
Determination of Critical Crop Water Stress Index of Tea under Drought Stress Based on the Intercellular CO2 Concentration

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Soil Physical Properties of Experimental Site
2.3. Fertilizer Management during the Experiment
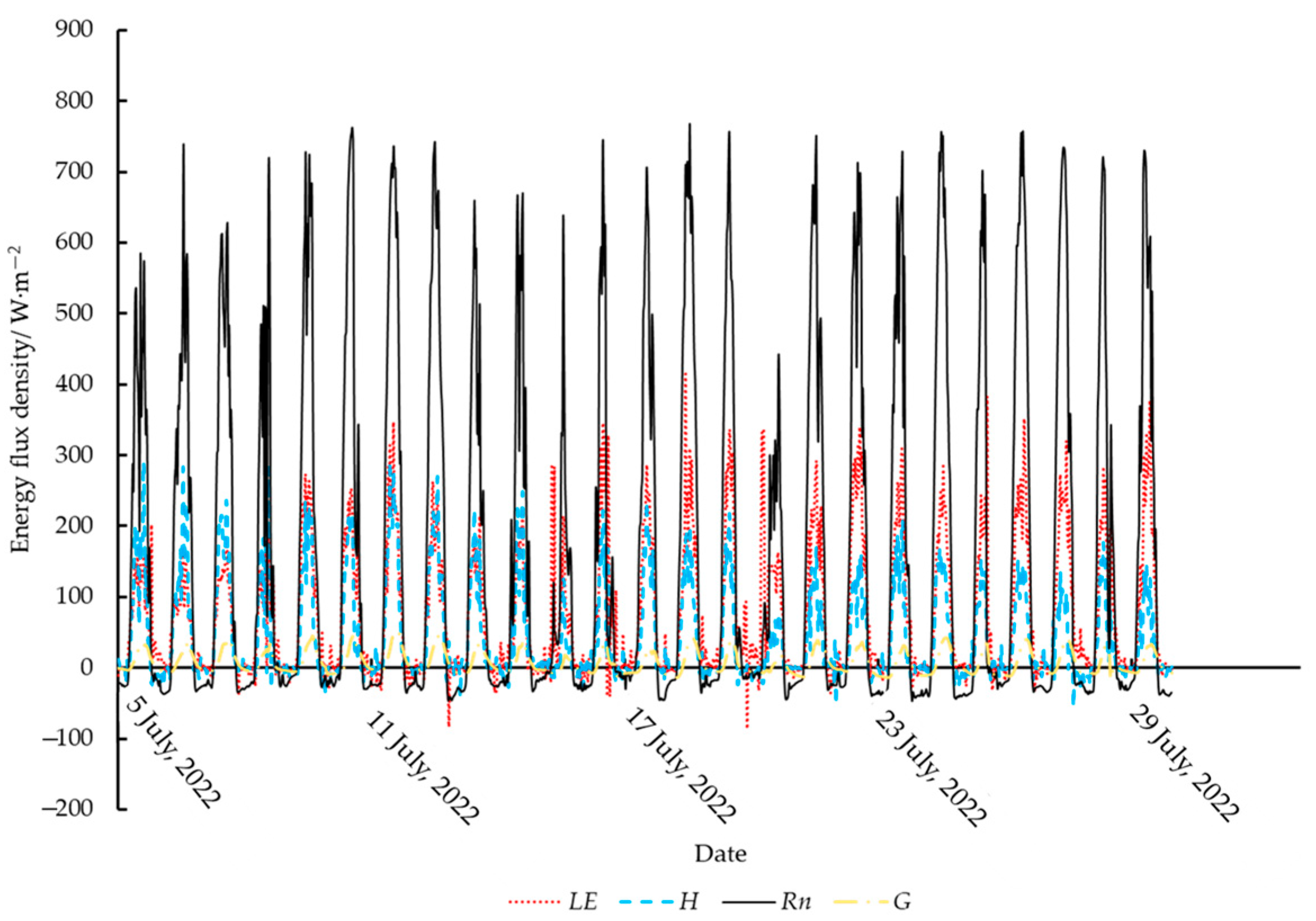
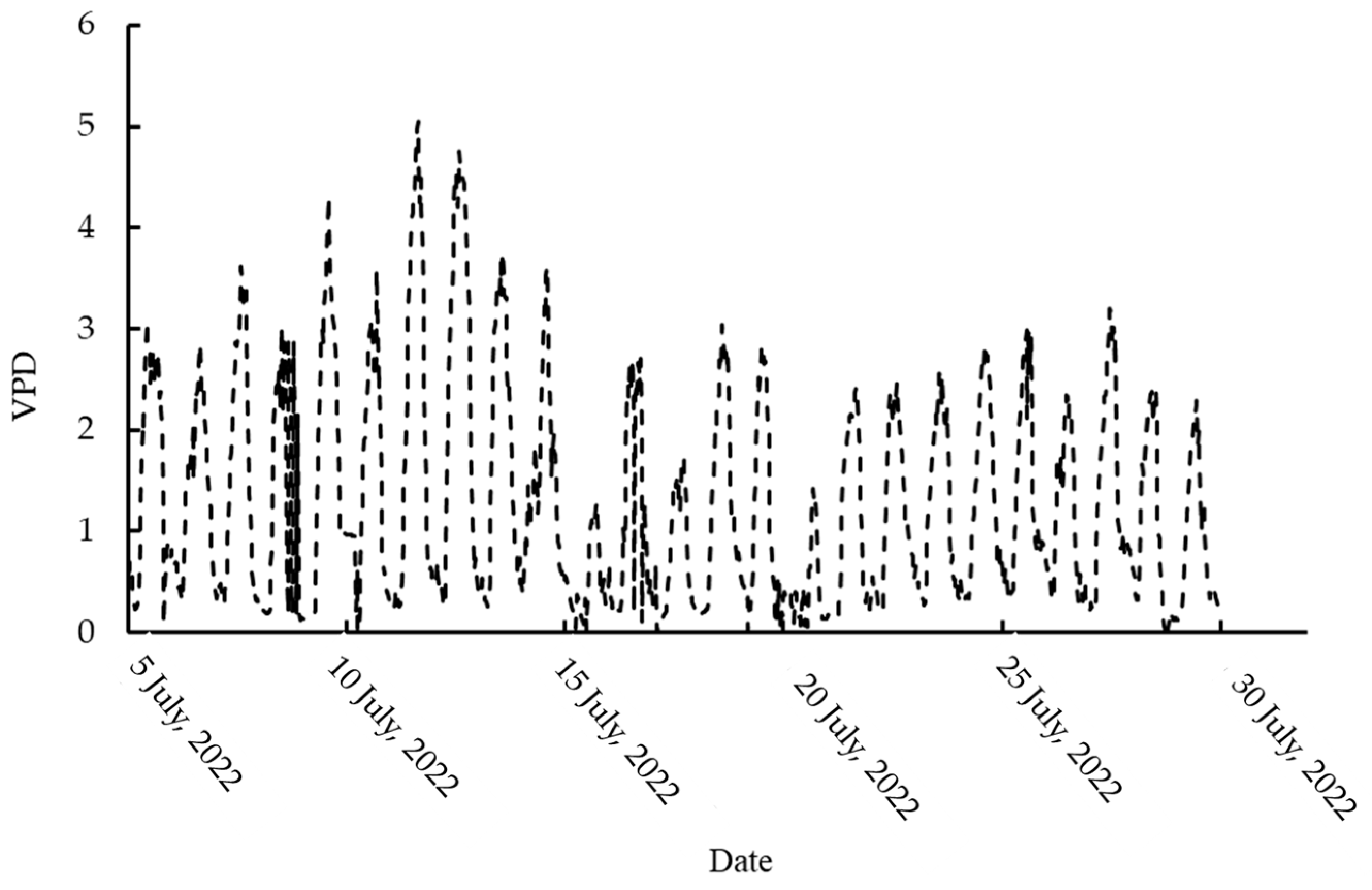
2.4. Micrometeorological Data Collection
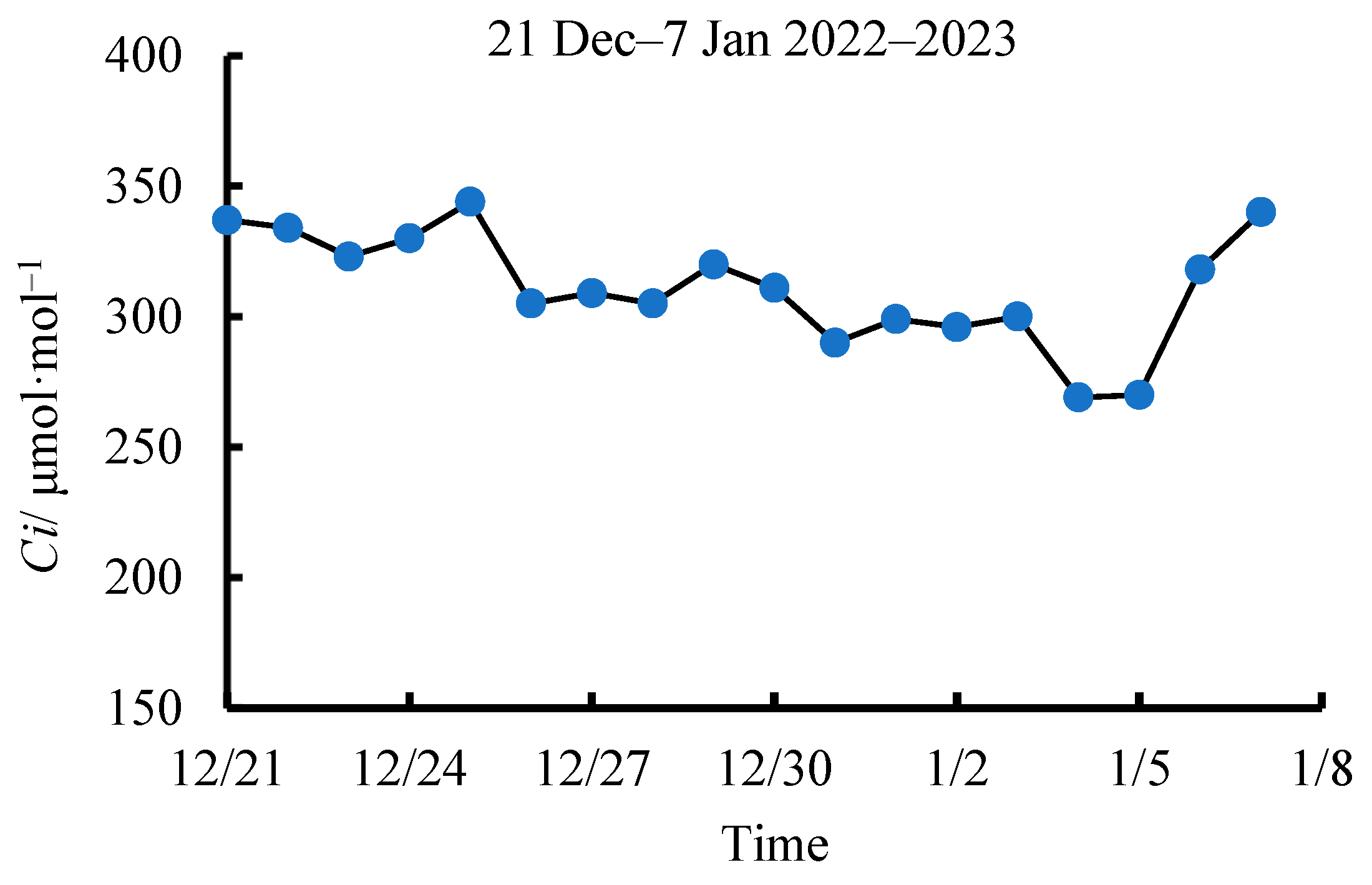
2.5. Intercellular CO2 Concentration (Ci)
2.6. Crop Water Stress Index (CWSI)
3. Results
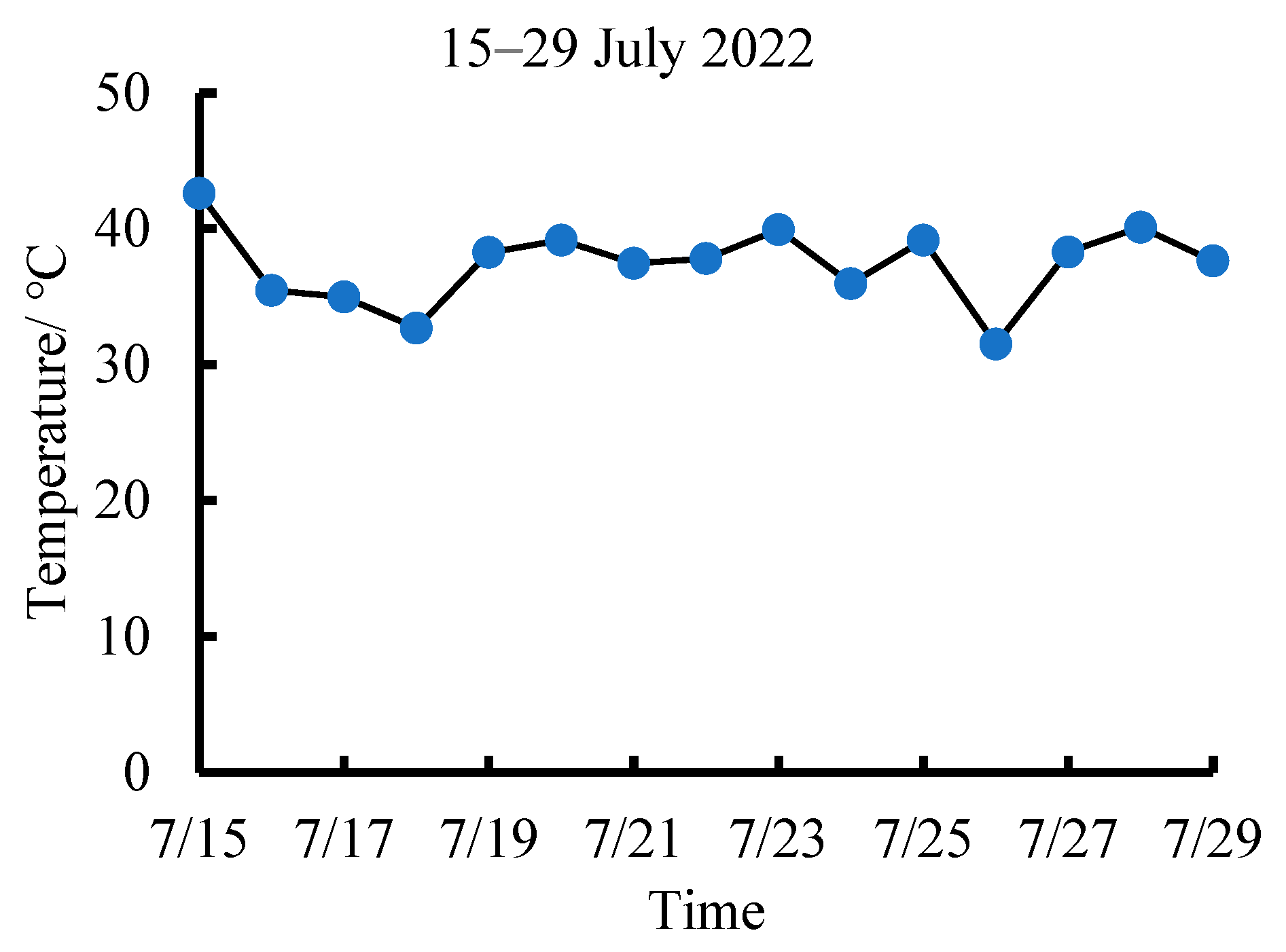
3.1. Variations in the Diurnal Canopy–Air Temperature Difference of Tea Plants
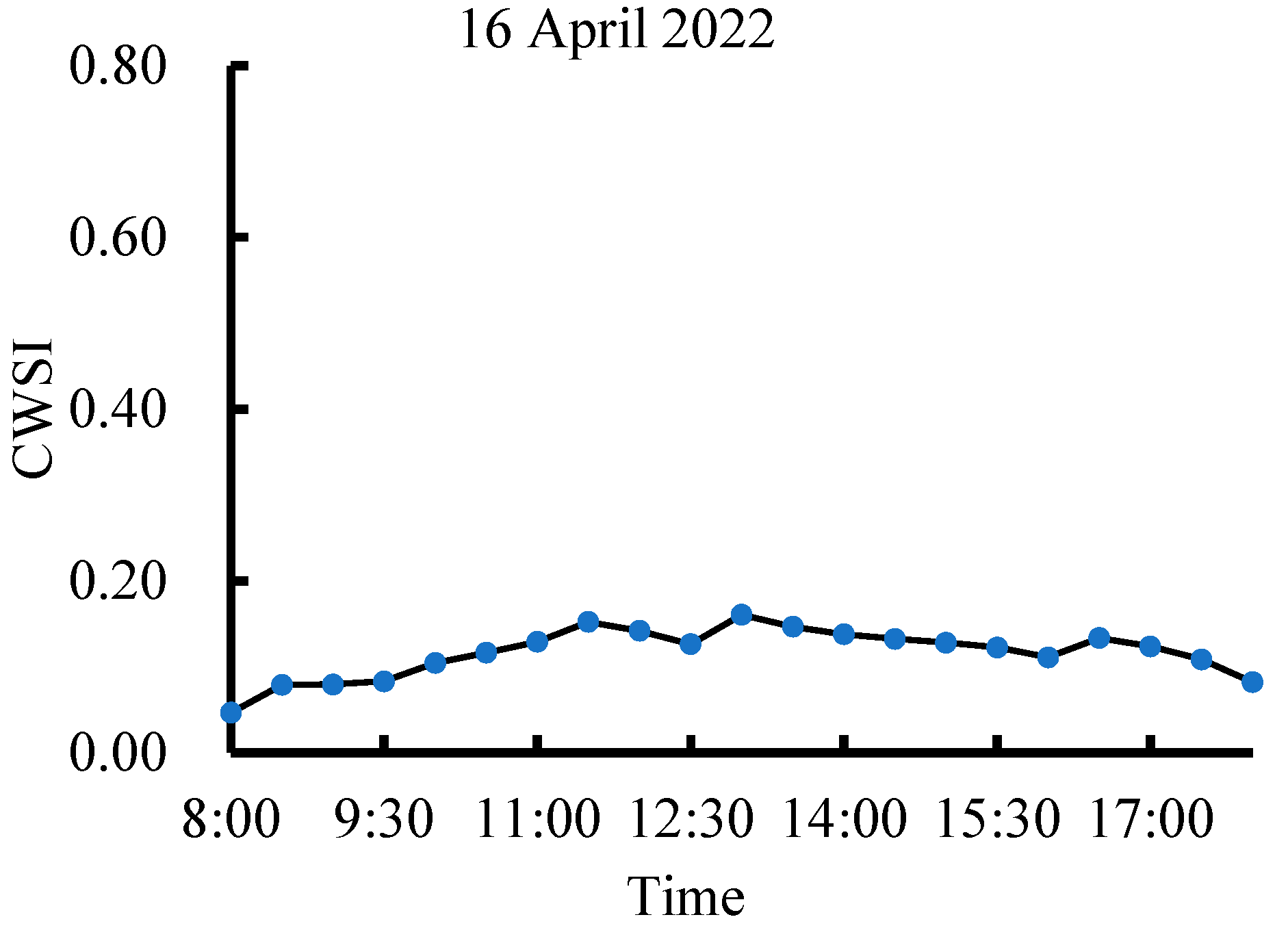
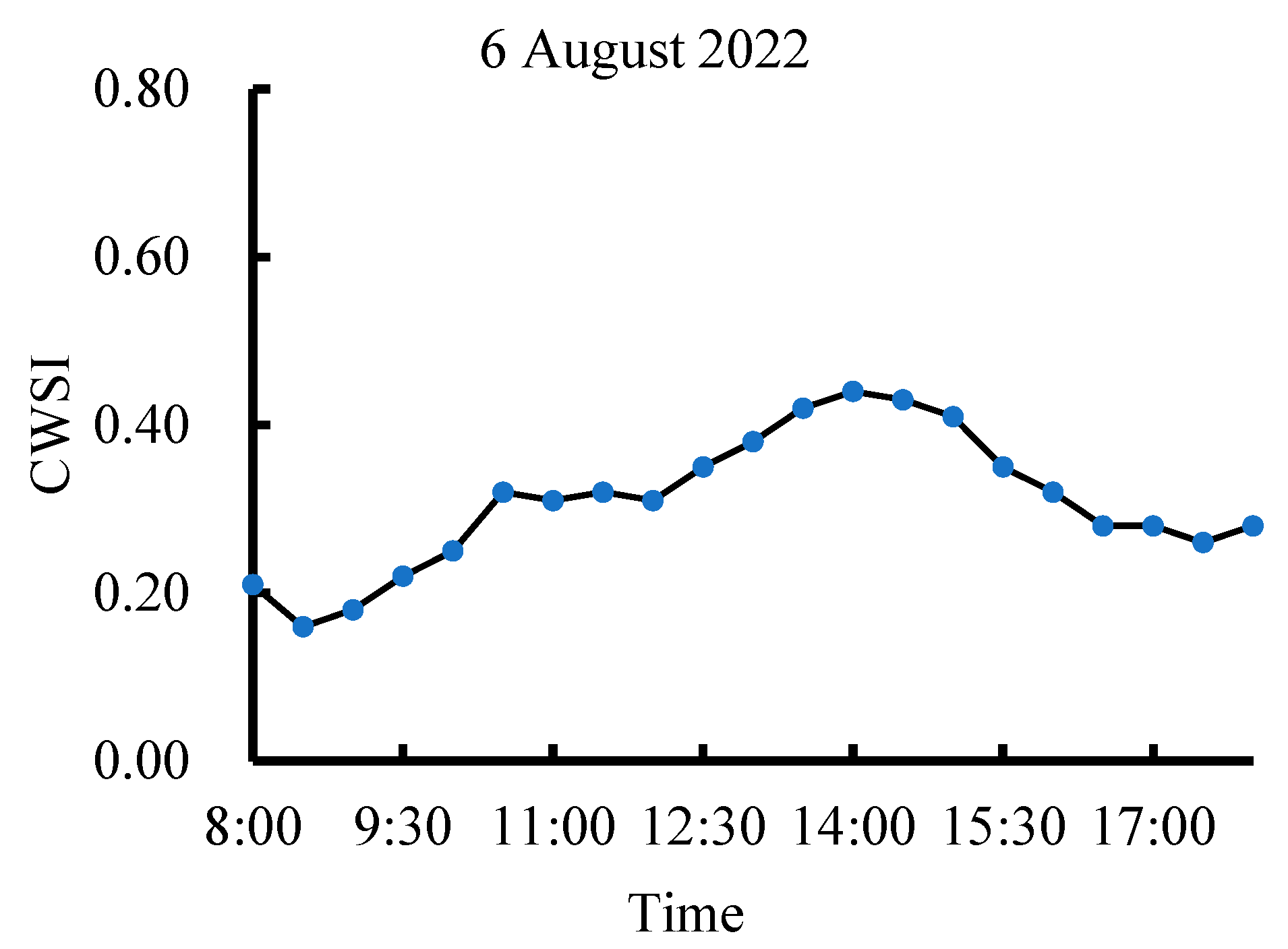
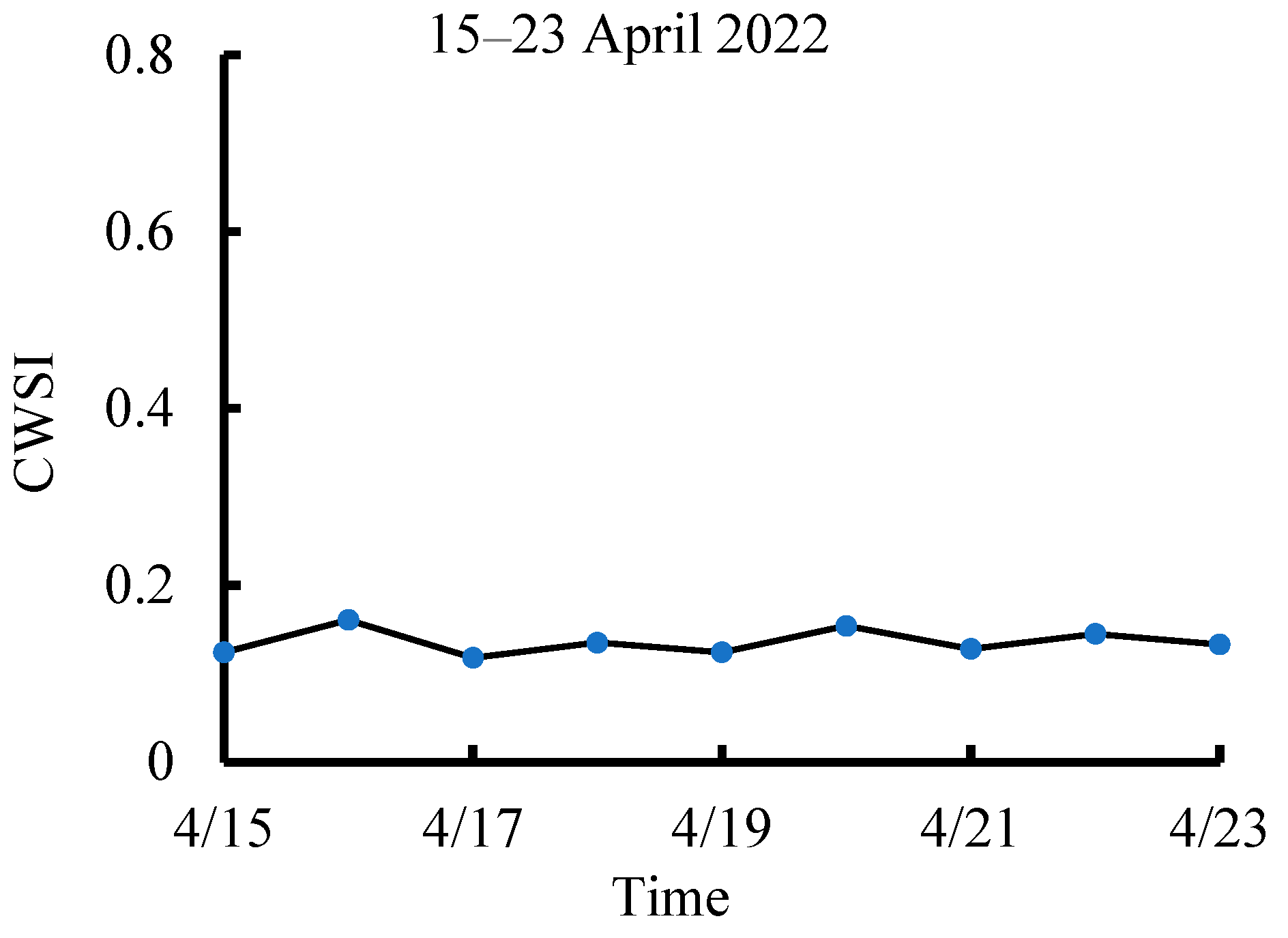
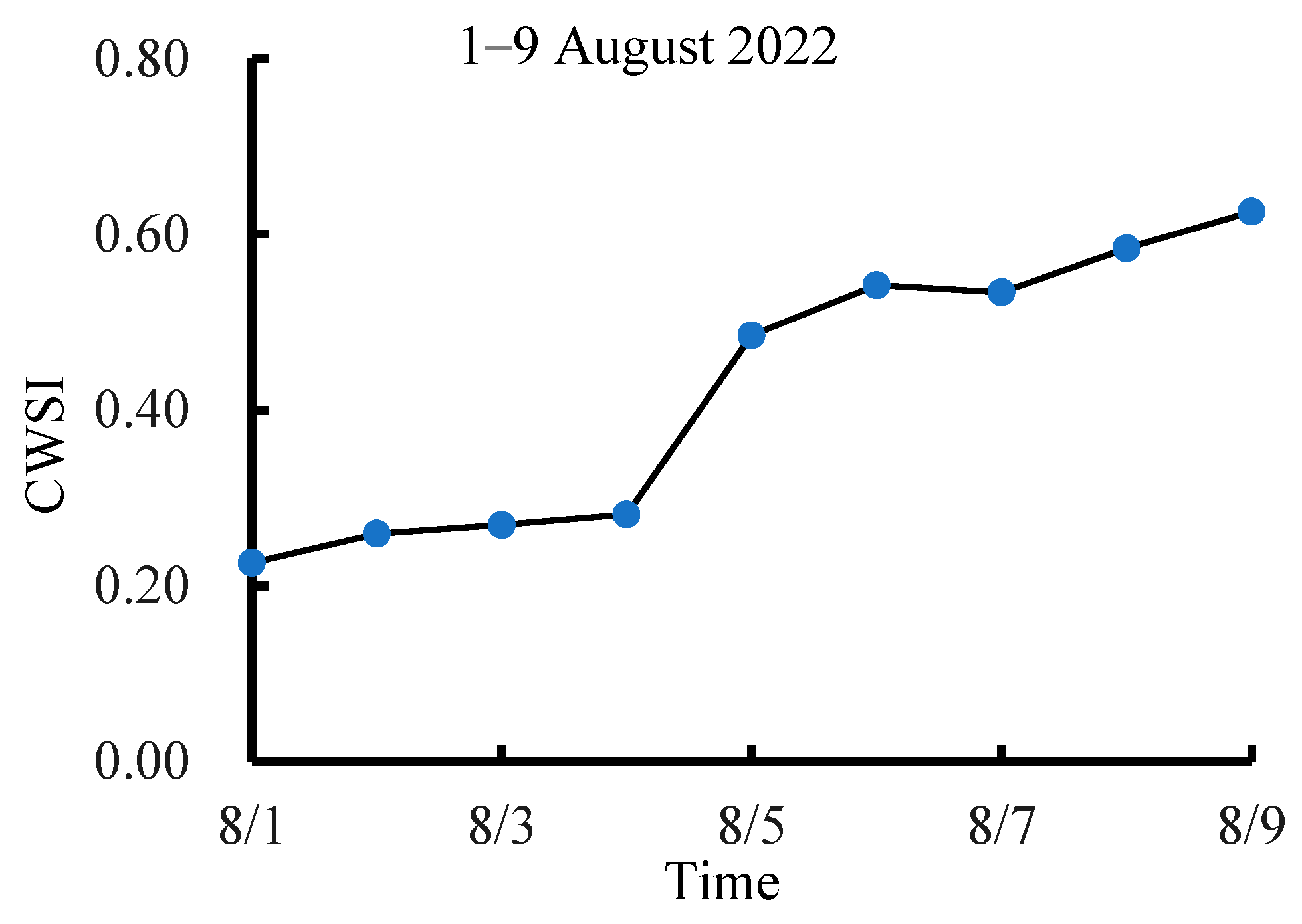
3.2. Dynamic Change in CWSI Characteristics at Different Growth Stages of Tea Plants
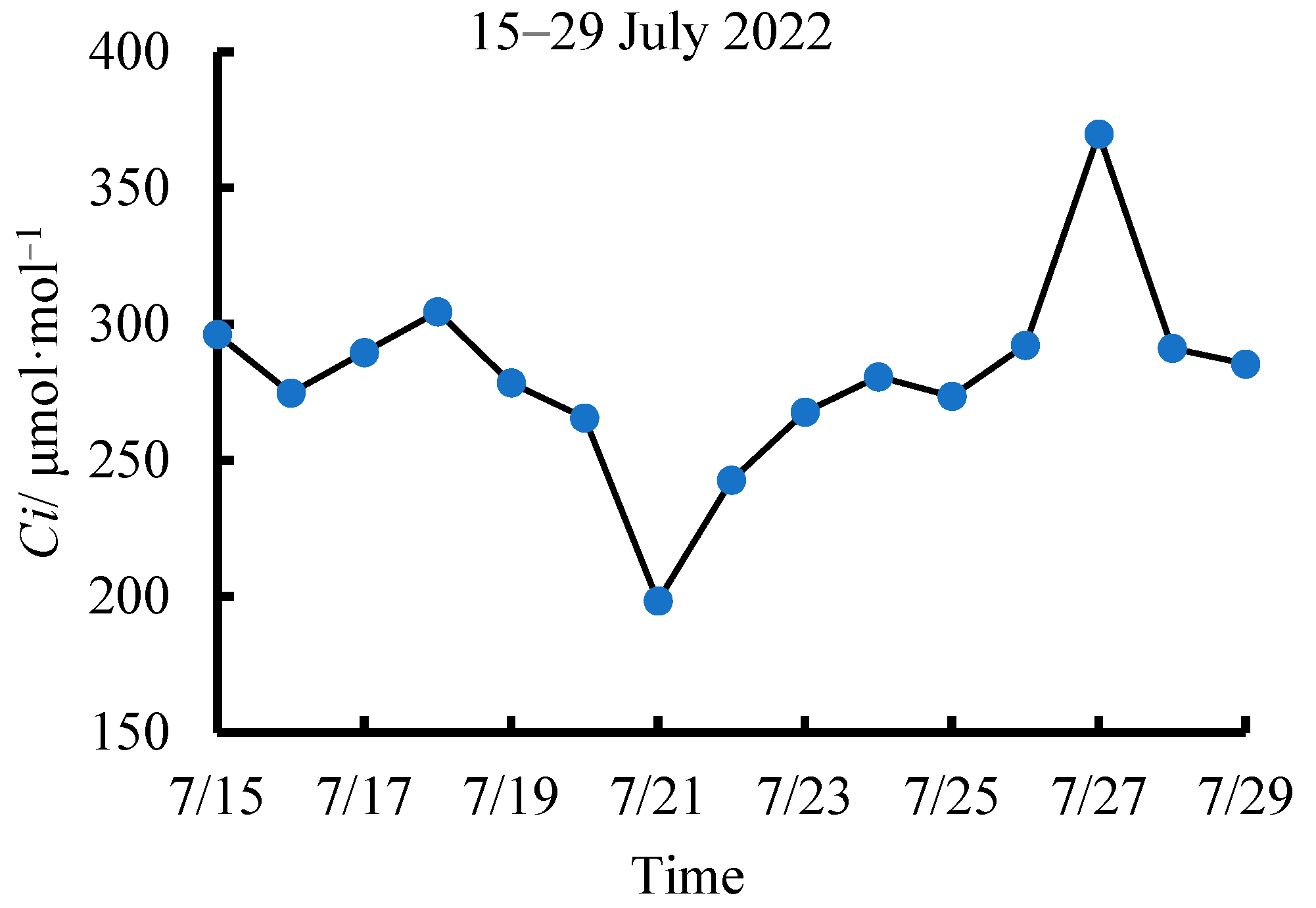
3.3. Determining the Critical CWSI Based on Ci
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, S.; Im, J.; Jang, E.; Rhee, J. Drought Assessment and Monitoring through Blending of Multi-Sensor Indices Using Machine Learning Approaches for Different Climate Regions. Agric. For. Meteorol. 2016, 216, 157–169. [Google Scholar] [CrossRef]
- Han, M.; Zhang, H.; DeJonge, K.C.; Comas, L.H.; Gleason, S. Comparison of Three Crop Water Stress Index Models with Sap Flow Measurements in Maize. Agric. Water Manag. 2018, 203, 366–375. [Google Scholar] [CrossRef]
