1. Introduction
Farmers in northern Europe often replace winter barley with winter wheat as the preceding crop for winter oilseed rape (
Brassica napus L.; WOSR) causing a delayed WOSR sowing due to the later wheat harvest. Luteman and Dixon [
1] and Sieling et al. [
2] observed a reduced autumn and winter growth by delayed plant establishment, especially if it was later than 10 September, increasing the risk of winter kill [
3,
4,
5]. While Luteman and Dixon [
1] found only small yield penalties, Uzan et al. [
6] reported a decline in seed yield of canola cultivars with a delay in sowing date under Mediterranean environment conditions due to shortening the length of the reproductive period and consequently the potential grain-filling period. Also, Scott et al. [
7] showed large yield penalties if sowing occurred after mid-September.
In order to compensate for the poor autumn growth, an additional nitrogen (N) supply of about 30–50 kg·N·ha
−1 in autumn is intended to ensure crop N supply, adequate crop growth before winter and, consequently, a good overwinter survival. Autumn N, especially in early-sown oilseed rape, leads to a better above-ground growth [
2] and enhances N accumulation before winter [
8]. It is, however, debatable whether this increases seed yield and overwintering. Results of Ogilvy and Bastiman [
9] revealed that, although plots receiving nitrogen in the seedbed or at the two-leaf stage were more vigorous before winter compared with unfertilized plants, neither the number of plants established, the survival over winter, nor the seed yield were affected by this treatment. According to Engström et al. [
10] no effects on seed yield occurred after applying 30 or 60 kg·N·ha
−1 at sowing to WOSR in Sweden. In addition, Sieling and Kage [
11] observed that WOSR yield increased by 0.2 t·ha
−1 after an autumn N supply of 40 kg·N·ha
−1. In experiments in France, severe N deficiencies in autumn, described in terms of the nitrogen nutrition index, reduced shoot biomass, tap root biomass, leaf area index, and radiation-use efficiency compared with N sufficient treatments [
12]. However, no difference in seed yield occurred. The authors assumed that sufficient N release due to natural mineralization provided enough growth in autumn to ensure sufficient regrowth in spring. In other experiments, autumn N gave a small yield response where the preceding cereal straw was baled or incorporated instead of burning [
13,
14]. In addition, N application to boost crop growth rate may lead to plants being more susceptible to freezing in winter due to vigorous vegetative growth prior to the first killing frost or to increased leaching risk of N not taken up by the crop.
Spring N fertilization clearly increases WOSR seed yield. WOSR demands high amounts of N fertilizer often exceeding 200 kg N·ha
−1 to achieve maximum yields in high yielding environments with yield levels >4 t·ha
−1. Scott et al. [
15] and Zhao et al. [
16] suggested a maximum yield response to a N rate around 200 kg·N·ha
−1, which was in good agreement with the results of Bilsborrow et al. [
17] who obtained >85% of the maximum recorded yield with an application of 150 kg·N·ha
−1. Several approaches from, for example, France and Germany take the amount of N in the WOSR canopy at the end of autumn growth and/or at the beginning of spring growth into account [
18,
19,
20]. In one study of insignificant leaf losses over winter, a well-developed rapeseed canopy showed a higher leaf area index at the beginning of spring growth than a poor canopy, which, in consequence, allows for reduced N fertilization rates [
21]. Henke et al. [
19] showed that even if canopy N is lost over winter due to frost, optimum N requirements in spring decreased with increasing canopy N in late autumn. These results assumed a negative interaction between autumn N supply, promoting above-ground dry matter accumulation and N uptake, and spring N application [
2].
WOSR has the ability to take up more N than winter cereals between sowing and spring [
22]; however, relatively little of this N ends up in the seed [
23]. Compared to the N requirement, N offtake by the WOSR seeds is comparatively low, leading to low nitrogen harvest indices and high N balance surpluses (fertilizer N minus N offtake by the seeds), which can lead to high N leaching rates during periods of heavy rainfall [
23,
24,
25,
26,
27]. Thus, along with an increasing rate of N-fertilization, the rate of fertilizer-N recovery (apparent N use efficiency (NUE)) declines substantially [
26]. Due to only small yield effects, N fertilization in autumn is less N efficient in spring, thus most of the autumn N remains in the system. In field experiments reported by Sieling and Kage [
11], 40 kg·N·ha
−1 in autumn increased N offtake by the seeds by about 4 kg·N·ha
−1, thus 36 kg·N·ha
−1 were left in the soil changing the N balance.
