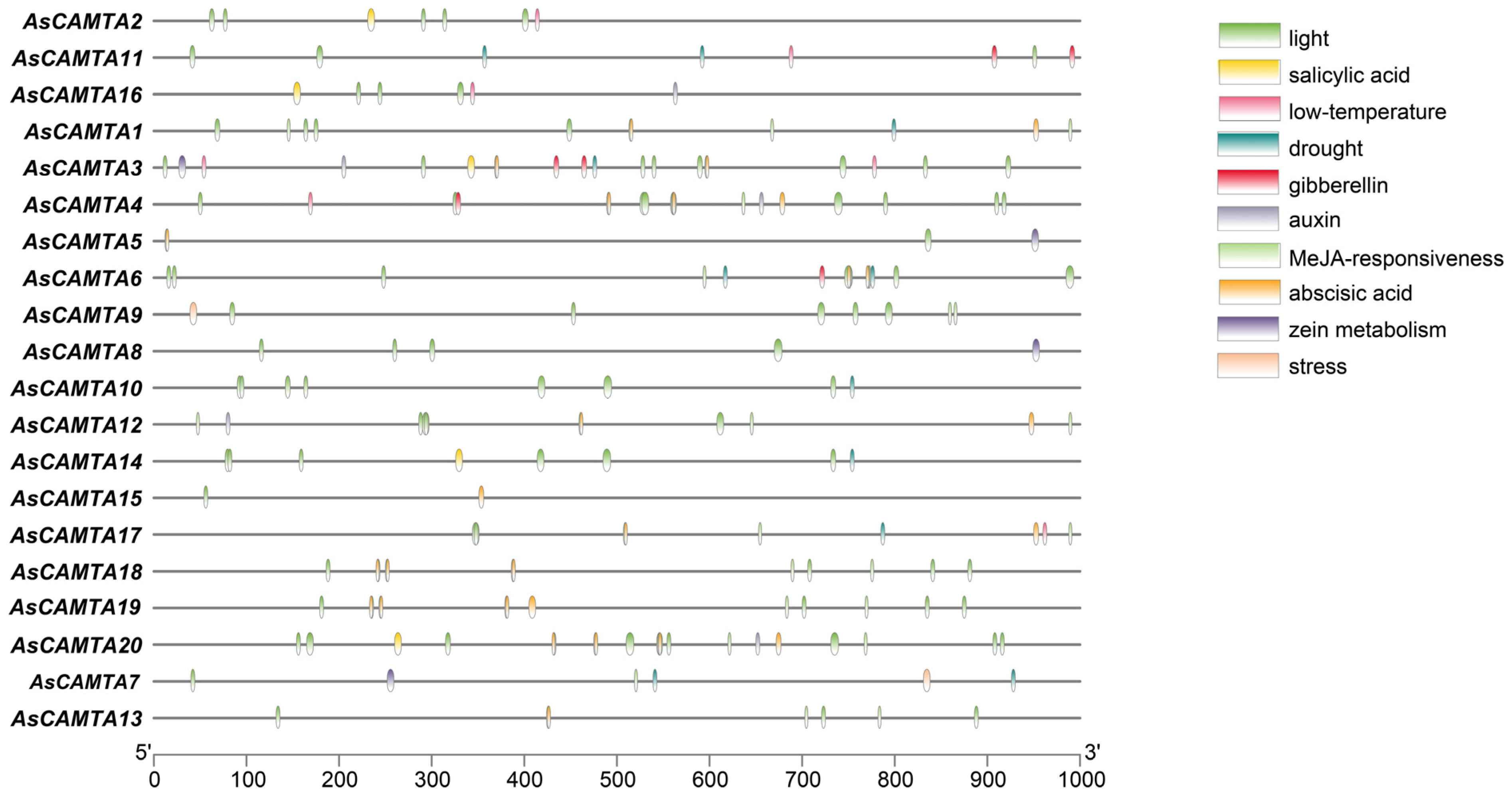
3.6. Cis-Acting Element Analysis of AsCAMTA Genes
The PlantCARE predictive analysis of the promoter sequence information of 1000 bp upstream of the AsCAMTAs gene resulted in a large number of elements related to plant growth and development (e.g., light, hormones, abiotic threats, etc.), but there were differences in the type and number of elements contained in each member (Figure 5). The results showed that the promoter region contains two basic cis-acting elements, such as drought-responsive elements; salt-responsive elements; TATA-box and CAAT-box; light-responsive elements, such as G-box; hormone-responsive elements, such as abscisic acid-responsive element (ABRE); ethylene-responsive element (ERE); stress-responsive elements (STRE); anaerobic-responsive element (ARE); etc. This suggests that these genes may be involved in the growth of oats, which play an important role in oat growth and development and abiotic stresses.