- Luan, Y.; Xu, J.; Lv, Y.; Liu, X.; Wang, H.; Liu, S. Improving the Performance in Crop Water Deficit Diagnosis with Canopy Temperature Spatial Distribution Information Measured by Thermal Imaging. Agric. Water Manag. 2021, 246, 106699. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Peña-Angulo, D.; Beguería, S.; Domínguez-Castro, F.; Tomás-Burguera, M.; Noguera, I.; Gimeno-Sotelo, L.; El Kenawy, A. Global Drought Trends and Future Projections. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 2022, 380, 20210285. [Google Scholar] [CrossRef]
- Hu, H.; Lu, Y.; Hu, Y.; Ding, R. Evaluation of Two Surface Renewal Methods for Calculating the Sensible Heat Flux over a Tea Field Ecosystem in Hilly Terrain. Agronomy 2023, 13, 1302. [Google Scholar] [CrossRef]
- Ma, S.; Zheng, L.; Liu, X.; Zhang, K.; Hu, L.; Hua, Y.; Huang, J. Genome-Wide Identification of Brassicaceae Hormone-Related Transcription Factors and Their Roles in Stress Adaptation and Plant Height Regulation in Allotetraploid Rapeseed. Int. J. Mol. Sci. 2022, 23, 8762. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, Y.; Zhang, J.; Qian, X.; Li, X.; Sun, X. Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses. Int. J. Mol. Sci. 2024, 25, 1079. [Google Scholar] [CrossRef]
- Jin, J.; Zhao, M.; Jing, T.; Zhang, M.; Lu, M.; Yu, G.; Wang, J.; Guo, D.; Pan, Y.; Hoffmann, T.D.; et al. Volatile Compound-Mediated Plant–Plant Interactions under Stress with the Tea Plant as a Model. Hortic. Res. 2023, 10, uhad143. [Google Scholar] [CrossRef]
- Rigden, A.J.; Mueller, N.D.; Holbrook, N.M.; Pillai, N.; Huybers, P. Combined Influence of Soil Moisture and Atmospheric Evaporative Demand Is Important for Accurately Predicting US Maize Yields. Nat. Food 2020, 1, 127–133. [Google Scholar] [CrossRef]
- Wilson, T.G.; Kustas, W.P.; Alfieri, J.G.; Anderson, M.C.; Gao, F.; Prueger, J.H.; McKee, L.G.; Alsina, M.M.; Sanchez, L.A.; Alstad, K.P. Relationships between Soil Water Content, Evapotranspiration, and Irrigation Measurements in a California Drip-Irrigated Pinot Noir Vineyard. Agric. Water Manag. 2020, 237, 106186. [Google Scholar] [CrossRef]