In general, it is assumed that WOSR requires at least six to eight leaves, a root collar diameter of 1 cm, a tap root length of 20 cm, and about 1 g of dry matter (DM) per plant to secure optimal winter survival [
28]. Under the climate conditions of northern Europe, sowing date and N availability in autumn are the main factors influencing WOSR growth before winter, which can be managed by the farmers [
2]. However, no data are available on which to base thresholds for optimal autumn growth.
Based on data from a three-year field experiment, this study aims (i) to investigate how delayed sowing affects seed yield, N offtake with the seeds, and N use efficiency of WOSR; (ii) to test the ability of autumn and spring N fertilization to compensate for the negative effects of a delayed sowing method; and (iii) to estimate the minimum autumnal growth for optimal seed yield. We hypothesize that delaying sowing reduces seed yield, N offtake, and N use efficiency, while autumn N supply may increase these factors, especially in late sown canopies. In addition, spring N fertilization may be reduced if additional N was applied in autumn.
3. Discussion
The experimental design allowed for an investigation of the effects of sowing method, autumn N, and spring N supply, as well as their interactions, on yield and N offtake by the seeds of WSOR. Due to varying sowing methods and autumn N supplies, 16 different WOSR canopies were established in autumn, each combined with increasing spring N fertilization in order to estimate N response curves separately for each canopy. The results confirmed the