Leguminous Alley Cropping Improves the Production, Nutrition, and Yield of Forage Sorghum

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
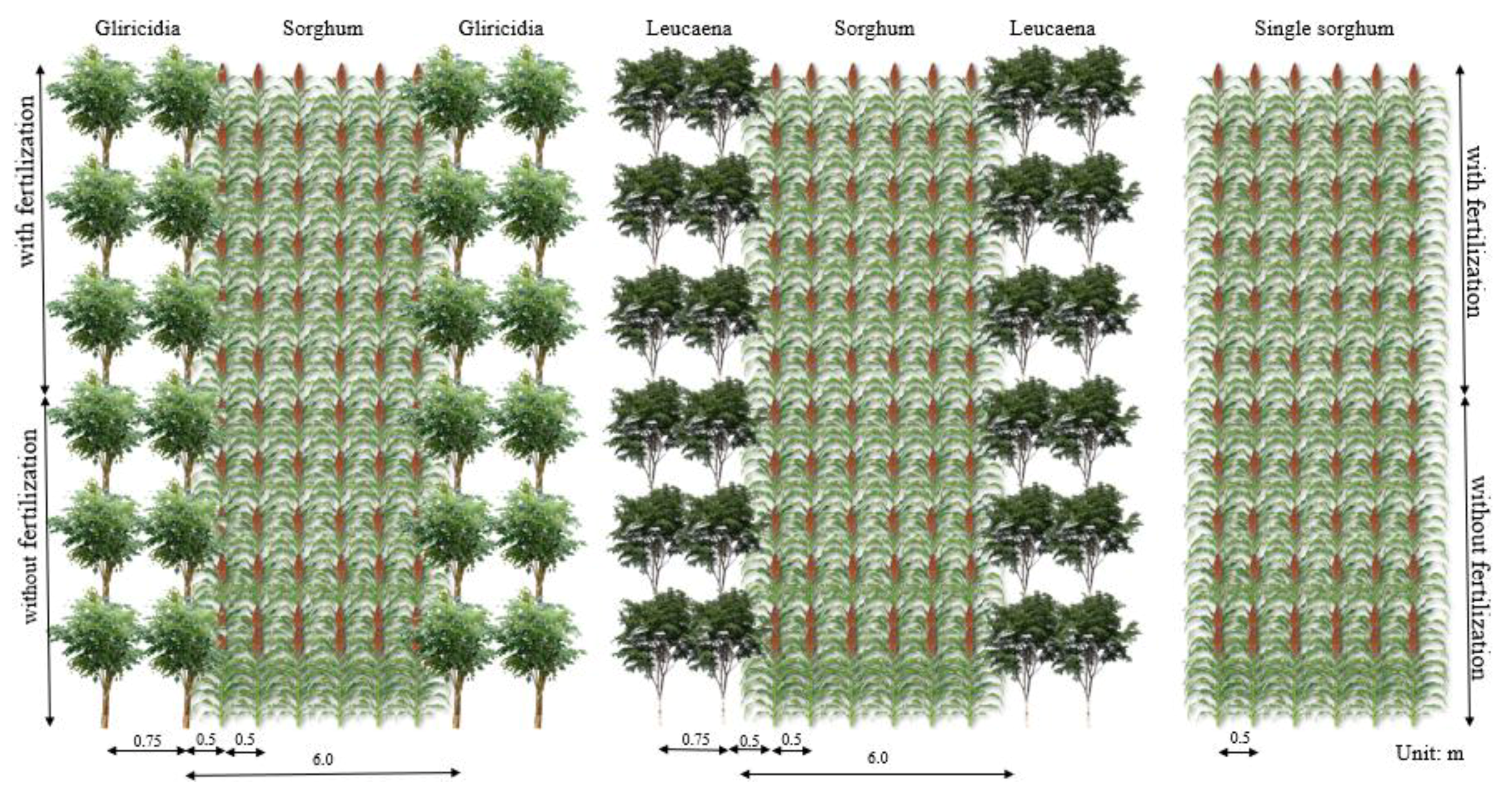
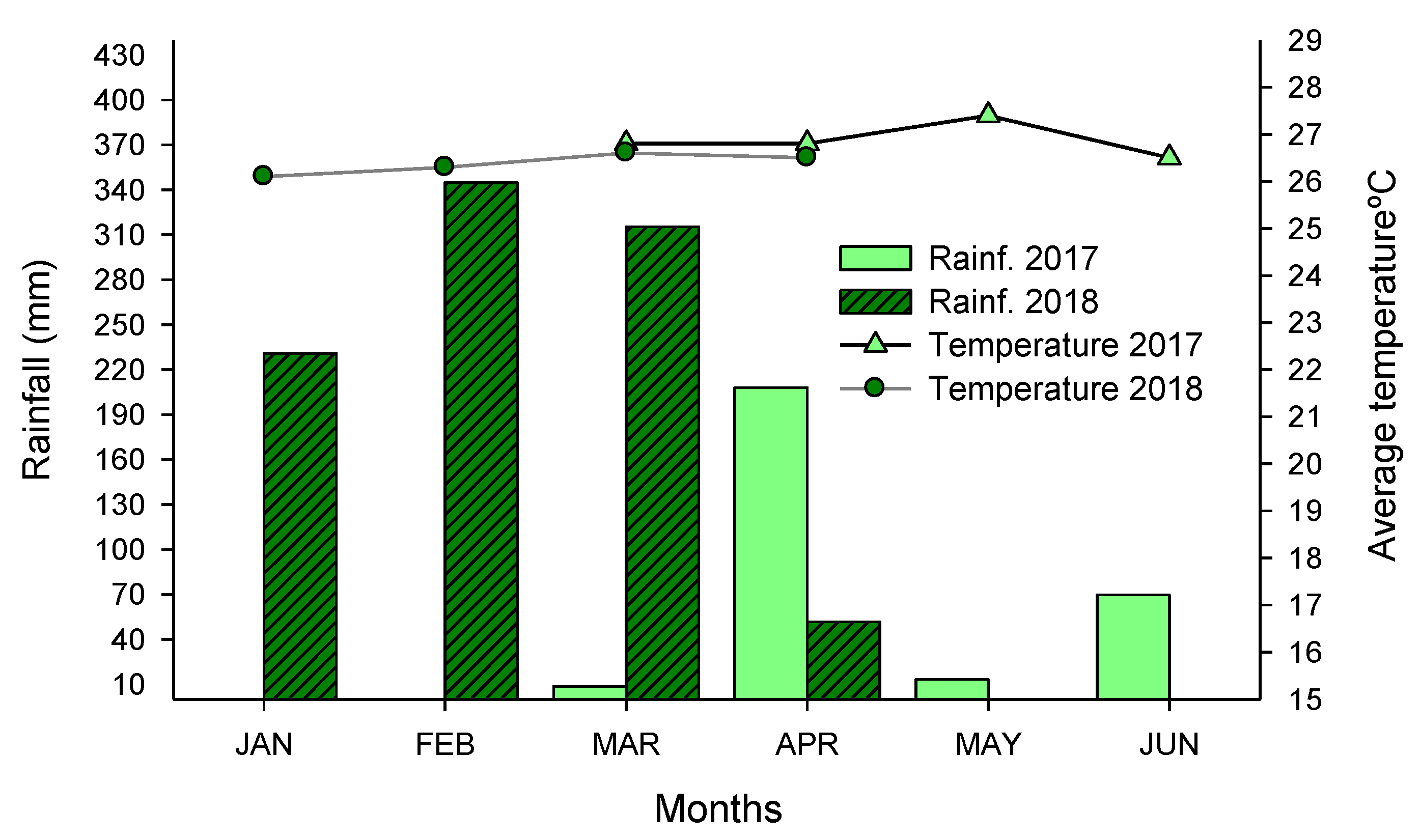
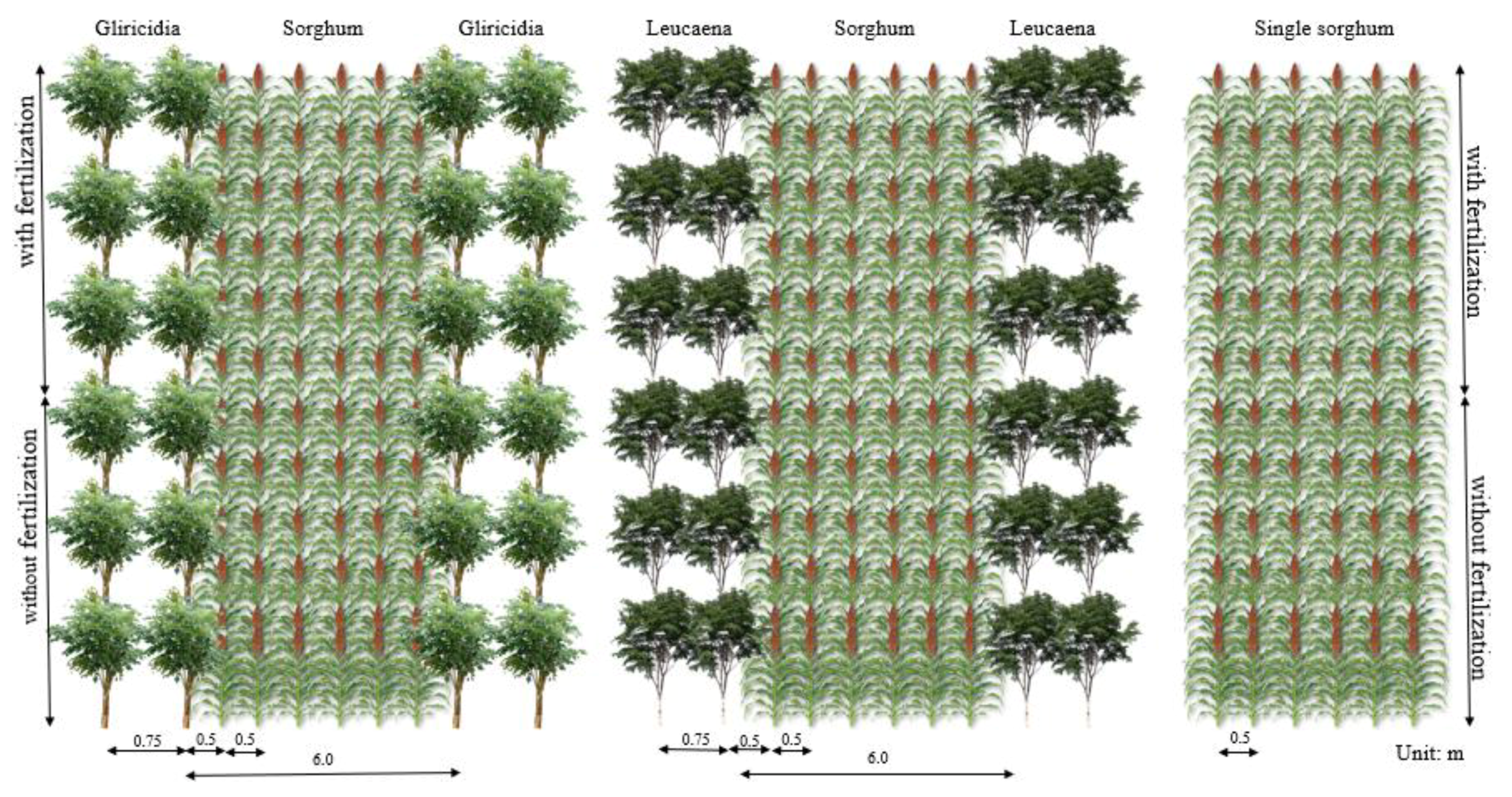
2.1. Experimental Area and Treatments
2.2. Establishment of Gliricidia and Leucaena Alleys and Forage Sorghum
2.3. Analysis of Plant Tissue and Sorghum Production
2.4. Statistical Analysis
3. Results
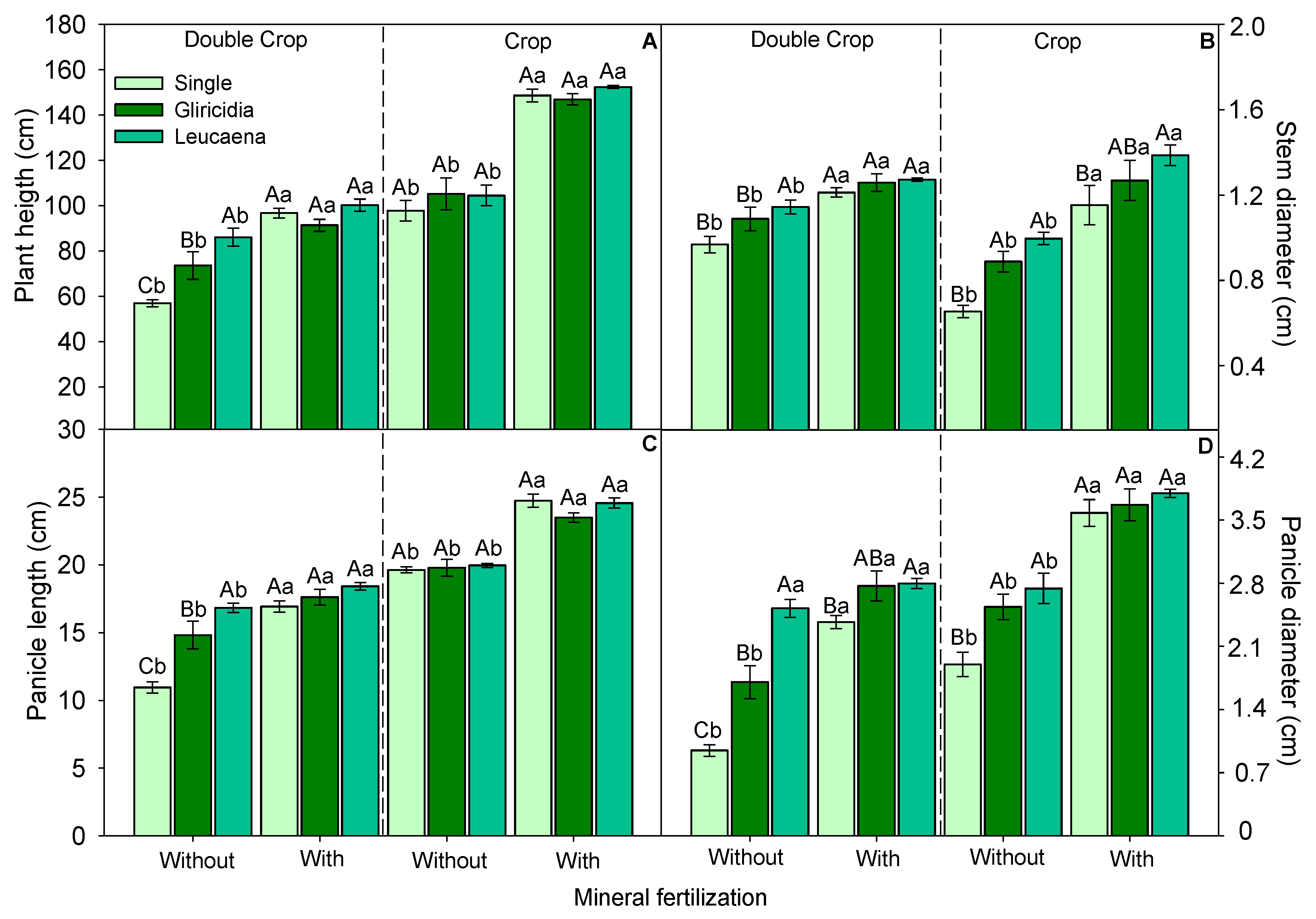
3.1. Sorghum Growth
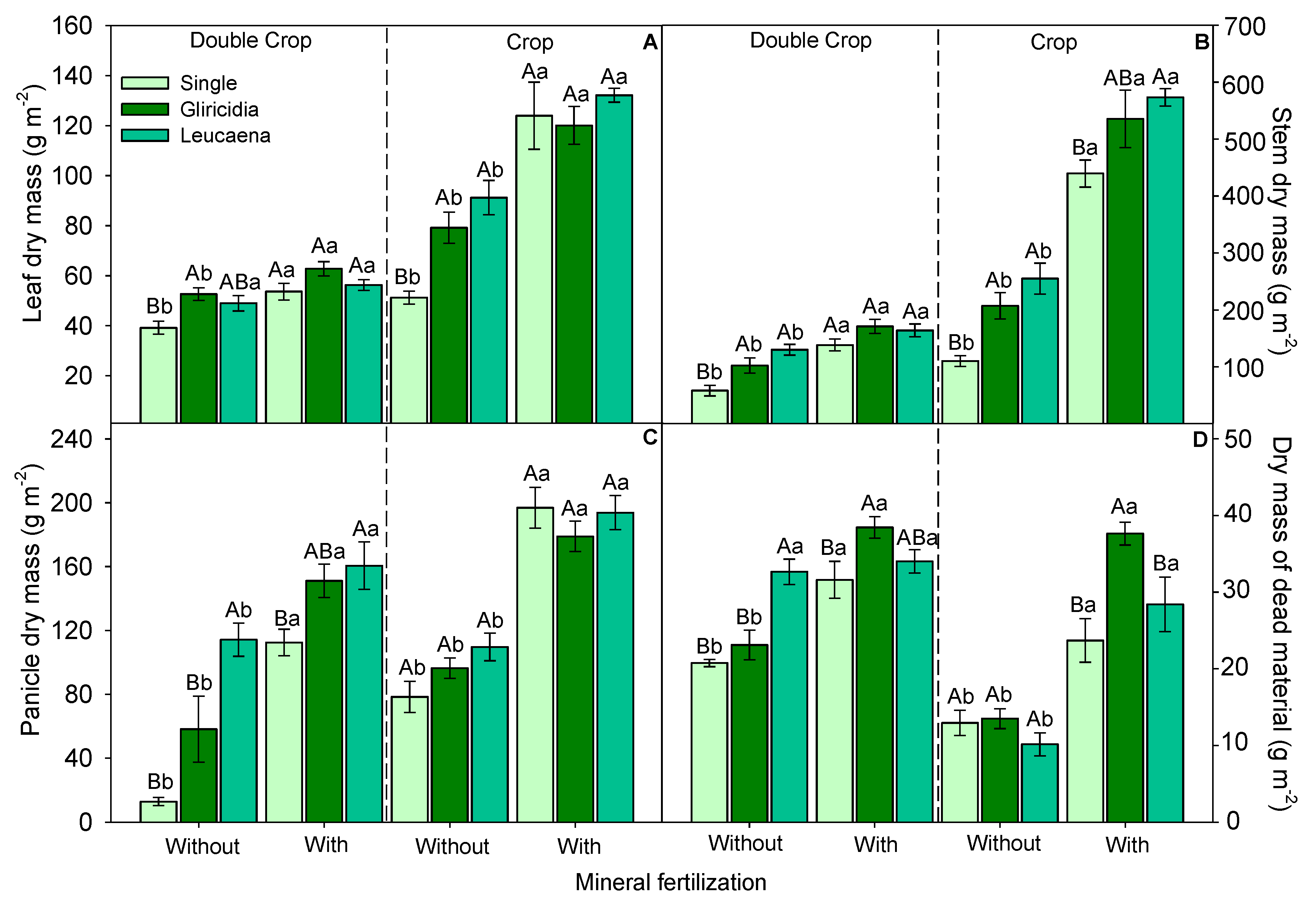
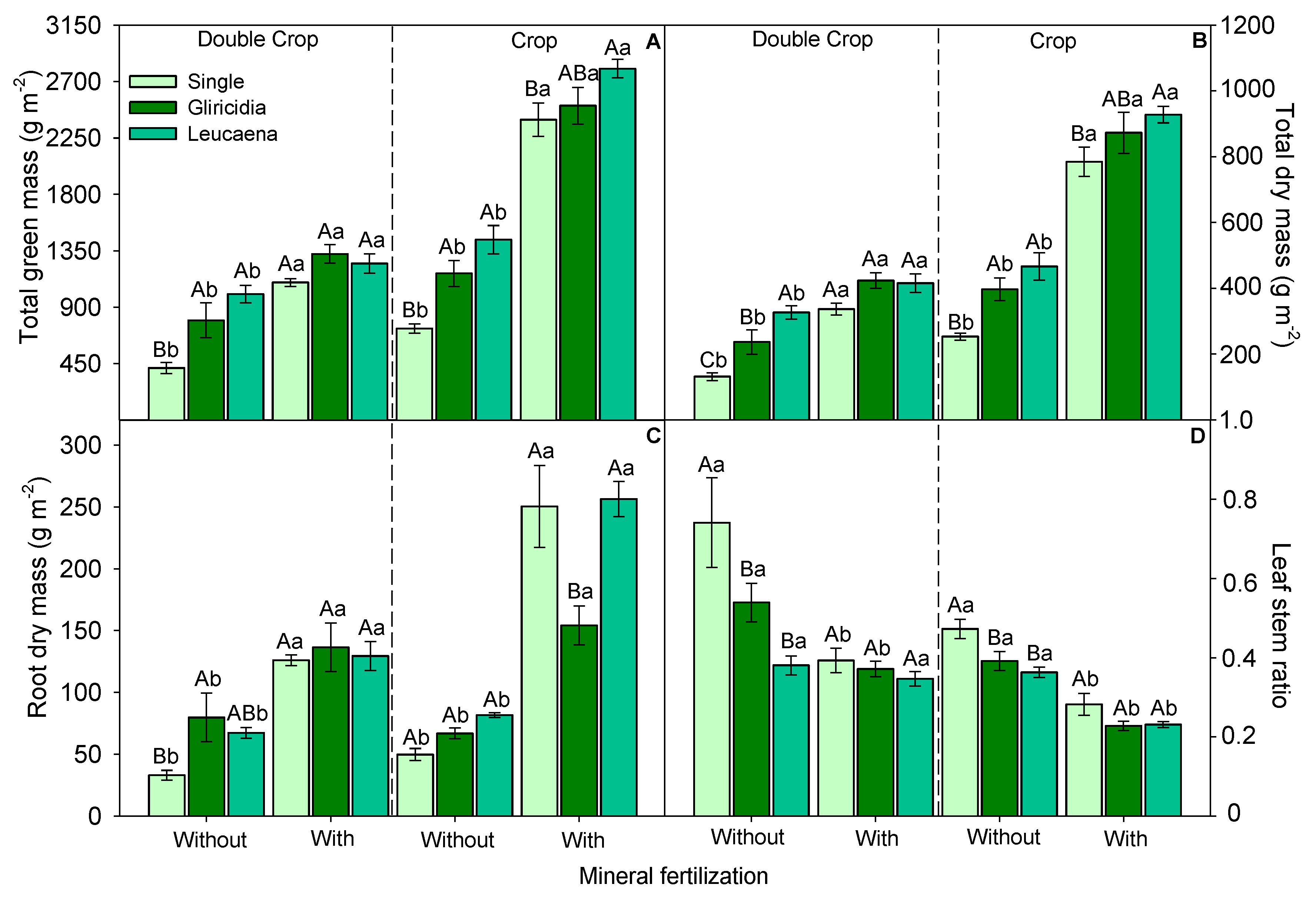
3.2. Morphological Components and Biomass Production of Forage Sorghum
3.3. Leaf Macronutrient Contents
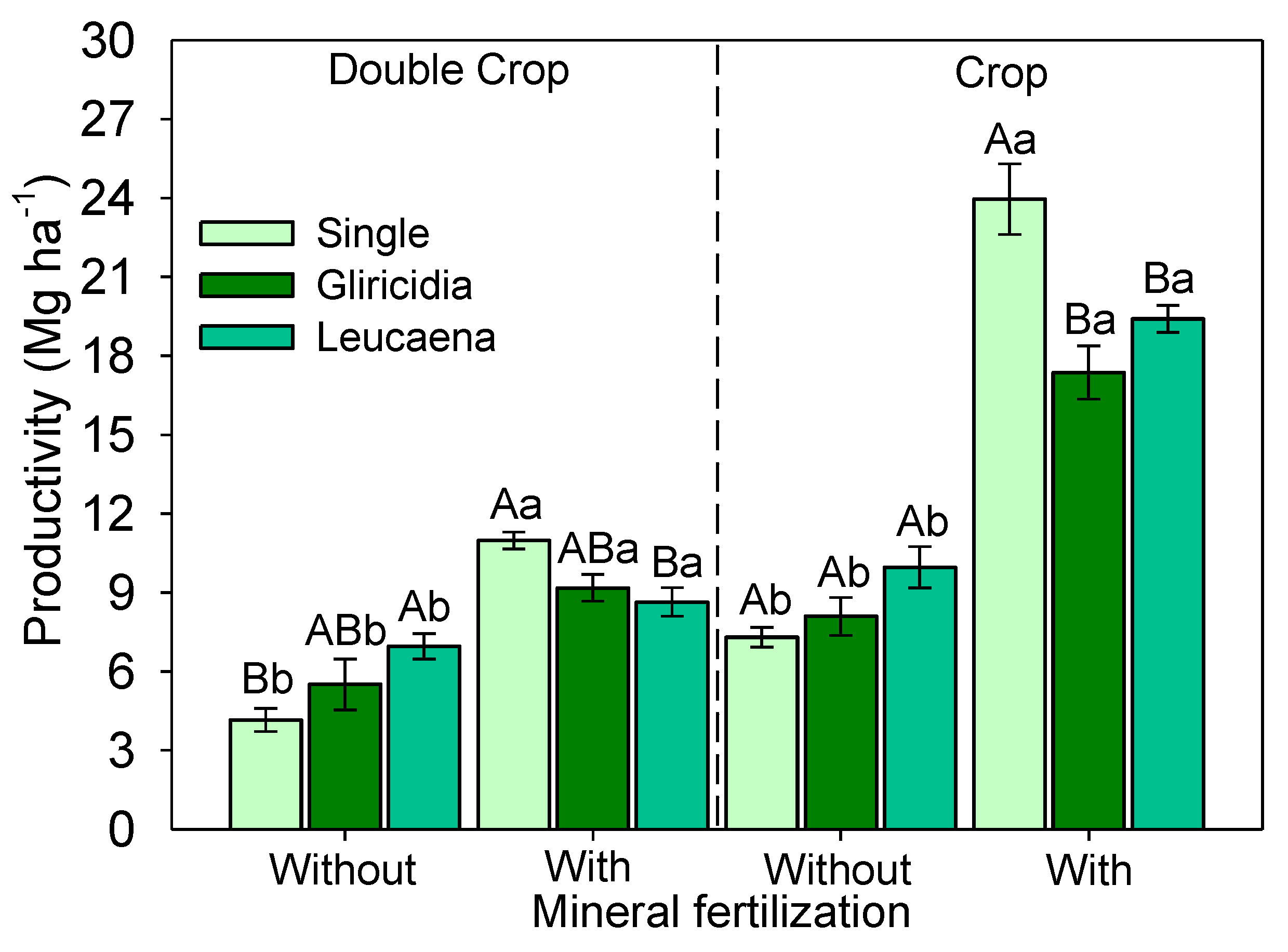
3.4. Sorghum Yield
4. Discussion
4.1. Morphological Components of Growth and Biomass Production of Forage Sorghum
4.2. Macronutrient Leaf Contents and Productivity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária). Sistema Brasileiro de Classificação de Solos, 3rd ed.; Centro Nacional de Pesquisa de Solos: Brasília, Brazil, 2013; p. 353. [Google Scholar]
- Ekepu, D.; Tirivanhu, P. Assessing socio-economic factors influencing adoption of legume-based multiple cropping systems among smallholder sorghum farmers in Soroti, Uganda. S. Afr. J. Agric. Ext. (SAJAE) 2016, 44, 195–215. [Google Scholar] [CrossRef]
- Omari, R.A.; Aung, H.P.; Hou, M.; Yokoyama, T.; Onwona-Agyeman, S.; Oikawa, Y.; Fujii, Y.; Bellingrath-Kimura, S.D. Influence of Different Plant Materials in Combination with Chicken Manure on Soil Carbon and Nitrogen Contents and Vegetable Yield. Pedosphere 2016, 26, 510–521. [Google Scholar] [CrossRef]
- Williams, D.M.; Blanco-Canqui, H.; Francis, C.A.; Galusha, T.D. Organic Farming and Soil Physical Properties: An Assessment after 40 Years. Agron. J. 2017, 109, 600. [Google Scholar] [CrossRef]