- Braunworth, W.S.; Mack, H.J. The Possible Use of the Crop Water Stress Index as an Indicator of Evapotranspiration Deficits and Yield Reductions in Sweet Corn. J. Am. Soc. Hortic. Sci. 1989, 114, 542–546. [Google Scholar] [CrossRef]
- Sezen, S.M.; Yazar, A.; Daşgan, Y.; Yucel, S.; Akyıldız, A.; Tekin, S.; Akhoundnejad, Y. Evaluation of Crop Water Stress Index (CWSI) for Red Pepper with Drip and Furrow Irrigation under Varying Irrigation Regimes. Agric. Water Manag. 2014, 143, 59–70. [Google Scholar] [CrossRef]
- Yan, H.; Deng, S.; Zhang, C.; Wang, G.; Zhao, S.; Li, M.; Liang, S.; Jiang, J.; Zhou, Y. Determination of Energy Partition of a Cucumber Grown Venlo–Type Greenhouse in Southeast China. Agric. Water Manag. 2023, 276, 108047. [Google Scholar] [CrossRef]
- Yan, H.; Yu, J.; Zhang, C.; Wang, G.; Huang, S.; Ma, J. Comparison of Two Canopy Resistance Models to Estimate Evapotranspiration for Tea and Wheat in Southeast China. Agric. Water Manag. 2021, 245, 106581. [Google Scholar] [CrossRef]
- Yan, H.; Li, M.; Zhang, C.; Zhang, J.; Wang, G.; Yu, J.; Ma, J.; Zhao, S. Comparison of Evapotranspiration Upscaling Methods from Instantaneous to Daytime Scale for Tea and Wheat in Southeast China. Agric. Water Manag. 2022, 264, 107464. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, W.; Yan, H.; Ni, Y.; Akhlaq, M.; Zhou, J.; Xue, R. Effect of Micro-Spray on Plant Growth and Chlorophyll Fluorescence Parameter of Tomato under High Temperature Condition in a Greenhouse. Sci. Hortic. 2022, 306, 111441. [Google Scholar] [CrossRef]