current recommendation that under the climatic conditions of northern Europe WOSR should ideally be sown between the middle and end of August, whereas plant establishment after mid-September clearly decreased seed yield, which was in agreement with [
7]. It should be noted that, if the weather conditions in the following spring are favorable, even an WOSR crop sown in the third week of September with two to four leaves at the end of autumn growth was able to achieve more than 5 t·ha
−1 [
29]; however, since long-term weather forecasts are not yet available, the risk of yield penalties increases with shifting WOSR sowing to the end of September.
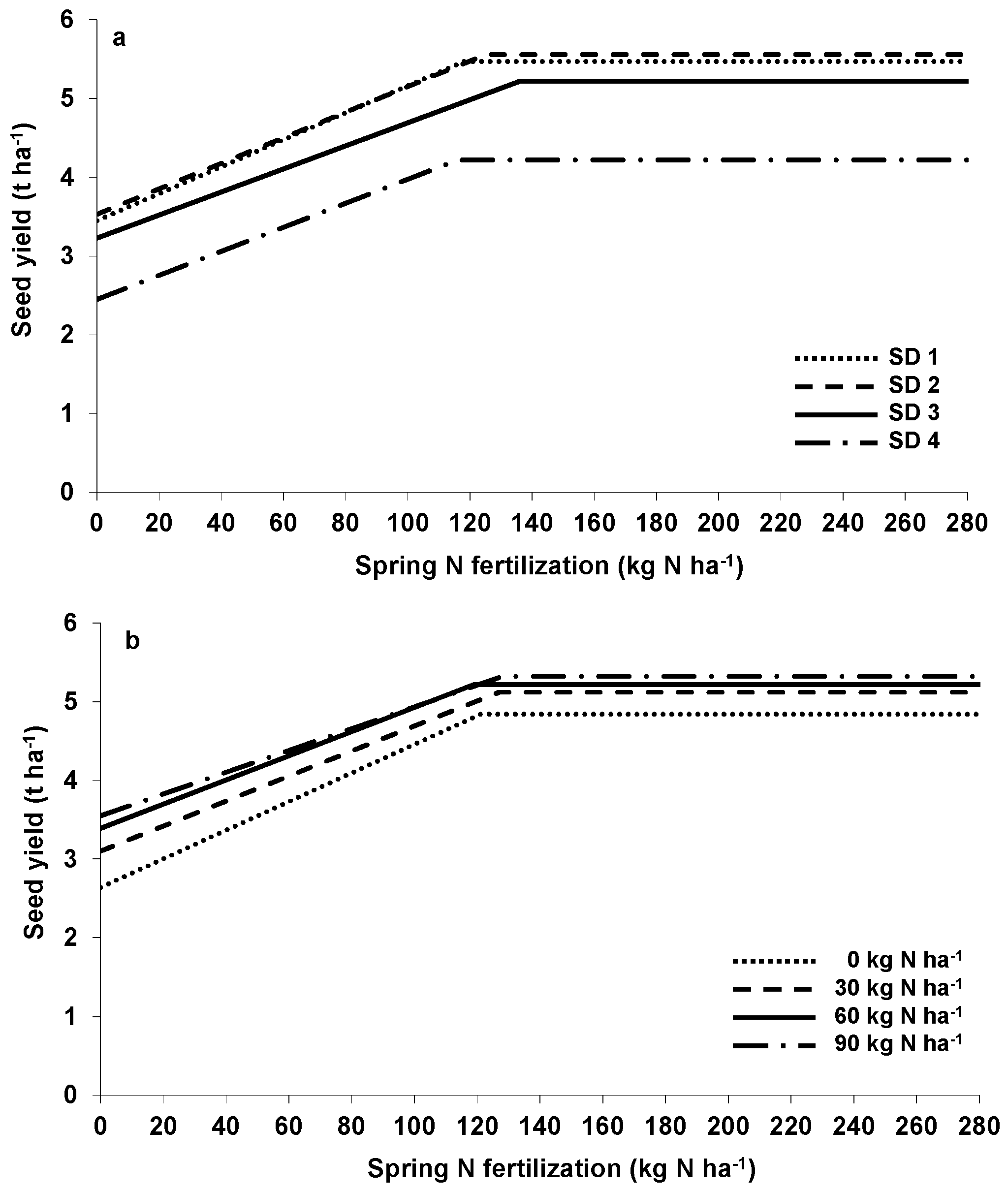
In combination with late sowing, farmers often apply additional N in autumn to ensure adequate WOSR growth before winter. No significant interaction between the sowing method and autumn as well as spring N fertilization on seed yield could be observed in our experiments (
Table 1). Applying at least 30 kg·N·ha
−1 in autumn increased WOSR yield at all sowing methods averaged over three years, although unfertilized WOSR canopies established in August already accumulated about 100 g·DM·m
−2 and 34 kg·N·ha
−1 at the end of autumn growth compared to 3 g·DM·m
−2 and <2 kg·N·ha
−1 in the late sown ones [
29]. In addition, the N amount in early sown canopies clearly exceeded the threshold for optimal autumn N uptake, as shown in