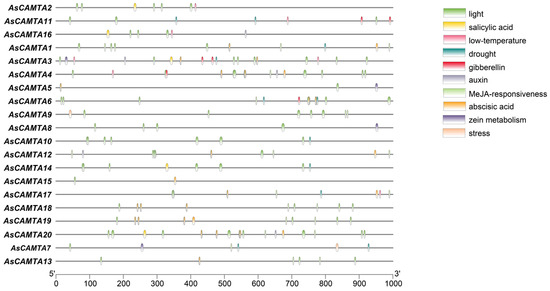
Figure 5.
Cis-acting elements in promoters of AsCAMTA family members.
3.7. Expression Analysis of AsCAMTAs in A. sativa under Two Abiotic Stresses
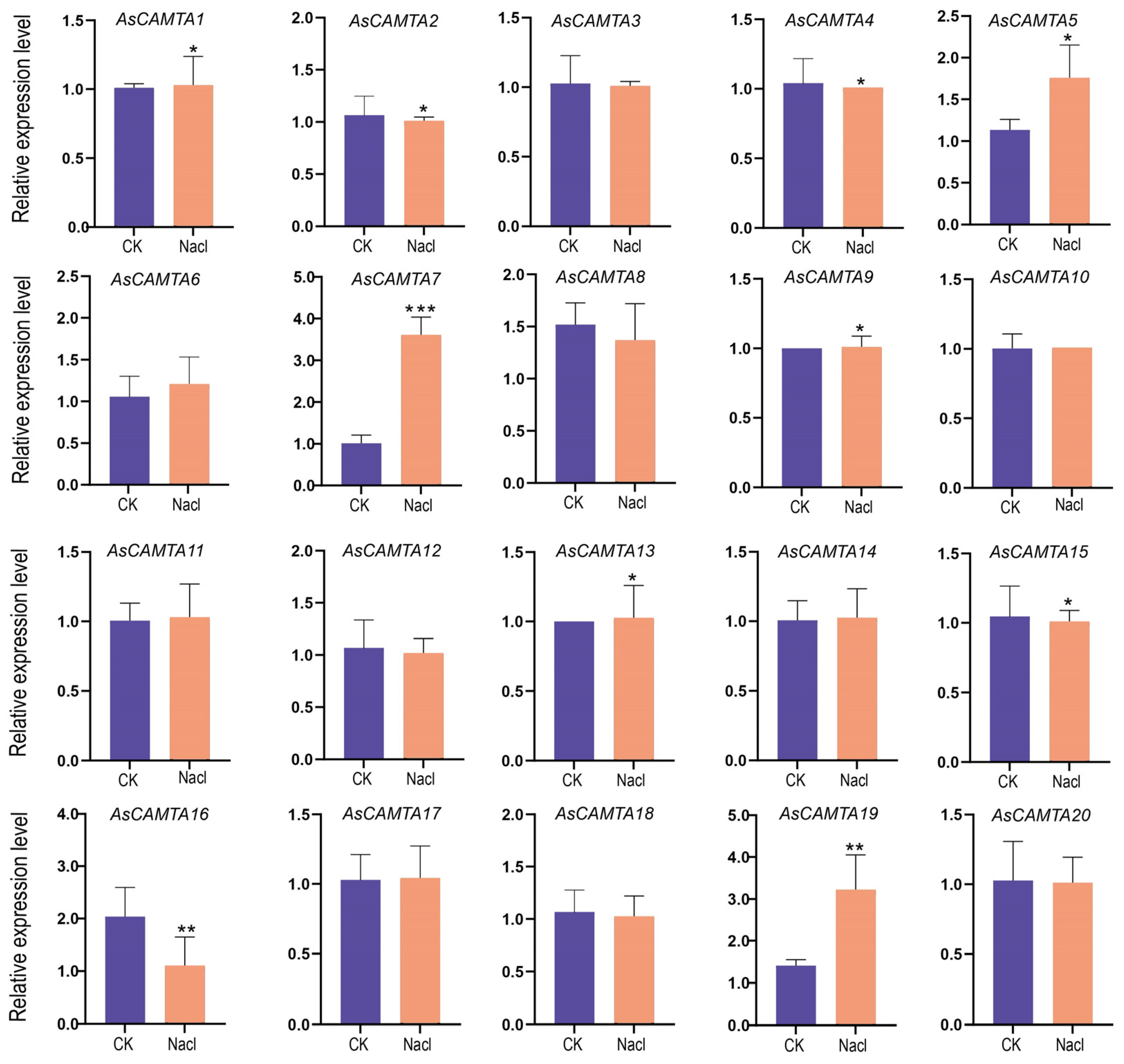
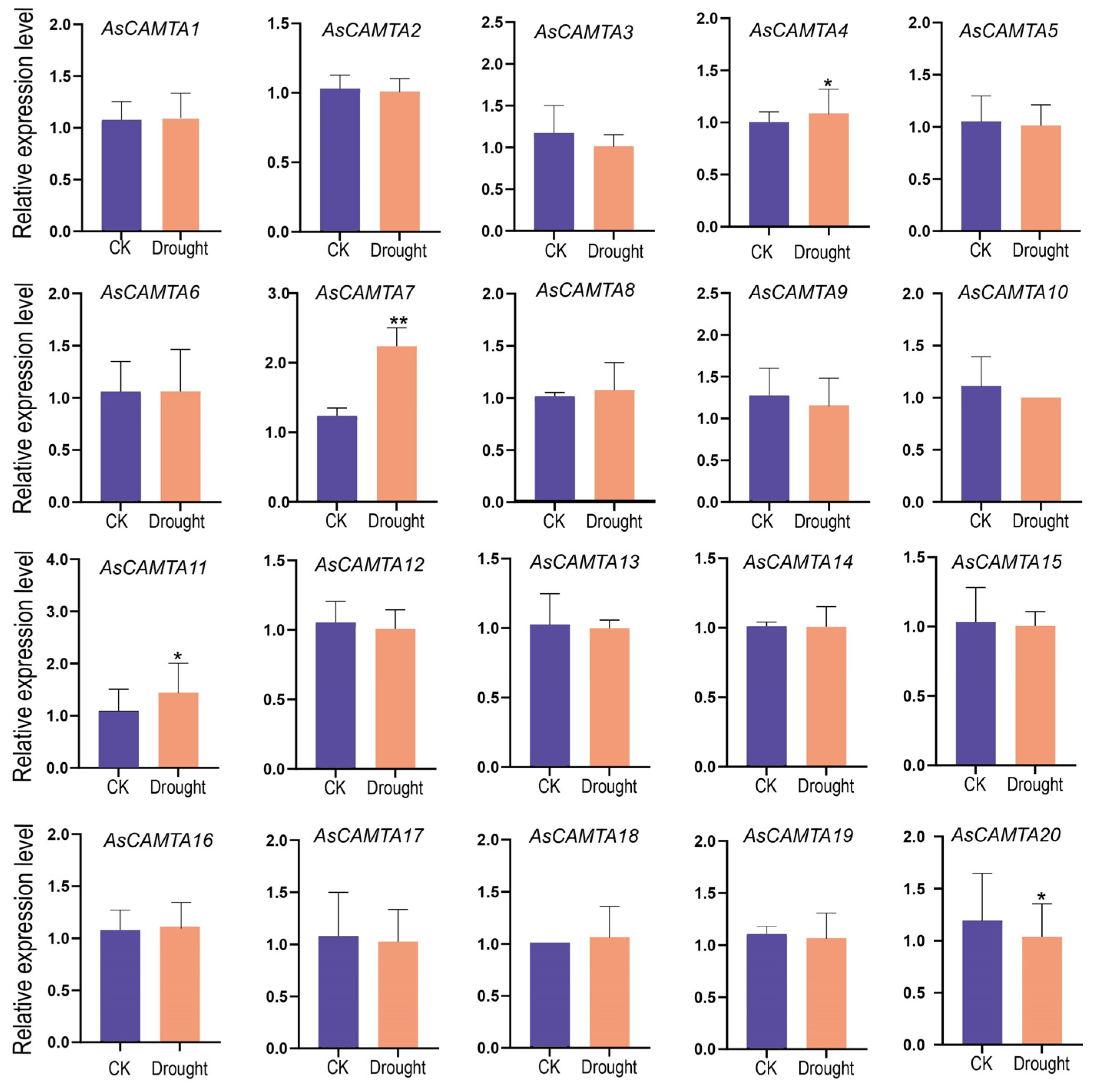
The expression profiles of 20 AsCAMTAs were analyzed using qRT-PCR under two abiotic stresses: salt and drought, as shown in Figure 6 and Figure 7. The study observed distinct expression patterns among the AsCAMTAs under these stress conditions, with salt stress eliciting a more pronounced