- Huang, S.; Yan, H.; Zhang, C.; Wang, G.; Acquah, S.J.; Yu, J.; Li, L.; Ma, J.; Opoku Darko, R. Modeling Evapotranspiration for Cucumber Plants Based on the Shuttleworth-Wallace Model in a Venlo-Type Greenhouse. Agric. Water Manag. 2020, 228, 105861. [Google Scholar] [CrossRef]
- Jackson, R.D.; Kustas, W.P.; Choudhury, B.J. A Reexamination of the Crop Water Stress Index. Irrig. Sci. 1988, 9, 309–317. [Google Scholar] [CrossRef]
- Jackson, R.D.; Reginato, R.J.; Idso, S.B. Wheat Canopy Temperature: A Practical Tool for Evaluating Water Requirements. Water Resour. Res. 1977, 13, 651–656. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy Temperature as a Crop Water Stress Indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Bo, L.; Guan, H.; Mao, X. Diagnosing Crop Water Status Based on Canopy Temperature as a Function of Film Mulching and Deficit Irrigation. Field Crops Res. 2023, 304, 109154. [Google Scholar] [CrossRef]
- Liu, L.; Gao, X.; Ren, C.; Cheng, X.; Zhou, Y.; Huang, H.; Zhang, J.; Ba, Y. Applicability of the Crop Water Stress Index Based on Canopy-Air Temperature Differences for Monitoring Water Status in a Cork Oak Plantation, Northern China. Agric. For. Meteorol. 2022, 327, 109226. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho–Pereira, J.M.; Gonçalves, B.C.; Lopes, J.I.; Correia, C.M. Physiological Responses of Different Olive Genotypes to Drought Conditions. Acta Physiol. Plant. 2009, 31, 611–621. [Google Scholar] [CrossRef]
- Pilon, C.; Snider, J.L.; Sobolev, V.; Chastain, D.R.; Sorensen, R.B.; Meeks, C.D.; Massa, A.N.; Walk, T.; Singh, B.; Earl, H.J. Assessing Stomatal and Non-Stomatal Limitations to Carbon Assimilation under Progressive Drought in Peanut (Arachis hypogaea L.). J. Plant Physiol. 