Figure 4. However, the pathway(s) which allowed autumn N to significantly increase seed yield for the early sowing method treatment require(s) further study. On the one hand, it can be argued that despite the often relatively small N uptake before winter, the additional N supply in autumn enables the plants to better utilize (N) resources in spring, thus indicating a physiological pathway. On the other hand, autumn N might act as advanced spring N fertilization if N is not leached out of the potential rooting zone during winter. In this case, an interaction between autumn and spring N treatments should be expected; however, it was not observed in our experiments (
Figure 1b,
Figure 2b, and
Table 2). In contrast, Henke et al. [
19] provided an approach which took autumn N uptake into account when estimating spring N fertilization, thus assuming an autumn N by spring N interaction. The authors suggested reducing the spring N amount if N in the canopy exceeds 50 kg·N·ha
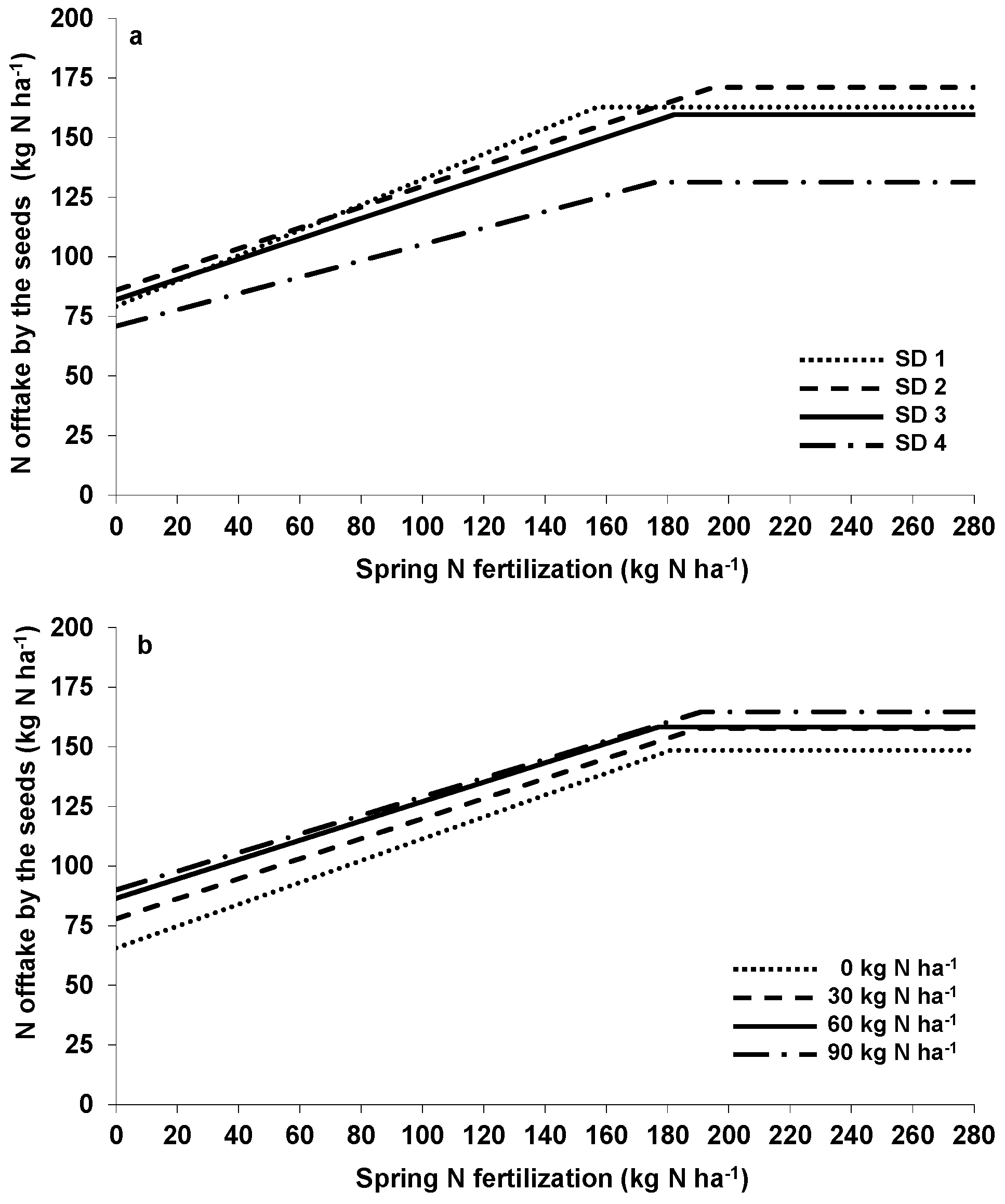
−1 at the end of autumn growth. With only small increases in N offtake by the seeds (
Table 3) and the resulting small NUE (
Table 4) in mind, autumn N should not exceed 30 kg·N·ha
−1.
In order to describe the N effects, a “Linear Response and Plateau” approach (LRP) was chosen, since using quadratic N response curves resulted in quite high N amounts required to achieve yield maximum due to a less steep slope compared to wheat [
30,
31]. Although the quadratic approach seems to be safer in terms of preventing financial losses under conditions of uncertainty, the risk of high positive N balances rises and the introduction of penalty functions may therefore favor the LPR model, especially against the background of high N surpluses in WOSR growing systems [
31].