response. Specifically, AsCAMTA5, AsCAMTA7, and AsCAMTA19 exhibited significant expression levels in response to salt stress, while AsCAMTA7 and AsCAMTA11 were notably up-regulated under drought stress. The pronounced expression of the AsCAMTA7 gene under both stress conditions suggests its potential as a pivotal regulator in A. sativa’s adaptation to drought and salt stresses.
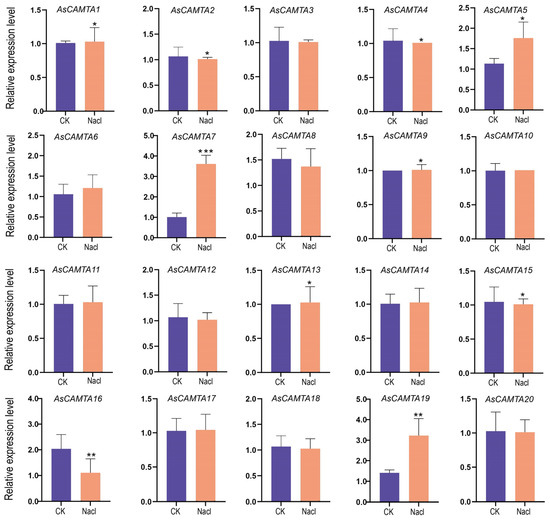
Figure 6.
The expression patterns of 20 AsCAMTA genes were analyzed under salt stress by real-time quantitative RT-PCR. *, ** and *** represent significance levels of p < 0.05, p < 0.01 and p < 0.001, respectively.