2018, 231, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of Drought Stress on Photosynthetic Response, Leaf Water Potential, and Stem Sap Flow in Two Cultivars of Bi-Leader Apple Trees (Malus × Domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Buckley, T.N.; Mott, K.A. Modelling Stomatal Conductance in Response to Environmental Factors. Plant Cell Environ. 2013, 36, 1691–1699. [Google Scholar] [CrossRef]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal Closure Is Induced by Hydraulic Signals and Maintained by ABA in Drought-Stressed Grapevine. Sci. Rep. 2015, 5, 12449. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Barón, M.; Bota, J.; Ducruet, J.-M.; Gallé, A.; Galmés, J.; Jiménez, M.; Pou, A.; Ribas-Carbó, M.; Sajnani, C.; et al. Photosynthesis Limitations during Water Stress Acclimation and Recovery in the Drought-Adapted Vitis Hybrid Richter-110 (V. berlandieri × V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef]
- Grassi, G.; Magnani, F. Stomatal, Mesophyll Conductance and Biochemical Limitations to Photosynthesis as Affected by Drought and Leaf Ontogeny in Ash and Oak Trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence—A Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Signarbieux, C.; Feller, U. Non–Stomatal Limitations of Photosynthesis in Grassland Species under Artificial Drought in the Field. Environ. Exp. Bot. 2011, 71, 192–197. [Google Scholar] [CrossRef]
- Idso, S.B. Non-Water-Stressed Baselines: A Key to Measuring and Interpreting Plant Water Stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the Stress-Degree-Day Parameter for Environmental Variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Reginato, R.J. Remote-Sensing of Crop Yields. Science 1977, 196, 19–25. [Google Scholar] [CrossRef]