- Marin, A.M.P.; Menezes, R.S.C.; Salcedo, I.H. Produtividade de milho solteiro ou em aléias de gliricídia adubado com duas fontes orgânicas. Pesquisa Agropecuária Brasileira 2007, 42, 669–677. [Google Scholar] [CrossRef]
- Oliveira, F.R.A.D.; Souza, H.A.D.; Carvalho, M.A.R.D.; Costa, M.C.G. Green fertilization with residues of leguminous trees for cultivating maize in degraded soil. Rev. Caatinga 2018, 31, 798–807. [Google Scholar] [CrossRef]
- De Moura, E.G.; Albuquerque, J.M.; Aguiar, A.D.C.F. Growth and productivity of corn as affected by mulching and tillage in Alley cropping systems. Sci. Agricola 2008, 65, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.C.R.; Freitas, A.D.S.D.; Menezes, R.S.C.; Sampaio, E.V.D.S.B. Nitrogen symbiotically fixed by cowpea and gliricidia in traditional and agroforestry systems under semiarid conditions. Pesquisa Agropecuária Brasileira 2015, 50, 178–184. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, V.R.; E Silva, P.S.L.; Pontes, F.S.T.; De Paiva, H.N.; Antonio, R.P. Growth Of Arboreal Leguminous Plants And Maize Yield In Agroforestry Systems. Revista Árvore 2016, 40, 679–688. [Google Scholar] [CrossRef]
- Loss, A.; Pereira, M.G.; Ferreira, E.P.; Dos Santos, L.L.; Beutler, S.J.; Júnior, A.S.D.L.F. Frações oxidáveis do carbono orgânico em argissolo vermelho-amarelo sob sistema de aleias. Revista Brasileira de Ciência do Solo 2009, 33, 867–874. [Google Scholar] [CrossRef]
- Aguiar, A.D.C.F.; Amorim, A.P.; Coelho, K.P.; De Moura, E.G. Environmental and agricultural benefits of a management system designed for sandy loam soils of the humid tropics. Revista Brasileira de Ciência do Solo 2009, 33, 1473–1480. [Google Scholar] [CrossRef] [Green Version]
- Beedy, T.; Snapp, S.; Akinnifesi, F.; Sileshi, G. Impact of Gliricidia sepium intercropping on soil organic matter fractions in a maize-based cropping system. Agric. Ecosyst. Environ. 2010, 138, 139–146. [Google Scholar] [CrossRef]
- Primo, D.C.; Menezes, R.S.C.; Sampaio, E.V.D.S.B.; Garrido, M.D.S.; Junior, J.C.B.D.; Souza, C.S. Recovery of N applied as 15N-manure or 15N-gliricidia biomass by maize, cotton and cowpea. Nutr. Cycl. Agroecosyst. 2014, 100, 205–214. [Google Scholar] [CrossRef]