Based on the LRP approach, the minimum amount of spring N required for maximum seed yield ranged, based on average of the three experimental years, between 120 and 140 kg·N·ha
−1, regardless of the sowing method and the autumn N supply, which was lower than local recommendation according to the “Düngeverordnung” [
32], the national implementation of the EU Nitrate Directive, as well as results from other authors [
26,
33]. Since N fertilization also increased seed N concentration, optimal N supply for the maximum N offtake by the seeds was higher (about 180–190 kg·N·ha
−1,
Figure 2a,b and
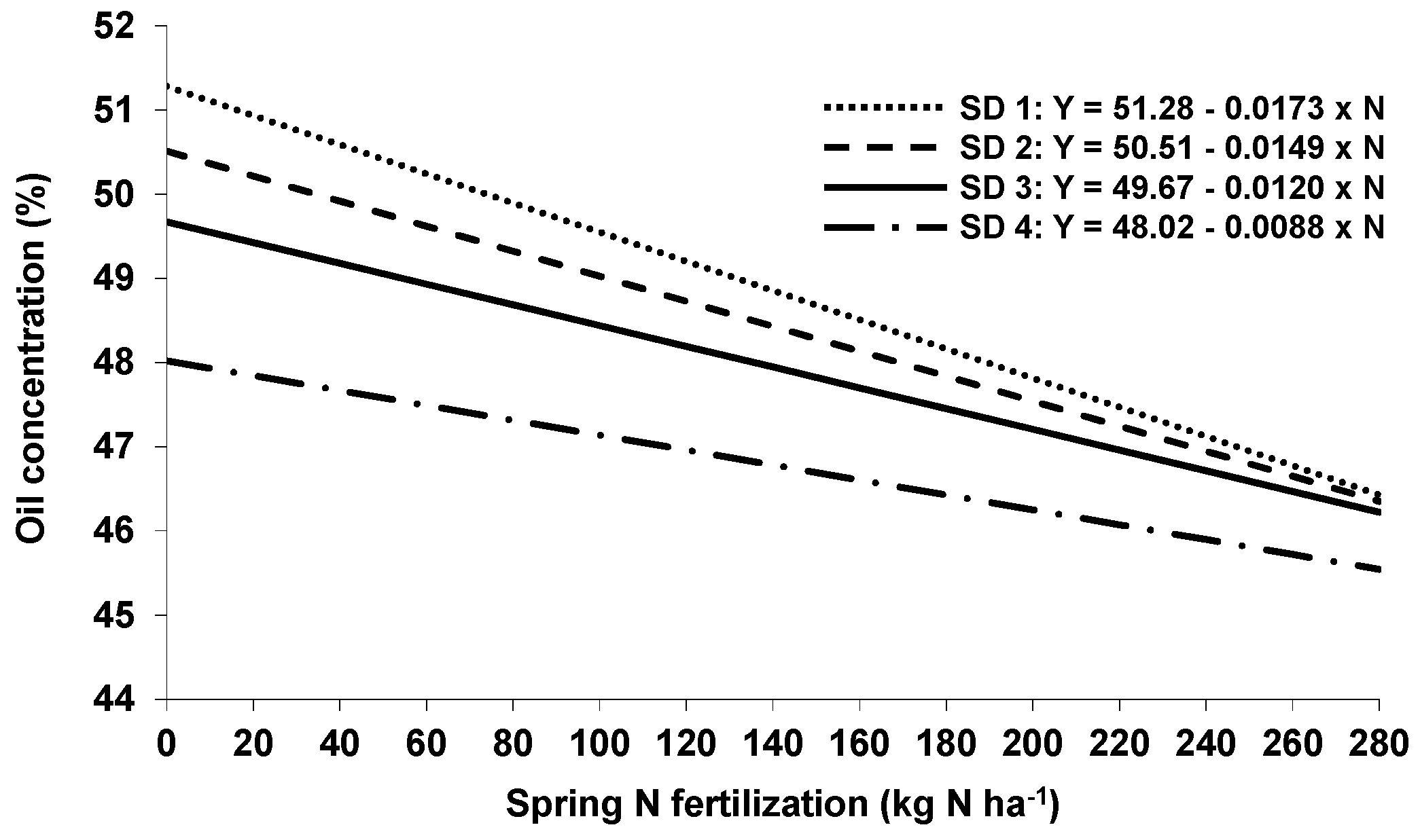
Table 2). In addition, delayed sowing methods also led to a higher seed N concentration, whereas autumn N had no effects [
29].