- Pereira, L.S.; Allen, R.G.; Smith, M.; Raes, D. Crop Evapotranspiration Estimation with FAO56: Past and Future. Agric. Water Manag. 2015, 147, 4–20. [Google Scholar] [CrossRef]
- Kirnak, H.; Irik, H.A.; Unlukara, A. Potential Use of Crop Water Stress Index (CWSI) in Irrigation Scheduling of Drip–Irrigated Seed Pumpkin Plants with Different Irrigation Levels. Sci. Hortic. 2019, 256, 108608. [Google Scholar] [CrossRef]
- Yuan, G.; Luo, Y.; Sun, X.; Tang, D. Evaluation of a Crop Water Stress Index for Detecting Water Stress in Winter Wheat in the North China Plain. Agric. Water Manag. 2004, 64, 29–40. [Google Scholar] [CrossRef]
- Song, Z.; Wang, L.; Lee, M.; Yue, G.H. The Evolution and Expression of Stomatal Regulators in C3 and C4 Crops: Implications on the Divergent Drought Tolerance. Front. Plant Sci. 2023, 14, 1100838. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T. Dynamics of Changing Intercellular CO2 Concentration (Ci) during Drought and Determination of Minimum Functional Ci. Plant Physiol 1996, 111, 179–185. [Google Scholar] [CrossRef]
- Baker, J.T.; Mahan, J.R.; Gitz, D.C.; Lascano, R.J.; Ephrath, J.E. Comparison of Deficit Irrigation Scheduling Methods That Use Canopy Temperature Measurements. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2013, 147, 40–49. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, C.; Coenders Gerrits, M.; Acquah, S.J.; Zhang, H.; Wu, H.; Zhao, B.; Huang, S.; Fu, H. Parametrization of Aerodynamic and Canopy Resistances for Modeling Evapotranspiration of Greenhouse Cucumber. Agric. For. Meteorol. 2018, 262, 370–378. [Google Scholar] [CrossRef]
- DeJonge, K.C.; Taghvaeian, S.; Trout, T.J.; Comas, L.H. Comparison of Canopy Temperature-Based Water Stress Indices for Maize. Agric. Water Manag. 2015, 156, 51–62. [Google Scholar] [CrossRef]