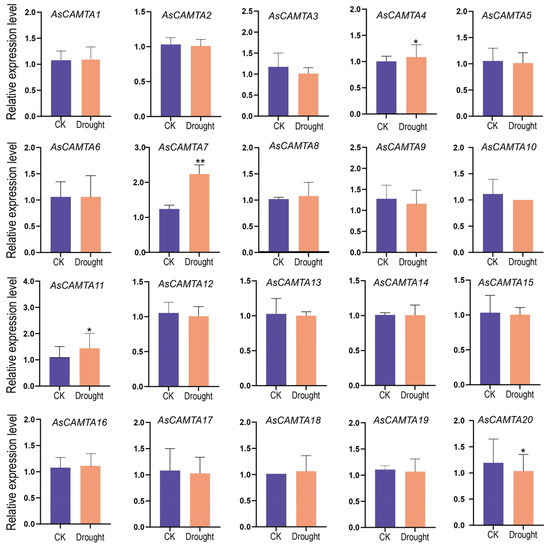
Figure 7.
The expression patterns of 20 AsCAMTA genes were analyzed under drought stress by real-time quantitative RT-PCR. * and ** represent significance levels of p < 0.05 and p < 0.01, respectively.
In addition, the expression levels of AsCAMTA7 and AsCAMTA11 were up-regulated under drought stress. The AsCAMTA7 gene was significantly expressed under two stress conditions, so it was speculated that it may be a key gene regulating drought stress and salt stress in A. sativa.