- Primo, D.; Menezes, R.; Filho, R.; Dutra, E.; Silva, E.; Alves, R.; Sampaio, E.; Antonino, A.; Lucena, E. Characteristics Physico-Chemical and Carbon Balance in Fluvic Entisol after Six Years Fertilization with Manure and Gliricidia. J. Exp. Agric. Int. 2018, 23, 1–11. [Google Scholar] [CrossRef]
- Moura-Silva, A.G.; Aguiar, A.D.C.F.; Jorge, N.; Agostini-Costa, T.D.S.; Moura, E.G. Food quantity and quality of cassava affected by leguminous residues and inorganic nitrogen application in a soil of low natural fertility of the humid tropics. Bragantia 2017, 76, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Mundus, S.; Menezes, R.S.C.; Neergaard, A.; Garrido, M.S. Maize growth and soil nitrogen availability after fertilization with cattle manure and/or gliricidia in semi-arid NE Brazil. Nutr. Cycl. Agroecosyst. 2008, 82, 61–73. [Google Scholar] [CrossRef]
- Calvo, C.L.; Foloni, J.S.S.; Brancalião, S.R. Produtividade de fitomassa e relação C/N de monocultivos e consórcios de guandu-anão, milheto e sorgo em três épocas de corte. Bragantia 2010, 69, 77–86. [Google Scholar] [CrossRef]
- De Oliveira, L.B.; Pires, A.J.V.; Viana, A.E.S.; Matsumoto, S.N.; De Carvalho, G.G.P.; Ribeiro, L.S.O. Produtividade, composição química e características agronômicas de diferentes forrageiras. Revista Brasileira de Zootecnia 2010, 39, 2604–2610. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.D.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Sousa, D.M.G.; Lobato, E. Cerrado: Correção do Solo E Adubação, 2nd ed.; Embrapa Cerrados: Brasília, Brazil, 2004; p. 416. [Google Scholar]
- Boaretto, A.E.; van Raij, B.; da Silva, F.C.; Chitolina, J.C.; Tedesco, M.J.; do Carmo, C.A.F. Amostragem acondicionamento e preparo de amostras de planta para análise química. In Manual de Análises Químicas de Solos, planta e fertilizantes; da Silva, F.C., Ed.; Embrapa informação tecnológica: Brasília, Brazil, 2009; pp. 61–85. [Google Scholar]