- Irmak, S.; Haman, D.Z.; Bastug, R. Determination of Crop Water Stress Index for Irrigation Timing and Yield Estimation of Corn. Agron. J. 2000, 92, 1221–1227. [Google Scholar] [CrossRef]
- Taghvaeian, S.; Comas, L.; DeJonge, K.C.; Trout, T.J. Conventional and Simplified Canopy Temperature Indices Predict Water Stress in Sunflower. Agric. Water Manag. 2014, 144, 69–80. [Google Scholar] [CrossRef]
- Wang, D.; Gartung, J. Infrared Canopy Temperature of Early-Ripening Peach Trees under Postharvest Deficit Irrigation. Agric. Water Manag. 2010, 97, 1787–1794. [Google Scholar] [CrossRef]
- Agam, N.; Cohen, Y.; Berni, J.A.J.; Alchanatis, V.; Kool, D.; Dag, A.; Yermiyahu, U.; Ben-Gal, A. An Insight to the Performance of Crop Water Stress Index for Olive Trees. Agric. Water Manag. 2013, 118, 79–86. [Google Scholar] [CrossRef]
- Park, S.; Ryu, D.; Fuentes, S.; Chung, H.; O’Connell, M.; Kim, J. Dependence of CWSI-Based Plant Water Stress Estimation with Diurnal Acquisition Times in a Nectarine Orchard. Remote Sens. 2021, 13, 2775. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Bozkurt Çolak, Y.; Yazar, A.; Alghory, A.; Tekin, S. Evaluation of Crop Water Stress Index and Leaf Water Potential for Differentially Irrigated Quinoa with Surface and Subsurface Drip Systems. Irrig. Sci. 2021, 39, 81–100. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal Limitations to Photosynthesis and Their Critical Water Conditions in Different Growth Stages of Maize under Water Stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Wang, Q.; He, Q.; Zhou, G. Applicability of Common Stomatal Conductance Models in Maize under Varying Soil Moisture Conditions. Sci. Total Environ. 2018, 628–629, 141–149. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Depth (cm) | Size Composition | Bulk Density (g·cm−3) | Saturated VWC (%) | Field Capacity (%) | Soil Texture | ||