N-use efficiency (NUE) for autumn (spring) N was calculated by comparing the seed yield according to the average of the spring (autumn) N treatments in relation to the amount of applied N in autumn (spring). In general, NUE of WOSR is lower compared to wheat or barley [
23,
30,
34], which often has been associated with a low foliar N remobilization, thus with a high N concentration in WOSR leaves dropping off during the growth period, especially after flowering [
35,
36,
37]. NUE decreased with increasing N amounts (
Table 4). WOSR utilized autumn N to a lower extent than spring applied N [
9,
11]. Autumn NUE was highest when WOSR was sown in the first week of September, which was mainly due to the lower N offtake of the unfertilized plots compared to earlier sowing, while the autumn N fertilized ones showed similar values (
Table 3). Therefore, NUE alone is not suitable for evaluating the sustainability of a cropping system as the N offtake and, in consequence, the N balance (N surplus) have to be taken into account. From an environmental point of view, N application in autumn, if any, should not exceed 30 kg·N·ha
−1, since most of the N remained in the system, increasing the risk of losses in the subsequent crops [
27]. According to the presented results using the LPR approach, applying 120–130 kg·N·ha
−1 in spring were sufficient to achieve the highest seed yields. However, using quadratic N response curves, 180–200 kg·N·ha
−1 were required to get the economic optimal seed yield [
29], representing an application rate that was similar to that used by the farmers.
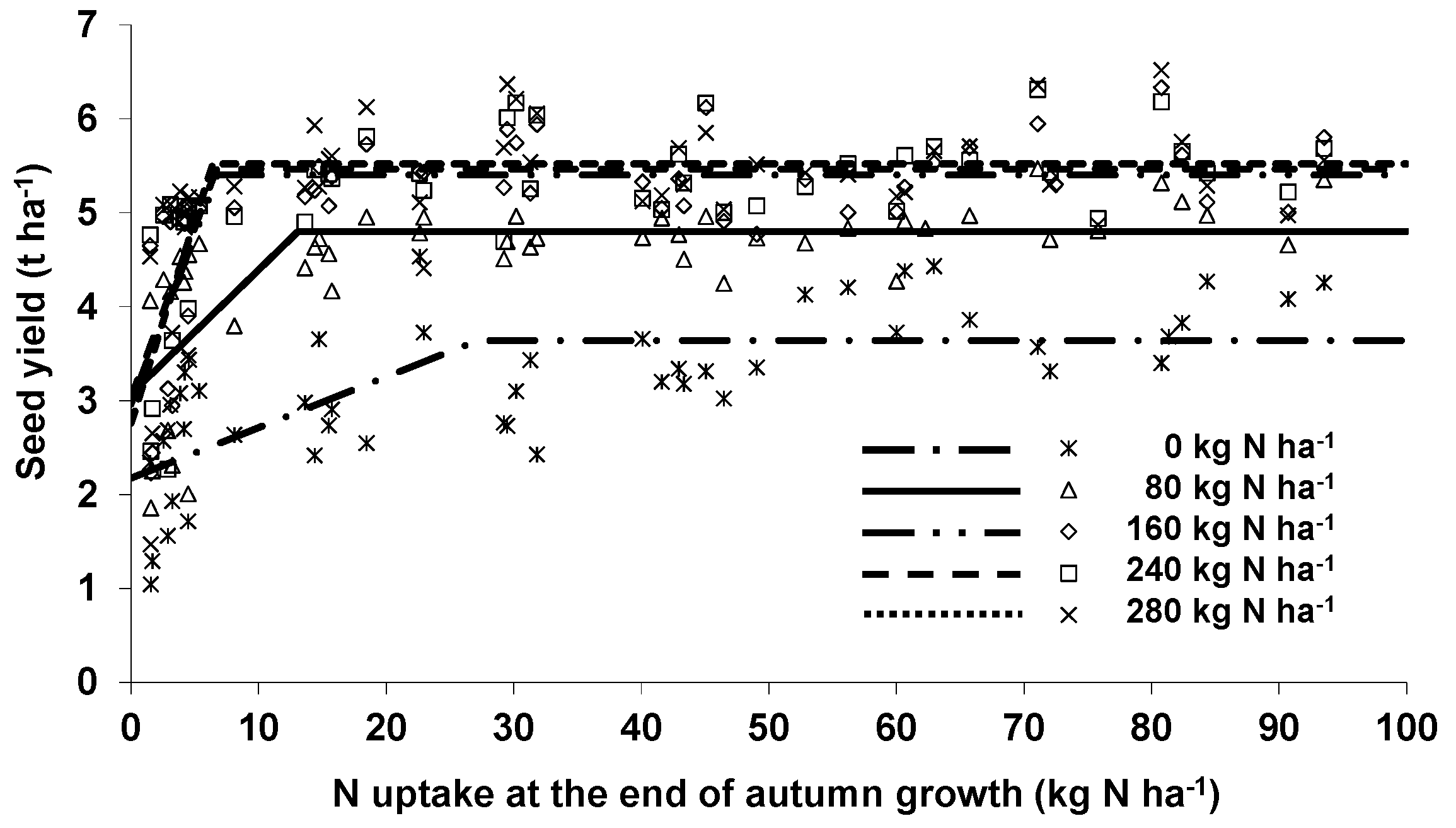
The estimation of the minimum amount of N taken up by the canopy at the end of autumn in order to achieve maximum seed yield was based on the LPR approach, where only the spring N fertilization was additionally considered. Including autumn N supply did not improve the relationship (except if the canopy remained unfertilized in spring), indicating that the above-ground N accumulation at the end of autumn growth adequately covered the effects of autumn N application. Optimal autumn N uptake depended on the amount of spring N. If there are plans for 160 kg·N·ha