- Andrade Neto, R.C.; Miranda, N.O.; Duda, G.P.; Góes, G.B.; Lima, A.S. Crescimento e produtividade do sorgo forrageiro BR 601 sob adubação verde. Revista Brasileira de Engenharia Agrícola e Ambiental 2010, 14, 124–130. [Google Scholar] [CrossRef]
- Silva, A.R.D.; Collier, L.S.; Flores, R.A.; Santos, V.M.D.; Silva, L.L.D.; Oliveira, V.A.; Barbosa, J.M. Productive Yield of Cowpea and Maize in Single Crop and Mixtures in an Agroforestry System. Am. Eur. J. Agric. Environ. Sci. 2015, 15, 85–92. [Google Scholar]
- Silva, P.S.L.; Oliveira, V.R.D.; Silva, P.I.B.; Chicas, L.D.S.; Tomaz, F.L.D.S. Effects of ground cover from branches of arboreal species on weed growth and maize yield. Revista Ciência Agronômica 2015, 46, 809–817. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, W.P.; De Carvalho, G.J.; Neto, D.D.O.A.; Teixeira, L.G.V. Desempenho agronômico de plantas de cobertura usadas na proteção do solo no período de pousio. Pesquisa Agropecuária Brasileira 2013, 48, 157–166. [Google Scholar] [CrossRef]
- Heinrichs, R.; Vitti, G.C.; Moreira, A.; Fancelli, A.L. Produção e estado nutricional do milho em cultivo intercalar com adubos verdes. Revista Brasileira de Ciência do Solo 2002, 26, 225–230. [Google Scholar] [CrossRef]
- Fernandes, M.F.; Barreto, A.C. Cultivo de Gliricidia sepium e Leucaena leucocephala em alamedas visando a melhoria dos solos dos tabuleiros costeiros. Pesquisa Agropecuária Brasileira 2001, 36, 1287–1293. [Google Scholar]
- Martins, J.C.R.; Menezes, R.S.C.; Sampaio, E.V.S.B.; Dos Santos, A.F.; Nagai, M.A. Produtividade de biomassa em sistemas agroflorestais e tradicionais no Cariri Paraibano. Revista Brasileira de Engenharia Agrícola e Ambiental 2013, 17, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Gama, T.D.C.M.; Volpe, E.; Lempp, B.; Galdeia, E.C. Recuperação de pasto de capim-braquiária com correção e adubação de solo e estabelecimento de leguminosas. Revista Brasileira de Saúde e Produção Animal 2013, 14, 635–647. [Google Scholar] [CrossRef]