4. Discussion
The CAMTA family, a key player in plant adversity signaling, demonstrates broad phylogenetic diversity across eukaryotes. To date, the CAMTA family of transcription factors has been identified in a variety of plants, for example, 6 in Arabidopsis [36], 6 in tea plants [37], 5 in peaches [38], and 15 in wheat [39], emphasizing the evolutionary conservation and expansion of this gene family across different species. Our study extends this comparative framework to oats, wherein we identified 20 CAMTA genes distributed across 10 chromosomes. This number is comparable to that in Triticum aestivum and Brassica napus [40], suggesting a correlation with genome size and polyploidy, particularly given oats’ hexaploid nature which likely contributes to an increased number of homologous genes.
The biophysical diversity observed within the AsCAMTA proteins, with isoelectric points ranging from 5.36 to 8.46, indicates significant functional versatility and aligns with previous findings in wheat [23]. Notably, the uneven chromosomal distribution of AsCAMTA genes, predominantly in regions of high gene density, may reflect segmental duplications within the oat genome, a hypothesis supported by the observed chromosomal rearrangements [41]. Physicochemical property analyses showed that these proteins differed in length, isoelectric point, molecular weight, and hydrophilicity, suggesting a broad functional specificity of the family. The predictions of subcellular localization indicate that they function predominantly in the nucleus, consistent with their transcription factor functions. The gene structure and motif analyses highlight the conserved motifs that are essential for CAMTA protein function, and changes in motif composition and intron distribution reflect evolutionary divergence.
Polyploidization and gene duplication are pivotal in shaping the genomic architecture of oats, facilitating the expansion of gene families such as AsCAMTA. These processes not only increase genetic diversity but also enhance the plant’s ability to adapt to diverse environmental stresses. The identification of segmental duplications within the AsCAMTA gene family, such as those between AsCAMTA1 and AsCAMTA2, and AsCAMTA16 and AsCAMTA17, supports the role of duplicative events in the adaptive evolutionary capacity of oats. A. insularis and A. longiglumis from different subgenomes further emphasize the complexity and evolutionary dynamics of the oat genome [42].
The evolution of large eukaryotic genomes, the distribution of conserved motifs, intron gain or loss, and intron density have a significant impact on the evolution of large eukaryotic genomes, the distribution of conserved motifs, and exon and intron structures, which was relatively uniform among the different AsCAMTA gene subgroups. The pattern of intron gain and loss, and the density of these introns, play crucial roles in the evolution of large eukaryotic genomes. By categorizing the AsCAMTA genes into seven groups based on their structural similarities, this study not only enhances our understanding of the genetic architecture of oats but also contributes to broader genomic studies on plant adaptation and evolution.
The promoters of oat CAMTA genes possess numerous cis-acting elements that are crucial for plant development and adversity responses, suggesting that AsCAMTAs could be integral to abiotic stress resistance in oats. In Arabidopsis, CAMTA genes contribute significantly to frost resistance by inducing the expression of CBF2 and other cold-responsive genes. For instance, AtCAMTA1 enhances frost resistance by regulating stress-responsive promoters, including those of RD26 and ERD7. These findings emphasize the roles of CAMTA genes in cold stress management, which is further complicated in C4 plants like Cenchrus fungigraminus, where CAMTA expression varies with drought conditions, decreasing after rewatering, indicative of its role in drought response regulation [43]. Similar patterns are observed in other species; for example, in Capsicum annuum, CAMTA expression changes under low temperature and salinity stress, suggesting a broad role in stress signal transduction [44]. In Hevea brasiliensis, HbCAMTA3 is markedly up-regulated in response to low-temperature stress, highlighting its potential as a candidate gene for enhancing cold stress tolerance [45].