−1 or more to be applied in spring, an N uptake of 10 kg·N·ha
−1 before winter should enable the canopies to produce optimal seed yield. Even under the less favorable conditions occurring in autumn 2010, 13 kg·N·ha
−1 seemed to be sufficient [
29]. Assuming a quadratic relationship, about 40 kg·N·ha
−1 were required to achieve 95% of the yield maximum [
29] being similar to the 50 kg·N·ha
−1 suggested by Henke et al. [
19] to represent an average rapeseed canopy which will be fertilized in spring according to the official recommendation. However, a threshold of 1 g single plant DM for minimum autumn growth which is often used by the farmers results in about 20 kg·N·ha
−1 assuming 40 plants·m
−2 and 5% N concentration in the above-ground biomass.
4. Materials and Methods
4.1. Site and Soil
The field experiment was carried out on a pseudogleyic sandy loam (Luvisol: 100 g·kg−1 clay, pH 6.6, 86 mg·kg−1 P, 79 mg·kg−1 K, 150 g·kg−1 Mg, 13.8 g·kg−1 Corg, 1.1 g·kg−1 Norg) at the Hohenschulen Experimental Farm (10.0° E, 54.3° N, 30 m above see level) of the Kiel University, located in northern Germany 15 km west of Kiel (Schleswig-Holstein).
The climate of NW Germany can be described as humid. Total annual rainfall averages 750 mm at the experimental site, with ca. 400 mm received during April–September, the main growing season, and ca. 350 mm during October–March. In all three years, the mean air temperature in October was below the long-term average. Especially in autumn 2010, the low temperature in August until November, in combination with very high rainfall in August and September, resulted in poor autumn growth in all treatments (
Table 5).