- Akinnifesi, F.K.; Makumba, W.; Sileshi, G.; Ajayi, O.C.; Mweta, D. Synergistic effect of inorganic N and P fertilizers and organic inputs from Gliricidia sepium on productivity of intercropped maize in Southern Malawi. Plant Soil 2007, 294, 203–217. [Google Scholar] [CrossRef]
- Chirwa, P.W.; Black, C.R.; Ong, C.K.; Maghembe, J. Nitrogen Dynamics in Cropping Systems in Southern Malawi Containing Gliricidia sepium, Pigeonpea and Maize. Agrofor. Syst. 2006, 67, 93–106. [Google Scholar] [CrossRef]
- De Paula, P.D.; Campello, E.F.C.; Guerra, J.G.M.; Santos, G.D.A.; De Resende, A.S. DECOMPOSIÇÃO DAS PODAS DAS LEGUMINOSAS ARBÓREAS Gliricidia sepium E Acacia angustissima EM UM SISTEMA AGROFLORESTAL. Ciência Florest. 2015, 25, 791–800. [Google Scholar] [CrossRef]
- Queiroz, L.R.; Coelho, F.C.; Barroso, D.G.; Galvão, J.C.C. Cultivo de milho consorciado com leguminosas arbustivas perenes no sistema de aléias com suprimento de fósforo. Rev. Ceres. 2008, 55, 409–415. [Google Scholar]
- Hauggaard-Nielsen, H.; Jensen, E.S. Facilitative Root Interactions in Intercrops. Plant Soil 2005, 274, 237–250. [Google Scholar] [CrossRef]
- Makumba, W.; Janssen, B.; Oenema, O.; Akinnifesi, F.; Mweta, D.; Kwesiga, F. The long-term effects of a gliricidia–maize intercropping system in Southern Malawi, on gliricidia and maize yields, and soil properties. Agric. Ecosyst. Environ. 2006, 116, 85–92. [Google Scholar] [CrossRef]
- Primo, D.C.; Menezes, R.S.C.; Oliveira, F.F.D.; Dubeux Júnior, J.C.B.; Sampaio, E.V.S.B. Timing and placement of cattle manure and/or gliricidia affects cotton and sunflower nutrient accumulation and biomass productivity. Anais da Academia Brasileira de Ciências 2018, 90, 415–424. [Google Scholar] [CrossRef] [PubMed]