Our analysis of AsCAMTA gene expression in oats following the salt and drought treatments indicates that these genes may play a comprehensive role in mediating plant responses to multiple abiotic stresses. This is evidenced by the induction of most AsCAMTA genes under both salt and drought conditions, although responses varied among individual genes. Particularly, AsCAMTA11 and AsCAMTA7, which cluster in the same phylogenetic group, showed distinct responses under drought stress, with AsCAMTA11 displaying a higher number of drought response elements. This suggests a specific regulatory role for AsCAMTA11 in drought stress management in oats. The quantitative RT-PCR results further revealed that the expression levels of AsCAMTA genes were generally higher under salt stress than drought, indicating a possible greater sensitivity of CAMTA proteins to salinity. This differential expression suggests that CAMTA proteins could be more effectively targeted for improving salt tolerance in oats.
Despite these insights, the specific functions of the AsCAMTA gene family in oats remain largely unexplored. Therefore, future studies should focus on functionally characterizing these genes, particularly through transgenic or gene editing approaches, to confirm their roles in stress responses and potential in crop improvement strategies.
5. Conclusions
This study provided a comprehensive analysis of the CAMTA gene family in A. sativa, and a total of 20 distinct CAMTA genes (AsCAMTA1 to AsCAMTA20) were identified. Bioinformatics data were used to analyze the evolutionary relationships, gene structure, gene duplication, promoters, and regulatory networks of the CAMTA gene family. Chromosomal mapping and gene duplication analysis emphasized an uneven distribution across chromosomes. The analysis of cis-acting elements revealed their influential role in the expression pattern of CAMTA genes under abiotic stresses. The expression analysis under abiotic stresses revealed differential expression patterns, identifying the potential key regulators of the stress response. Collectively, these findings enhance our understanding of the CAMTA gene family’s evolutionary and functional dynamics in A. sativa, laying the groundwork for further functional studies and crop improvement strategies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14092053/s1, Table S1: The primer information of AsCAMTA genes used in the study. File S1: The CAMTA proteins sequences in the study.
Author Contributions
Conceptualization, S.C. and M.Y.; methodology, Y.Y., J.L. and S.C.; software, Y.Y. and J.L.; investigation, Y.Y. and J.L.; data curation, J.L. and M.Y.; writing—original draft preparation, Y.Y. and J.L.; writing—review and editing, M.Y. and S.C.; supervision, S.C.; funding acquisition, M.Y. and S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Key Research & Development Program of Sichuan province (2021YFYZ0013, 2023NZZJ0002), and the Fundamental Research Funds for the Central Universities, Southwest Minzu University (ZYN2024090).
Data Availability Statement
The data presented in this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fu, Y.B.; Peterson, G.W.; Williams, D.; Richards, K.W.; Fetch, J.M. Patterns of AFLP variation in a core subset of cultivated hexaploid oat germplasm. Theor. Appl. Genet. 2005, 111, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape stringApp: Network analysis and visualization of proteomics data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef]
- Bräutigam, M.; Lindlöf, A.; Zakhrabekova, S.; Gharti-Chhetri, G.; Olsson, B.; Olsson, O. Generation and analysis of 9792 EST sequences from cold acclimated oat, Avena sativa. BMC Plant Biol. 2005, 5, 18. [Google Scholar] [CrossRef]
- Zhao, D.; Wright, D.L.; Marois, J.J.; Mackowiak, C.L.; Brennan, M. Improved growth and nutrient status of an oat cover crop in sod-based versus conventional peanut-cotton Rotations. Agron. Sustain. Dev. 2010, 30, 497–504. [Google Scholar] [CrossRef]