|---|---|---|---|---|---|---|---|
| <0.002 (mm) | 0.002–0.02 (mm) | 0.02–2 (mm) | |||||
| 0~10.0 | 1.2% | 32.3% | 66.5% | 1.34 | 35.5 | 26.5 | Sandy loam Sandy loam Sandy loam Loam |
| 10.0~20.0 | 2.0% | 29.3% | 68.7% | 1.36 | 36.9 | 27.1 | |
| 20.0~30.0 | 1.4% | 31.5% | 67.1% | 1.38 | 36.7 | 26.6 | |
| 30.0~40.0 | 6.1% | 42.6% | 51.3% | 1.56 | 40.4 | 25.9 | |
| 40.0~50.0 | 9.4% | 41.2% | 49.4% | 1.64 | 41.8 | 25.5 | Loam Loam |
| 50.0~60.0 | 8.4% | 41.4% | 50.2% | 1.64 | 41.5 | 25.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Zheng, J.; Hu, H.; Pan, Q.; Cui, L.; Hu, Y. Determination of Critical Crop Water Stress Index of Tea under Drought Stress Based on the Intercellular CO2 Concentration. Agronomy 2024, 14, 2154. https://doi.org/10.3390/agronomy14092154
Lu Y, Zheng J, Hu H, Pan Q, Cui L, Hu Y. Determination of Critical Crop Water Stress Index of Tea under Drought Stress Based on the Intercellular CO2 Concentration. Agronomy. 2024; 14(9):2154. https://doi.org/10.3390/agronomy14092154
Chicago/Turabian StyleLu, Yongzong, Jialiang Zheng, Huijie Hu, Qingmin Pan, Longfei Cui, and Yongguang Hu. 2024. "Determination of Critical Crop Water Stress Index of Tea under Drought Stress Based on the Intercellular CO2 Concentration" Agronomy 14, no. 9: 2154. https://doi.org/10.3390/agronomy14092154
APA StyleLu, Y., Zheng, J., Hu, H., Pan, Q., Cui, L., & Hu, Y. (2024). Determination of Critical Crop Water Stress Index of Tea under Drought Stress Based on the Intercellular CO2 Concentration. Agronomy, 14(9), 2154. https://doi.org/10.3390/agronomy14092154