- De Moura, E.G.; Serpa, S.S.; Dos Santos, J.G.D.; Sobrinho, J.R.S.C.; Aguiar, A.D.C.F. Nutrient use efficiency in alley cropping systems in the Amazonian periphery. Plant Soil 2010, 335, 363–371. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.; Murphy, A. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017; p. 888. [Google Scholar]
- Korwar, G.R. Fodder production potential of leucaena hedgerows on an Alfisol and a Vertisol in the semi-arid tropics. In Proceedings of the International Workshop on Nitrogen Fixing Trees for Fodder Production, Morillion, AR, USA, 20–25 March 1995; pp. 146–153. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | |
|---|---|
| pH (H2O) | 5.3 |
| Organic matter (g kg−1) | 6.0 |
| Available P (mg dm−3) | 7.48 |
| Available K (mg dm−3) | 8.0 |
| Ca2+ (cmolc dm−3) | 2.47 |
| Mg2+ (cmolc dm−3) | 1.19 |
| Al3+ (cmolc dm−3) | 0.04 |
| H+ + Al3+ (cmolc dm−3) | 1.78 |
| CEC (cmolc dm−3) | 5.46 |
| Sand (g kg−1) | 893.5 |
| Silt (g kg−1) | 6.5 |
| Clay (g kg−1) | 100.0 |
| Legume | N (g kg−1) | P (g kg−1) | K (g kg−1) | Ca (g kg−1) | Mg (g kg−1) | Dry Mass (Mg ha−1) |
|---|---|---|---|---|---|---|
| Gliricidia | 32.8 | 2.8 | 17.6 | 14.6 | 5.5 | 5.4 |
| Leucaena | 33.1 | 1.7 | 11.4 | 10.5 | 3.2 | 6.0 |
| Middle Square | Double Crop | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variation Source | PH | SD | PD | PL | LDM | SDM | PDM | DMDM | TGM | TDM | RDM |
| Cultivation systems (C) | 687 * | 0.03 * | 2.5 * | 34 * | 318 * | 6573 * | 14,027 * | 131 * | 389,177 * | 49,341 * | 2108 * |
| Mineral fertilization (M) | 4276 * | 0.24 * | 6.3 * | 89 * | 846* | 27,919 * | 47,521 * | 632 * | 1,763,023 * | 192,993 * | 37,439 * |
| (C × M) | 484 * | 0.008 * | 0.8 * | 12 * | 33 * | 1445 * | 2102 | 126 * | 124,341 * | 9758 * | 966 * |
| CV (%) | 8.5 | 6.7 | 11.8 | 6.9 | 12.3 | 19.7 | 27.8 | 10.6 | 17.4 | 17.7 | 29.0 |
| Crop | |||||||||||
| Cultivation systems (C) | 68.5 ns | 0.2 * | 0.7 * | 1.1 ns | 1443 * | 50,903 * | 658ns | 155 * | 777,522 * | 81,883 * | 8905 * |
| Mineral fertilization (M) | 16450 * | 1.3 * | 12.4 * | 150 * | 19,885 * | 793,331 * | 67,794 * | 2353 * | 1,590,2883 * | 1,797,184 * | 178,427 * |