- Xu, W.; Dai, W.; Yan, H.; Li, S.; Shen, H.; Chen, Y.; Xu, H.; Sun, Y.; He, Z.; Ma, M. Arabidopsis NIP3;1 Plays an important role in arsenic uptake and Root-to-Shoot translocation under arsenite stress conditions. Mol. Plant 2015, 8, 722–733. [Google Scholar] [CrossRef]
- Wu, B.; Munkhtuya, Y.; Li, J.; Hu, Y.; Zhang, Q.; Zhang, Z. Comparative transcriptional profiling and physiological responses of two contrasting oat genotypes under salt stress. Sci. Rep. 2018, 8, 16248. [Google Scholar] [CrossRef] [PubMed]
- Boczkowska, M.; Zebrowski, J.; Nowosielski, J.; Kordulasińska, I.; Nowosielska, D.; Podyma, W. Environmentally-related genotypic, phenotypic and metabolic diversity of oat (Avena sativa L.) landraces based on 67 Polish accessions. Genet. Resour. Crop Ev. 2017, 64, 1829–1840. [Google Scholar] [CrossRef]
- Shi, S.; Nan, L.; Smith, K. The current status, problems, and prospects of Alfalfa (Medicago sativa L.) breeding in China. Agronomy 2017, 7, 1. [Google Scholar] [CrossRef]
- Cui, X.Y.; Du, Y.T.; Fu, J.; Yu, T.F.; Wang, C.T.; Chen, M.; Chen, J.; Ma, Y.Z.; Xu, Z.S. Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses. BMC Plant Biol. 2018, 18, 93. [Google Scholar] [CrossRef]
- Cheng, S.B.; Yang, X.Z.; Zou, L.; Wu, D.D.; Lu, J.L.; Cheng, Y.R.; Wang, Y.; Zeng, J.; Kang, H.Y.; Sha, L.N.; et al. Comparative physiological and root transcriptome analysis of two annual ryegrass cultivars under drought stress. J. Plant Physiol. 2022, 277, 153807. [Google Scholar] [CrossRef]
- Ling, L.; Li, M.; Chen, N.; Xie, X.; Han, Z.; Ren, G.; Yin, Y.; Jiang, H. Genome-wide identification of NAC gene family and expression analysis under abiotic stresses in Avena sativa. Genes 2023, 14, 1186. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Qiu, Q.; Zou, B.; Wu, Q.; Ye, X.; Wan, Y.; Huang, J.; Wu, X.; Sun, Y.; Yan, H.; et al. Comparative transcriptome and genome analysis unravels the response of tatary buckwheat root to nitrogen deficiency. Plant Physiol. Bioch. 2023, 196, 647–660. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Shariq Iqbal, M.; Singh, S.P.; Buaboocha, T. Ca2+/calmodulin complex triggers CAMTA transcriptional machinery under stress in plants: Signaling cascade and molecular regulation. Front. Plant Sci. 2020, 11, 598327. [Google Scholar] [CrossRef]
- Xiao, P.; Feng, J.-W.; Zhu, X.-T.; Gao, J. Evolution analyses of CAMTA transcription factor in plants and its enhancing effect on cold-tolerance. Front. Plant Sci. 2021, 12, 758187. [Google Scholar] [CrossRef]
- Reddy, A.S.N. Calcium: Silver bullet in signaling. Plant Sci. 2001, 160, 381–404. [Google Scholar] [CrossRef]
- Snedden, W.A.; Fromm, H. Calmodulin as a versatile calcium signal transducer in plants. New Phytol. 2001, 151, 35–66. [Google Scholar] [CrossRef]
- Bouché, N.; Yellin, A.; Snedden, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Biol. 2005, 56, 435–466. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.S.N.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Du, L. Calcium signaling: Decoding mechanism of calcium signatures. New Phytol. 2018, 217, 1394–1396. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. An early ethylene up-regulated gene encoding a calmodulin-binding protein pnvolved in plant senescence and death. J. Biol. Chem. 2000, 275, 38467–38473. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.; Aysha, J.; Ketehouli, T.; Yang, J.; Du, L.; Wang, F.; Li, H. Calmodulin binding transcription activators: An interplay between calcium signalling and plant stress tolerance. J. Plant Physiol. 2021, 256, 153327. [Google Scholar] [CrossRef]
- Wang, D.; Wu, X.; Gao, S.; Zhang, S.; Wang, W.; Fang, Z.; Liu, S.; Wang, X.; Zhao, C.; Tang, Y. Systematic analysis and identification of drought-responsive genes of the CAMTA gene family in wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2022, 23, 4542. [Google Scholar] [CrossRef]