4.2. Treatments
In 2009/2010, 2010/2011, and 2012/2013, a field experiment with winter oilseed rape (WOSR) cv. Visby was established to test the effects of sowing method (SD) and N application in autumn and spring on crop growth in autumn, seed yield, and plant N recovery. Heavy rainfall in August 2011 prevented a proper crop establishment, therefore, no oilseed rape was grown in that year. In order to create sufficiently differentiated canopies, the four sowing dates ranged between the first week of August and the third week of September (
Table 6). The seed density was increased with delaying the sowing dates resulting in 35, 40, 50, and 70 seeds·m
−2, respectively, according to common farmers’ practice. Since sowing date was confounded by seed rate, the term ‘sowing method’ is used throughout the paper. In addition, each sowing method received four autumn N treatments (0, 30, 60, and 90 kg·N·ha
−1 as calcium ammonium nitrate with 27% N; for application dates, see
Table 6), resulting in 16 sowing method by N supply combinations. Each of these 16 different canopies was fertilized with five N amounts (0/0, 40/40, 80/80, 120/120, and 140/140 kg·N·ha
−1 as calcium ammonium nitrate with 27% N) in spring in order to estimate separate N response curves. N was applied in split-dressings at the beginning of spring growth and at stem elongation (for application dates, see
Table 6).
The sowing methods as main plots were randomized with four replicates, while the N treatments were randomized within the main plots. This experimental design allowed for analyzing the sowing method effects separately for each year. The single plot size was 3 m × 12 m. Winter barley was the preceding crop; its straw remained on the plots. In general, the plots were ploughed within one day before sowing. Crop management not involving the treatments (e.g., P and K supply, soil tillage, pesticide application) were applied according to local recommendations to achieve optimal yield.
4.3. Measurements
In all sowing methods plant samples were taken at the end of autumn growth. Plants were dug up from an area of 1 m2. After washing, above-ground biomass was analyzed for dry matter (DM) and N concentration using the near infrared spectroscopy (NIRS) analysis. Autumn N uptake was calculated by multiplying above-ground DM and the respective N concentration.
In order to minimize border effects, only a core of 1.75 m × 6 m (= 10.5 m2) was combine harvested at maturity to determine seed yield (standardized to t·ha−1 with 91% DM based on the moisture content of a seed subsample). Oil and N concentration of the seeds (at 100% DM) were determined by NIRS. N offtake by the seeds results from the product of seed yield DM and N concentration.
Apparent N use efficiency (NUE) was defined as the difference between the N offtake by the seeds in the fertilized and unfertilized treatment in relation to the N amount. Since no interaction between autumn and spring N fertilization occurred, NUE of the autumn (spring) N application was calculated based on the average of the spring (autumn) N treatments.
4.4. Statistical Analysis
First, the effects of sowing methods (SD), autumn and spring N fertilization were analyzed using a mixed model approach (SAS 9.4, Proc MIXED, SAS Institute Inc., Cary, NC, US) with “year” and “replication” as random terms. Since the SD by autumn N by spring N interaction was not significant, only the interactions between two factors as average on the remaining factor are presented.
Second, the effects of spring N application on seed yield and N offtake by the seeds were estimated by using a “Linear-Plateau” approach (Proc NLIN) separately for the sowing methods and the autumn N treatments, respectively.
The ‘Linear-Plateau’ model is specified by three parameters a, b, and P as follows:
where Y denotes the DM yield (t·ha
−1) or the N offtake by the seeds (kg·N·ha
−1), X the application rate of N (kg·N·ha
−1) or the N uptake in autumn, a the intercept, b the linear coefficient, and ε the error term. Nopt is the nitrogen application rate at the intersection of the linear model and the plateau yield P (
Table 2). Function parameters were compared by a modified t-test based on Zar [
38].
Seed oil concentration was analyzed by performing an analysis of covariance (Proc GLM).
A p-value of 0.05 was considered to be significant in the statistical analyses.








{kind=link}
{kind=link}
{kind=link}
{kind=link}