| (C × M) | 55.1 ns | 0.01 * | 0.2 * | 1.2 ns | 841 * | 93 * | 1028 ns | 112 * | 82,518 * | 3378 * | 8815 * |
| CV (%) | 8.0 | 12.3 | 9.8 | 4.1 | 15.9 | 17.0 | 15.1 | 25.2 | 12.1 | 13.7 | 26.1 |
| Middle Square | Double Crop | ||||||
|---|---|---|---|---|---|---|---|
| Variation Source | Stem/Leaf Relation | N | P | K | Ca | Mg | Yield |
| Cultivation systems (C) | 0.1 * | 1.7 * | 0.01 ns | 70.3 * | 0.6 * | 0.5 * | 5259 * |
| Mineral fertilization (M) | 0.2 * | 0.004 ns | 0.007 ns | 37.8 * | 0.001 * | 0.3 * | 123,413 * |
| (C × M) | 0.06 * | 0.05 * | 0.001 ns | 52.6 | 0.08 * | 0.3 * | 167,987 * |
| CV (%) | 26.8 | 7.0 | 3.4 | 12.7 | 13.3 | 21.3 | 16.4 |
| Crop | |||||||
| Cultivation systems (C) | 0.01 * | 85.8 * | 0.7 * | 0.5 ns | 0.5 * | 0.09 * | 218,755 * |
| Mineral fertilization (M) | 0.19* | 0.07 ns | 0.02 ns | 120 * | 0.5 * | 0.5 * | 104,247 * |
| (C × M) | 0.002 * | 1.4 ns | 0.05 * | 2.8 ns | 0.1 * | 0.006 ns | 443,964 * |
| CV (%) | 14.3 | 10.8 | 14.3 | 9.2 | 13.8 | 15.0 | 12.2 |
| Cultivation System | Mineral Fertilization | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Absent | Present | Average | Absent | Present | Average | Absent | Present | Average | Absent | Present | Average | Absent | Present | Average | |||
| N (g kg−1) | P (g kg−1) | K (g kg−1) | Ca (g kg−1) | Mg (g kg−1) | |||||||||||||
| Double crop | |||||||||||||||||
| Single | 7.2 Aa | 7.3 Aa | - | 1.6 Aa | 1.6 Aa | 1.67A | 10.8 Cb | 17.9 Aa | - | 1.46 Aa | 1.26 Ab | - | 1.2 Aa | 0.6 Ab | - | ||
| Gliricidia | 6.8 Aa | 6.7 ABa | - | 1.5 Aa | 1.6 Aa | 1.59B | 16.2 Ba | 18.0 Aa | - | 0.95 Ba | 1.1 ABa | - | 0.7 Ba | 0.6 Aa | - | ||
| Leucaena | 6.5 Aa | 6.3 Ba | - | 1.5 Aa | 1.6 Aa | 1.61B | 20.7 Aa | 18.6 Aa | - | 0.81 Ba | 0.9 Ba | - | 0.4 Ca | 0.5 Aa | - | ||
| Average | - | - | - | 1.6 a | 1.6 a | - | - | - | - | - | - | - | - | - | - | ||
| Crop | |||||||||||||||||
| Single | 20.9 Ba | 20.0 Ba | 20.5 B | 2.1 Aa | 1.8 Ba | - | 18.3 Ab | 21.1 Aa | 19.7 A | 1.4 Aa | 1.1 Ab | - | 0.9 Aa | 0.6 Ab | 0.78 A | ||