- Cunha, S.R.; Mohler, P.J. Ankyrin protein networks in membrane formation and stabilization. J. Cell Mol. Med. 2009, 13, 4364–4376. [Google Scholar] [CrossRef]
- Bouché, N.; Scharlat, A.; Snedden, W.; Bouchez, D.; Fromm, H. A novel family of calmodulin-binding transcription activators in multicellular organisms. J. Biol. Chem. 2002, 277, 21851–21861. [Google Scholar] [CrossRef]
- Kim, Y.; Park, S.; Gilmour, S.J.; Thomashow, M.F. Roles of CAMTA transcription factors and salicylic acid in configuring the low-temperature transcriptome and freezing tolerance of Arabidopsis. Plant J. 2013, 75, 364–376. [Google Scholar] [CrossRef]
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar] [CrossRef]
- Pandey, N.; Ranjan, A.; Pant, P.; Tripathi, R.K.; Ateek, F.; Pandey, H.P.; Patre, U.V.; Sawant, S.V. CAMTA 1 regulates drought responses in Arabidopsis thaliana. BMC Genom. 2013, 14, 216. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, F.; Zhou, H.; Liu, N.; Niu, X.; Yan, C.; Zhang, L.; Han, S.; Hou, C.; Wang, D. TaCAMTA4, a calmodulin-interacting protein, involved in defense response of wheat to Puccinia triticina. Sci. Rep. 2019, 9, 641. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc 2.0: An improved package of web-servers for predicting subcellular localization of proteins in various organisms. Nat. Sci. 2010, 2, 1090–1103. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Gamage, D.G.; Gunaratne, A.; Periyannan, G.R.; Russell, T.G. Applicability of instability index for in vitro protein stability prediction. Protein Pept. Lett. 2019, 26, 339–347. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhao, M.; Xing, F.; Mao, G.; Wang, Y.; Dai, Y.; Niu, M.; Yuan, H. Identification and expression analysis of CAMTA genes in tea plant reveal their complex regulatory role in stress responses. Front. Plant Sci. 2022, 13, 910768. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, Z.; Cao, X.; Duan, W.; Wei, C.; Zhang, C.; Jiang, D.; Li, M.; Chen, K.; Qiao, Y.; et al. Genome-wide analysis of calmodulin binding transcription activator (CAMTA) gene family in peach (Prunus persica L. Batsch) and ectopic expression of PpCAMTA1 in Arabidopsis camta2,3 mutant restore plant development. Int. J. Mol. Sci. 2022, 23, 10500. [Google Scholar] [CrossRef]
- Yang, F.; Dong, F.; Hu, F.; Liu, Y.; Chai, J.; Zhao, H.; Lv, M.; Zhou, S. Genome-wide identification and expression analysis of the calmodulin-binding transcription activator (CAMTA) gene family in wheat (Triticum aestivum L.). BMC Genet. 2020, 21, 105. [Google Scholar] [CrossRef]
- Rahman, H.; Xu, Y.P.; Zhang, X.R.; Cai, X.Z. Brassica napus genome possesses extraordinary high number of CAMTA genes and CAMTA3 contributes to PAMP triggered immunity and resistance to Sclerotinia sclerotiorum. Front. Plant Sci. 2016, 7, 581. [Google Scholar] [CrossRef]
- Kamal, N.; Tsardakas Renhuldt, N.; Bentzer, J.; Gundlach, H.; Haberer, G.; Juhász, A.; Lux, T.; Bose, U.; Tye-Din, J.A.; Lang, D.; et al. The Mosaic oat genome gives insights into a uniquely healthy cereal crop. Nature 2022, 606, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, H.; Guo, L.; Deng, C.; Wang, C.; Wang, Y.; Kang, L.; Zhou, P.; Yu, K.; Dong, X.; et al. Reference genome assemblies reveal the origin and evolution of allohexaploid oat. Nat. Genet. 2022, 54, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Shi, F.; Fukuda, K.; Yang, Y. Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Sci. Plant Nutr. 2010, 56, 725–733. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.-K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Ther. Innov. Regul. Sci. 2020, 1, 100017. [Google Scholar] [CrossRef]
- Zhu, T.; Wu, S.; Zhang, D.; Li, Z.; Xie, K.; An, X.; Ma, B.; Hou, Q.; Dong, Z.; Tian, Y.; et al. Genome-wide analysis of maize GPAT gene family and cytological characterization and breeding application of ZmMs33/ZmGPAT6 gene. Theor. Appl. Genet. 2019, 132, 2137–2154. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).