| Gliricidia | 23.8 ABa | 24.1 ABa | 23.9 A | 2.0 Aa | 2.0 Ba | - | 16.8 Ab | 21.7 Aa | 19.2 A | 1.1 Ba | 0.7 Bb | - | 0.7 Ba | 0.4 Bb | 0.5 B | ||
| Leucaena | 26.1 Aa | 26.5 Aa | 26.3 A | 2.4 Aa | 2.49 Aa | - | 17.3 Ab | 21.6 Aa | 19.4 A | 0.8 Ca | 0.7 Ba | - | 0.8 ABa | 0.5 ABb | 0.7 AB | ||
| Average | 23.6 a | 23.5 a | - | - | - | - | 17.4 b | 21.4 a | - | - | - | - | 0.8 a | 0.5 b | - | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Costa Leite, R.; Donizetti dos Santos, J.G.; da Costa Leite, R.; Fernandes Sousa, L.; de Sousa Soares, G.O.; Fernandes Rodrigues, L.; Santana da Silva Carneiro, J.; Clementino dos Santos, A. Leguminous Alley Cropping Improves the Production, Nutrition, and Yield of Forage Sorghum. Agronomy 2019, 9, 636. https://doi.org/10.3390/agronomy9100636
da Costa Leite R, Donizetti dos Santos JG, da Costa Leite R, Fernandes Sousa L, de Sousa Soares GO, Fernandes Rodrigues L, Santana da Silva Carneiro J, Clementino dos Santos A. Leguminous Alley Cropping Improves the Production, Nutrition, and Yield of Forage Sorghum. Agronomy. 2019; 9(10):636. https://doi.org/10.3390/agronomy9100636
Chicago/Turabian Styleda Costa Leite, Robson, José Geraldo Donizetti dos Santos, Rubson da Costa Leite, Luciano Fernandes Sousa, Guilherme Octávio de Sousa Soares, Luan Fernandes Rodrigues, Jefferson Santana da Silva Carneiro, and Antonio Clementino dos Santos. 2019. "Leguminous Alley Cropping Improves the Production, Nutrition, and Yield of Forage Sorghum" Agronomy 9, no. 10: 636. https://doi.org/10.3390/agronomy9100636
APA Styleda Costa Leite, R., Donizetti dos Santos, J. G., da Costa Leite, R., Fernandes Sousa, L., de Sousa Soares, G. O., Fernandes Rodrigues, L., Santana da Silva Carneiro, J., & Clementino dos Santos, A. (2019). Leguminous Alley Cropping Improves the Production, Nutrition, and Yield of Forage Sorghum. Agronomy, 9(10), 636. https://doi.org/10.3390/agronomy9100636